
Grand Fir Nutrient Management in the Inland Northwestern USA


Abstract
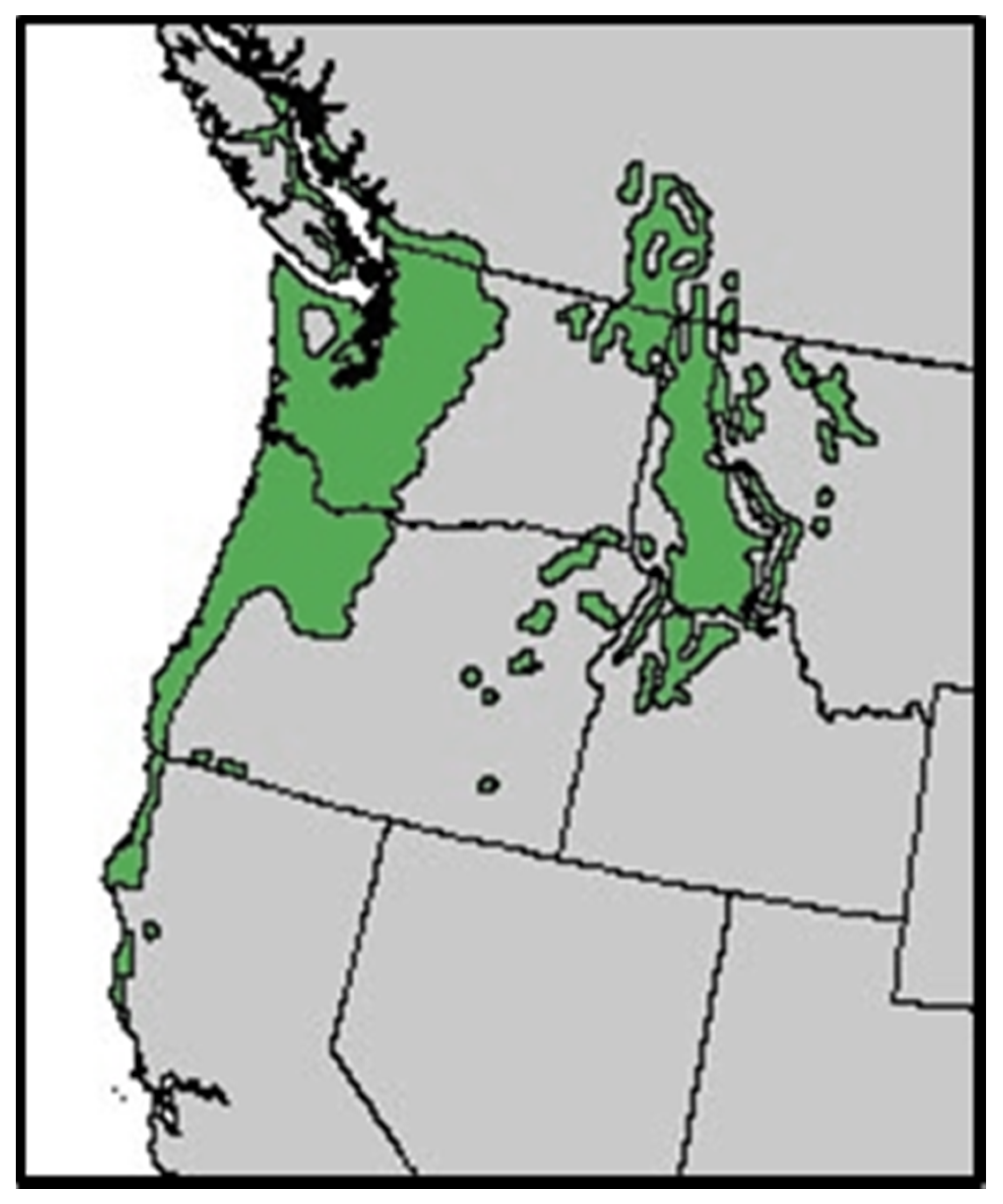
