1. Introduction
Biodiversity conservation and enhancement are key considerations in urban forest research and policy, given rapid rates of urbanization worldwide and growing concerns about biodiversity declines [
1]. Many studies have explored the range of benefits and services that diverse urban forests provide, including greater ecosystem productivity, higher resilience to environmental perturbations, and increased wildlife habitat [
2,
3,
4]. Consequently, diversity targets have been incorporated into the core objectives of urban forest management plans [
5,
6].
Despite the recognition that diversity is crucial to urban forest health and city sustainability, urban foresters and scholars continue to disagree on the importance of native tree species contributions to overall forest diversity; some argue that native tree species promote the establishment of other native organisms and increase the ecological integrity of urban ecosystems [
1,
3,
7], while others question the prioritization of nativeness over non-native, non-invasive species that are more resilient to urban stresses and are vital for urban biodiversity enhancement [
8,
9]. Furthermore, biotic homogenization has been identified as a pressing challenge to biodiversity enhancement in cities, as widespread exotic species can outcompete and replace native ones in anthropogenic environments, potentially contributing to species extirpations and threatening local, unique ecosystems [
2,
10,
11]. These arguments and concerns illustrate the importance of elucidating the factors shaping urban forest diversity as well as determining how diversity benefits can be strategically and effectively maximized.
Suburban development is of particular importance to urban biodiversity research, as studies have shown that both native- and non-native tree-species richness can peak in neighbourhoods developed at the periphery of cities, located between the countryside and the urban core [
1,
12]. This trend has been attributed to the high level of landscape and habitat heterogeneity (e.g., private gardens, streetscapes, public green spaces including parks) that can occur in suburban and peri-urban areas [
13,
14,
15]. Research has revealed that the type of land cover (e.g., fields, grassland, forests, roads, ruderal habitat) can dictate suburban plant diversity, as habitat edges often promote greater species richness as well as the establishment of rare species [
16,
17]. Furthermore, relatively undisturbed forest patches have been shown to be more suitable for protecting native plant and tree species compared to areas that have undergone significant anthropogenic change [
18]. Diversity on residential properties is also influenced by home owner attitudes towards species selection, which often focus on aesthetics and promote exotic and ornamental species plantings [
2,
4]. Patterns in landscape structure and species composition emphasize the need to consider the planning and design of subdivisions during and after development, especially if urban forest diversity is to be considered a priority.
The heterogeneity of suburban landscapes, including the multiple land types on which trees can grow, necessitates the involvement of various urban forest professionals and practitioners in urban forest management. Trees found in parks and along streets generally fall under the jurisdiction of the municipality, and are subject to policies and targets determined by urban foresters and planners pertaining to tree planting guidelines, species selection, and diversity goals [
5,
6]. The land types found in suburban areas represent differences in tree establishment and management, reinforcing the importance of considering other factors such as policy and administration along with biophysical characteristics and home owner preferences when exploring how urban forest diversity is shaped.
Approximately two out of three Canadians live in suburban areas, which are currently experiencing much higher population growth rates compared to inner-city neighbourhoods located closer to city centres [
19]. The extensive suburbanization occurring across the country highlights the need to consider how suburban development influences urban forest diversity and potentially provides opportunities for biodiversity enhancement in urban areas. Although the effects of urban development on urban forest biodiversity have been examined in a single Canadian city [
