
Do Indigenous Street Trees Promote More Biodiversity than Alien Ones? Evidence Using Mistletoes and Birds in South Africa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Methods
2.3. Data Analyses
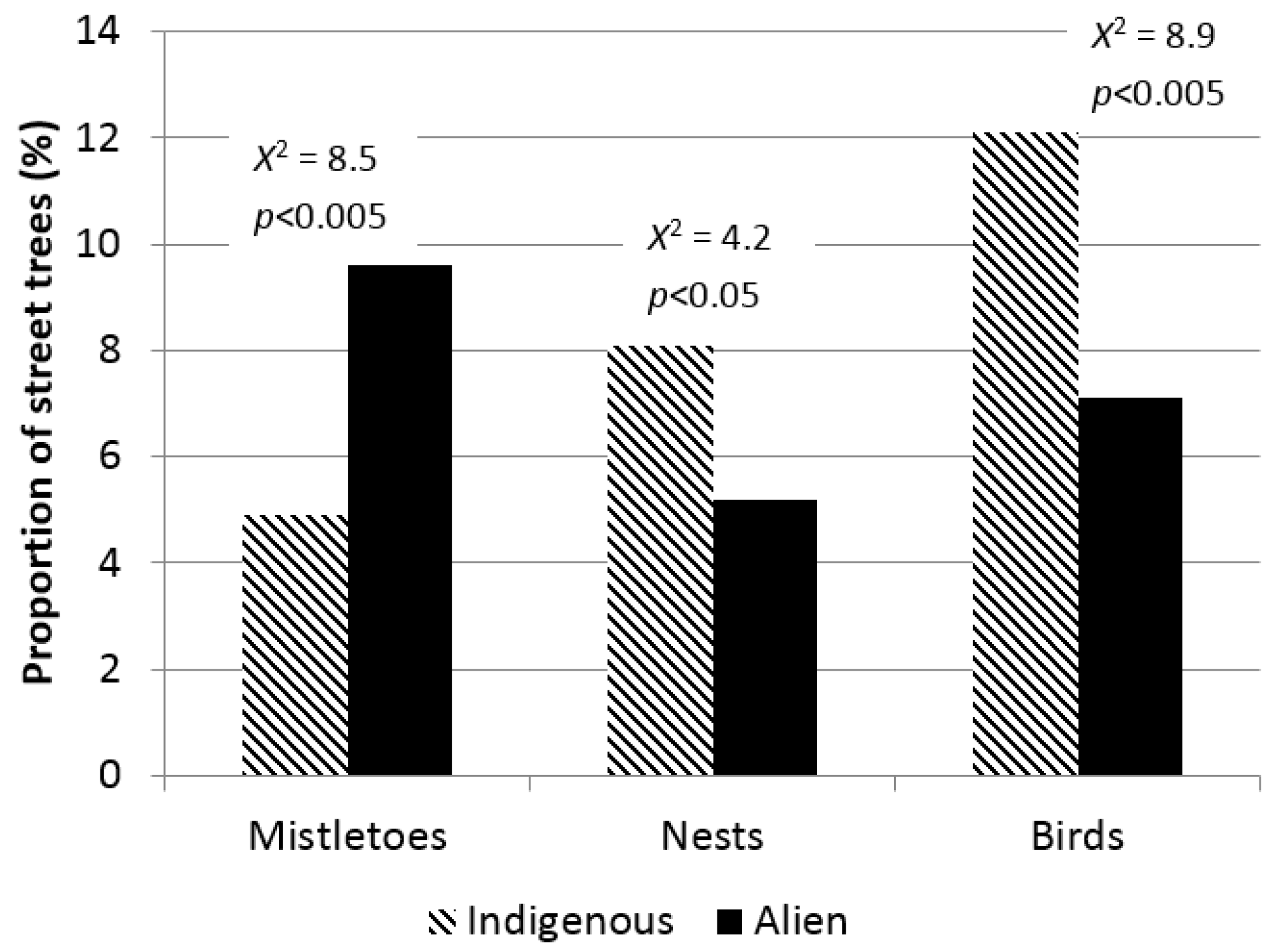
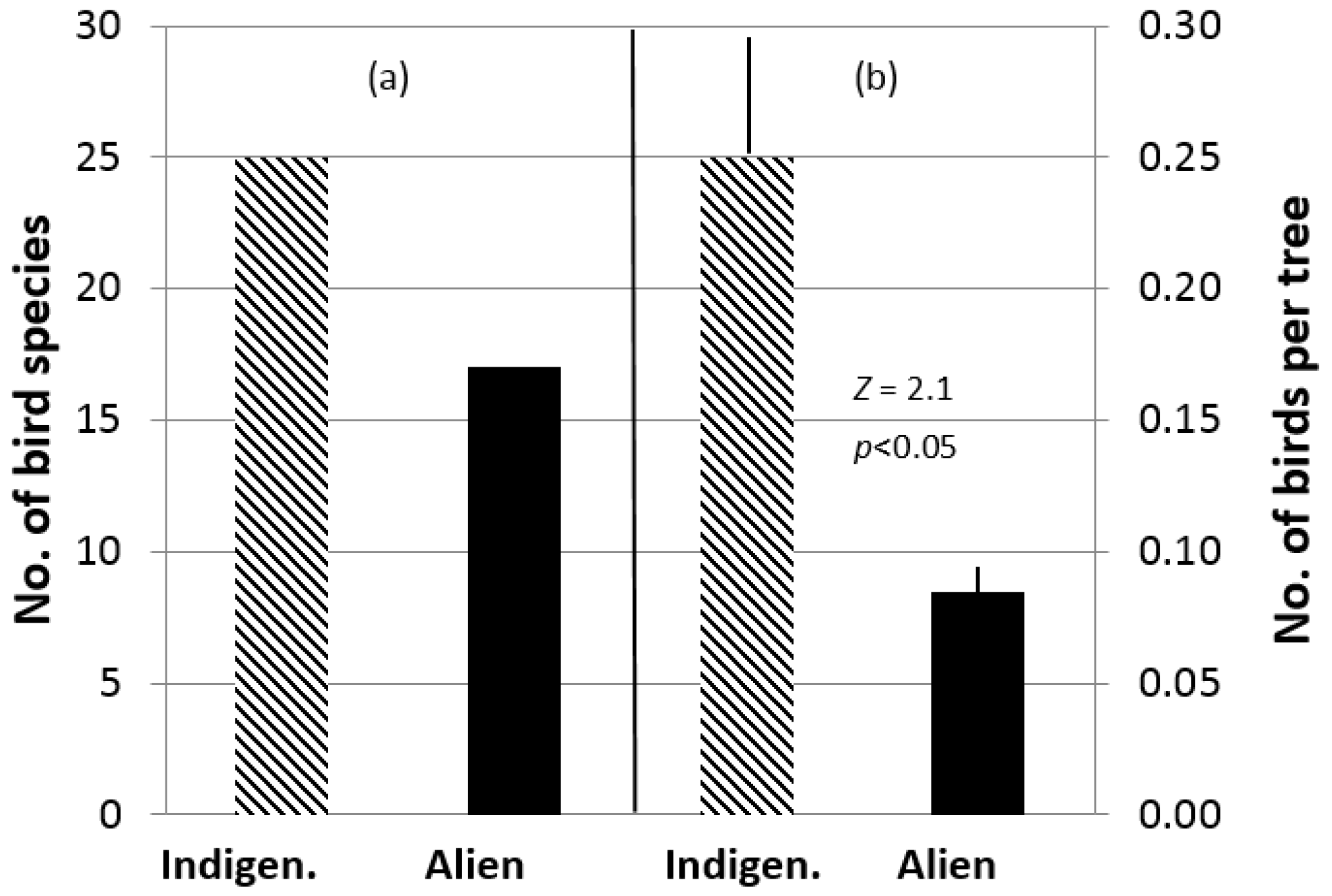
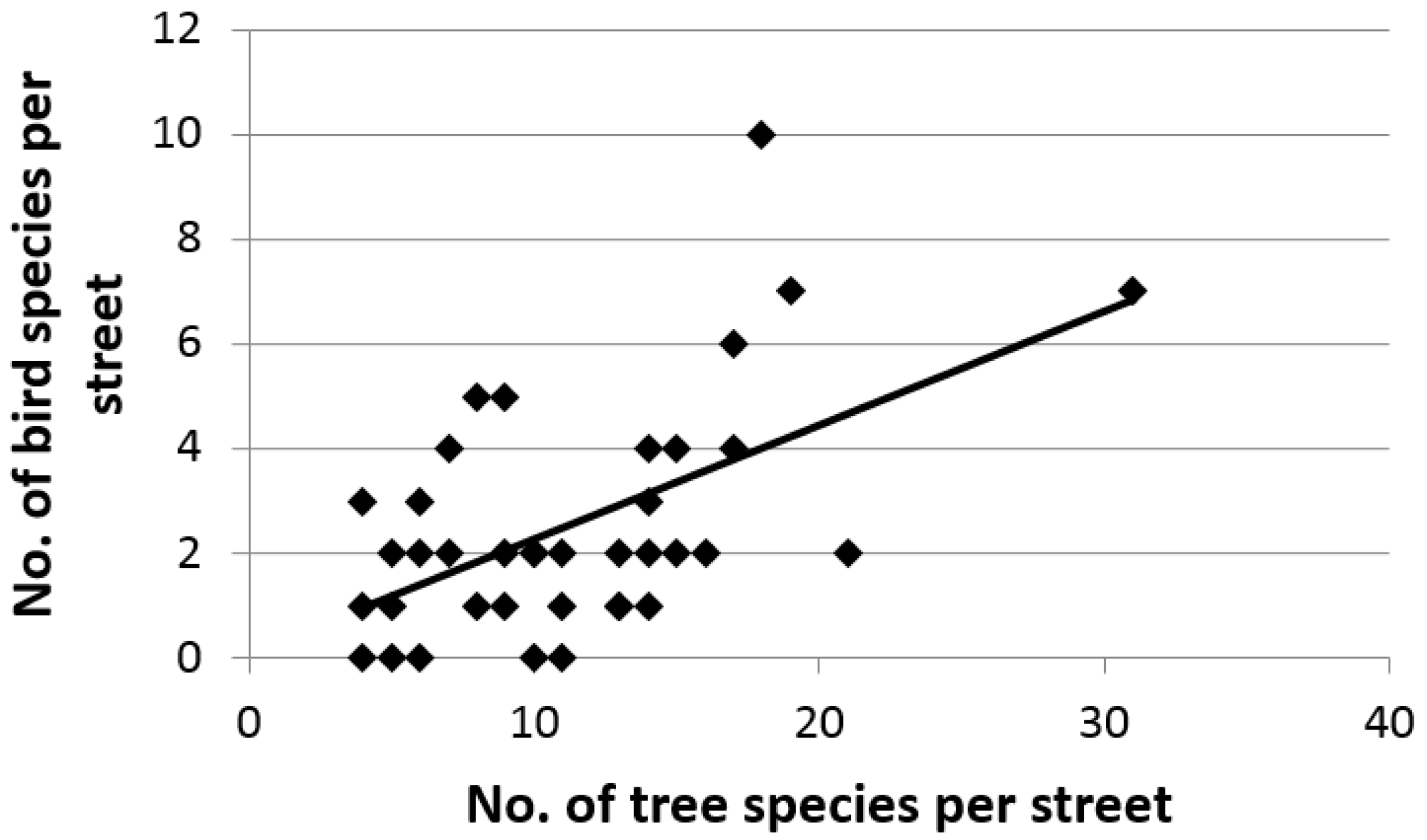
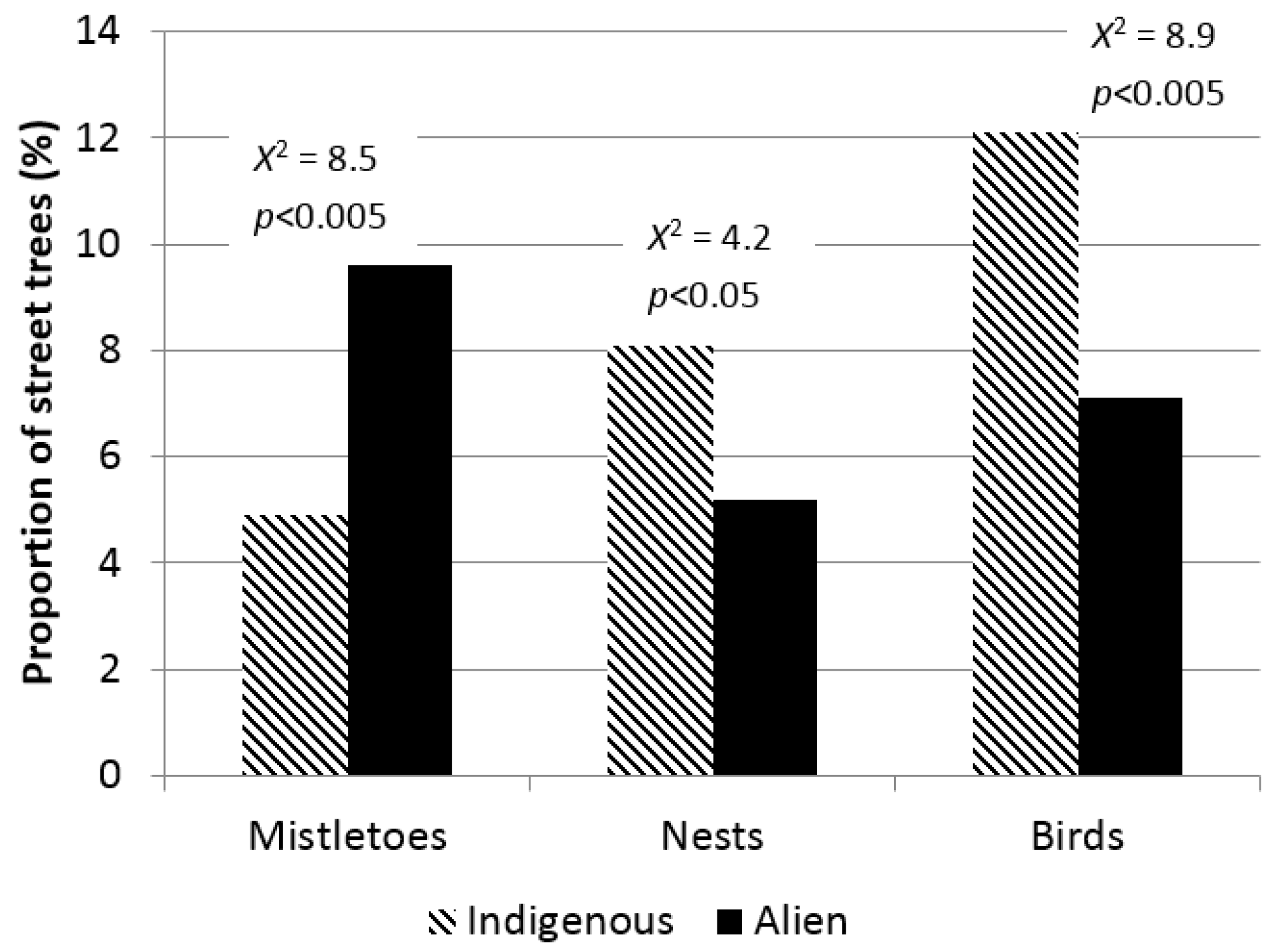
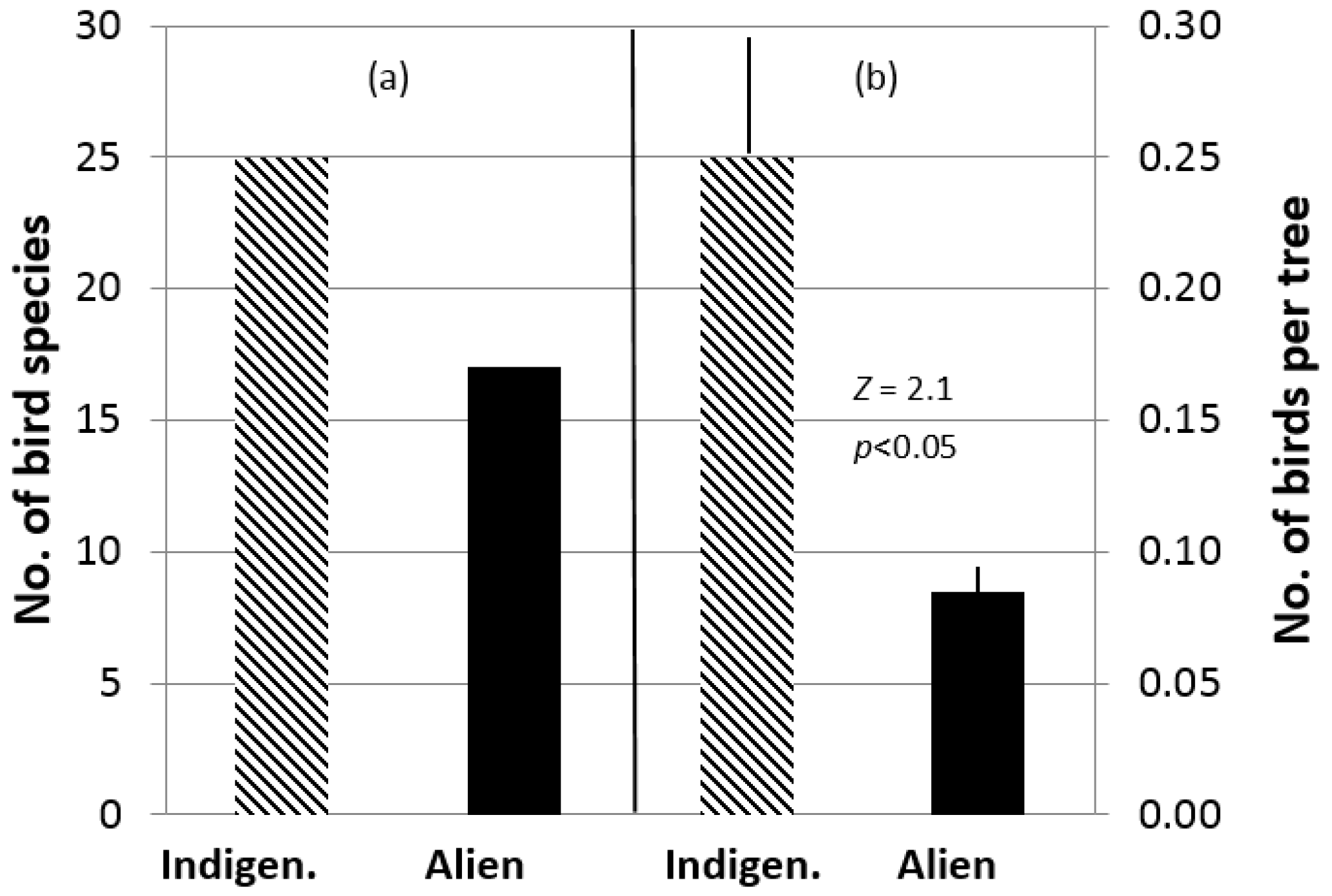
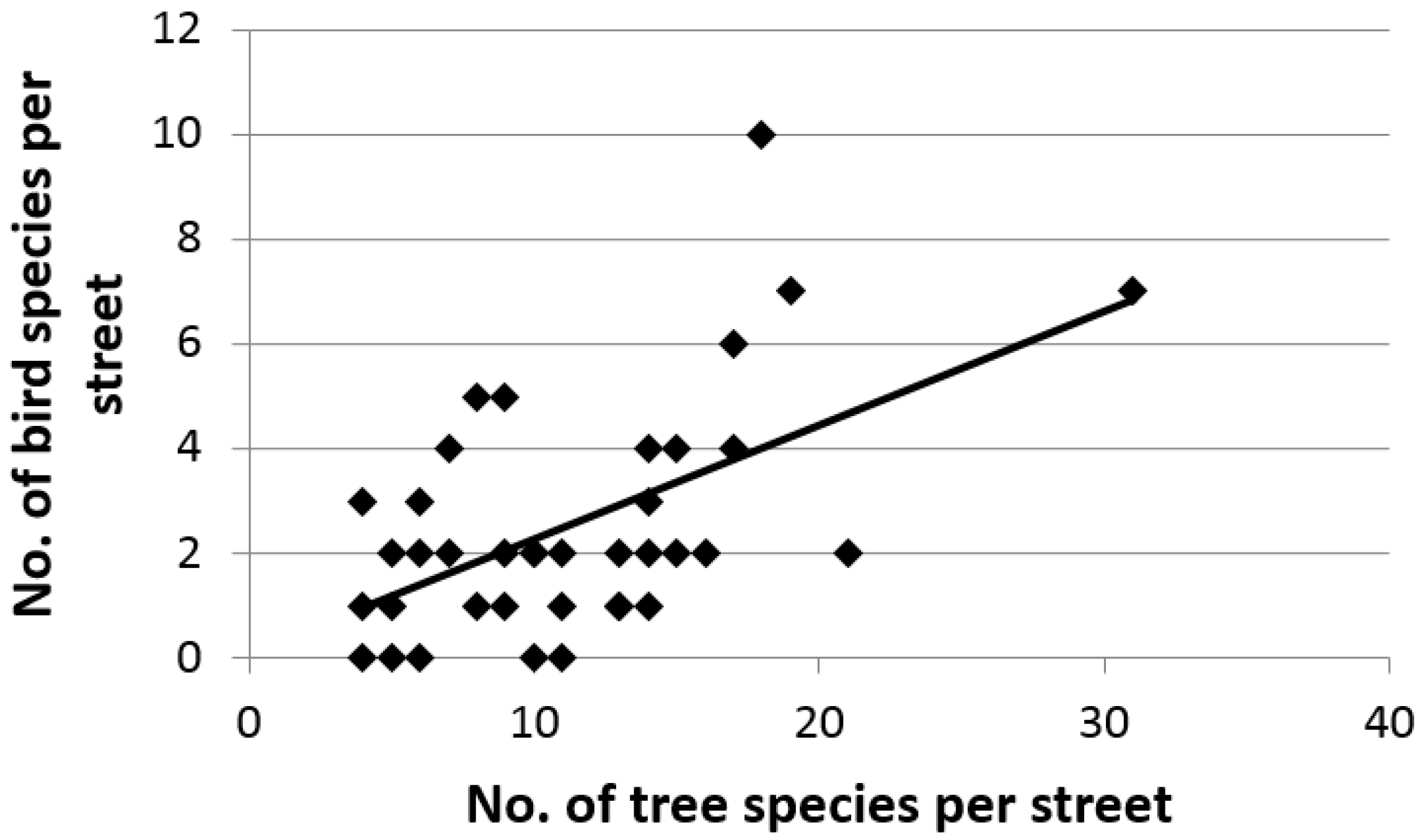