:1. Introduction
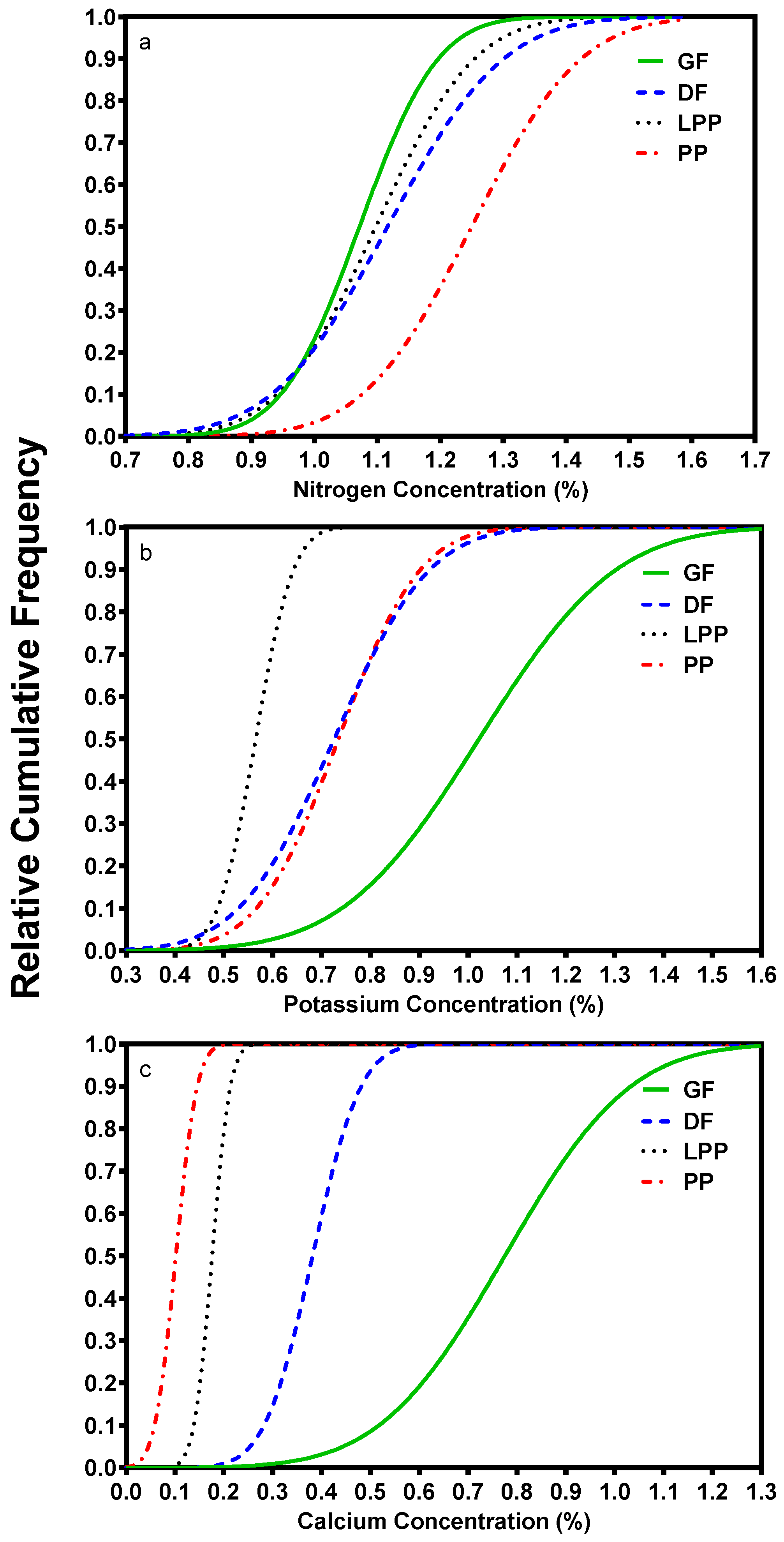