1,
4], researchers have yet to compare forest diversity losses and/or gains associated with development and previous land use in different cities. The goal of this study was thus to explore whether contrasting development patterns in two Canadian cities also determine differences in suburban tree diversity.
2. Materials and Methods
2.1. Study Sites
The two cities chosen for this study are Halifax, Nova Scotia (44.6478°N, 63.5714°W), and London, Ontario (42.9837°N, 81.2497°W). Although both cities are found in the eastern region of the country, they are situated approximately 1500 km apart (
Appendix Figure A1). Halifax is located in the Acadian forest region, a mixed broadleaf and temperate forest comprised of both broadleaf and coniferous species including red spruce (
Picea rubens), yellow birch (
Betula alleghaniensis), sugar maple (
Acer saccharum), and balsam fir (
Abies balsamea) [
20,
21]. Some relatively undisturbed hinterland forest stands are found in suburban and peri-urban neighbourhoods in Halifax [
6]. London is located at the northern range of the Carolinian forest region, characterized by a wide variety of broadleaf tree species like hackberry (
Celtis occidentalis), black walnut (
Juglans nigra), hickory (
Carya spp.), and tulip tree (
Liriodendron tulipfera). London is surrounded primarily by agricultural fields and some scattered remnant woodland corridors [
22].
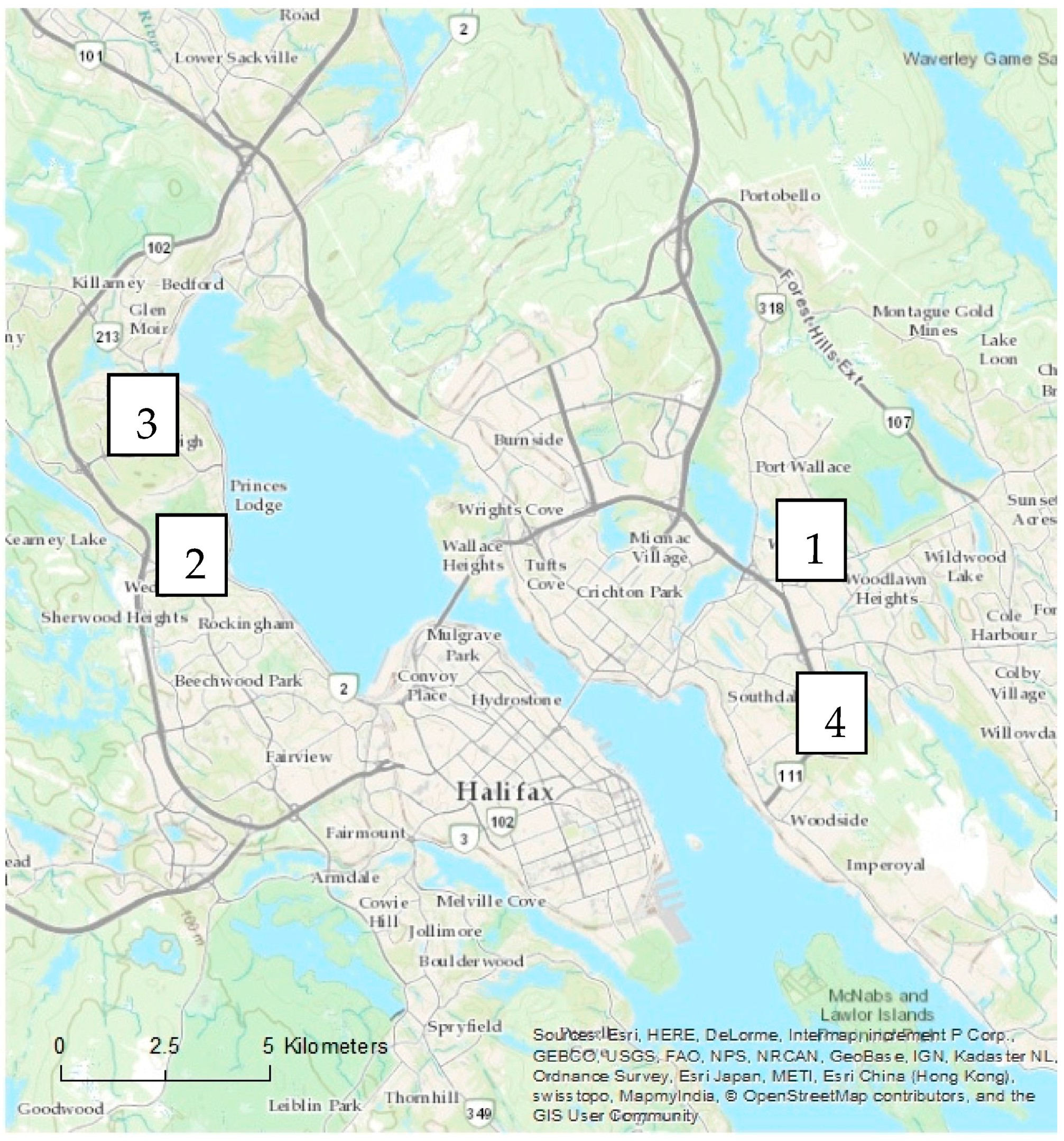
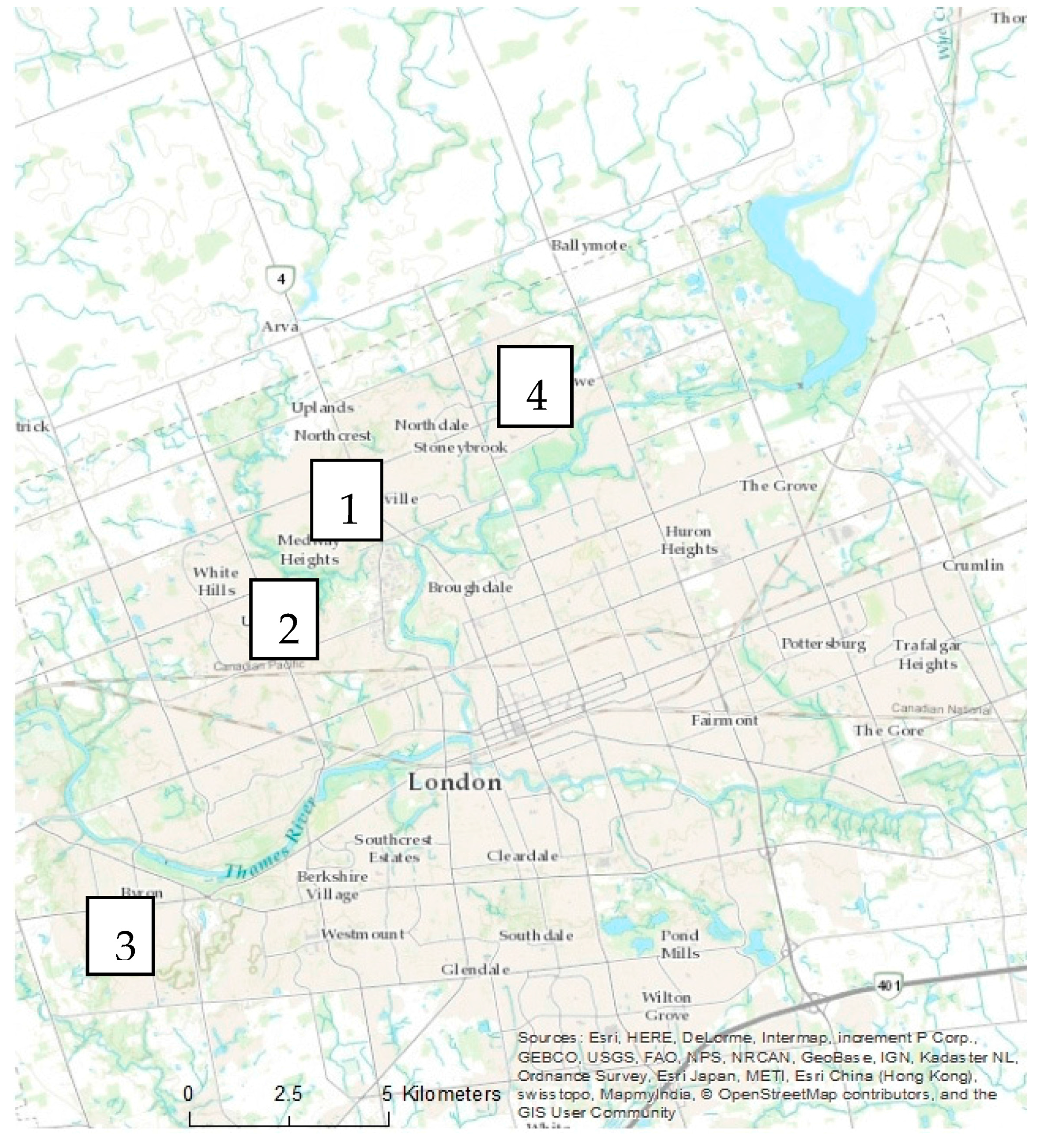
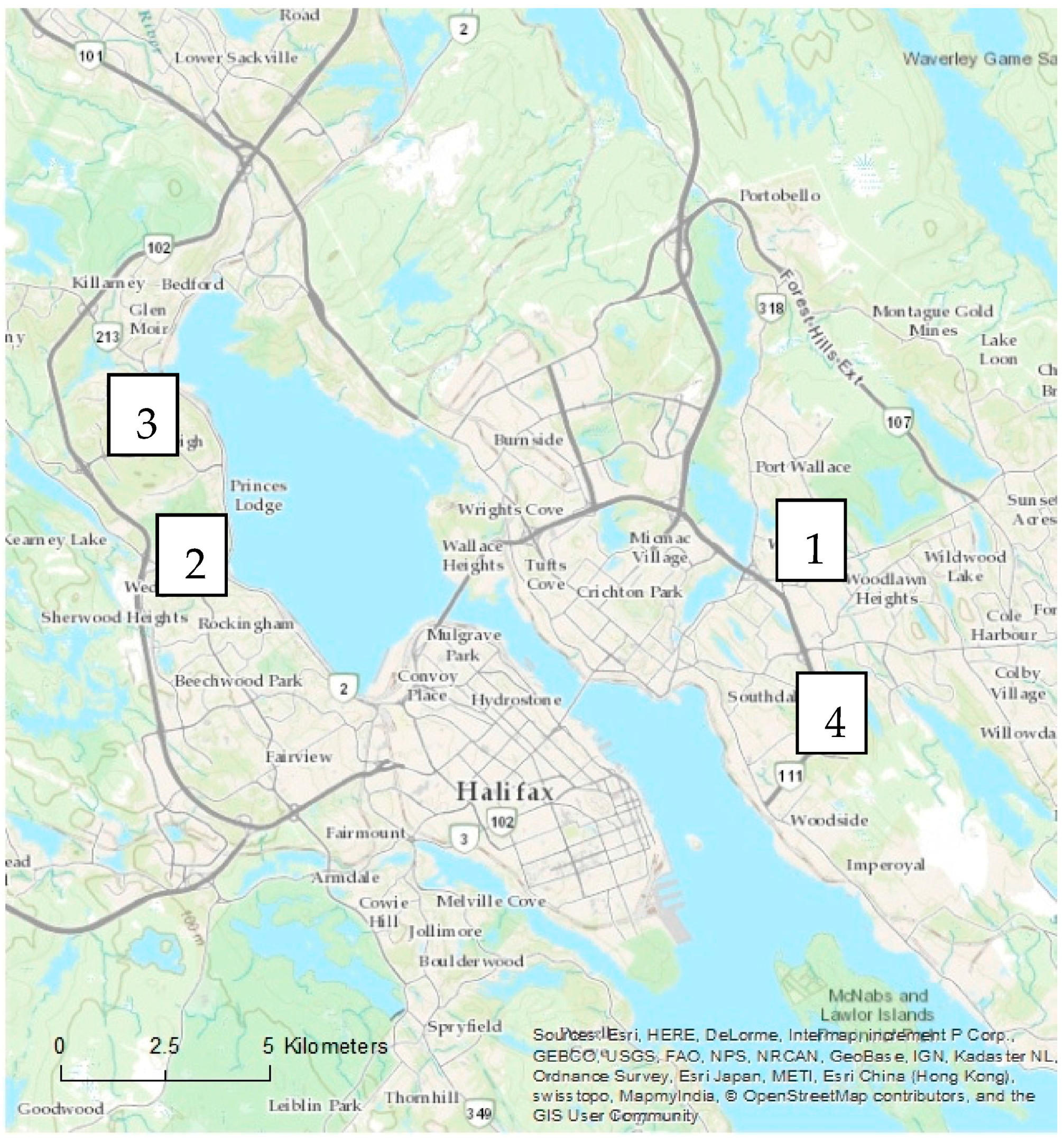
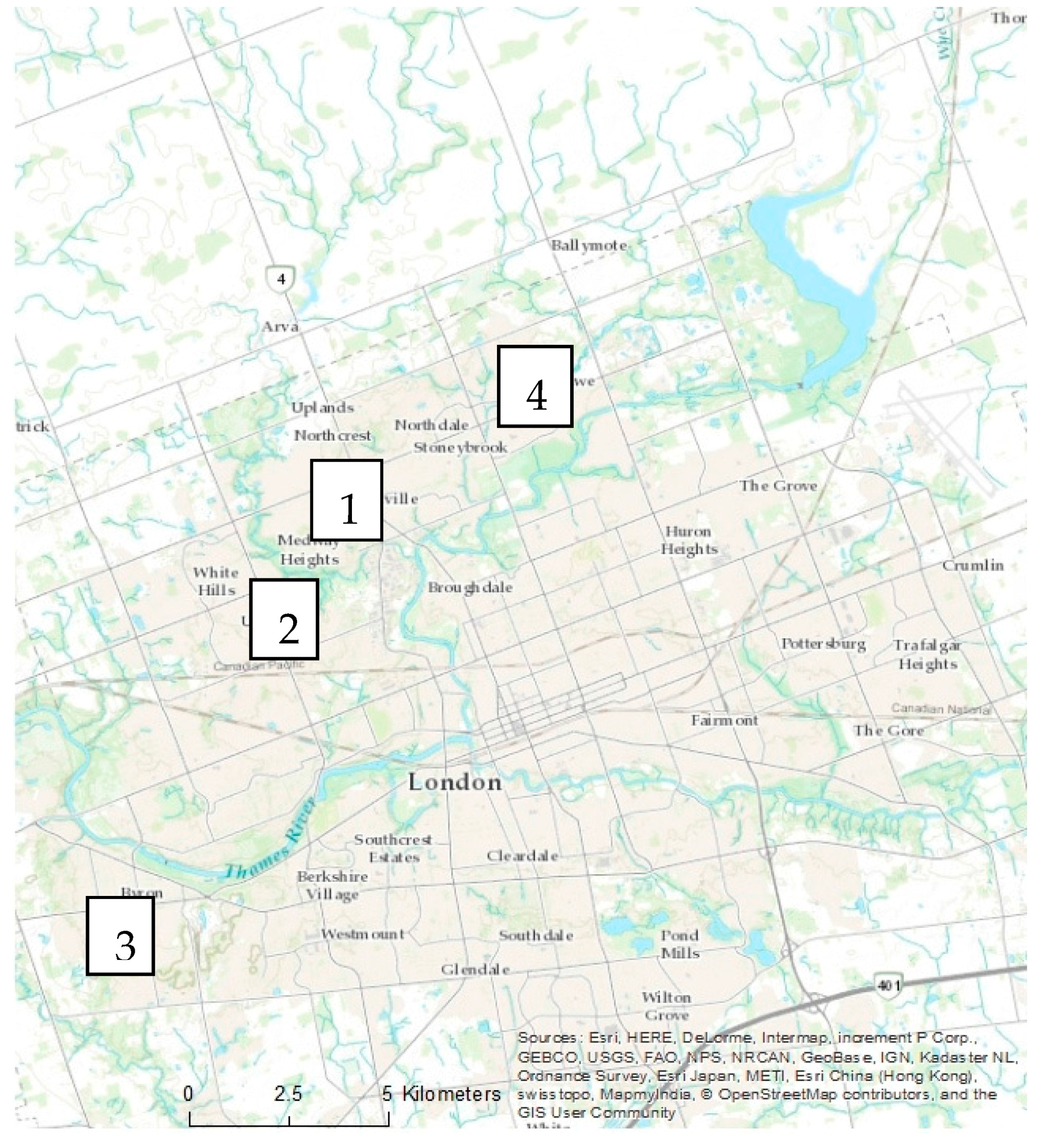
In each city, four residential neighbourhoods representing two age categories were examined (
Appendix Figure A2 and
Figure A3,
Table 1 and
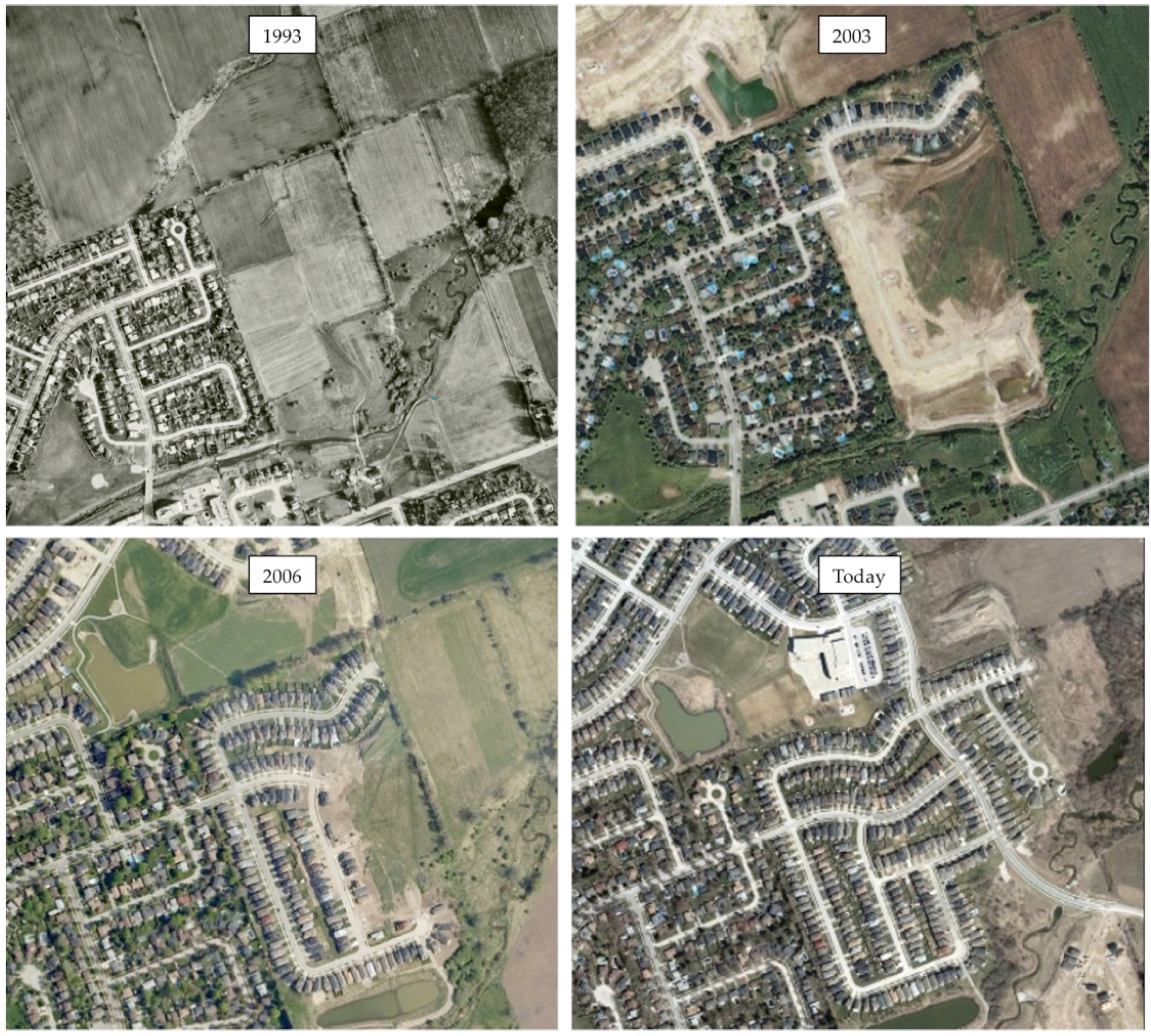
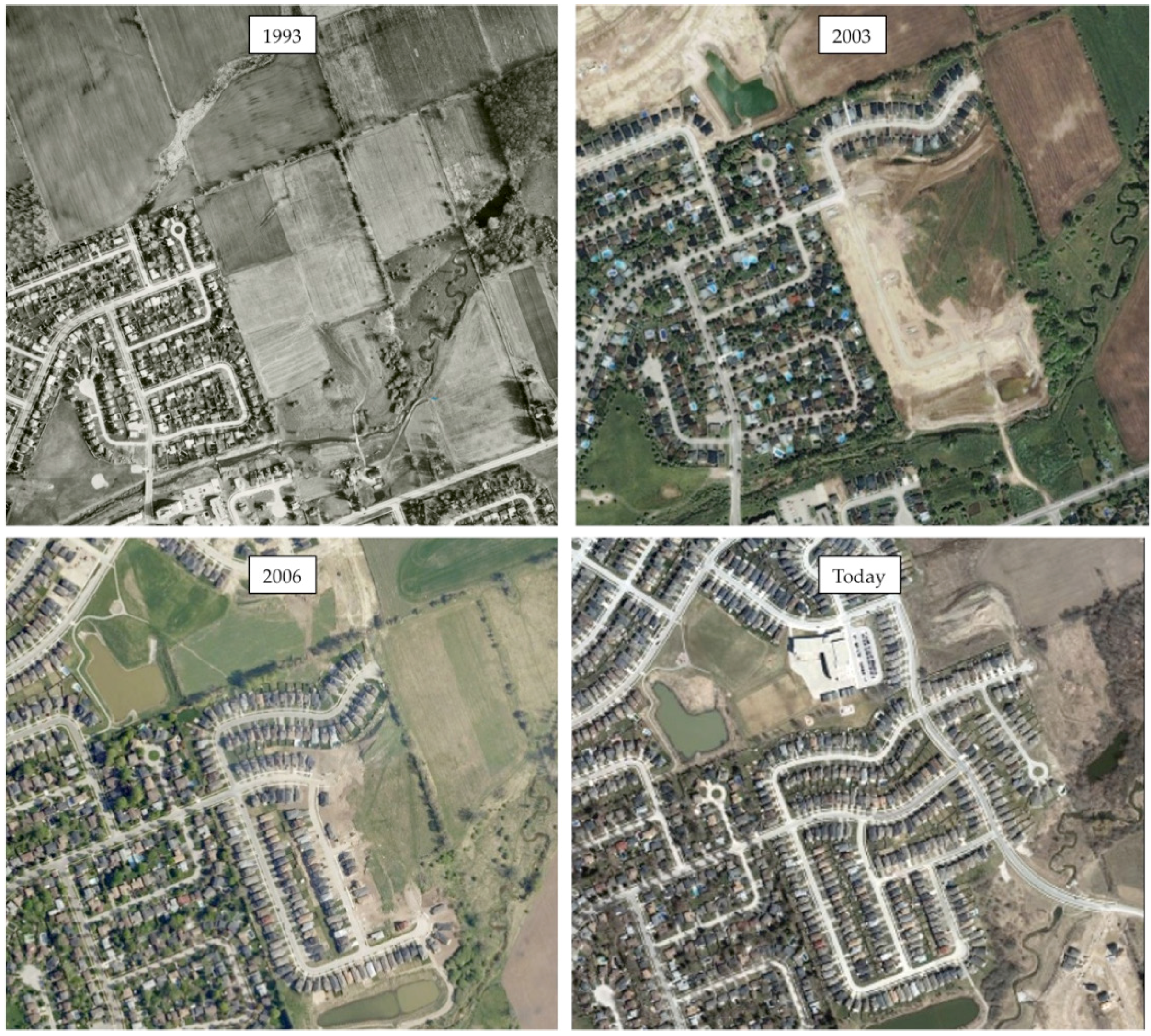
Table 2). Neighbourhood selection was based on the pre-urbanized landscape, which was determined by consulting with urban foresters and planners as well as historical aerial imagery (
Figure 1 and
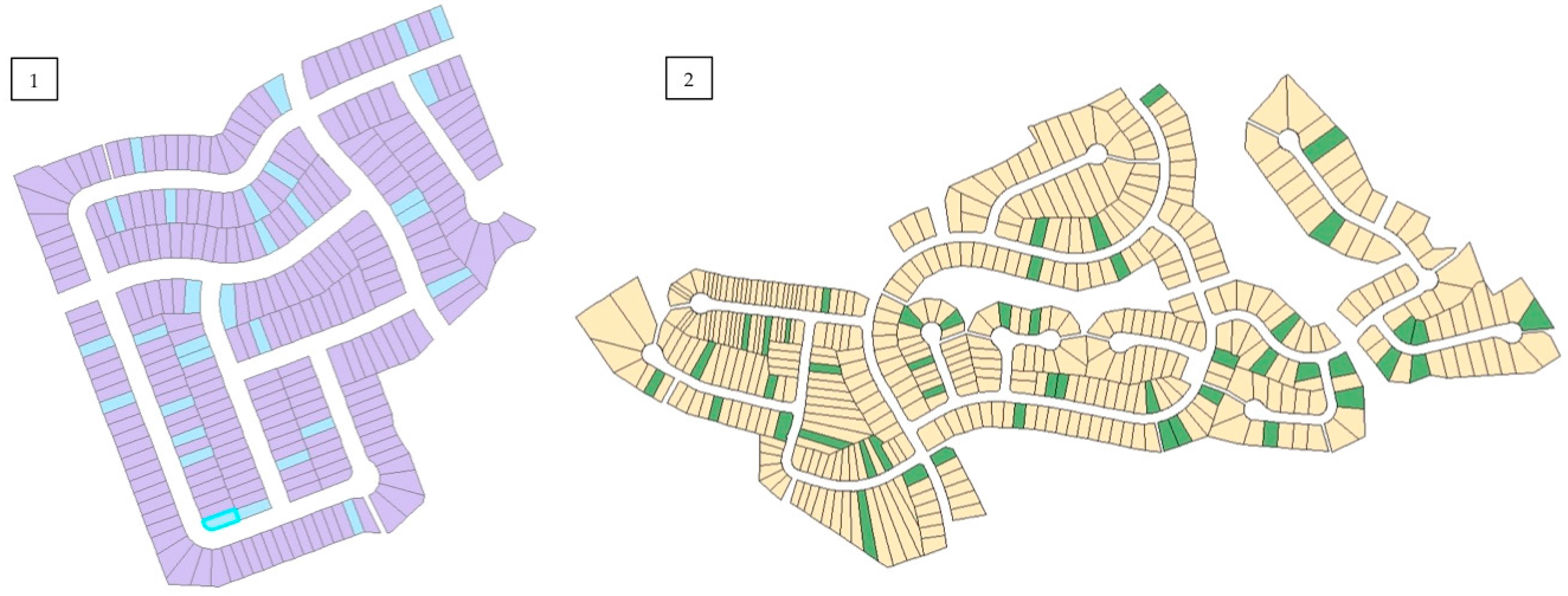
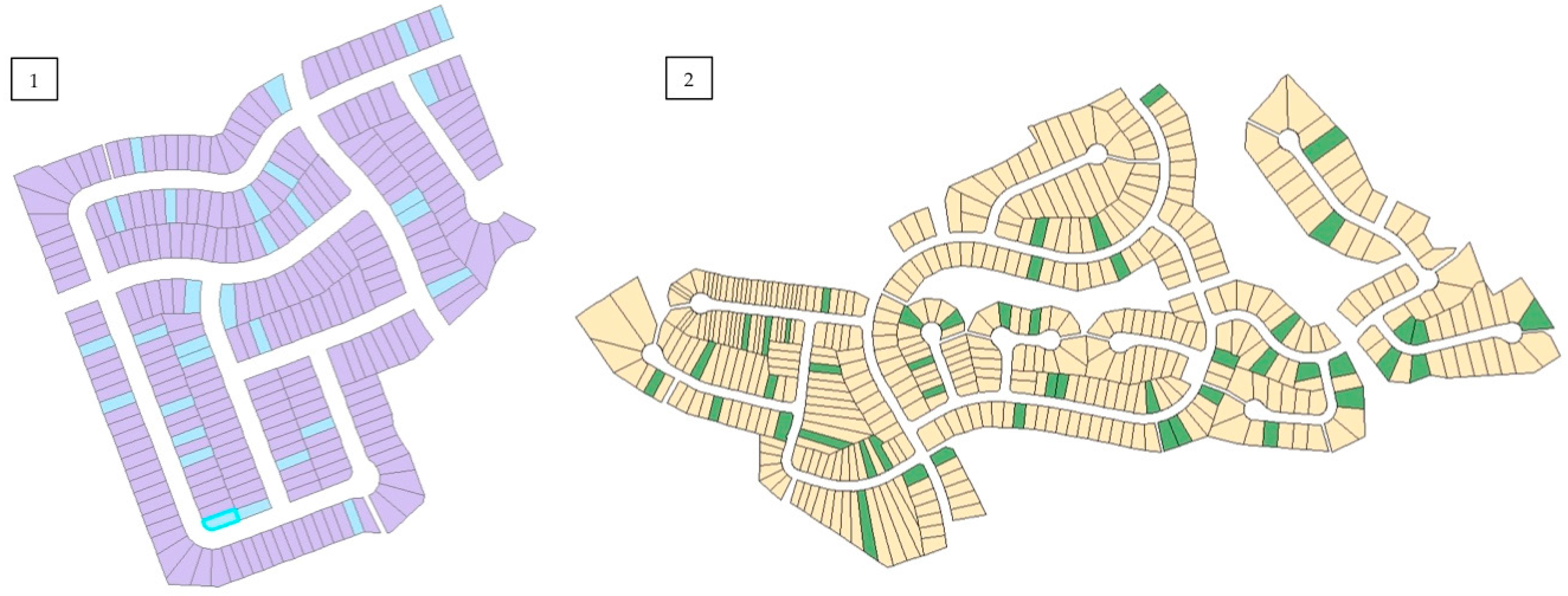
Figure 2). Other criteria for neighbourhood selection included size and general similarities in urban morphology related to street and sidewalk design, as well as presence of street trees and remnant woodland in Halifax. Approximately 10% of private property lots were randomly selected based on total residential property land area on ArcGIS© (version 10) using the parcel data layers from each city’s spatial database (
Figure 3). The residents of selected properties were given a letter describing the study and requesting permission to access their property. If permission was not granted, the home immediately next to the original selected property was chosen. All streets within the boundaries of each neighbourhood were examined. In Halifax, four remnant stands located adjacent to each residential neighbourhood were sampled using five plots of 10 m × 10 m (0.20 ha total remnant forest sampled). In London, two remnant woodland sites located outside city limits were sampled using three plots of 20 m × 20 m (0.24 ha total remnant forest sampled).
2.2. Data Collection
All sites in Halifax were visited between June and July 2015, while all sites in London were visited between June and August 2015. The street trees in each neighbourhood were censused. All trees within the boundaries of randomly selected residential properties and within each plot in remnant woodland stands were measured. For each tree, the species was identified and diameter at breast height (DBH) was measured at 1.4 m from the base of the trunk using a diameter tape or calipers, depending on the size of the tree. Dead trees and shrubs (woody plants with height <5 m at maturity) were not measured.
Street trees were defined as trees either found on the road verge located between the street and the sidewalk, or, in the case where there was no sidewalk, located between the curb and private residential properties. Private property trees were defined as those found on landscaped and maintained areas of residential properties.
2.3. Data Analysis
Tree species identified were denoted as “native” or “non-native” based on whether the natural range of the species is found in the province; London is found in Ontario, which has 85 native species, while Halifax is located in Nova Scotia with 42 native species [
