
Pattern and Drivers of White-Tailed Deer (Odocoileus virginianus) Herbivory on Tree Saplings across a Plateau Landscape

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Methods
2.3. Sapling Density Patterns
2.4. A Model of Sapling Density Based on Landscape Predictors
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ΔAICc | Difference in Akaike’s information criteria with small sample size correction |
| AIC | Akaike’s information criteria |
| AICc | Akaike’s information criteria with small sample size correction |
| CI | confidence interval |
| DEM | digital elevation model |
| MSE | Mean square error |
| NAPP | National Aerial Photography Program |
| RMSE | Root mean square error |
| SE | Standard error |
| UoS | University of the South |
Appendix

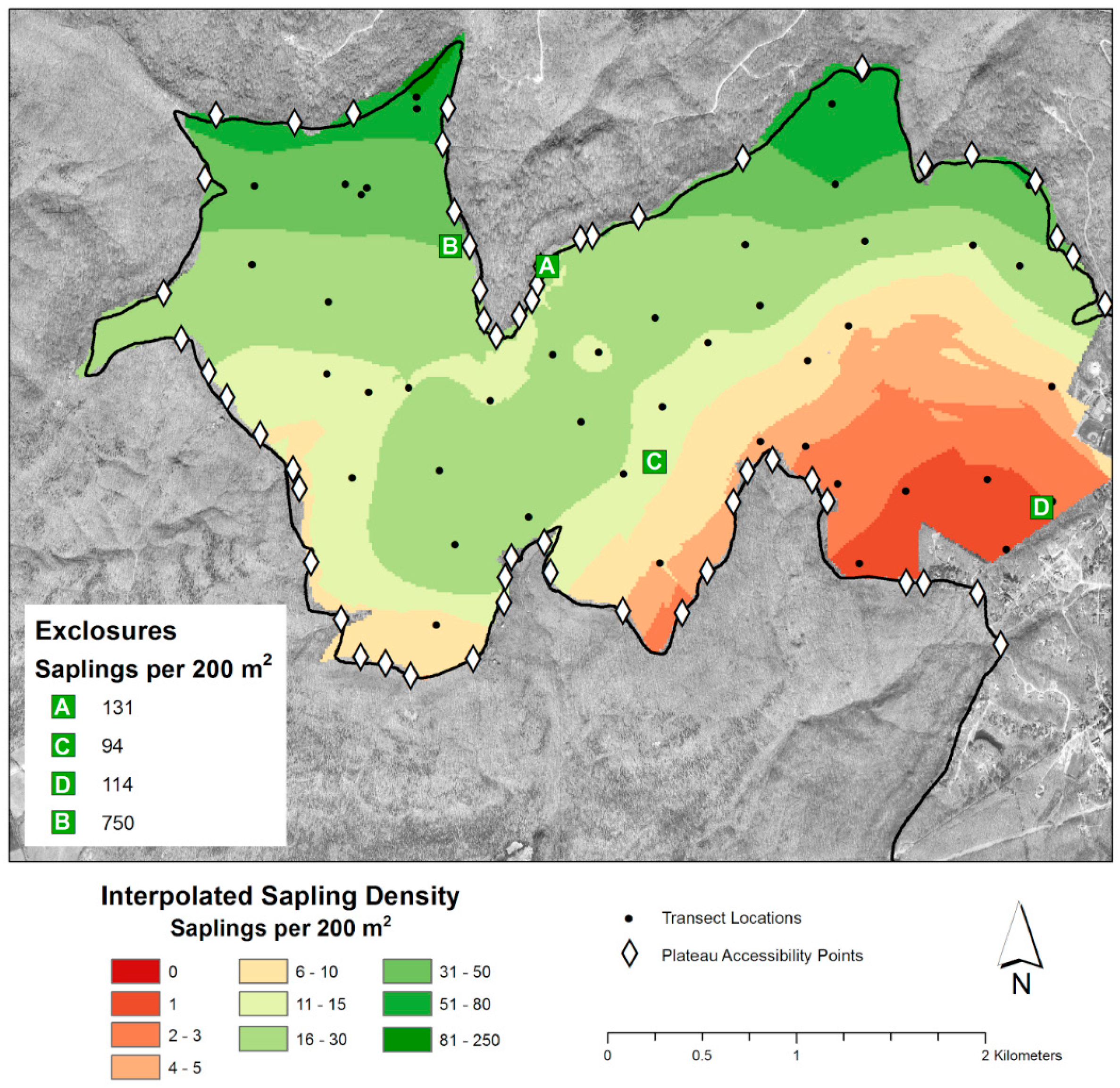
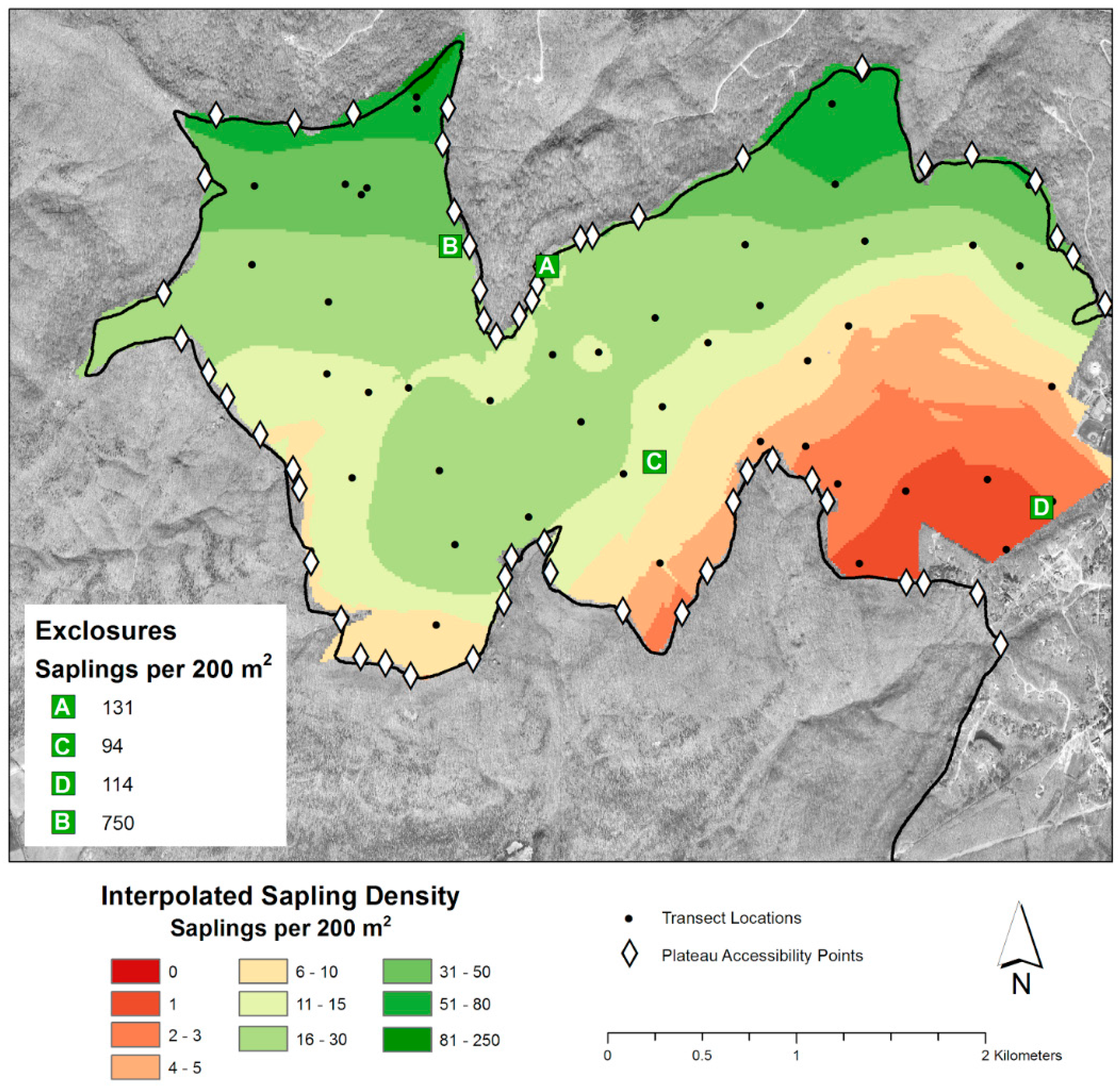{kind=link}
{kind=link}
| Edge | Agriculture | Clearcut | Road | Residential | Cleared Road | Lake | Latitude | Plateau Access Zone | Number Plateau Access Zones | Width Plateau Access Zones | 2014 Deer Cull | 3 Year Deer Cull | 5 Year Deer Cull | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Edge | NA | |||||||||||||
| Agriculture | 0.12 (0.28) | NA | ||||||||||||
| Clearcut | 0.23 (0.03) | −0.03 (0.75) | NA | |||||||||||
| Road | 0.08 (0.45) | 0.98 (0) | −0.16 (0.14) | NA | ||||||||||
| Residential | 0.07 (0.51) | 0.97 (0) | −0.18 (0.08) | 0.99 (0) | NA | |||||||||
| Cleared Road | 0.56 (<0.001) | −0.19 (0.07) | 0.25 (0.02) | −0.22 (0.04) | −0.22 (0.04) | NA | ||||||||
| Lake | 0.06 (0.59) | 0.16 (0.14) | −0.05 (0.66) | 0.13 (0.22) | 0.12 (0.25) | −0.23 (0.03) | NA | |||||||
| Latitude | −0.08 (0.46) | 0.25 (0.02) | −0.56 (<0.001) | 0.41 (<0.001) | 0.45 (<0.001) | −0.21 (0.04) | 0.09 (0.40) | NA | ||||||
| Plateau Access Zone | −0.12 (0.27) | −0.15 (0.16) | −0.25 (0.02) | −0.12 (0.24) | −0.09 (0.41) | 0.13 (0.24) | −0.44 (<0.001) | −0.05 (0.66) | NA | |||||