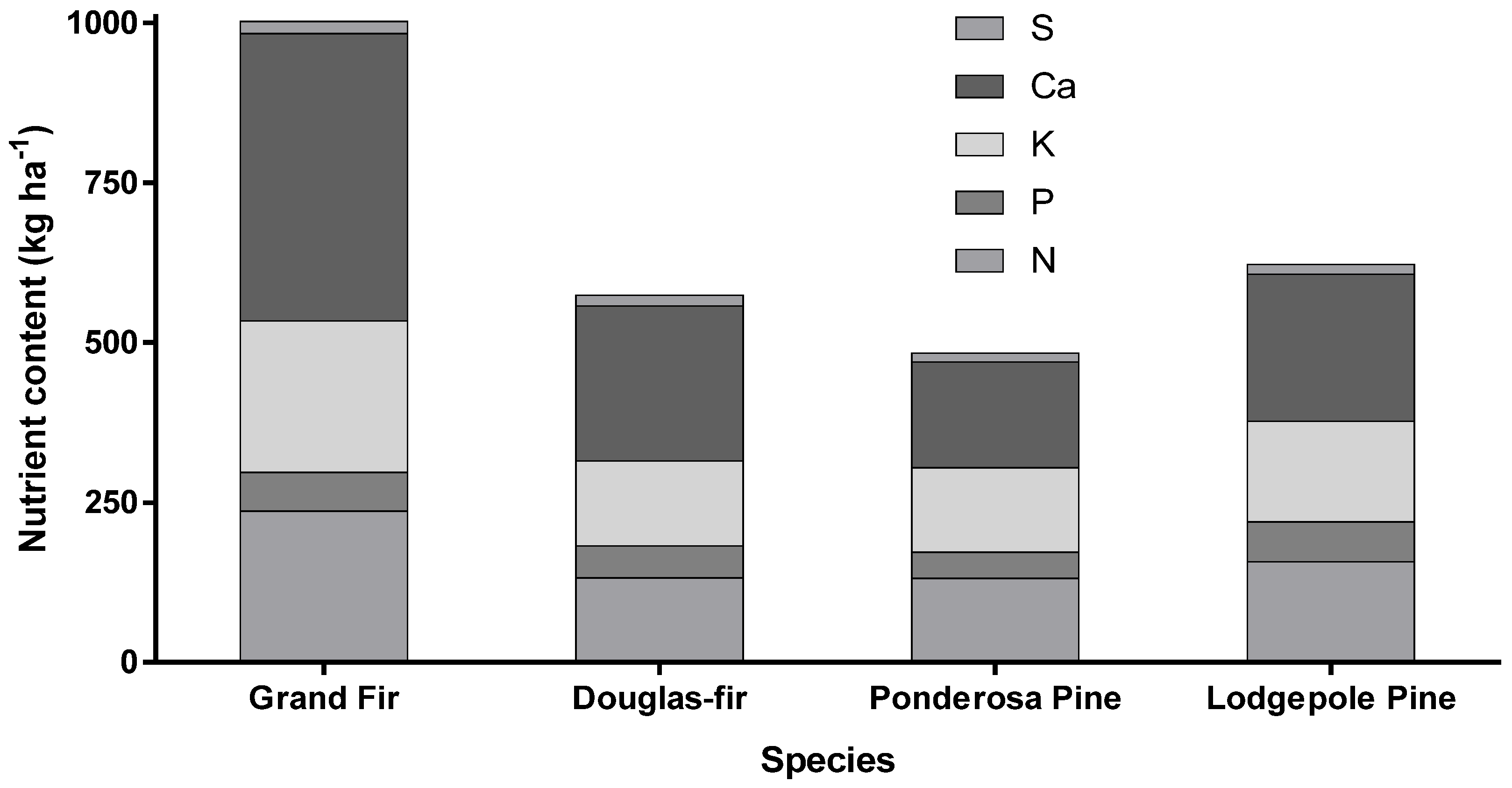
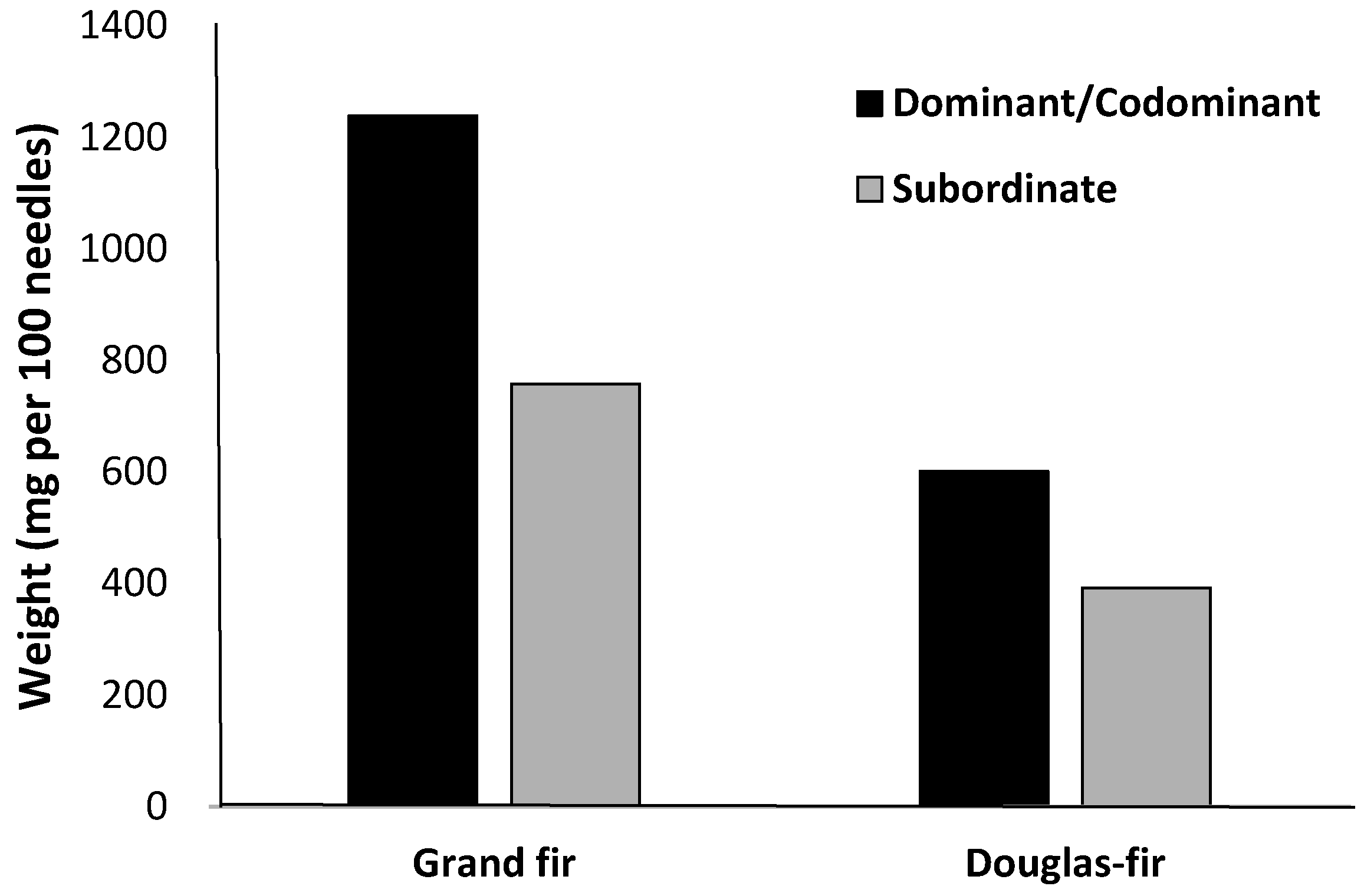