21,
23]. Naturalized species that were introduced after European colonization, like Norway maple (
Acer platanoides), were considered non-native.
For each land type and neighbourhood, species richness was calculated by summing all identified tree species. Proportions of native and non-native species were calculated by dividing the count of native/non-native trees by the total count of trees measured.
Two indices were used to calculate species diversity [
24]. The first is the Shannon-Weaver index, which takes into account both species richness and evenness:
where
represents the count proportion of the
ith species. Two-sample
t-tests were carried out to determine whether results from this calculation varied significantly across neighbourhoods within and between cities.
The second diversity measure is the Simpson index of dominance, which describes the evenness of a community based on the probability that two randomly selected individuals belong to different species:
where
represents the count proportion of the
ith species.
Four separate chi-squared analyses were carried out on SPSS© (version 22) using 2 × 2 contingency tables to identify the factors that influence proportions of native and non-native trees on residential properties. For these calculations, tree data was combined from the two neighbourhoods of equal age class within the same city. Two variables were tested: the pre-urbanized landscape and the decade of neighbourhood development (or neighbourhood age). The tests compared counts of native and non-native trees in old neighbourhoods with different pre-urbanized landscapes, in new neighbourhoods with different pre-urbanized landscapes, in both old and new neighbourhoods in Halifax, and in both old and new neighbourhoods in London.
3. Results
3.1. Site Descriptions
The size of the neighbourhoods ranged from approximately 25 to 50 hectares. Newer neighbourhoods were generally comprised of smaller residential property lots compared to older neighbourhoods. Properties in all neighbourhoods examined contained detached or semi-detached homes surrounded by lawn. Many home owners chose to maintain garden beds or other landscaped areas to grow shrubs, hedges, herbaceous plants, and trees. Both newer and older neighbourhoods in Halifax retained some woodland area during development, including corridors located behind and between homes, and in some cases along pedestrian footpaths. Newer neighbourhoods in London were primarily surrounded by fields with some scattered forest corridors (
Figure 4). Some areas of an older neighbourhood in London bordered a ravine with some woodland, while the other older neighbourhood housed a small patch of regenerated woodland.
In Halifax, the remnant woodland sites were found adjacent to the subdivision, and were determined to be forested immediately prior to development, and remained so throughout the development process. Red maple (Acer rubrum) and red spruce dominated remnant sites in Halifax. Dominant trees were estimated to range between 50 and 100 years old. Other species sampled included yellow birch, striped maple (Acer pensylvanicum), and eastern hemlock (Tsuga canadensis). Two non-native species, Norway maple and European alder (Alnus glutinosa), were sampled in remnant plots located in close proximity to Birch Cove and Tam O’Shanter, respectively.
In London, remnant woodland was sampled at two sites located outside city limits. Dominant species included red maple, black cherry (Prunus serotina), and red oak (Quercus rubra); these trees were estimated to be between 100–200 years old. More uncommon species sampled included blue beech (Carpinus caroliniana), flowering dogwood (Cornus florida) and tulip tree.
3.2. Species Diversity
3.2.1. General Summary
In total, 82 tree species were identified in Halifax and 104 tree species were identified in London (
Table 3). In both cities, the total species richness of private properties was greatest, followed by streetscapes and remnant woodland. Across all land types, London had a higher number of native species compared to Halifax. However, the selected neighbourhoods in Halifax housed a larger proportion of native species (33 native species sampled/42 total native species within the area of Nova Scotia = 79%) compared to London (52/85 = 61%).
