
How natural Forest Conversion Affects Insect Biodiversity in the Peruvian Amazon: Can Agroforestry Help?


Abstract
:1. Introduction
2. Experimental Section
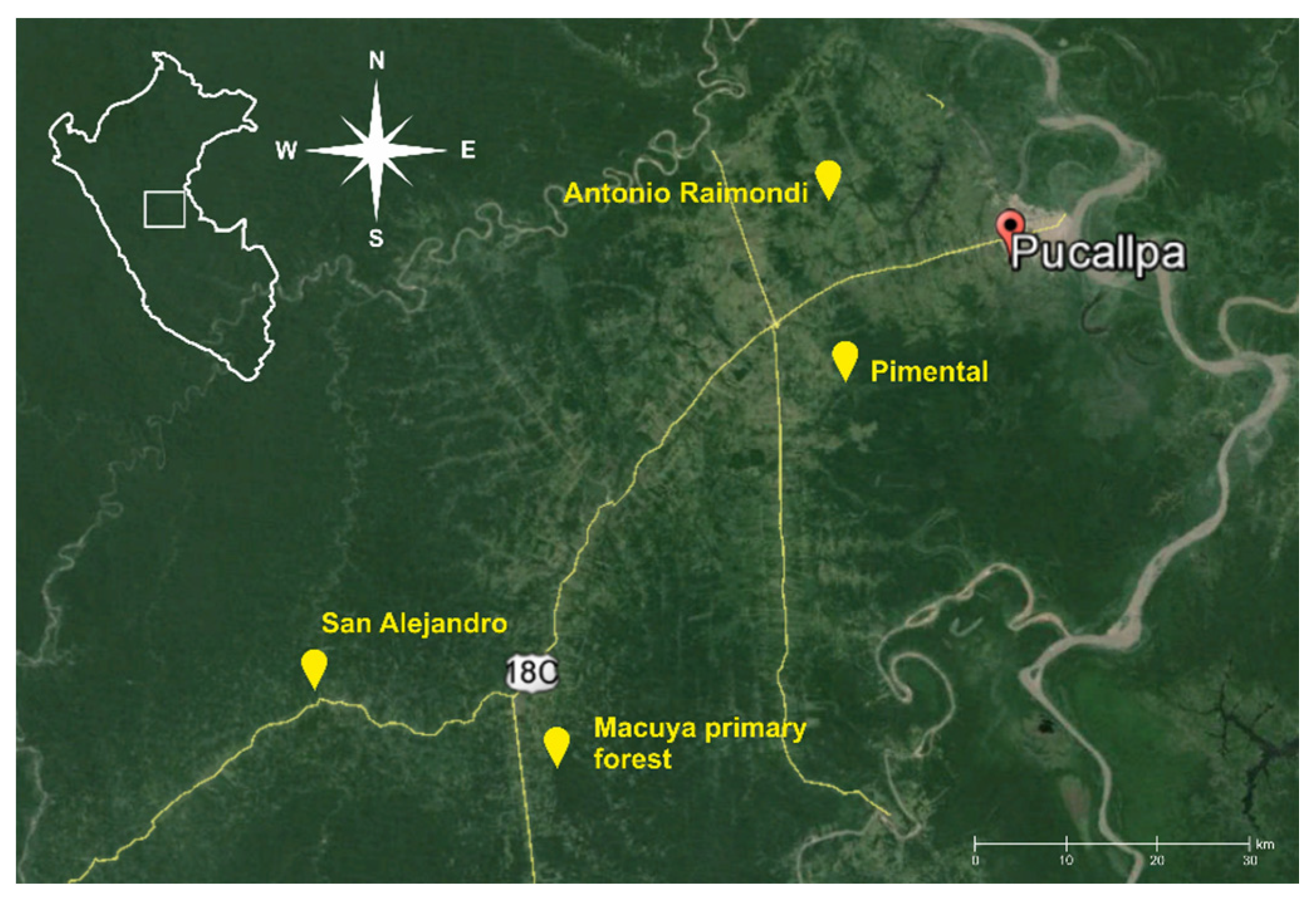
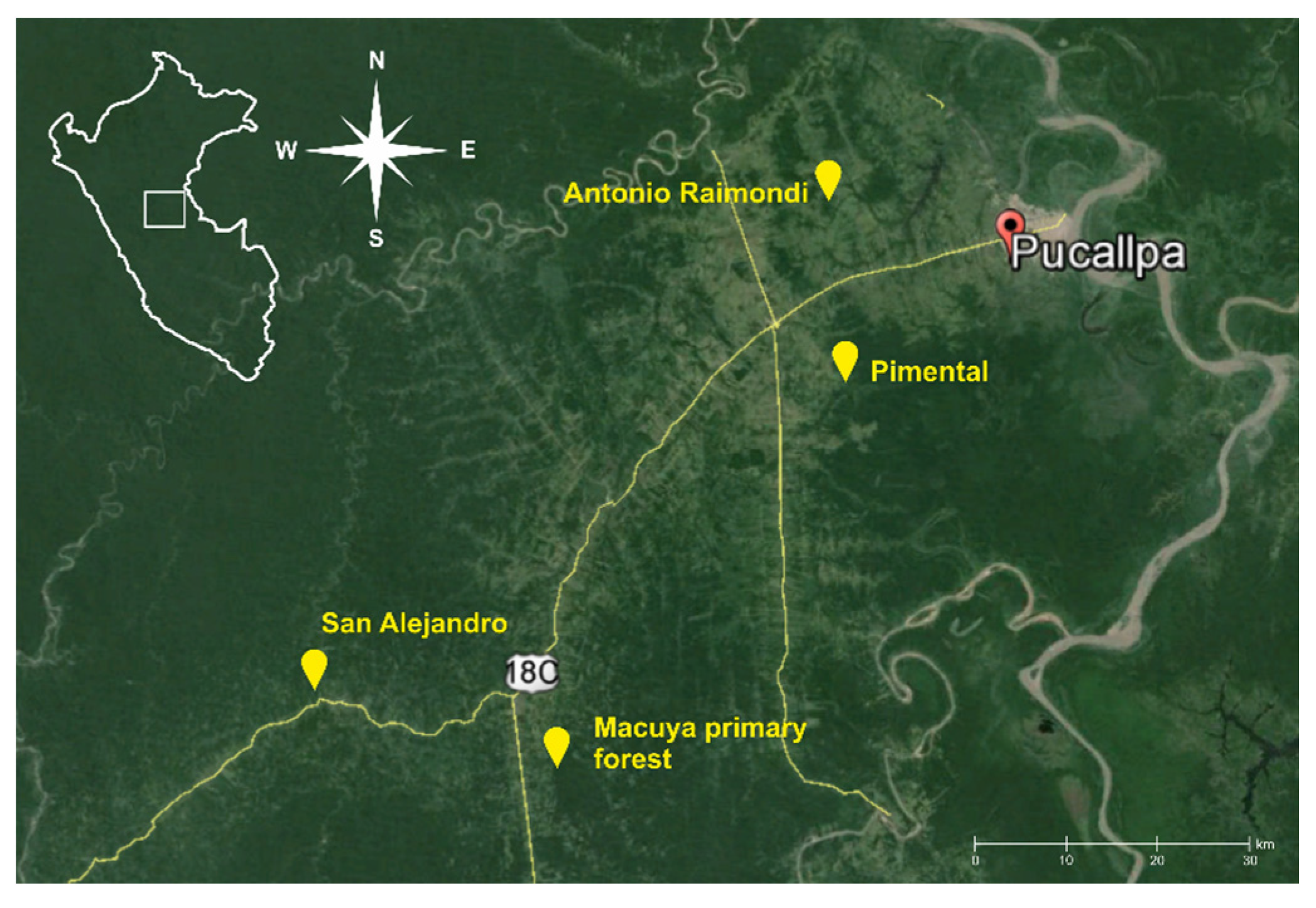
2.1. Study Sites
2.2. Data Collection
2.3. Data Evaluation
3. Results
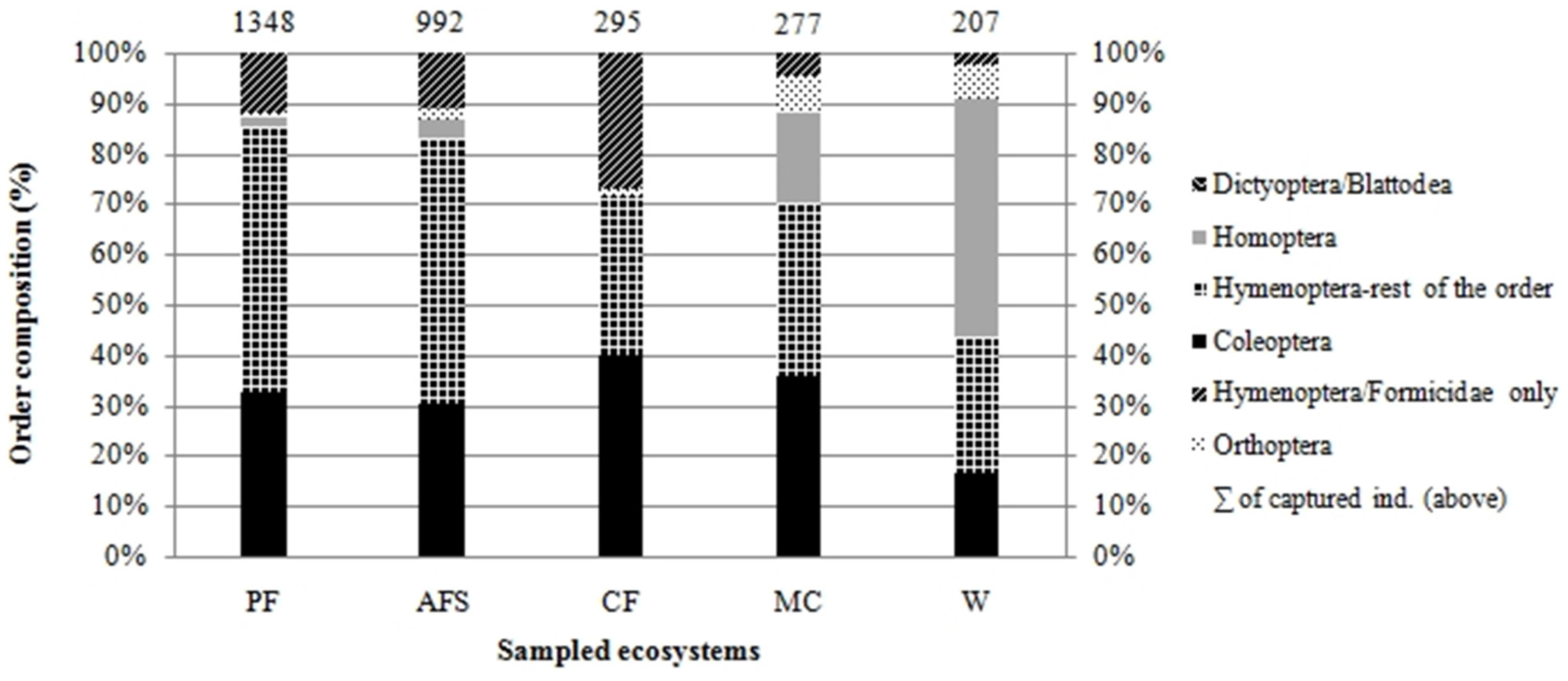
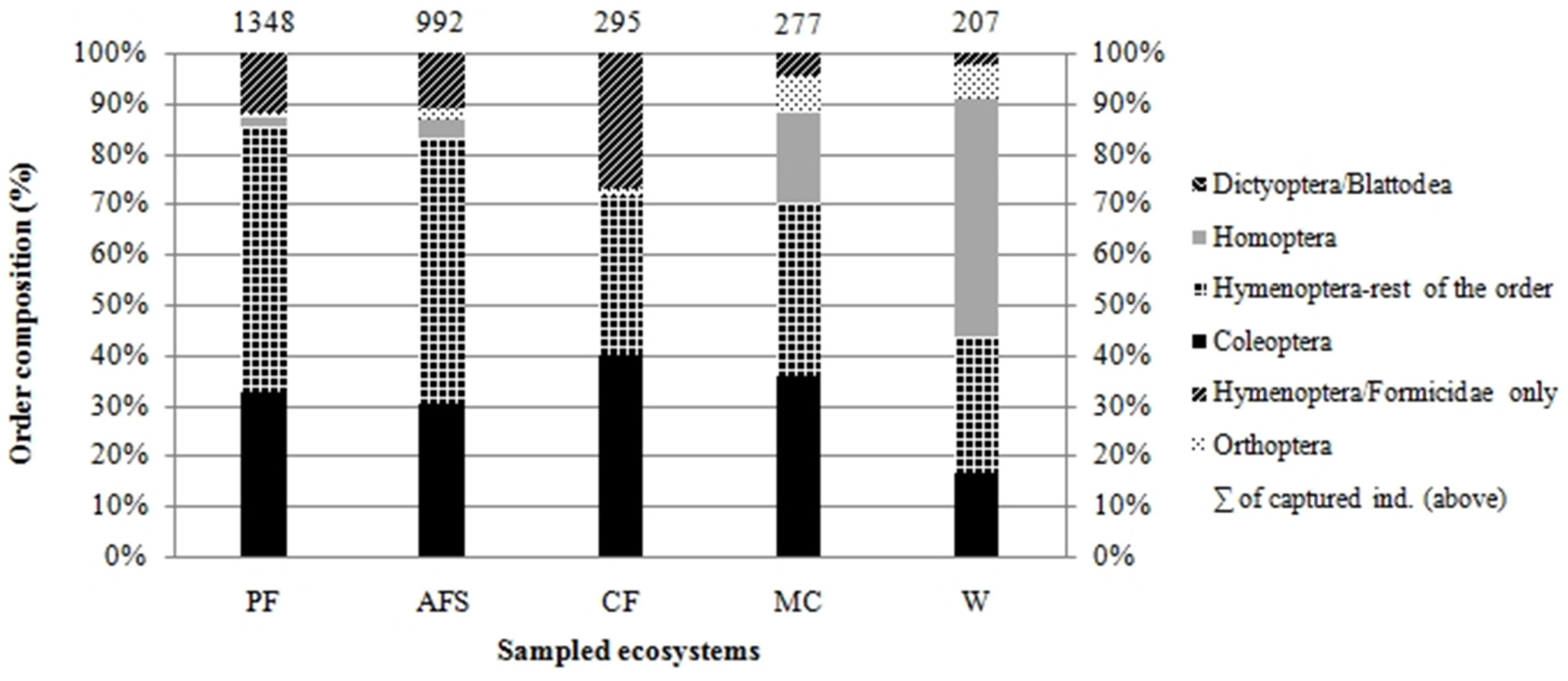
3.1. Species Abundance and Composition