| Number Plateau Access Zones | −0.11 (0.30) | −0.23 (0.03) | 0.24 (0.02) | −0.28 (0.01) | −0.32 (0.002) | 0.05 (0.62) | −0.17 (0.10) | −0.53 (<0.001) | 0.14 (0.20) | NA | ||||
| Width Plateau Access Zones | 0.15 (0.17) | 0.34 (0.001) | −0.05 (0.66) | 0.37 (<0.001) | 0.38 (<0.001) | 0.05 (0.68) | −0.04 (0.72) | 0.35 (<0.001) | −0.13 (0.22) | −0.39 (<0.001) | NA | |||
| 2014 Deer Cull | −0.05 (0.65) | −0.60 (<0.001) | −0.15 (0.16) | −0.54 (<0.001) | −0.52 (<0.001)) | 0.31 (0.003) | −0.18 (0.08) | −0.02 (0.82) | 0.28 (0.01) | 0.01 (0.94) | −0.13 (0.23) | NA | ||
| 3 Year Deer Cull | −0.07 (0.53) | −0.58 (<0.001) | −0.17 (0.10) | −0.54 (<0.001) | −0.51 (<0.001) | 0.18 (0.09) | −0.13 (0.23) | −0.04 (0.70) | 0.35 (<0.001) | −0.03 (0.77) | −0.15 (0.15) | 0.93 (0) | NA | |
| 5 Year Deer Cull | −0.08 (0.48) | −0.56 (<0.001) | −0.11 (0.33) | −0.54 (<0.001) | −0.51 (<0.001) | 0.20 (0.06) | −0.16 (0.13) | −0.11 (0.30) | 0.35 (<0.001) | −0.02 (0.87) | −0.14 (0.19) | 0.84 (0) | 0.95 (0) | NA |
References
- Stromayer, K.A.; Warren, R.J. Are overabundant deer herds in the Eastern United States creating alternate stable states in forest plant communities? Wildl. Soc. B 1997, 25, 227–234. [Google Scholar]
- Russell, F.L.; Zippin, D.B.; Fowler, N.L. Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: A review. Am. Midl. Nat. 2001, 146, 1–26. [Google Scholar]
- Côté, S.D.; Rooney, T.P.; Tremblay, J.P.; Dussault, C.; Waller, D.M. Ecological impacts of deer overabundance. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef]
- Latham, R.E.; Beyea, J.; Benner, M.; Dunn, C.A.; Fajvan, M.A.; Freed, R.R.; Grund, M.; Horsley, S.B.; Rhoads, A.F.; Shissler, B.P. Managing white-tailed deer in forest habitat from an ecosystem perspective. In Report by the Deer Management Forum for Audubon Pennsylvania and Pennsylvania Habitat Alliance; Audubon Pennsylvania: Harrisburg, PA, USA, 2005; p. 340. [Google Scholar]
- White, M.A. Long-term effects of deer browsing: Composition, structure and productivity in a northeastern Minnesota old-growth forest. For. Ecol. Manag. 2012, 269, 222–228. [Google Scholar] [CrossRef]