3.2.2. Street Trees
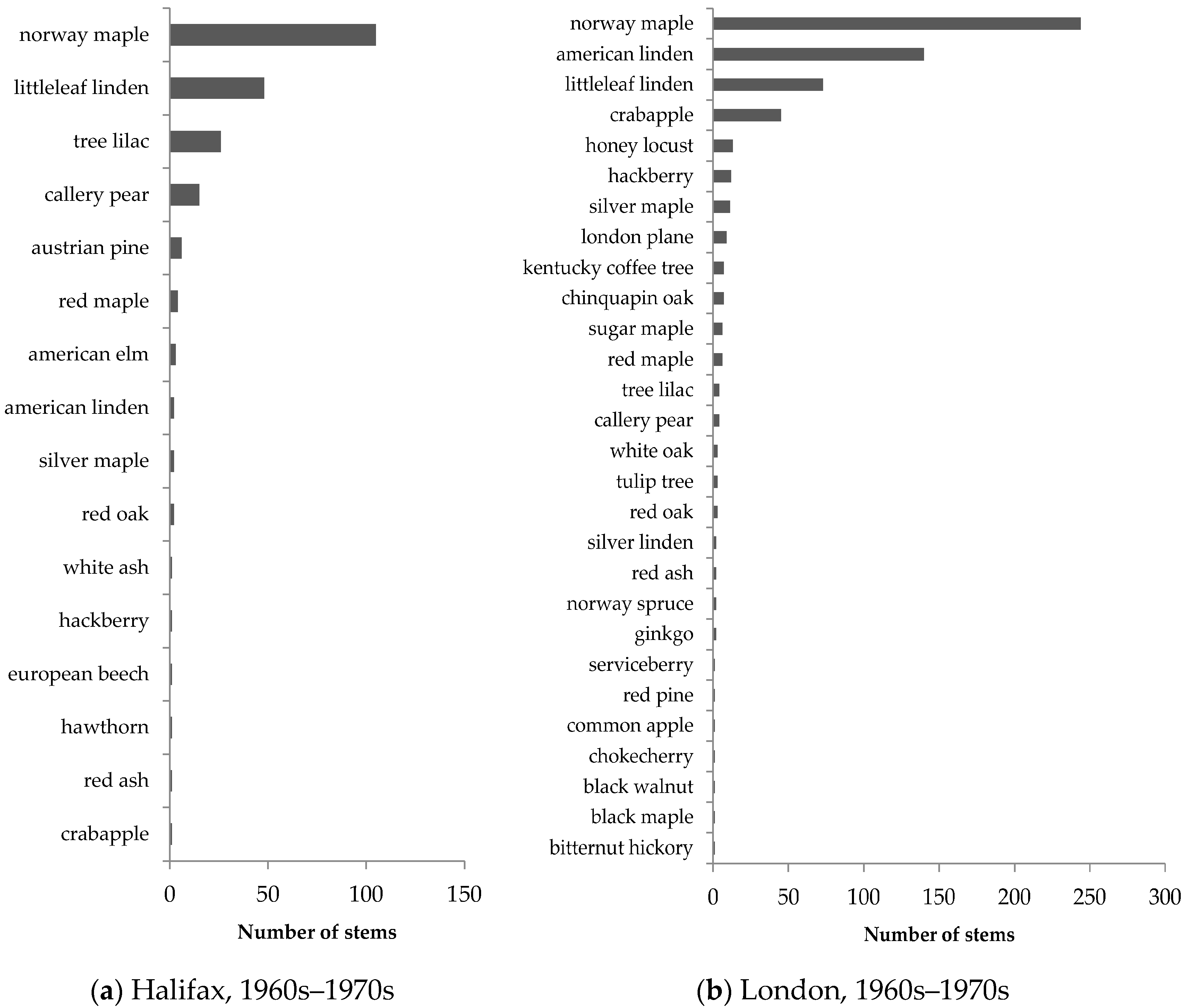
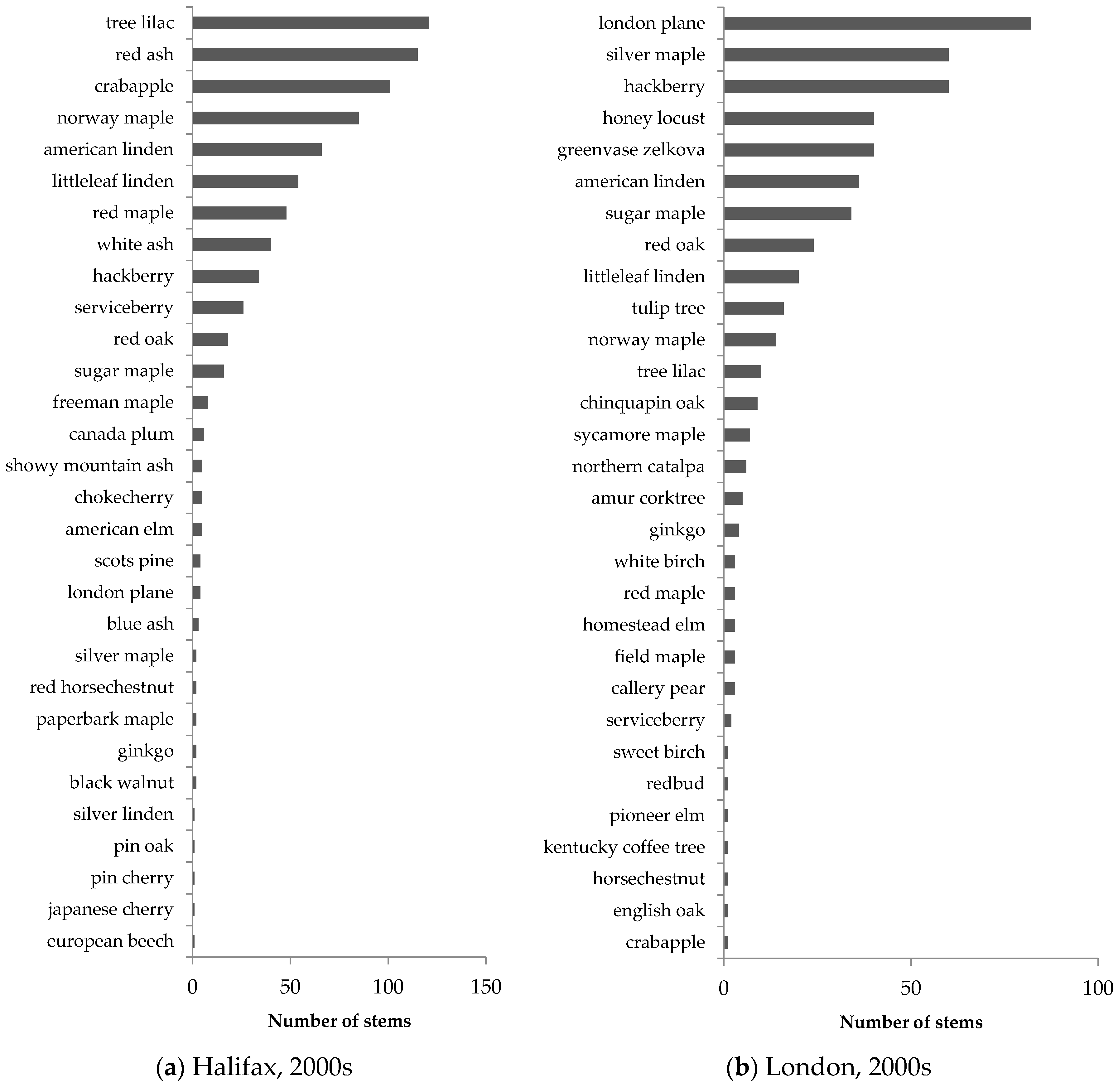
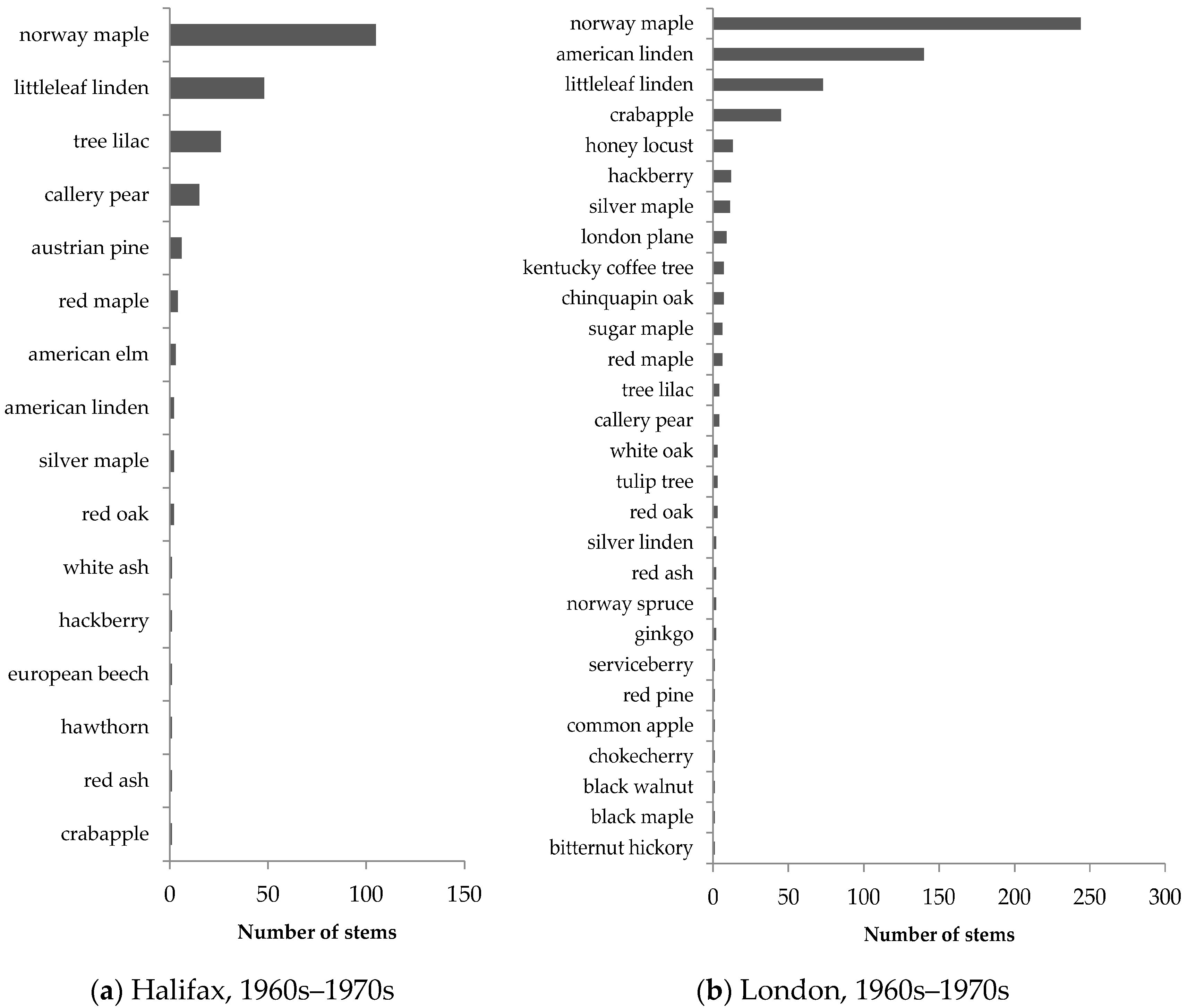
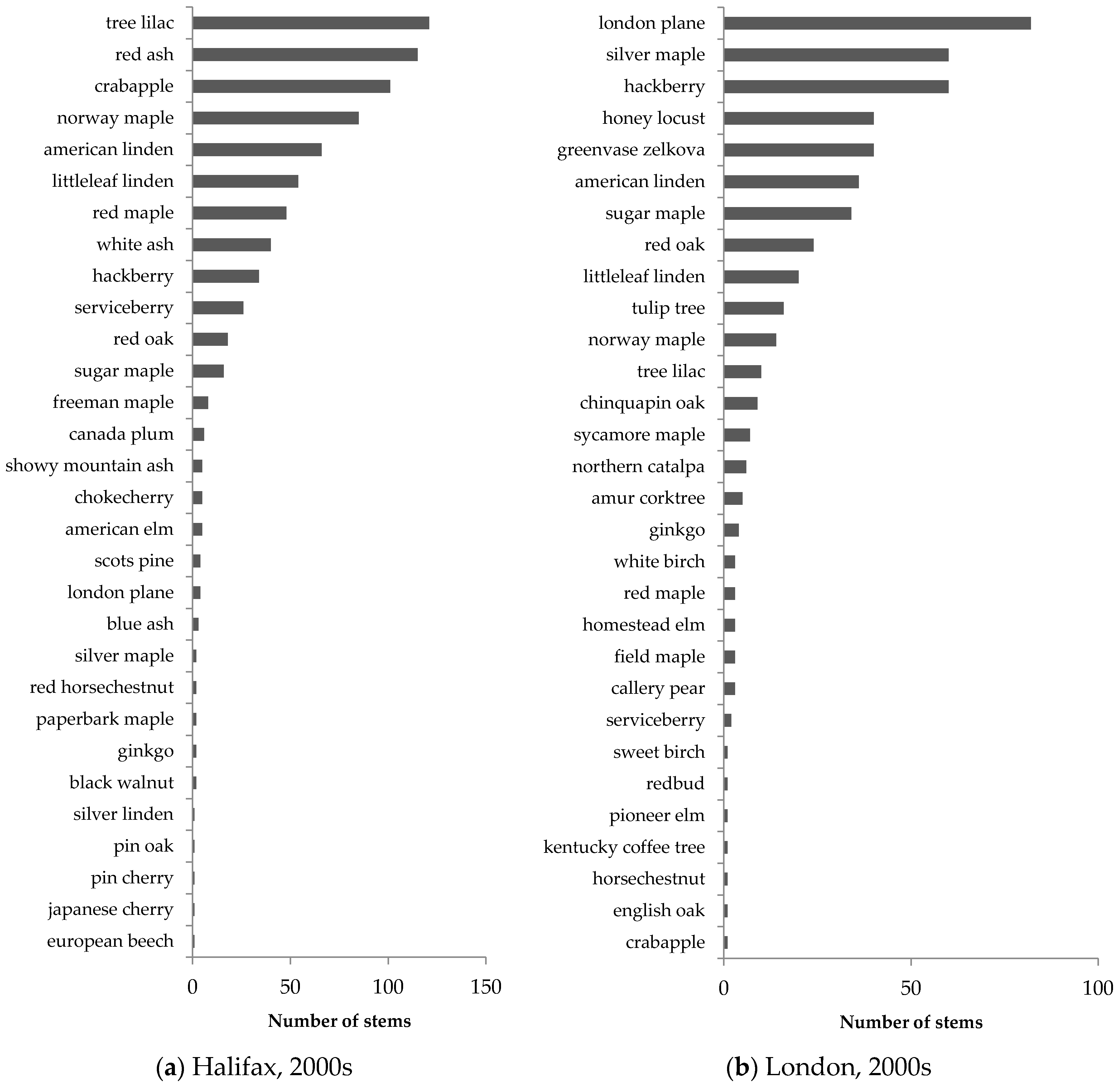
Similar patterns in street-tree composition were observed in Halifax and London (
Figure 5 and
Figure 6). Older neighbourhoods exhibited low species evenness and were dominated by non-native species, including Norway maple, littleleaf linden (
Tilia cordata), crabapple (
Malus spp.), and tree lilac (
Syringa reticulata). In streetscapes, Norway maple accounted for 48% of all older trees in Halifax and 40% of all older trees in London, illustrating its popularity in street-tree planting in Canadian neighbourhoods developed 40–50 years ago.
The results from the Shannon-Weaver diversity index calculation, which takes into account species richness and evenness, indicated that the diversity of streetscapes in older neighbourhoods in both cities is significantly different from streetscape diversity in newer neighbourhoods (
Table 4). A two-sample
t-test was used to compare differences in the Shannon index calculations (e.g., diversity) of street trees between older (mean = 1.30, SE = ±0.040) and newer (mean = 2.56, SE = ±0.105) neighbourhoods in Halifax (
t = 11.17,
p = 0.008, d.f. = 2). The same test found a significant difference between the diversity of street trees in older neighbourhoods (mean = 1.83, SE = ±0.040) and newer neighbourhoods (mean = 2.58, SE = ±0.055) in London (
t = 10.95,
p = 0.008, d.f. = 2). Older streetscapes in London were slightly more diverse than older streetscapes in Halifax (
t = 9.37,
p = 0.011, d.f. = 2), while a similar result was not found between newer streetscapes in both cities.
Street trees in newer neighbourhoods exhibited much greater species evenness, as illustrated by the more gradual slope in the species abundance distribution (
Figure 6). The representation of Norway maple was also much smaller, accounting for approximately 11% of all newer street trees in Halifax and 3% of all newer street trees in London. Native stem count proportions were also higher in newer neighbourhoods compared to older ones, indicating that native species are more recently preferred for planting in city streets (
Table 4).