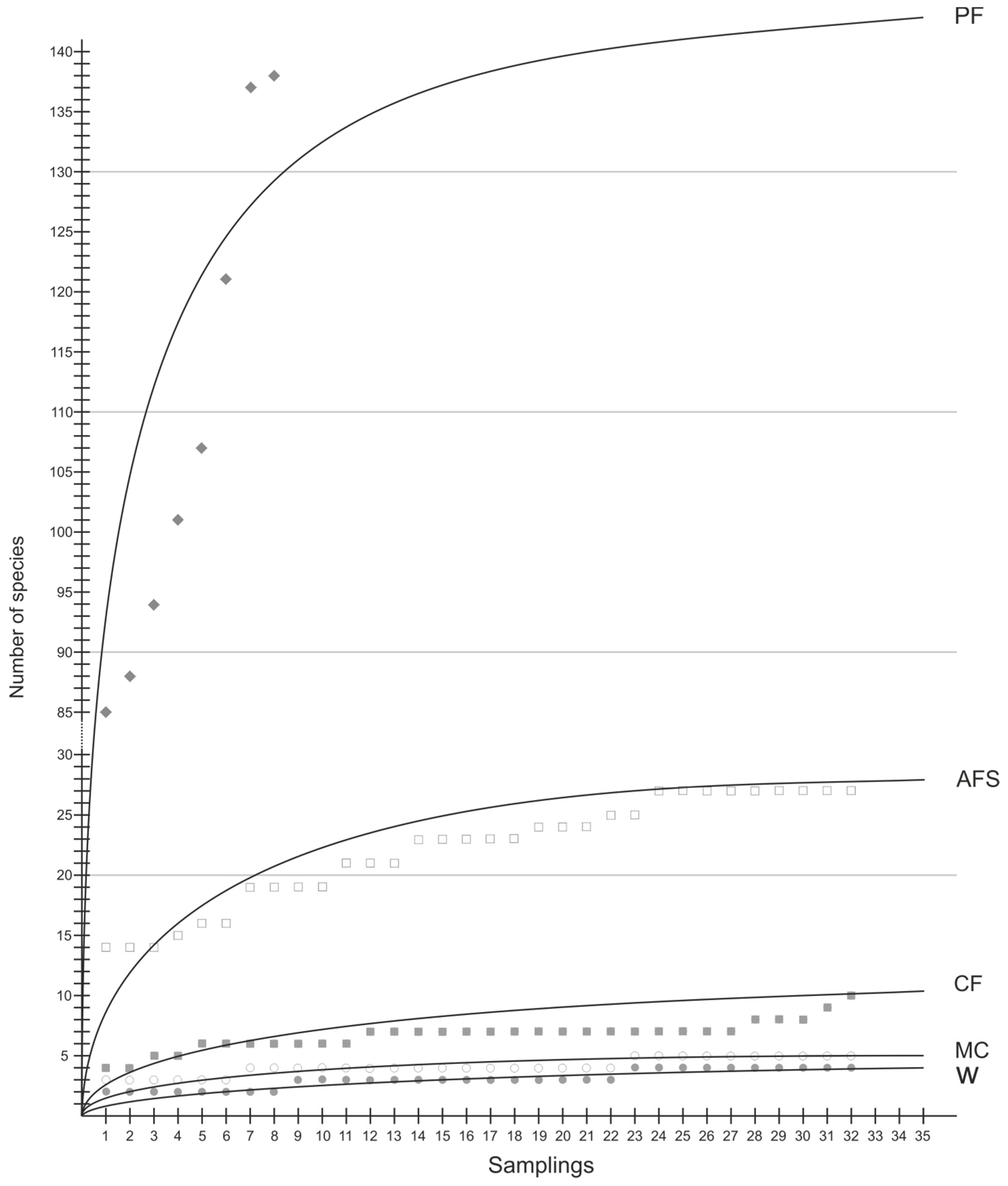
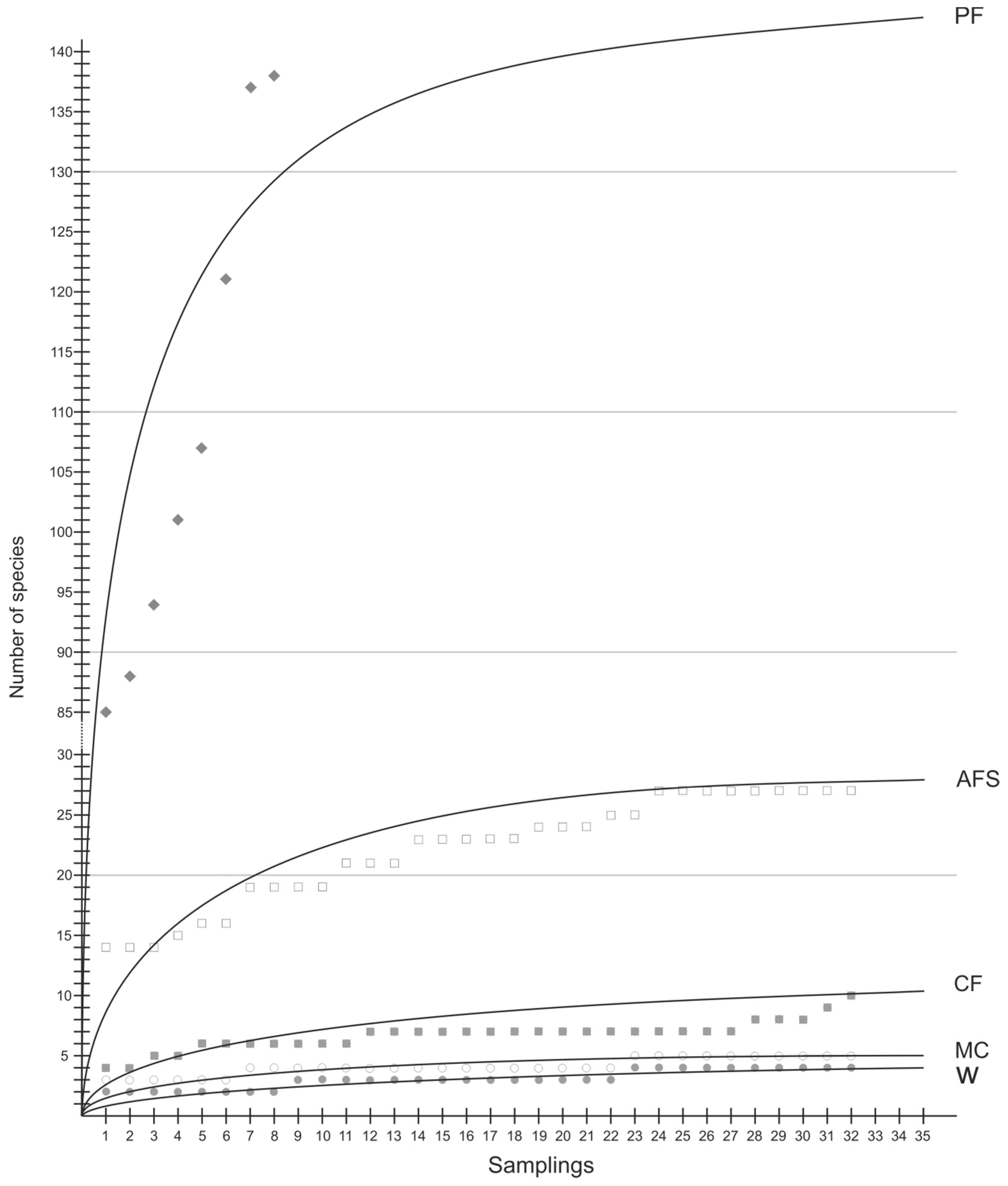
3.2. Species Richness and Diversity
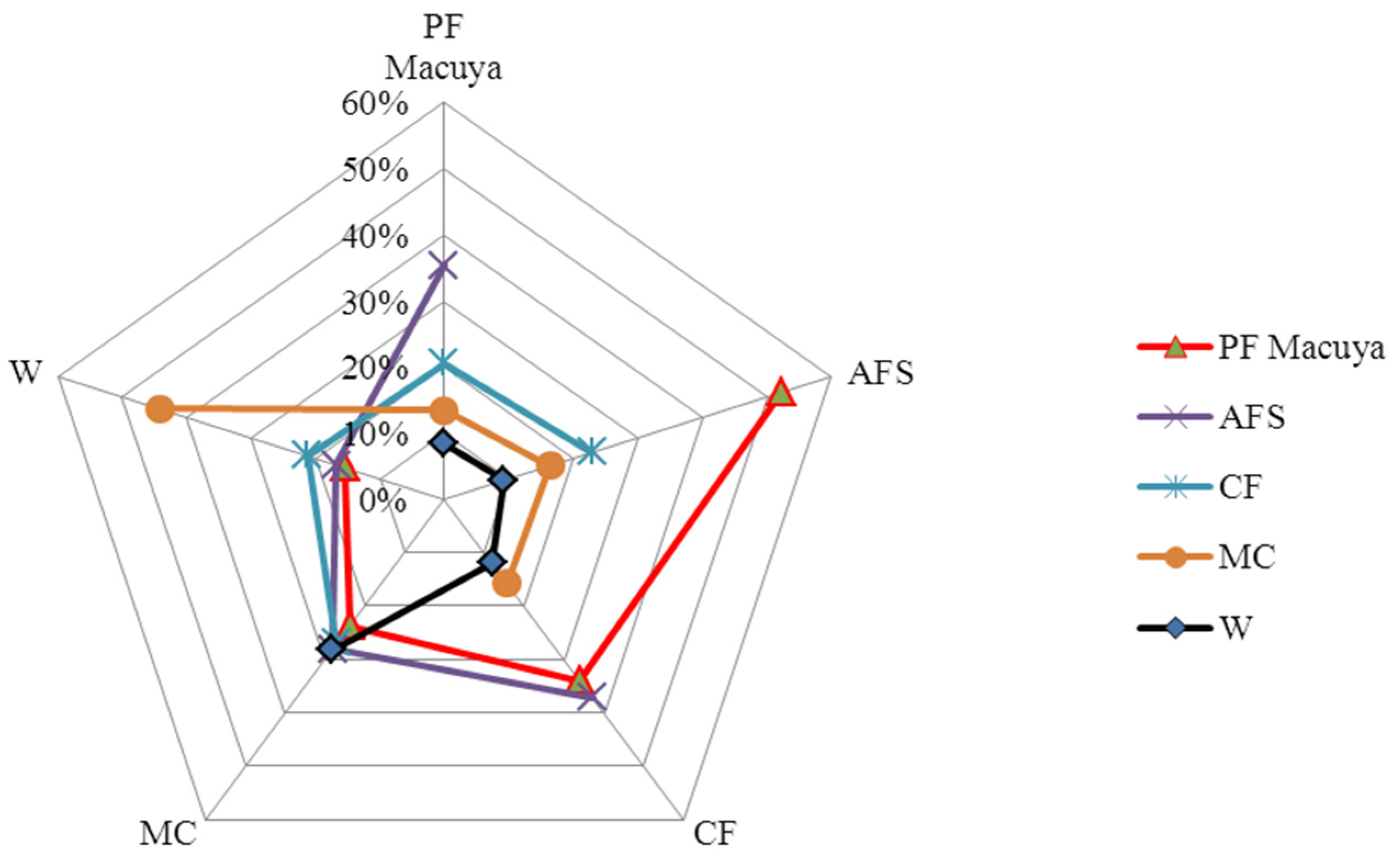
3.3. Similarity among the Habitats
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonesca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Millenium Ecosystem Assessment. Ecosystems and Human Well-Being: Biodiversity Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- Clay, J.W.; Clement, C.R. Selected Species and Strategies to Enhance Income Generation from Amazonian Forests. FAO: Misc/93/6 Working Paper: Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/3/a-v0784e.pdf (accessed on 28 August 2015).
- Oliviera, P.J.C.; Asner, G.P.; Knapp, D.E.; Almeyda, A.; Galván-Gildemeister, R.; Keene, S.; Raybin, R.F.; Smith, R.C. Land use Allocation Protects the Peruvian Amazon. Science 2007, 317, 1233–1237. [Google Scholar] [CrossRef] [PubMed]
- Hyman, G.; Fujisaka, S. Final Report Desakota, Part II G4. Case Study: Central Peruvian Amazon—A test Case for Desakota Development in the Amazon. International Centre for Tropical Agriculture (CIAT); p. 9. Available online: http://www.dfid.gov.uk/r4d/Output/179310/Default.aspx (accessed on 13 September 2008).
- Hyman, G.; Puig, J.; Bolanos, S. Multi-source remote sensing and GIS for exploring deforestation patterns and processes in the Central Peruvian Amazon. In Proceedings of the 29th International Symposium on Remote Sensing of Environment, Buenos Aires, Argentina, 8–12 April 2002.
- Swallow, B.; van Noordwijk, M.; Dewi, S.; Murdiyarso, D.; White, D.; Gockowski, J.; Hyman, G.; Budidarsono, S.; Robiglio, V.; Meadu, V.; et al. Opportunities for Avoided Deforestation with Sustainable Benefits. Available online: http://www.asb.cgiar.org/PDFwebdocs/Report-on-Opportunitiesfor-Avoided-Deforestation-Sustainable-Benefits-web-low.pdf (accessed on 6 October 2007).
- Takasaki, Y. Economic Model of Shifting Cultivation: A Review; Tsukuba Economics Working Papers No. 2011-006; University of Tsukuba: Tsukuba, Japan; p. 18.