- Bugalho, M.N.; Ibáñez, I.; Clark, J.S. The effects of deer herbivory and forest type on tree recruitment vary with plant growth stage. For. Ecol. Manag. 2013, 308, 90–100. [Google Scholar] [CrossRef]
- Kobe, R.K. Intraspecific variation in sapling mortality and growth predicts geographic variation in forest composition. Ecol. Monogr. 1996, 66, 181–201. [Google Scholar] [CrossRef]
- Wyckoff, P.H.; Clark, J.S. The relationship between growth and mortality for seven co-occurring tree species in the southern Appalachian Mountains. J. Ecol. 2002, 90, 604–615. [Google Scholar] [CrossRef]
- Fisichelli, N.; Frelich, L.E.; Reich, P.B. Sapling growth responses to warmer temperatures ‘cooled’ by browse pressure. Glob. Chang. Biol. 2012, 18, 3455–3463. [Google Scholar] [CrossRef]
- Alverson, W.S.; Waller, D.M.; Solheim, S.L. Forests too deer: Edge effects in northern Wisconsin. Conserv. Biol. 1988, 2, 348–358. [Google Scholar] [CrossRef]
- Healy, W.M. Influence of deer on the structure and composition of oak forests in central Massachusetts. In The Science of Overabundance: Deer Ecology and Population Management; McShea, W.J., Underwood, H.B., Rappole, J.H., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1997; pp. 246–266. [Google Scholar]
- Anderson, R.C.; Loucks, O.L. White-tail deer (Odocoileus virginianus) influence on structure and composition of Tsuga canadensis forests. J. Appl. Ecol. 1979, 16, 855–861. [Google Scholar] [CrossRef]
- Rossell, C.R.; Gorsira, B.; Patch, S. Effects of white-tailed deer on vegetation structure and woody seedling composition in three forest types on the Piedmont Plateau. For. Ecol. Manag. 2005, 210, 415–424. [Google Scholar] [CrossRef]
- Rooney, T.P.; McCormick, R.J.; Solheim, S.L.; Waller, D.M. Regional variation in recruitment of hemlock seedlings and saplings in the upper Great Lakes, USA. Ecol. Appl. 2000, 10, 1119–1132. [Google Scholar] [CrossRef]