3.2.3. Residential Property Trees
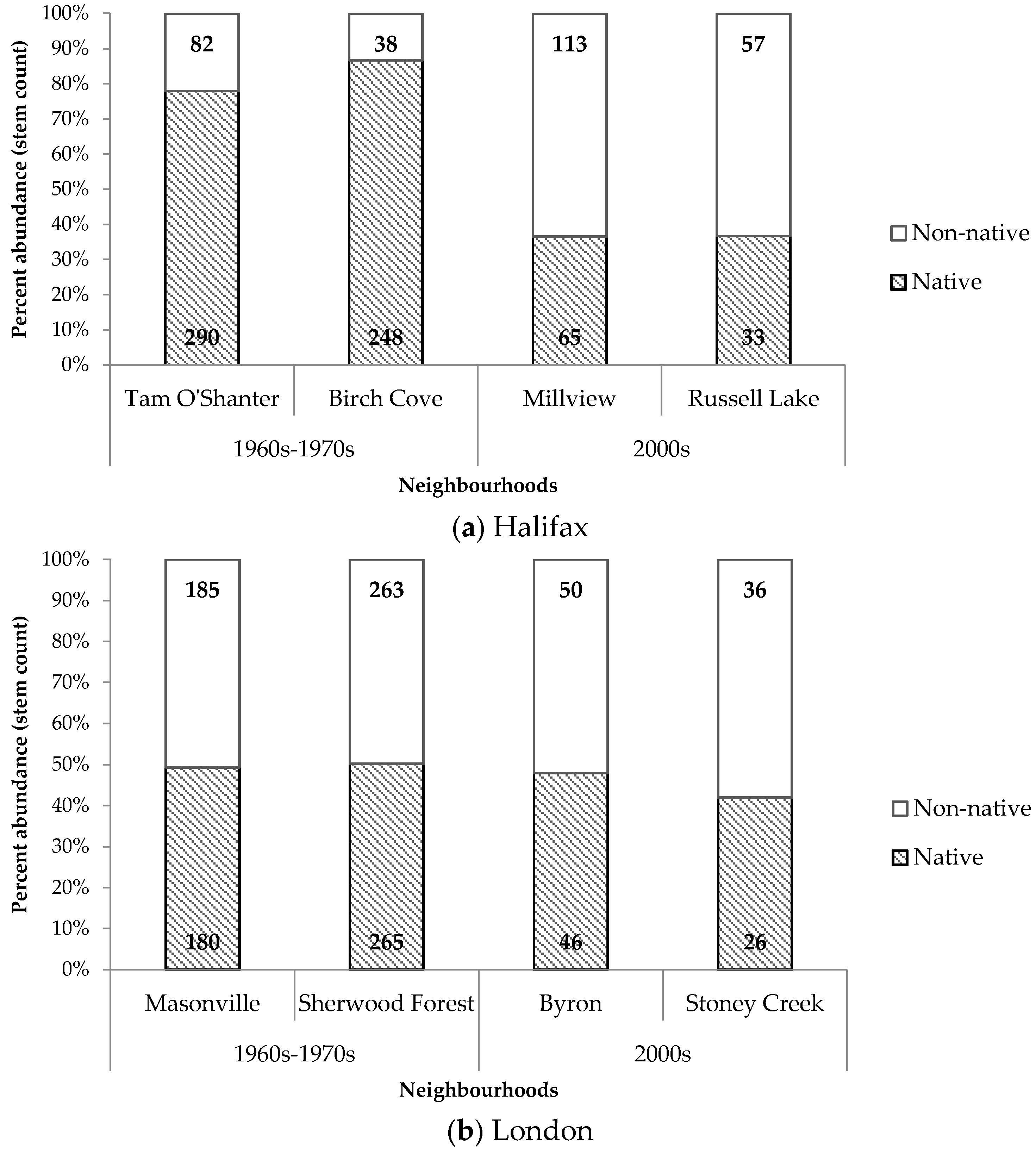
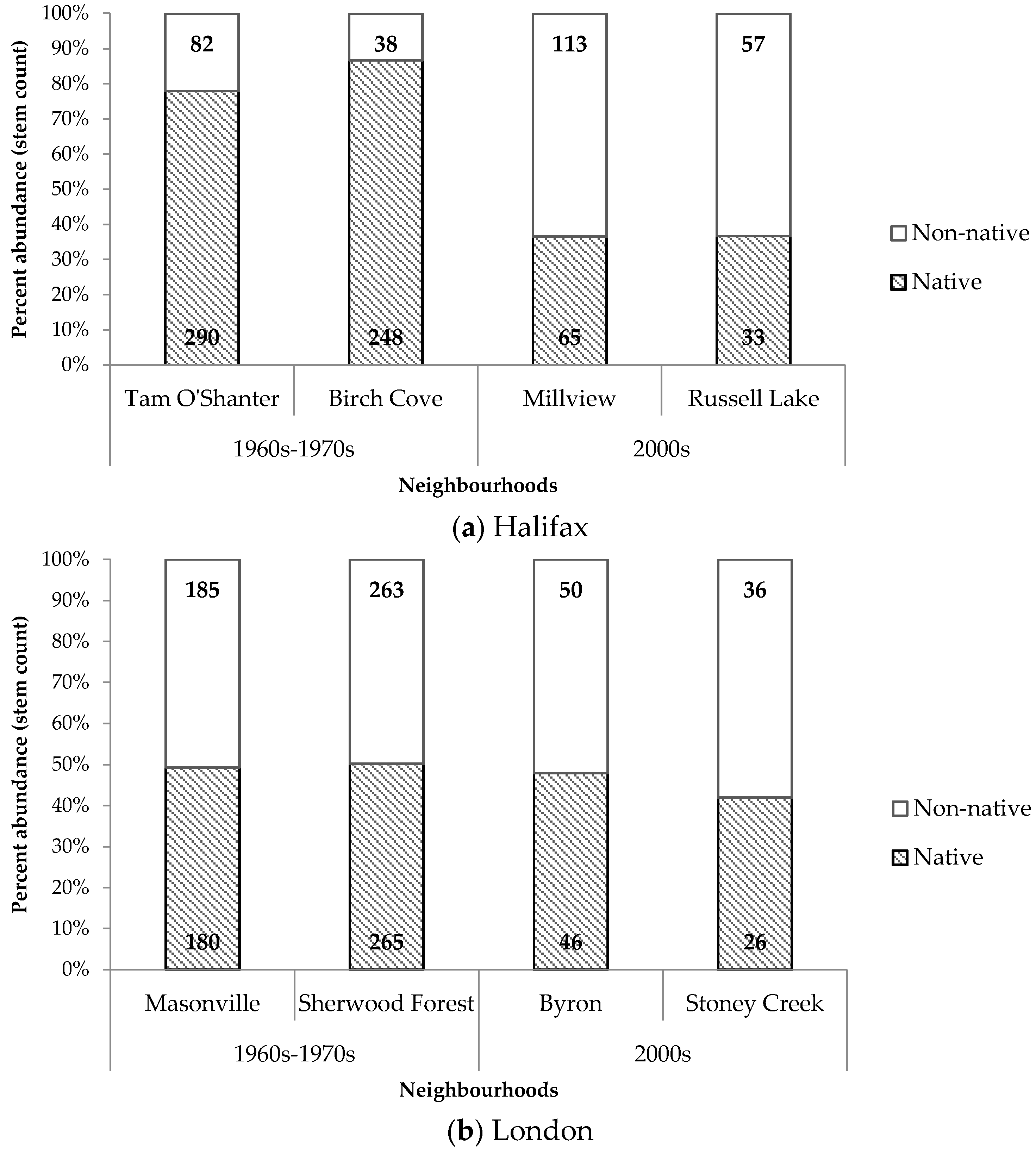
Contrasting patterns in the composition and diversity of residential property trees were observed between the two cities. In Halifax, older neighbourhoods had a significantly greater proportion of native trees compared to newer neighbourhoods (
Figure 7). Older neighbourhoods in Halifax also displayed higher proportions of native trees compared to older neighbourhoods in London. In London, there were no notable difference between proportions of native and non-native trees across old and new neighbourhoods.
No statistical difference was observed between the diversity of property trees in older and newer neighbourhoods within each city (
Table 5). However, a statistical difference did exist (
t = 5.77,
p = 0.029, d.f. = 2) between older neighbourhoods in Halifax (mean = 2.44, SE = ±0.155) and older neighbourhoods in London (mean = 3.46, SE = ±0.085). A similar relationship was not observed between newer neighbourhoods in both cities.
3.3. Size-Class Diversity
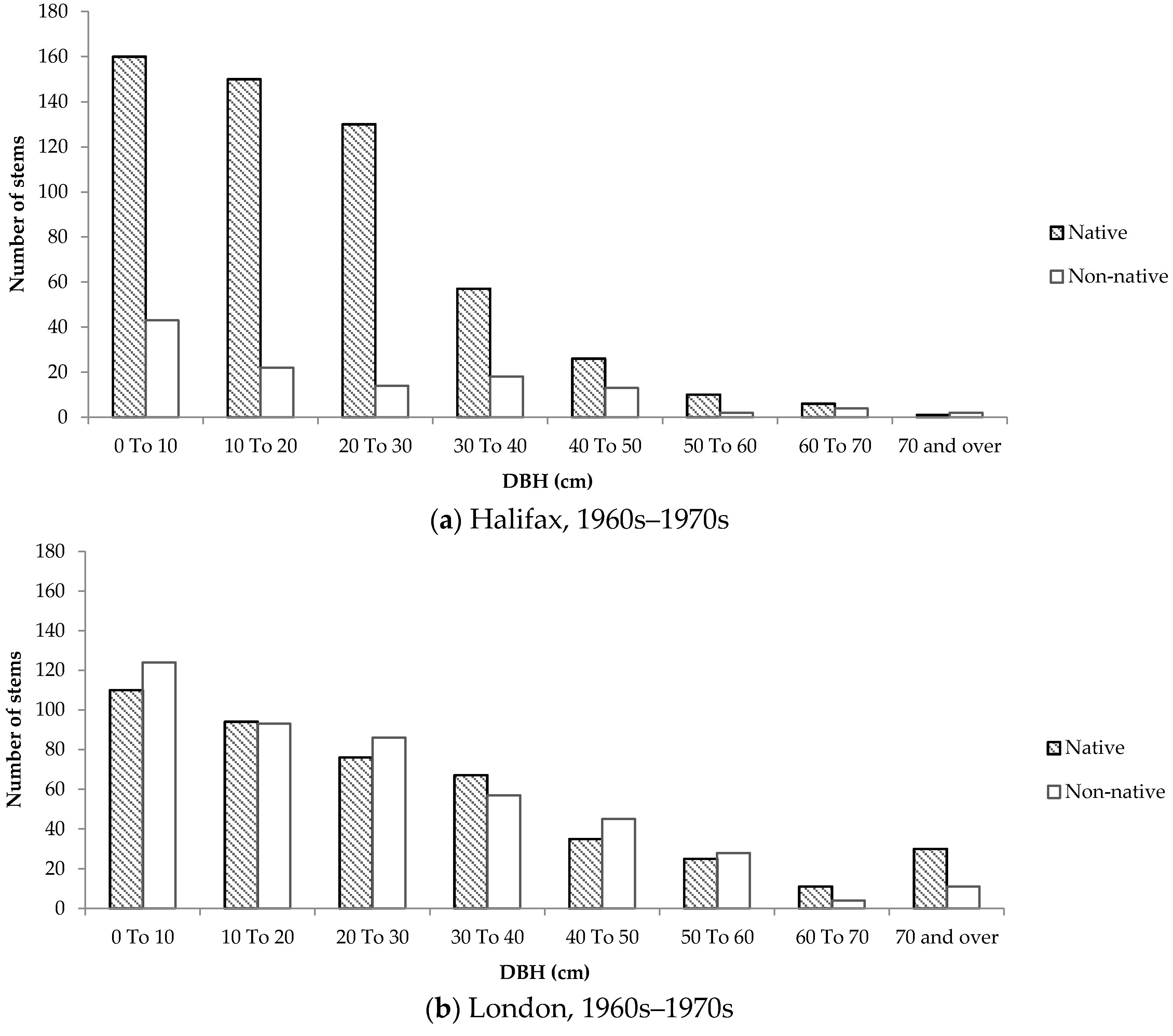
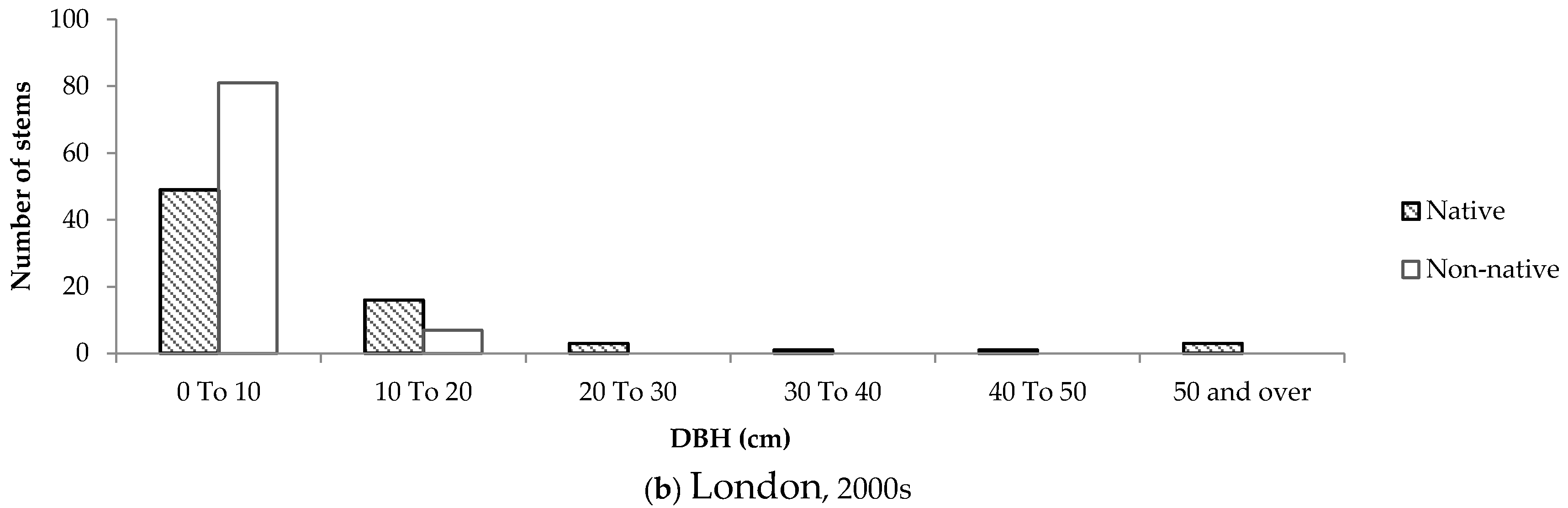
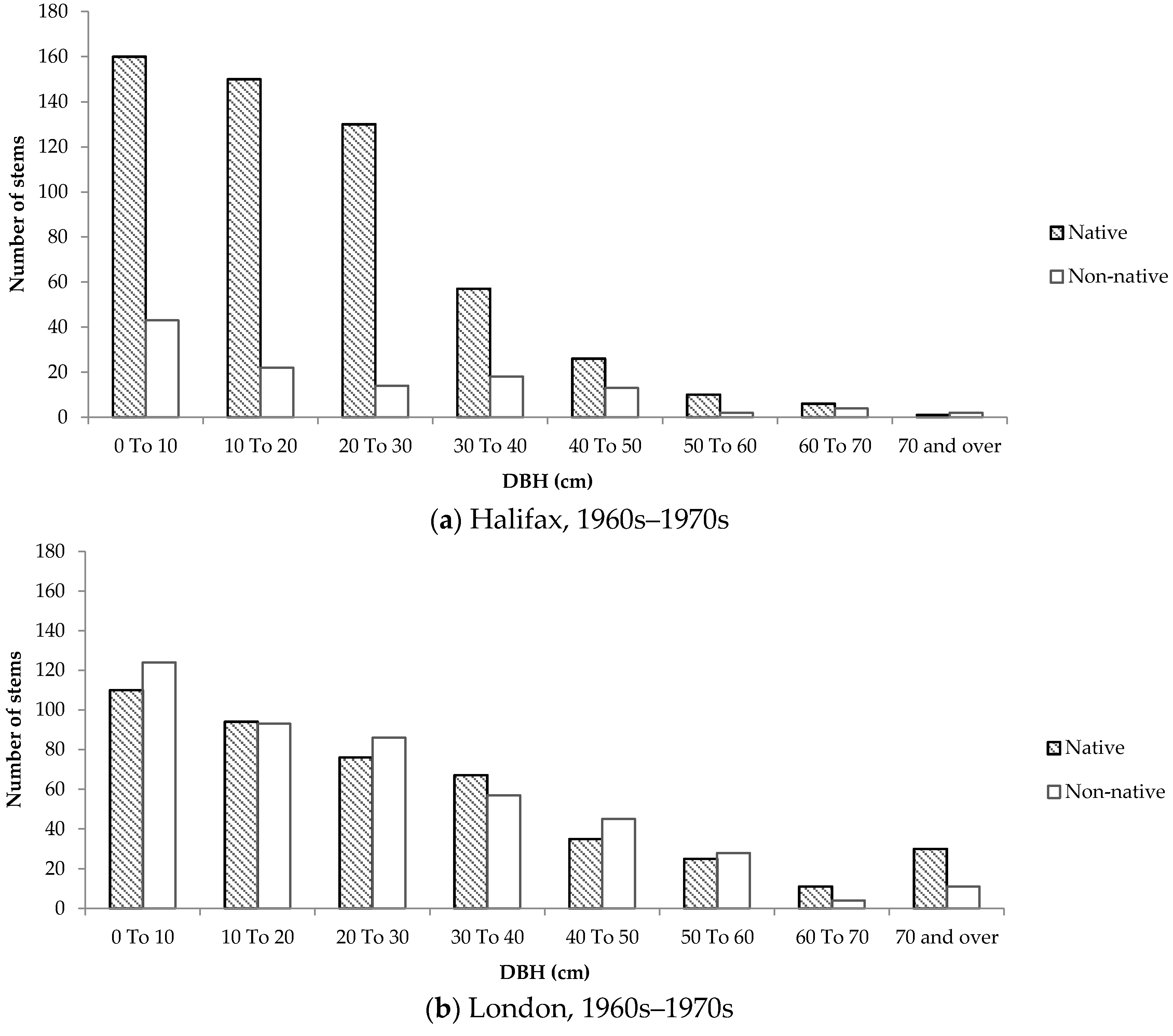