- Fujisaka, S.; White, D. Pasture or permanent crops after slash-and-burn cultivation? Land-use choice in three Amazon colonies. Agrofor. Syst. 1998, 42, 45–59. [Google Scholar] [CrossRef]
- Kleinman, P.J.A.; Pimentel, D.; Bryant, R.B. The ecological sustainability of slash-and-burn agriculture. Agric. Ecosyst. Environ. 1995, 52, 235–249. [Google Scholar] [CrossRef]
- De Jong, W.; Freitas, L.; Baluarte, J.; van de Kop, P.; Salazar, A.; Inga, E.; Melendez, W.; Germaná, C. Secondary forest dynamics in the Amazon floodplain in Peru. For. Ecol. Manag. 2001, 150, 135–146. [Google Scholar] [CrossRef]
- Puri, S.; Panwar, P. Agroforestry: System and Practices; New India Publishing Agency: New Delhi, India, 2007; p. 657. [Google Scholar]
- Brown, K.S., Jr.; Hutchings, R.W. Chapter 7. Disturbance, Fragmentation, and the Dynamics of Diversity in Amazonian Forest Butterflies. In Tropical Forest Remnants—Ecology, Management and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R.O., Jr., Eds.; The University of Chicago Press: Chicago, UK, 1997; p. 616. [Google Scholar]
- Godfray, H.C.J.; Lewis, O.T.; Memmott, J. Studying insect diversity in the tropics. Philos. Trans. R. Soc. Lond. 1999, 354, 1811–1824. [Google Scholar] [CrossRef] [PubMed]
- Stamps, W.; Linit, M. Plant diversity and arthropod communities: Implications for temperate agroforestry. Agrofor. Syst. 1997, 39, 73–89. [Google Scholar] [CrossRef]
- Andow, D.A. Vegetational diversity and arthropod population response. Annu. Rev. Entomol. 1991, 36, 561–586. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology, 2nd ed.; Addison-Wesley Educational Publishers, Inc.: Boston, MA, USA, 1999. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2001; p. 260. [Google Scholar]
- Khan, A. Methodology for Assessing Biodiversity. Centre of Advanced Study in Marine Biology, Annamalai University. Available online: http://www.slideshare.net/MMASSY/methodology-forassessmentbiodiversity (accessed on 28 August 2015).
- Sörensen, T.A. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content, and its aplication to analyses of the vegetation on Danish commons. K. Dan. Vidensk. Selsk. 1948, 5, 1–34. [Google Scholar]
- Bos, M.M.; Steffan-Dewenter, I.; Tscharntke, T. The contribution of cacao agroforests to the conservation of lower canopy ant and beetle diversity in Indonesia. Biodivers. Conserv. 2007, 16, 2429–2444. [Google Scholar] [CrossRef]
- Rice, R.A.; Greenberg, R. Cacao cultivation and the conservation of biological diversity. Ambio 2000, 29, 167–172. [Google Scholar] [CrossRef]
- Urquhart DH. Cocoa; Longmans Green and Co: London, UK, 1961. [Google Scholar]
- Bhagwat, S.A.; Willis, K.J.; Birks, H.J.B.; Whittaker, R.J. Agroforestry: A refuge for tropical biodiversity? Trends Ecol. Evol. 2008, 23, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Delabie, J.H.C.; Jahyny, B.; Cardoso do Nascimento, I.; Mariano, C.S.F. Contribution of cocoa plantations to the conservation of native ants (Insecta: Hymenoptera: Formicidae) with a special emphasis on the Atlantic Forest fauna of southern Bahia, Brazil. Biodivers. Conserv. 2007, 16, 2359–2384. [Google Scholar] [CrossRef]
- Ambrecht, I.; Perfecto, I.; Vandermeer, J. Enigmatic biodiversity correlations: Ant diversity responds to diverse resources. Science 2004, 304, 284–286. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Village | Locality Description | GPS Position | Population | Natural Forest | Multistrata Agroforests | Cocoa Agroforests | Annual Crops | Weedy Vegetation |
|---|---|---|---|---|---|---|---|---|
| Macuya | University center for forest research, 12 km from Von Humboldt. Undulated terrain, regular water sources, logged over primary forest, 20+ years old forest on the edge. | S 8°54', W 74°59' | 5 | Natural forest selectively logged 20+ years, on the edges; trees, palms, shrubs, thick layer of fallen leaves and woody material; Closed canopy; tree density 9 trees/25 m2; 95% ground shade, several natural streams and small ponds of standing water. | -- | -- | -- | -- |