- Horsley, S.B.; Stout, S.L.; de Calesta, D.S. White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol. Appl. 2003, 13, 98–118. [Google Scholar] [CrossRef]
- Scribner, K.T.; Smith, M.H.; Chesser, R.K. Spatial and temporal variability of microgeographic genetic structure in white-tailed deer. J. Mammal. 1997, 78, 744–755. [Google Scholar] [CrossRef]
- Van Deelen, T.R.; Campa, H., III; Hamady, M.; Haufler, J.B. Migration and seasonal range dynamics of deer using adjacent deeryards in northern Michigan. J. Wildl. Manag. 1998, 62, 205–213. [Google Scholar] [CrossRef]
- Finder, R.A.; Roseberry, J.L.; Woolf, A. Site and landscape conditions at white-tailed deer/vehicle collision locations in Illinois. Landsc. Urban Plan. 1999, 44, 77–85. [Google Scholar] [CrossRef]
- Campbell, T.A.; Laseter, B.R.; Ford, W.M.; Miller, K.V. Topographic home ranges of white-tailed deer in the central Appalachians. Southeast. Nat. 2004, 3, 645–652. [Google Scholar] [CrossRef]
- Boulanger, V.; Baltzinger, C.; Saïd, S.; Ballon, P.; Picard, J.F.; Dupouey, J.L. Ranking temperate woody species along a gradient of browsing by deer. For. Ecol. Manag. 2009, 258, 1397–1406. [Google Scholar] [CrossRef]
- Walter, W.D.; VerCauteren, K.C.; Campa, H., III; Clark, W.R.; Fischer, J.W.; Hygnstrom, S.E.; Mathews, N.E.; Nielsen, C.K.; Schauber, E.M.; van Deelen, T.R.; et al. Regional assessment on influence of landscape configuration and connectivity on range size of white-tailed deer. Landsc. Ecol. 2009, 24, 1405–1420. [Google Scholar] [CrossRef]
- Quinn, A.C.; Williams, D.M.; Porter, W.F. Landscape structure influences space use by white-tailed deer. J. Mammal. 2013, 94, 398–407. [Google Scholar] [CrossRef]
- Waller, D.M.; Alverson, W.S. The white-tailed deer: A keystone herbivore. Wildl. Soc. B 1997, 25, 217–226. [Google Scholar]
- Williamson, S.J.; Hirth, D.H. An evaluation of edge use by white-tailed deer. Wildl. Soc. B 1985, 13, 252–257. [Google Scholar]