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Niemelä, J.; Saarela, S.-R.; Söderman, T.; Kopperoinen, L.; Yli-Pelkonen, V.; Väre, S.; Kotze, D.J. Using the ecosystem services approach for better planning and conservation of urban green spaces: A Finland case study. Biodiv. Conserv. 2010, 19, 3225–3243. [Google Scholar] [CrossRef]
- Roy, S.; Byrne, J.; Pickering, C. A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different climatic zones. Urban For. Urban Green. 2012, 11, 351–363. [Google Scholar] [CrossRef]
- Livesley, S.J.; McPherson, G.M.; Calfapietra, C. The urban forest and ecosystem services: Impacts on urban water, heat and pollution cycles at the tree, street and city scale. J. Environ. Qual. 2016, 45, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Todorova, A.; Asakawa, S.; Aikoh, T. Preferences for and attitudes towards street flowers and trees in Sapporo, Japan. Landsc. Urban Plan. 2004, 69, 403–416. [Google Scholar] [CrossRef]
- Antupit, S.; Gray, B.; Woods, S. Steps ahead: Making streets that work in Seattle, Washington. Landsc. Urban Plan. 1996, 35, 107–122. [Google Scholar] [CrossRef]
- Tzilkowski, W.M.; Wakeley, J.S.; Morris, L.J. Relative use of municipal street trees by birds during summer in state college, Pennsylvania. Urban Ecol. 1986, 9, 387–398. [Google Scholar] [CrossRef]
- Kubista, C.E.; Bruckner, A. Importance of urban trees and building as daytime roost for bats. Biologia 2015, 70, 1545–1552. [Google Scholar] [CrossRef]
- Kendle, A.D.; Rose, J.E. The aliens have landed! What are the justifications for ‘native only’ policies in landscape plantings? Landsc. Urban Plan. 2000, 47, 19–31. [Google Scholar] [CrossRef]
- Smith, R.M.; Thompson, K.; Hodgson, J.G.; Warren, P.H.; Gaston, K.J. Urban domestic gardens (IX): Composition and richness of the vascular plant flora, and implications for native biodiversity. Biol. Conserv. 2006, 129, 312–322. [Google Scholar] [CrossRef]
- Johnston, M.; Nail, S.; James, S. ‘Natives versus aliens’: The relevance of the debate to urban forest management in Britain. In Trees, People and the Built Environment, Proceedings of the Urban Trees Research Conference, Birmingham, UK, 13–14 April 2012; Johnston, M., Percival, G., Eds.; pp. 181–191.