| San Alejandro | Undulated with steep slopes, lots of water sources. Local climate allows effective cocoa plantation. | S 8°49', W 75°12' | 450 | -- | -- | Cocoa plantation with various shade trees: Inga sp., Calycophyllum sp., Dipteryx sp., Tabebuia sp. | Cassava mono-cropping; low vegetation density; no shade | Abundant weeds dominated by Imperata grass; compact 1.20 m tall grass vegetation |
| Pimental | Middle steep terrains and plains, dry with occasional water resources, slash-and-burn farmers, common perennial plantations. | S 8°31′, W 74°46′ | 300 | 20+ year old forest, closed canopy 5 m, dense vegetation, humid, relatively steep. Small pond of standing water | Inga edulis, pineapples, 6 years old, tree density up to 7 trees/25 m2; trees 9 m high; layer of fallen leaves; 80% ground shade, small stream and pond of standing water nearby | Piper nigrum, Guazuma crinita six years old; trees 12 m high; vegetation density 6 trees/25 m2; 50% ground shade, small stream and pond of standing water nearby | Cassava mono-cropping; vegetation density low; no shade. | Abundant weeds dominated by Imperata grass; compact 1.20 m tall grass vegetation |
| Antonio Raymondi | Slash-and-burn farmers, mainly colonizers. Deforested plains invaded by Imperata weeds, eroded soils and dry climate. | S 8°22′, W 74°42′ | 200 | -- | 5 m Inga edulis, pineapples, 6 years old, tree density up to 5 trees/25 m2; trees 6 m high; layer of fallen leaves; layer of fallen leaves; 75% ground shade | -- | Cassava cropping; vegetation density low; no shade. | Plentiful Abundant weeds dominated by Imperata; compact 1.20 m tall grass Abundant wminated by Imperata grass; compact 1.20 m tall grass vegetation |
| Diversity Indices | Natural Forests | Multistrata Agroforests | Cocoa Agroforests | Annual Crops | Weedy Grasslands | ANOVA p-value |
|---|---|---|---|---|---|---|
| Totals | ||||||
| Abundance No of ind. | 1348 | 992 | 297 | 277 | 207 | |
| Observed sp. richness No. of sp. | 758 | 540 | 189 | 97 | 64 | |
| Jackknife sp. richness | 853 | 594 | 203 | 105 | 70 | |
| Singletons | 157 | 119 | 70 | 24 | 2 | |
| Unique sp. | 126 | 57 | 18 | 11 | 8 | |
| Shannon-Weiner diversity index | 5.21 | 4.94 | 4.09 | 4.07 | 3.68 | |
| Simpson’s diversity index (1-D) | 0.991 | 0.987 | 0.950 | 0.973 | 0.969 | |
| Means per sample ± SD | ||||||
| No. of samples | 8 | 31 | 32 | 32 | 32 | |
| Observed sp. richness No. of sp./sam. | 109 ± 21.0 a | 18.8 ± 3.16 b | 6.72 ± 1.22 c | 4.13 ± 0.71 c | 3.06 ± 0.76 c | <0.001 |
| Jackknife sp. richness | 148 ± 28.4 a | 24.4 ± 4.10 b | 8.38 ± 1.62 c | 4.72 ± 1.11 c,d | 3.34 ± 0.90 d | <0.001 |
| Sp. density sp./m2 | 0.121 | 0.086 | 0.030 | 0.016 | 0.010 | |
| Shannon-Weiner diversity index | 0.648 ± 0.155 a | 0.181 ± 0.033 b | 0.166 ± 0.04 b,c | 0.141 ± 0.027 c | 0.139 ± 0.029 c | <0.001 |
| Simpson’s diversity index (1-D) | 0.909 ± 0.031 a | 0.883 ± 0.018 a | 0.808 ± 0.066 b | 0.606 ± 0.081 c | 0.554 ± 0.082 d | <0.001 |
| PF | AFS | CF | MC | W | |
|---|---|---|---|---|---|
| PF | 158 | 77 | 51 | 31 | |
| AFS | 52% | 71 | 49 | 26 | |
| CF | 34% | 37% | 28 | 18 | |
| MC | 24% | 28% | 27% | 30 | |
| W | 15% | 17% | 21% | 44% |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perry, J.; Lojka, B.; Quinones Ruiz, L.G.; Van Damme, P.; Houška, J.; Fernandez Cusimamani, E. How natural Forest Conversion Affects Insect Biodiversity in the Peruvian Amazon: Can Agroforestry Help? Forests 2016, 7, 82. https://doi.org/10.3390/f7040082
Perry J, Lojka B, Quinones Ruiz LG, Van Damme P, Houška J, Fernandez Cusimamani E. How natural Forest Conversion Affects Insect Biodiversity in the Peruvian Amazon: Can Agroforestry Help? Forests. 2016; 7(4):82. https://doi.org/10.3390/f7040082
Chicago/Turabian StylePerry, Jitka, Bohdan Lojka, Lourdes G. Quinones Ruiz, Patrick Van Damme, Jakub Houška, and Eloy Fernandez Cusimamani. 2016. "How natural Forest Conversion Affects Insect Biodiversity in the Peruvian Amazon: Can Agroforestry Help?" Forests 7, no. 4: 82. https://doi.org/10.3390/f7040082
APA StylePerry, J., Lojka, B., Quinones Ruiz, L. G., Van Damme, P., Houška, J., & Fernandez Cusimamani, E. (2016). How natural Forest Conversion Affects Insect Biodiversity in the Peruvian Amazon: Can Agroforestry Help? Forests, 7(4), 82. https://doi.org/10.3390/f7040082