- Ries, L.; Fletcher, R.J., Jr.; Battin, J.; Sisk, T.D. Ecological responses to habitat edges: Mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 491–522. [Google Scholar] [CrossRef]
- Wirth, R.; Meyer, S.T.; Leal, I.R.; Tabarelli, M. Plant herbivore interactions at the forest edge. In Progress in Botany; Springer-Verlag: Berlin/Heidelberg, Germany, 2008; pp. 423–448. [Google Scholar]
- Ruzicka, K.J.; Groninger, J.W.; Zaczek, J.J. Deer browsing, forest edge effects, and vegetation dynamics following bottomland forest restoration. Restoration Ecol. 2010, 18, 702–710. [Google Scholar] [CrossRef]
- Loft, E.R.; Menke, J.W.; Burton, T.S. Seasonal movements and summer habitats of female black-tailed deer. J. Wildl. Manag. 1984, 48, 1317–1325. [Google Scholar] [CrossRef]
- Romin, L.A.; Bissonette, J.A. Temporal and spatial distribution of highway mortality of mule deer on newly constructed roads at Jordanelle Reservoir, Utah. Great Basin Nat. 1996, 56, 1–11. [Google Scholar]
- Pauley, G.R.; Peek, J.M.; Zager, P. Predicting white-tailed deer habitat use in Northern Idaho. J. Wildl. Manag. 1993, 57, 904–913. [Google Scholar] [CrossRef]
- Stroh, E.D.; Struckhoff, M.A.; Grabner, K.W. A Floral Survey of Cliff Habitats along Bull Run at Manassas National Battlefield Park, Virginia; 2014 (No. 940); US Geological Survey: Columbia, MO, USA, 2015.
- Kie, J.G.; Bowyer, R.T.; Nicholson, M.C.; Boroski, B.B.; Loft, E.R. Landscape heterogeneity at differing scales: Effects on spatial distribution of mule deer. Ecology 2002, 83, 530–544. [Google Scholar] [CrossRef]
- Knoll, M.A.; Potter, D.B.; van de Ven, C. Geology, hydrology, and water use history atop the Cumberland Plateau in the Sewanee and Tracy City, Tennessee, area. Field Guides 2015, 39, 197–218. [Google Scholar]
- Cheston, C. The University of the South Forest: A demonstration of practical applied forestry for student instruction and regional use. In Forestry Department; The University of the South: Sewanee, TN, USA, 1953. [Google Scholar]