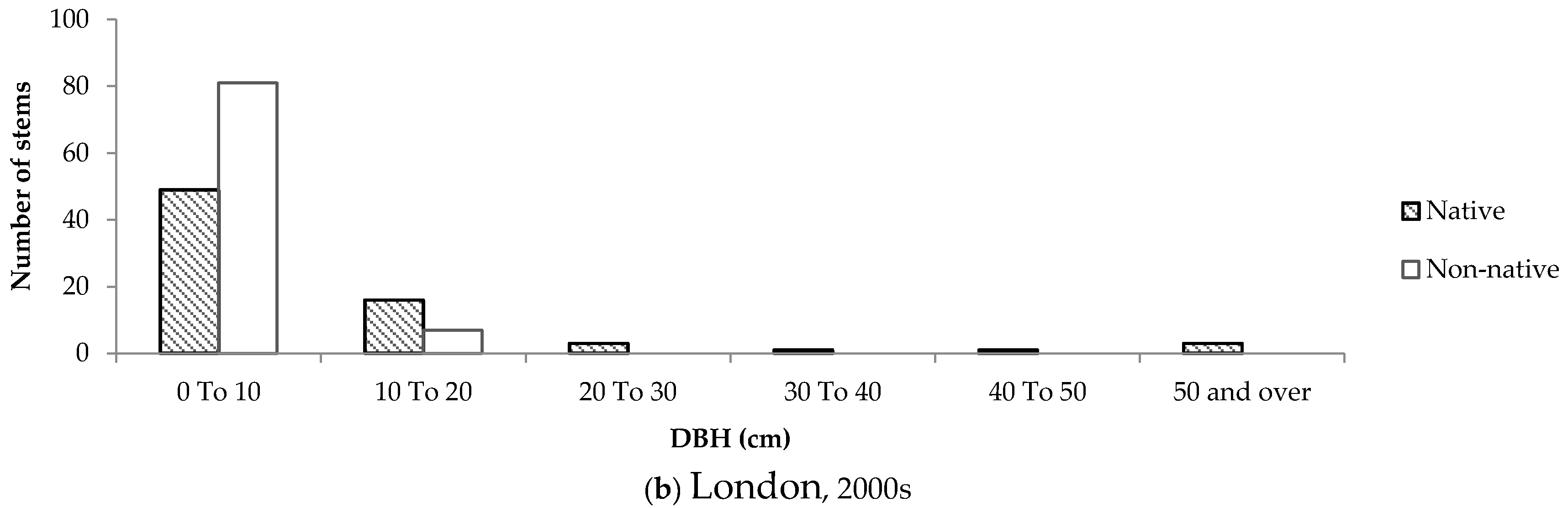
Size-class distributions were created to determine the composition of native and non-native trees according to DBH on residential properties, and to serve as a proxy for tree age. Although all size-class distributions were positively skewed, the distributions representing trees in older neighbourhoods differed considerably between the two cities.
In older neighbourhoods in Halifax, the distribution was more positively skewed compared to London. In Halifax, approximately 80% of small trees (DBH < 10cm) were native. In comparison, only 47% of trees of the same size class in older neighbourhoods in London were native. Furthermore, the number of native and non-native stems remained relatively equal and consistent across size classes in London (
Figure 8). Conversely, in Halifax almost 60% of all native trees had a DBH below 20 cm.
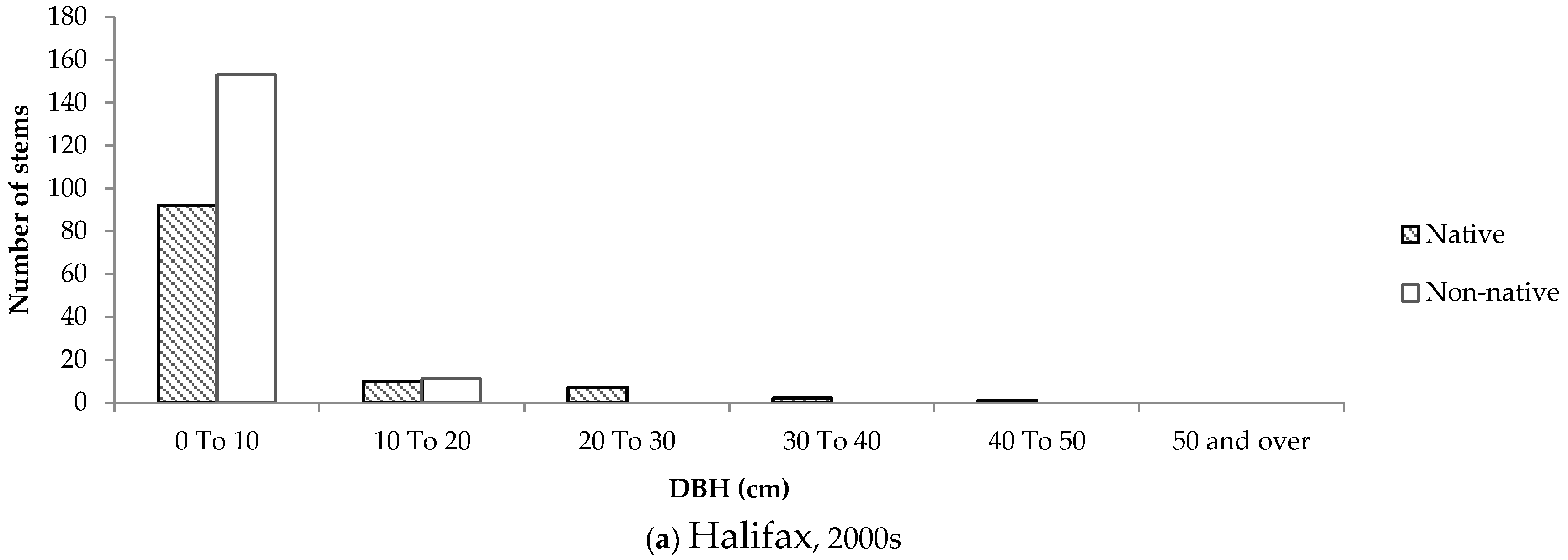
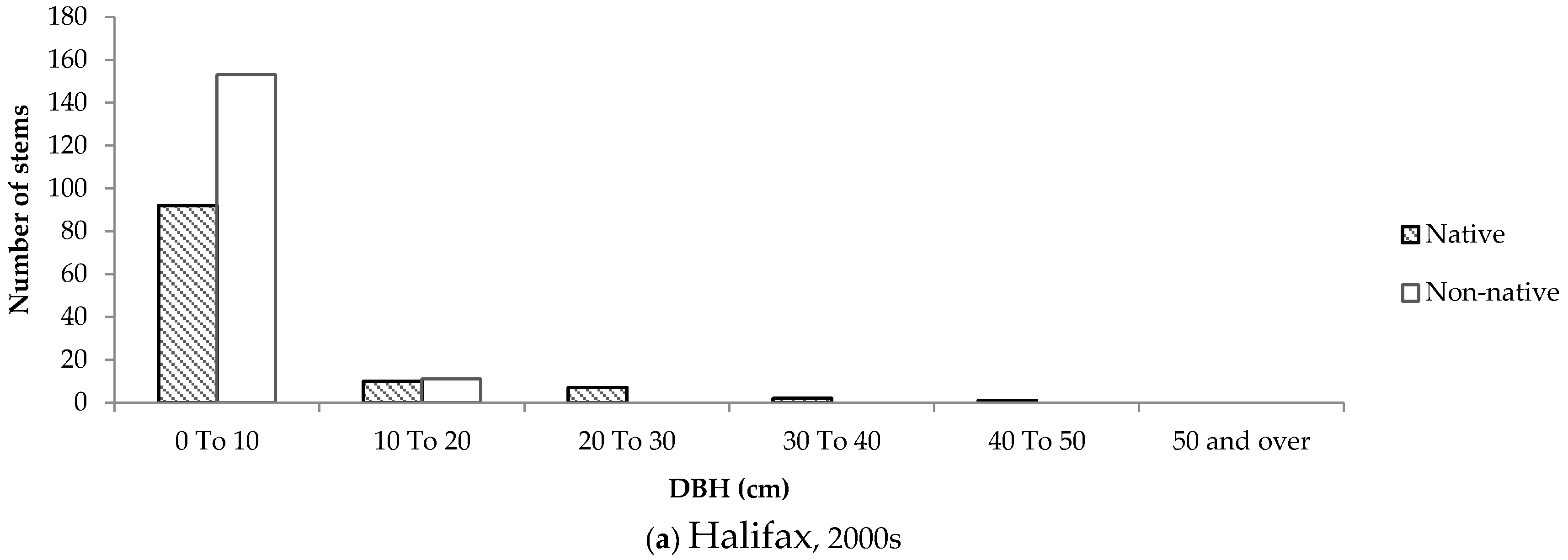
In newer neighbourhoods, the size-class distribution of native and non-native trees was similar across both cities (
Figure 9). Some older native trees were left standing in developments in both cities, while non-native trees dominated newly planted stems.