- Chalker-Scott, L. Nonnative, noninvasive woody species can enhance landscape biodiversity. Arboric. Urban For. 2015, 41, 173–186. [Google Scholar]
- Carthew, S.M.; Yáñez, B-M.; Ruykys, L. Straddling the divide: Den use by brushtail possums (Trichosurus vulpecula) in urban parklands. Urban Ecosyst. 2015, 18, 525–538. [Google Scholar] [CrossRef]
- Blanchon, D.; Pusateri, J.; Galbraith, M.; Thorpe, S. Sampling indigenous ground-living beetles in a stand of non-native tree privet (Ligustrum lucidum) in New Zealand raises new management questions. Ecol. Manag. Restor. 2011, 12, 234–236. [Google Scholar] [CrossRef]
- Chong, K.Y.; Teo, S.; Kurukulasuriya, B.; Chung, Y.F.; Rajathurai, S.; Tan, H.T.W. Not all green is as good: Different effects of the natural and cultivated components of urban vegetation on bird and butterfly diversity. Biol. Conserv. 2014, 171, 299–309. [Google Scholar] [CrossRef]
- Gairola, S.; Bhatt, A.; Govender, Y.; Baijnath, H.; Porcheş, S.; Ramdhani, S. Incidence and intensity of tree infestation by the mistletoe Erianthemum dregei (Eckl. & Zeyh.) V. Tieghem in Durban, South Africa. Urban For. Urban Green. 2013, 12, 315–322. [Google Scholar]
- French, K.; Major, R.; Hely, K. Use of native and exotic garden plants by suburban nectarivorous birds. Biol. Conserv. 2005, 121, 545–559. [Google Scholar] [CrossRef]
- Gray, E.R.; van Heezik, Y. Exotic trees can sustain native birds in urban woodlands. Urban Ecosyst. 2015. [Google Scholar] [CrossRef]
- Dickie, I.A.; Bennett, B.M.; Burrows, L.E.; Nunez, M.A.; Peltzer, D.A.; Porté, A.; Richardson, D.M.; Rejmánek, M.; Rundel, P.W.; van Wilgen, B.W. Conflicting values: Ecosystem services and invasive tree management. Biol. Invasions 2014, 16, 705–719. [Google Scholar] [CrossRef]
- Climatedata.eu. Available online: http://www.climatedata.eu/climate.php?loc=sfzz0020&lang=en (accessed on 13 March 2013).
- State of the Environment South Africa, 2007. Available online: http://soer.deat.gov.za/332.htm (accessed on 15 March 2015).
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19; South African National Biodiversity Institute (SANBI): Pretoria, South Africa, 2006; p. 816. [Google Scholar]
- McConnachie, M.M.; Shackleton, C.M. Public green space inequality in small towns in South Africa. Habit. Int. 2010, 34, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Kuruneri-Chitepo, C.; Shackleton, C.M. The distribution, abundance and composition of street trees in selected towns of the Eastern Cape, South Africa. Urban For. Urban Green. 2011, 10, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Gwedla, N.; Shackleton, C.M. The development visions and attitudes towards urban forestry of officials responsible for greening in South African towns. Land Use Policy 2015, 42, 17–26. [Google Scholar] [CrossRef]
- Richardson, E.; Shackleton, C.M. The extent, causes and local perceptions of street tree damage in small towns in the Eastern Cape, South Africa. Urban For. Urban Green. 2014, 13, 425–432. [Google Scholar] [CrossRef]
- Ikin, K.; Knight, E.; Lindenmayer, D.B.; Fischer, J.; Manning, A.D. The influence of native versus exotic streetscape vegetation on the spatial distribution of birds in suburbs and reserves. Divers. Distrib. 2013, 19, 294–306. [Google Scholar] [CrossRef]
- Bhullar, S.; Majer, J. Arthropods on street trees: A food resource for wildlife. Pac. Conserv. Biol. 2000, 6, 171–173. [Google Scholar] [CrossRef]
- Watson, D.M. The relative contribution of specialists and generalists to mistletoe dispersal: Insights from a neotropical rain forest. Biotropica 2013, 45, 195–202. [Google Scholar] [CrossRef]
- Izuddin, M.; Webb, E.L. The influence of tree architecture, forest remnants, and dispersal syndrome on roadside epiphyte diversity in a highly urbanized tropical environment. Biodivers. Conserv. 2015, 24, 2063–2077. [Google Scholar] [CrossRef]
- Palomino, D.; Carrascal, L.M. Birds on novel island environments: A case study with the urban avifauna of Tenerife (Canary Islands). Ecol. Res. 2005, 20, 611–617. [Google Scholar] [CrossRef]