- Reid, J.L.; Evans, J.P.; Hiers, J.K.; Harris, J.B.C. Ten years of forest change in two adjacent communities on the southern Cumberland Plateau, USA. J. Torrey Bot. Soc. 2008, 135, 224–235. [Google Scholar] [CrossRef]
- R Core Team. The R Project for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 1 January 2016).
- ESRI. 10.1; Environmental Systems Research Institute: Redlands, CA, USA, 2012.
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: An Information-Theoretic Approach; Springer-Verlag: New York, NY, USA, 1998. [Google Scholar]
- Sheskin, D.J. Handbook of Parametric and Nonparametric Statistical Procedures, 4th ed.; Chapman & Hall/CRC: Boca Raton, FL, USA, 2003; pp. 981–993. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-PLUS, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Akaike, H. Information measures and model selection. B Int. Statist. Inst. 1983, 50, 277–291. [Google Scholar]
- Hurvich, C.M.; Tsai, C. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Royall, R.M. Statistical Evidence: A Likelihood Paradigm; Chapman and Hall: New York, NY, USA, 1997. [Google Scholar]
- Bradley, K.L.; Damschen, E.I.; Young, L.M.; Kuefler, D.; Went, S.; Wray, G.; Haddad, N.M.; Knops, J.M.H.; Louda, S.M. Spatial heterogeneity, not visitation bias, dominates variation in herbivory. Ecology 2002, 84, 2214–2221. [Google Scholar] [CrossRef]
- Royo, A.A.; Stout, S.L.; Pierson, T.G. Restoring forest herb communities through landscape-level deer herd reductions: Is recovery limited by legacy effects? Biol. Conserv. 2010, 143, 2425–2434. [Google Scholar] [CrossRef]
- Tanentzap, A.J.; Bazely, D.R.; Koh, S.; Timciska, M.; Haggith, E.G.; Carleton, T.J.; Coomes, D.A. Seeing the forest for the deer: Do reductions in deer-disturbance lead to forest recovery? Biol. Conserv. 2011, 144, 376–382. [Google Scholar] [CrossRef]
- Long, Z.T.; Pendergast, T.H., IV; Carson, W.P. The impact of deer on relationships between tree growth and mortality in an old-growth beech-maple forest. For. Ecol. Manag. 2007, 252, 230–238. [Google Scholar] [CrossRef]