3.4. Chi-Squared Analysis
Four contingency tables were used to analyze the relationship between two factors (the pre-urbanized landscape and age of development) and counts of native and non-native trees on residential properties (
Table A1,
Table A2,
Table A3 and
Table A4). To review, the pre-urbanized landscape in Halifax is woodland, while the pre-urbanized landscape in London is field. A strong and statistically significant relationship was found between the type of pre-urbanized landscape and counts of native and non-native trees in neighbourhoods developed during the 1960s–1970s (
Table 6). Halifax, developed onto woodland, had a significantly greater number of native trees compared to non-native trees in older neighbourhoods. London, developed onto field, had virtually equivalent numbers of native and non-native trees. The equivalent relationship in newer neighbourhoods was not found to be significant. Another important relationship existed in Halifax, but not in London, between decade of development and counts of native and non-native trees. Newer neighbourhoods in Halifax were dominated by non-native trees, while the opposite occurred in older neighbourhoods. In comparison, count proportions of native and non-native trees remained consistent across new and old neighbourhoods in London.
4. Discussion
4.1. Development and Tree-Species Composition
The various land types that are found in suburban areas reflect differences in tree establishment, planting practices, maintenance, and ownership, and are thus influenced by different drivers of tree-species composition. The sites sampled in this study ranged from near-natural remnant forests to the urbanized streetscape. In both cities, streets in older neighbourhoods were dominated by only a few species, including the ubiquitous Norway maple. In comparison, streets in newer neighbourhoods had greater species richness and evenness as well as a better representation of native species, which have been shown to support insect and bird diversity more effectively than non-native species [
3,
25].
Remnant stands represented the most “natural” forest habitat found in suburban areas, and contained few (if any) non-native trees. Nevertheless, plots in Halifax supported two non-native tree species, both of which are invasive. Given the close proximity of the remnant sites to the adjacent residential area, it is probable that these species were planted on private property some time after development occurred, and managed to disperse into the remnant stands.
Many residential properties were intensively landscaped, and housed common ornamental non-native species like Norway maple, blue spruce (
Picea pungens), and Japanese maple (
Acer palmatum). Properties in newer neighbourhoods had fewer trees, of which a majority were non-native species. Conversely, properties in older neighbourhoods in Halifax held a large amount of native trees and resembled the remnant woodland sites more closely than properties in newer developments in terms of species composition. The size-class distributions suggest that some trees were kept during development in neighbourhoods in both cities; it is likely that large (and old) non-native trees found in London were planted on farms or along fence rows. These patterns illustrate the importance of examining changes in species distributions across remnant forests and adjacent green spaces, particularly as some non-native species can threaten native habitats and potentially displace native species [
2,
11].
The following sections elaborate on similarities and differences observed in the tree-species composition in streetscapes and on private properties in both cities.
4.2. Policy and Administration of Street Trees
Street trees are generally planted by a developer or contractor, and are maintained by the municipality. Despite the contrasting development patterns (e.g., the pre-urbanized landscape) of the selected neighbourhoods, the patterns exhibited in street-tree communities observed between the two cities are very similar. Given street-tree planting practices, it is certain that political and administrative factors play a large role in determining the types of trees found in city streetscapes.
The drastic difference between street-tree composition in newer
versus older neighbourhoods in both cities could reflect changes in urban forest management and policy that have occurred in the last few decades. The infiltration of diseases and insects like Dutch elm disease and the emerald ash borer resulted in mass tree mortality rates in Canada in the last century, and has heightened awareness of the importance of diversifying planting stock [
26,
27,
28]. Many urban forest management plans created more recently have developed targets to enhance native and overall species diversity, illustrating concerns about invasion events and urban forest canopy loss [
5]. Higher evenness and native species representation in newer neighbourhoods in both cities may reflect a growing diversification trend among urban forest practitioners across Canada.
The differences observed in street-tree diversity between older and newer neighbourhoods could also be due in part to changes in nursery stock availability over time. Large-scale projects, like street-tree planting in new subdivision developments, often acquire trees from regional wholesalers instead of local nurseries [
29]. At a given time, species selection may be limited due to location, resources, or customer demand; some tree species requested by the developer or urban forester may be substituted or outsourced, potentially resulting in more unfavourable species choices and reducing diversity enhancements [
30]. However, some research in the United States has shown that nurseries are offering more species choices today compared to decades ago [
31]. More recent demand for a wider range of species may therefore be influencing nurseries and wholesalers to diversify their stocks, in turn promoting higher species richness and evenness as well as native species representation in newer subdivision developments.
Various practitioners are both directly and indirectly involved in the process of street-tree species selection and planting, including municipal officials, arborists, urban planners, foresters, developers, and contractors. These groups reflect differences in expertise, values, priorities, and professional paradigms [
29,
32]; these in turn can dictate the species composition of street trees. For example, the popularity of tree lilac and London plane (
Platanus ×
acerifolia) in newer neighbourhoods could be due to the preference of a particular urban forester or planner, based on factors such as tolerance to urban stresses, maintenance, growth rate, shade value, and cost.
4.3. The Influence of Remnant Woodland on Private Properties
Although little research has been carried out on the subject, it has been shown that development patterns, including the landscape present prior to urbanization, can influence the composition of urban tree species. One study found that residential neighbourhoods developed from forested areas had higher tree species richness and a greater proportion of native species compared to regions developed from prairie land [
33]. Researchers have also suggested that the presence of remnant woodland located within or adjacent to residential areas can promote native species establishment and representation within the urban landscape [
34,
35].
The results of this study seem to illustrate these processes to some extent. The significant difference between both cities in native and non-native stem counts in older neighbourhoods suggests that native trees in Halifax may be dispersing from woodland areas retained during and after development onto adjacent residential properties. Furthermore, the much higher number of native trees found on older residential properties compared to newer ones in Halifax potentially demonstrates a lag effect; the tree-species composition of newer neighbourhoods may be influenced primarily by home owner planting preferences, while the species composition of older neighbourhoods may be shaped both by home owner planting decisions and the natural establishment of some trees over time as the development aged.
The size-class distributions for Halifax show that the majority of native trees are smaller, suggesting that many stems appeared on residential properties a while after development occurred. This phenomenon was not observed to the same degree in London, which could be due to the lack of remnant woodland in and around the selected neighbourhoods. The size-class distributions of native and non-native trees in London are very consistent across new and old neighbourhoods, therefore it is more likely that the species composition on private properties is influenced primarily by the planting attitudes and preferences expressed by home owners. Furthermore, the strikingly similar size-class distributions of trees in newer neighbourhoods in Halifax and London imply that the influence of remnant woodland is neither felt during nor immediately after development, but rather over time as trees are gradually given the chance to disperse into adjacent green space.
Despite the fact that private properties in Halifax had a better representation of native species, properties in London were generally more diverse, both in terms of species richness and evenness. This pattern could be attributed to differences in tree establishment; the tree-species composition of gardens in London mostly reflects individual planting preferences, and many home owners prefer non-native ornamental species for horticultural purposes, which contribute to overall species diversity [
2,
13,
36]. In a similar vein, this trend could also explain why the diversity measures characterizing newer neighbourhoods in Halifax and London are more similar compared to the older neighbourhoods, where naturalization is likely occurring in Halifax, but not in London, due to native species dispersal.
Depending on diversity targets and priorities, neighbourhoods in cities like Halifax may benefit from retaining and protecting forest buffers and patches during and after development. Not only are remnant areas important for promoting native tree-species establishment, but other organisms may benefit from retaining woodland during development, illustrating the crucial role that urban forests play in biodiversity conservation [
7].
4.4. Home Owner Attitudes and Opportunities for Diversity Enhancement
In both cities, residential properties were found to be most diverse compared to other land types, confirming the results from other studies on land tenure and tree-species composition [
2,
4,
37]. Home owner attitudes and preferences likely explain the diverse range of planting decisions; many social and economic factors taking place at the property- and neighbourhood-level can influence residential species diversity, such as income [
38], education [
39], ethnicity and nationality [
40], and planting fads [
41]. These patterns emphasize the potential for private yards and gardens to contribute to urban forest biodiversity [
34,
42]. It is thus important to consider how landscape conversion and development history could influence private property trees, particularly in the context of biodiversity management and enhancement.
In the context of native species diversity, it is worthwhile to explore home owner attitudes towards native species establishment, and whether residents knowingly allow native trees to establish on their property. Research has shown that although urban dwellers may in theory support the planting of native species, they might not be knowledgeable enough to identify native trees and recognize their ecological importance [
34]. Although results from this study have shown that native trees are probably dispersing onto private properties, the exact establishment mechanisms and potential decisions on the part of home owners have not been identified.
Subdivisions developed in cities like London, where most forested area has already been converted into agricultural fields, may benefit from targeted education initiatives and planting projects geared towards increasing native species representation and overall species richness, depending on the diversity goals of the municipality in question. Educating residents about the benefits of both native and non-native species may result in more informed planting choices, and in turn could influence the availability of local nursery stock [
29]. Education initiatives geared towards nursery workers and wholesalers could also influence resource availability and encourage the dissemination of information about tree diversity and its benefits to customers [
43]. London in particular has a relatively broad palette of native species to choose from, and encouraging a wider range of species selection on residential properties could give rise to diversity benefits at the city level.
Researchers have stressed the importance of increasing green space connectivity in urban and suburban areas for the purpose of increasing resilience to environmental change, providing wildlife habitat, and enhancing biodiversity [
44]. Regardless of pre-urbanized landscape, suburban areas may effectively enhance diversity if the design of residential properties and neighbourhoods is considered and incorporated into biodiversity initiatives or urban forest management plans implemented at the city level [
42]. It is worth noting that subdivision design should consider the possible negative impacts of residential encroachment onto remnant woodland, including loss and/or destruction of forest habitat, waste disposal and littering, and introductions of exotic species [
45]. Neighbourhood-level urban forest strategies could educate residents about remnant forest protection (particularly in riparian areas) and identify specific biodiversity issues and opportunities at the community level, encouraging partnerships between municipalities, planners, developers, researchers, and residents [
6,
27,
42]. Citizen engagement can be particularly useful in fostering public stewardship of the urban forest and in achieving municipal tree diversity goals [
6].
4.5. Limitations, Challenges, and Future Research
The advantages of this study design relate to the large sample size of trees measured on different land types, allowing for robust analyses and conclusions about species composition at the neighbourhood level. However, it is difficult to generalize the results of this study to other residential areas or other cities, as only four neighbourhoods in each city were examined. Furthermore, this study did not examine parks, a land type that has been extensively researched and plays an important role in urban forest diversity enhancement and management [
46,
47,
48].
Characterizing urban forest diversity is rendered more complex with the introduction of different land types (e.g., streets, private properties, remnant stands) that each require particular sampling protocols in order to effectively capture trends related to land use. Given the sampling design, the areas investigated for each land type in each neighbourhood are necessarily different. Species accumulation curves are included in the
supplementary material, but should be interpreted with caution given the fact that urban species composition does not always reflect natural forest establishment and succession patterns that conform to traditional species-area relationships [
49].