- MacGregor-Fors, I. Relation between habitat attributes and bird species richness in western Mexico suburbs. Landsc. Urban Plan. 2008, 84, 92–08. [Google Scholar] [CrossRef]
- Stagoll, K.; Lindenmayer, D.B.; Knight, E.; Fischer, J.; Manning, A.D. Large trees are keystone structures in urban parks. Conserv. Lett. 2012, 5, 115–122. [Google Scholar] [CrossRef]
- Nagendra, H.; Gopal, D. Tree diversity, distribution, history and change in urban parks: Studies in Bangalore, India. Urban Ecosyst. 2011, 14, 211–223. [Google Scholar] [CrossRef]
- Heilmann-Clausen, J.; Christensen, M. Does size matter? On the importance of various dead wood fractions for fungal diversity in Danish beech forests. For. Ecol. Manag. 2004, 201, 105–117. [Google Scholar] [CrossRef]
- Johansson, P.; Erlén, J. Influence of habitat quantity, quality and isolation on the distribution and abundance of two epiphytic lichens. J. Ecol. 2003, 91, 213–221. [Google Scholar]
- Vergnes, A.; Le Viol, I.; Clergeau, P. Green corridors in urban landscapes affect the arthropod communities of domestic gardens. Biol. Conserv. 2012, 145, 171–178. [Google Scholar] [CrossRef]
- Braaker, S.; Ghazoul, J.; Obrist, M.K.; Moretti, M. Habitat connectivity shapes urban arthropod communities: The key role of green roofs. Ecology 2014, 95, 1010–1021. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Alien | % of All Trees | Indigenous | % of All Trees |
|---|---|---|---|
| Jacaranda mimosifolia | 8.7 | Erythrina caffra | 6.0 |
| Schinus terebinthifolius | 8.6 | Celtis africana | 5.6 |
| Fraxinus spp. | 7.3 | Acacia karroo | 5.0 |
| Brachychiton acerfolium | 6.4 | Ekebergia capensis | 3.4 |
| Grevillea robusta | 5.9 | Harpephyllum caffrum | 3.2 |
| Total number of tree species | 61 | Total number of tree species | 40 |
| Total proportion (%) of all street trees | 64.6 | Total proportion (%) of all street trees | 35.4 |
| Species | Origin | No. Sampled | Mistletoes | Nests | Birds |
|---|---|---|---|---|---|
| Acacia karroo | Indig | 63 | 0.8 | 3.2 | 2.1 |
| Celtis africana | Indig | 70 | 0.2 | 1.9 | 1.0 |
| Ekebergia capensis | Indig | 43 | 0 | 0.4 | 2.1 |
| Erythrina caffra | Indig | 76 | 0 | 0.4 | 1.9 |
| Harpephyllum caffrum | Indig | 40 | 0 | 0 | 1.1 |
| Podocarpus falcatus | Indig | 34 | 0 | 0.5 | 1.3 |
| Brachychiton acerfolium | Alien | 81 | 0 | 0.2 | 0.4 |
| Brachychiton populneum | Alien | 24 | 0.5 | 0.7 | 0.5 |
| Eucalyptus ficifolia | Alien | 40 | 0 | 0 | 0.3 |
| Fraxinus sp | Alien | 92 | 6.4 | 1.9 | 0.4 |
| Grevillea robusta | Alien | 74 | 0 | 0.9 | 1.2 |
| Jacaranda mimosifolia | Alien | 110 | 0.1 | 0.3 | 0.7 |
| Quercus robur | Alien | 91 | 0.3 | 1.8 | 1.5 |
| Schinus terebenthifolius | Alien | 109 | 0 | 0.2 | 0.8 |
| Tipua tipu | Alien | 17 | 0 | 0 | 0.7 |
| Attribute | Mistletoes | Nests | Birds |
|---|---|---|---|
| Street | n.s | n.s | n.s |
| Distance to nearest green area | 0.00002 | 0.0023 | 0.0116 |
| Aspect | 0.0257 | n.s | n.s |
| Tree species | 0.0048 | n.s | n.s |
| Origin | 0.0003 | 0.0361 | 0.0016 |
| Tree circumference | n.s | n.s | 0.00001 |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shackleton, C. Do Indigenous Street Trees Promote More Biodiversity than Alien Ones? Evidence Using Mistletoes and Birds in South Africa. Forests 2016, 7, 134. https://doi.org/10.3390/f7070134
Shackleton C. Do Indigenous Street Trees Promote More Biodiversity than Alien Ones? Evidence Using Mistletoes and Birds in South Africa. Forests. 2016; 7(7):134. https://doi.org/10.3390/f7070134
Chicago/Turabian StyleShackleton, Charlie. 2016. "Do Indigenous Street Trees Promote More Biodiversity than Alien Ones? Evidence Using Mistletoes and Birds in South Africa" Forests 7, no. 7: 134. https://doi.org/10.3390/f7070134
APA StyleShackleton, C. (2016). Do Indigenous Street Trees Promote More Biodiversity than Alien Ones? Evidence Using Mistletoes and Birds in South Africa. Forests, 7(7), 134. https://doi.org/10.3390/f7070134