- Anderson, M.G.; Barnett, A.; Clark, M.; Ferree, C.; Sheldon, A.O.; Prince, J. Resilient Sites for Terrestrial Conservation in the Southeast Region; The Nature Conservancy, Eastern Conservation Science: Boston, MA, USA, 2014; p. 127. [Google Scholar]
- Saïd, S.; Servanty, S. The influence of landscape structure on female roe deer home-range size. Landsc. Ecol. 2005, 20, 1003–1012. [Google Scholar] [CrossRef]
- Millington, J.D.; Walters, M.B.; Matonis, M.S.; Liu, J. Effects of local and regional landscape characteristics on wildlife distribution across managed forests. For. Ecol. Manag. 2010, 259, 1102–1111. [Google Scholar] [CrossRef]
- Massé, A.; Côté, S.D. Linking habitat heterogeneity to space use by large herbivores at multiple scales: From habitat mosaics to forest canopy openings. For. Ecol. Manag. 2012, 285, 67–76. [Google Scholar] [CrossRef]
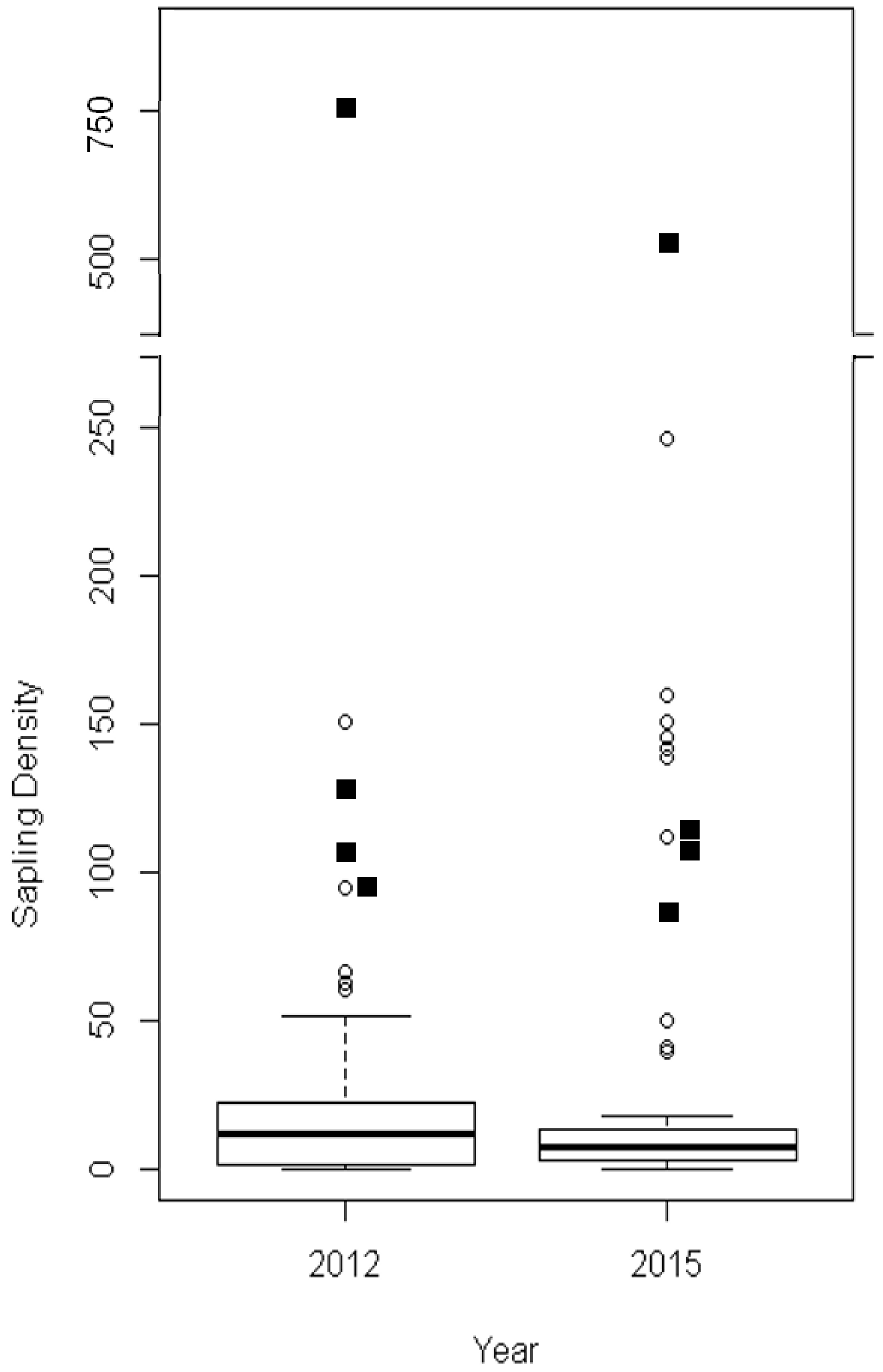
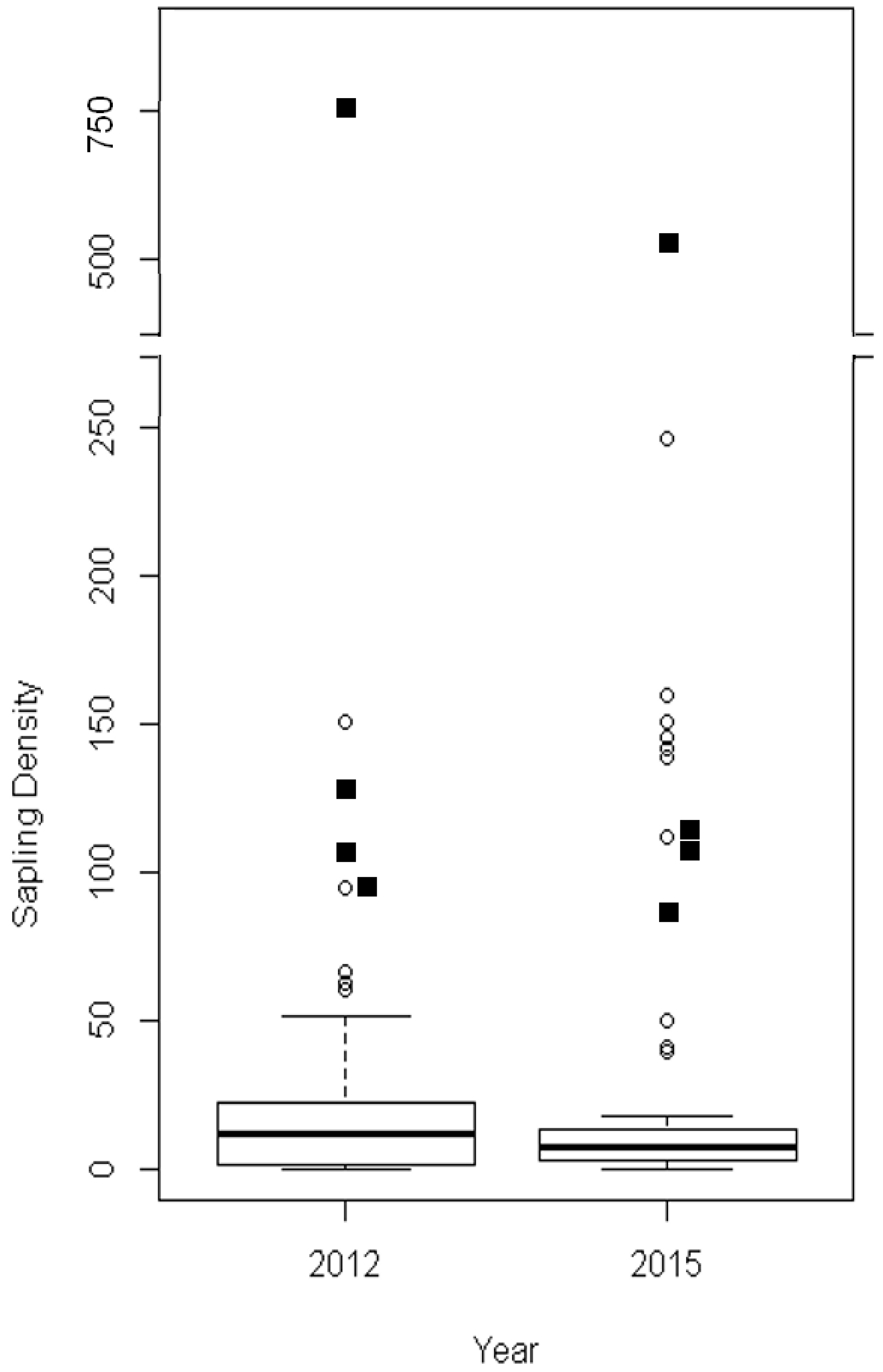
| Model Variable | Explanation |
|---|---|
| Edge Predictors | |
| Residential Development | Distance to residential development (m) |
| Road | Distance to road (m) |
| Cleared Road | Distance to cleared road (m) |
| Recent Clearcut | Distance to clearcut (m) present 2012–2015 |
| Livestock Pasture/Agriculture | Distance to agriculture (m) |
| Lake | Distance to lake (m) |
| Nearest Edge | Distance to nearest edge (m), defined as the nearest of all edge predictors |
| Plateau Accessibility Predictors | |
| Latitude | Latitude (decimal degrees) |
| Plateau Access Zone | Distance to closest plateau access zone (m) |
| Number of Nearby Plateau Access Zones | Number of plateau access zones within 500 m of closest bluff edge |
| Average Nearby Plateau Access Zone Width | Average width of all plateau access zones within 500 m of closest bluff edge |
| Deer Cull Predictors | |
| 5 Year Deer Cull | 5 year average number of deer killed (per km2) in associated cull area |
| 3 Year Deer Cull | 3 year average number of deer killed (per km2) in associated cull area |