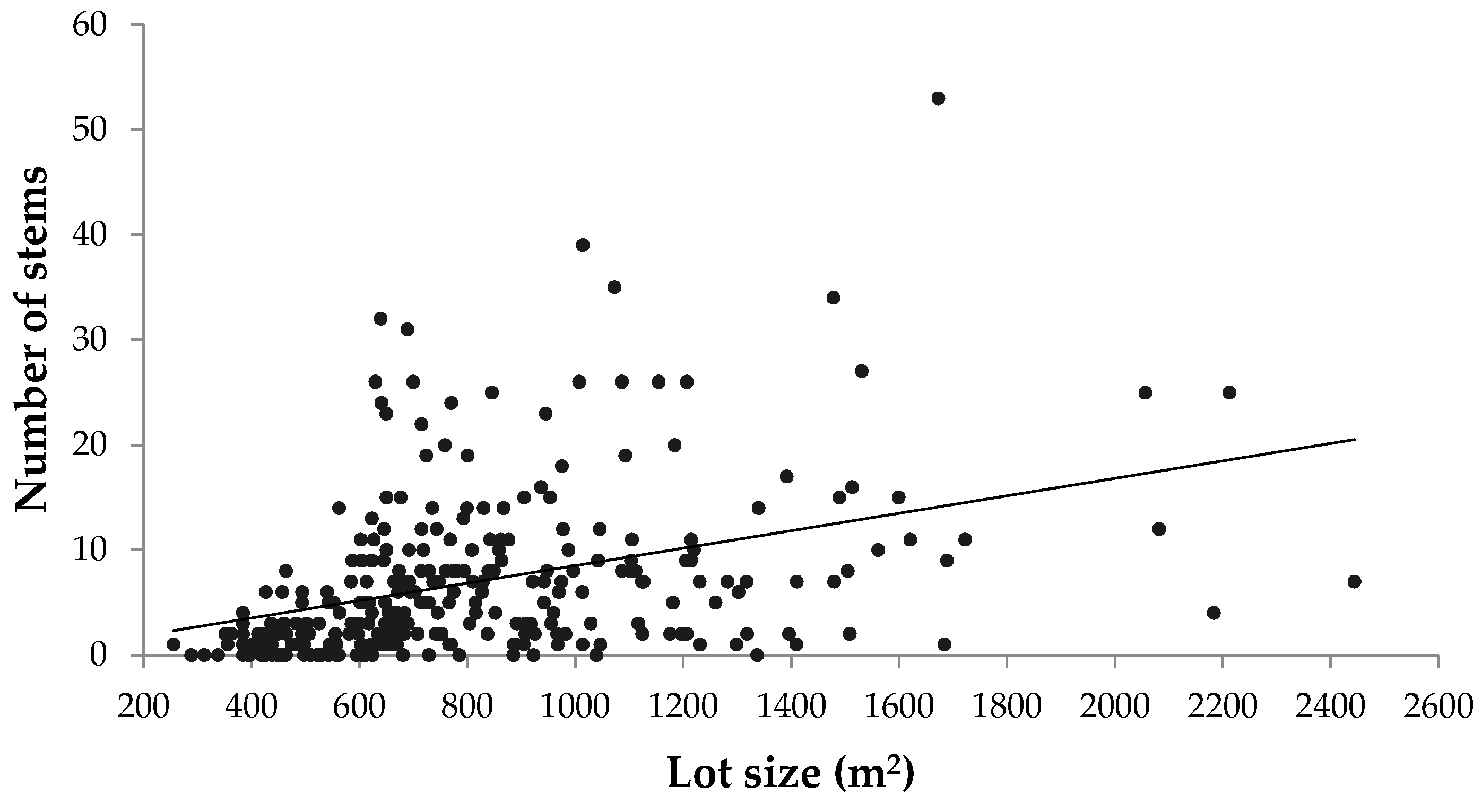
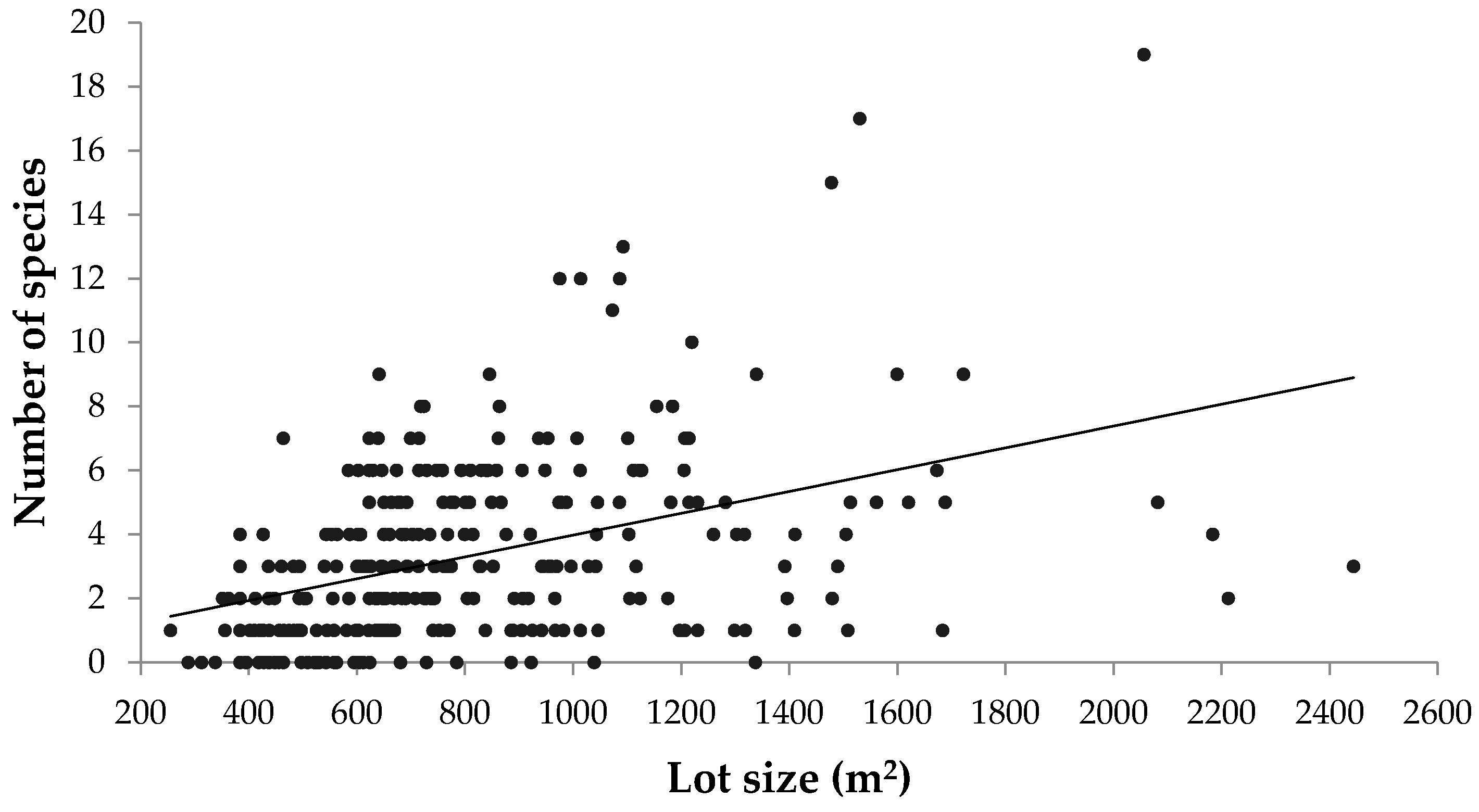
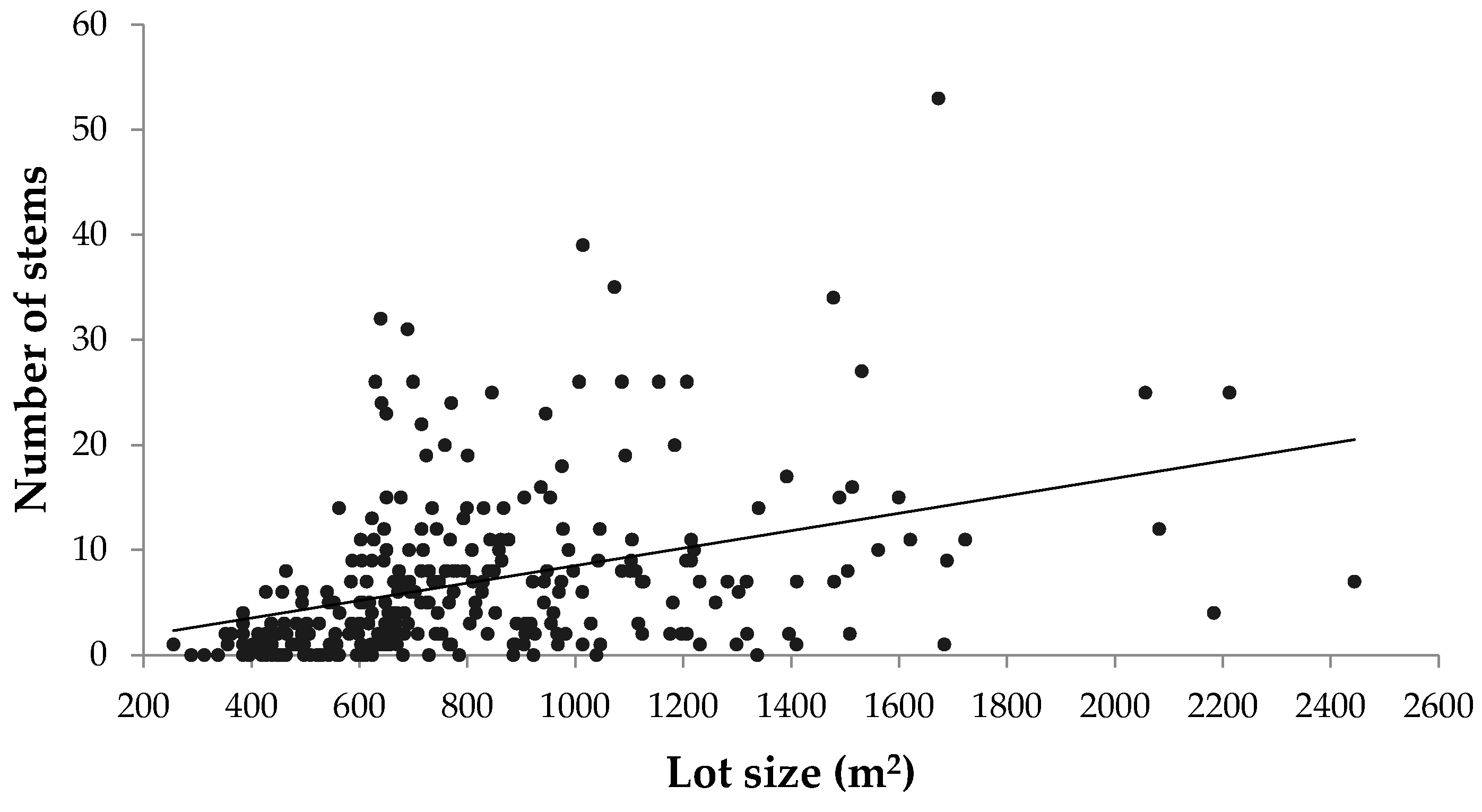
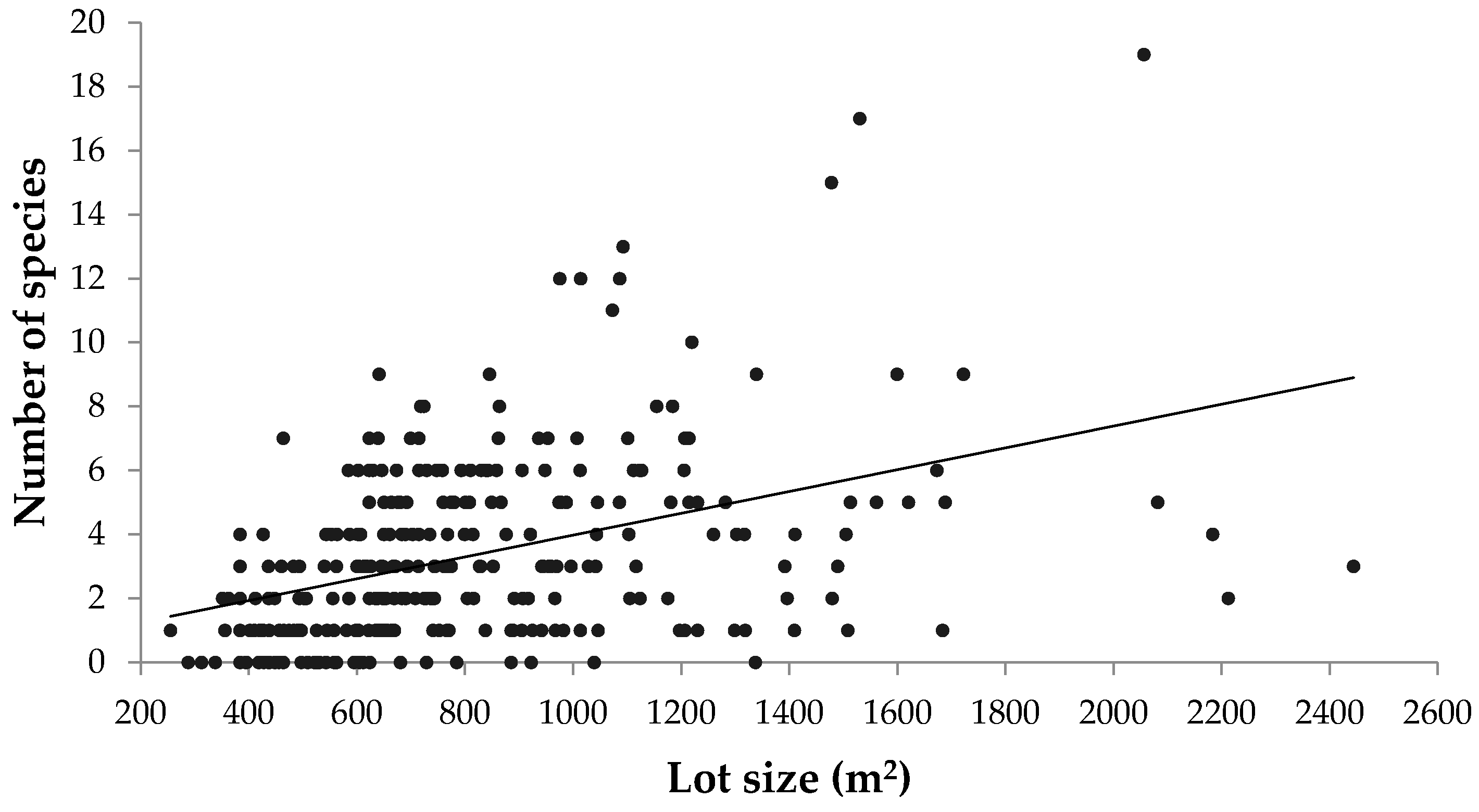
Although this study focused on the pre-urbanized landscape as a major driver of urban forest diversity, there are many other biophysical, socioeconomic, and political factors that influence urban tree diversity. In residential areas, lot size, which was not controlled for in this case, has been shown to correlate with urban tree diversity [
50,
51]. However, the results from this study showed a negligible correlation between property lot size and both number of stems and species richness (
Appendix Figure A4 and
Figure A5). Exploring how other drivers pertaining to urban morphology and neighbourhood design could promote urban forest diversity and green space connectivity would also be an important endeavour, particularly as suburban areas continue to expand in Canadian metropolitan areas.
Future research should explore home owner decisions about species selection for planting and the natural establishment of native trees on residential properties from remnant woodland. It would be worthwhile to determine whether residents purposefully encourage native species establishment on private property, and what kind of motivations are involved. Furthermore, creating and implementing education and planting initiatives geared towards home owners and community groups could shed light on the types of strategies that are most effective for biodiversity promotion in residential areas.
5. Conclusions
The results of this study emphasize the importance of considering both spatial and temporal contexts associated with urban forest creation and management. The land types found in suburban areas represent divergences in tree establishment, planting, and maintenance practices, and thus reflect various manifestations in tree-species composition. Urban forest policies and management traditions likely explain differences observed in street-tree species composition between older and newer neighbourhoods. The pre-urbanized landscape, on the other hand, appears to be an important driver of urban forest composition on residential properties, as native species can naturally establish onto adjacent green spaces from forest buffers and patches, and potentially alter tree diversity over time.
Studies like these can encourage urban forest practitioners to reflect on the ways in which cities are developed, and how green infrastructure is created and molded based on the preferences and priorities of multiple professionals and urban dwellers. Envisioning how urban areas can be built to enhance urban forest diversity also allows for the opportunity to contemplate the kind of diversity that we would like to see in our cities. Although tree communities in London were generally more diverse, Halifax had greater proportions of native trees on residential properties. Is one case better than the other? What elements of urban forest diversity should be prioritized when developing and implementing management plans? These are crucial considerations, especially given growing concerns about biodiversity loss and environmental disturbances like pests, diseases, and climate change, which threaten the success of trees in urban landscapes. Given these difficulties, managers and practitioners should work towards achieving the right kind of biodiversity, instead of simply aiming for more biodiversity.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}