| 2014 Deer Cull | Average number of deer killed (per km2) in associated cull area in 2014 |
| Model | Parameter | AICc | ΔAICc | MSE | RMSE |
|---|---|---|---|---|---|
| Sapling Density | |||||
| Latitude | 2 | 687.8 | 0 | 1.671 | 1.293 |
| Residential | 2 | 716.7 | 28.9 | 2.305 | 1.519 |
| 5 Year Deer Cull | 2 | 730.2 | 42.4 | 1.854 | 1.361 |
| Year | 2 | 750.3 | 62.5 | 2.474 | 1.572 |
| Parameter | Estimate (SE) | Lower 95% CI | Upper 95% CI | Unit Change | Scaled Estimate | Importance Weight |
|---|---|---|---|---|---|---|
| Sapling Density | ||||||
| Latitude | 142 (16.82) | 113.31 | 171.33 | 0.01 degrees | 1.42 | 0.99 |
| Residential | 9.22 × 10−4 (1.27 × 10−4) | 6.38 × 10−4 | 1.22 × 10−3 | 1000 m | 1.226 | <0.01 |
| 5 Year Deer Cull | −6.44 (1.10) | −8.58 | −4.12 | 0.1 deer/km2 | −0.64 | <0.01 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, J.P.; Oldfield, C.A.; Cecala, K.K.; Hiers, J.K.; Van De Ven, C.; Armistead, M.M. Pattern and Drivers of White-Tailed Deer (Odocoileus virginianus) Herbivory on Tree Saplings across a Plateau Landscape. Forests 2016, 7, 101. https://doi.org/10.3390/f7050101
Evans JP, Oldfield CA, Cecala KK, Hiers JK, Van De Ven C, Armistead MM. Pattern and Drivers of White-Tailed Deer (Odocoileus virginianus) Herbivory on Tree Saplings across a Plateau Landscape. Forests. 2016; 7(5):101. https://doi.org/10.3390/f7050101
Chicago/Turabian StyleEvans, Jonathan P., Callie A. Oldfield, Kristen K. Cecala, John Kevin Hiers, Chris Van De Ven, and Meg M. Armistead. 2016. "Pattern and Drivers of White-Tailed Deer (Odocoileus virginianus) Herbivory on Tree Saplings across a Plateau Landscape" Forests 7, no. 5: 101. https://doi.org/10.3390/f7050101
APA StyleEvans, J. P., Oldfield, C. A., Cecala, K. K., Hiers, J. K., Van De Ven, C., & Armistead, M. M. (2016). Pattern and Drivers of White-Tailed Deer (Odocoileus virginianus) Herbivory on Tree Saplings across a Plateau Landscape. Forests, 7(5), 101. https://doi.org/10.3390/f7050101