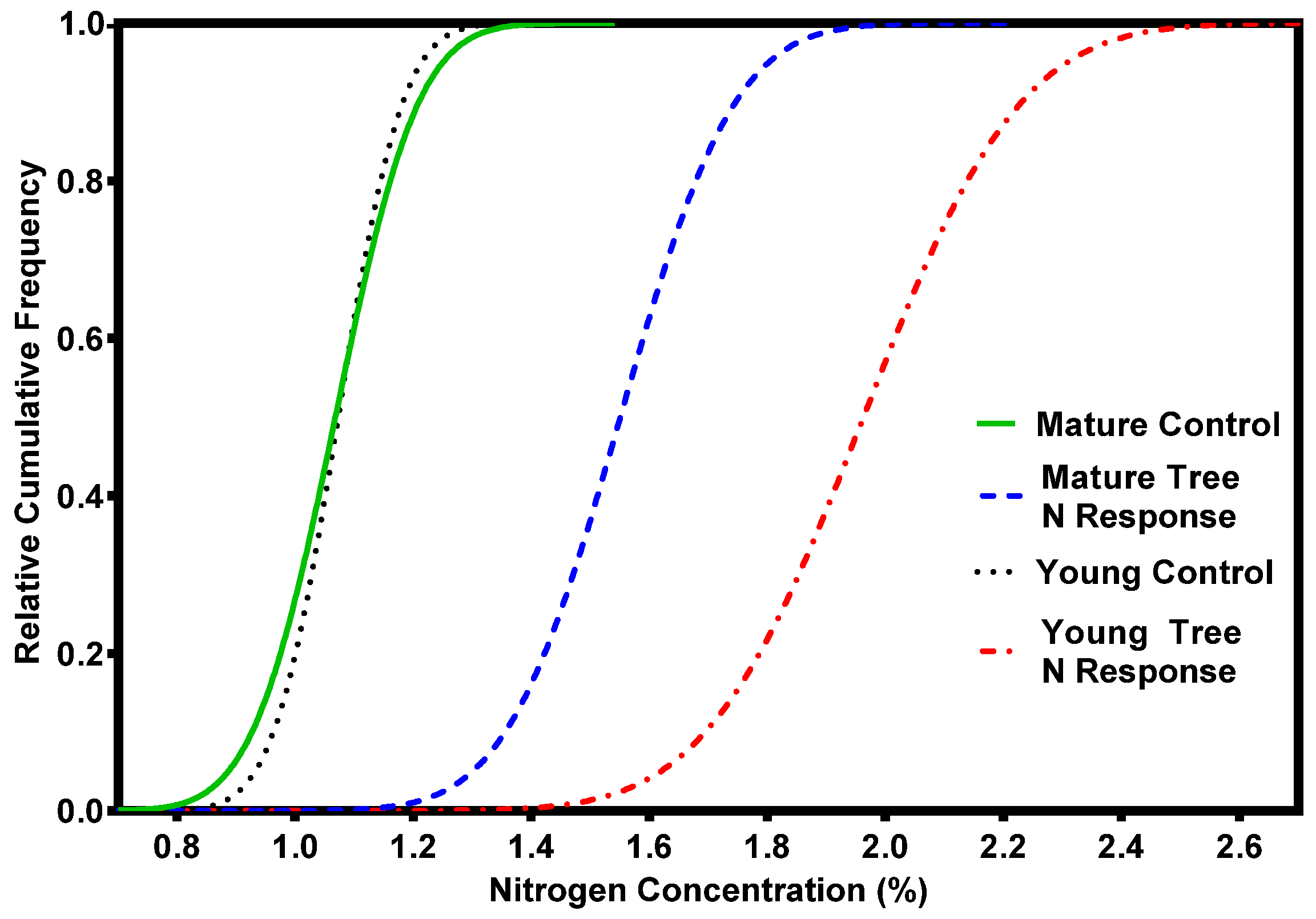
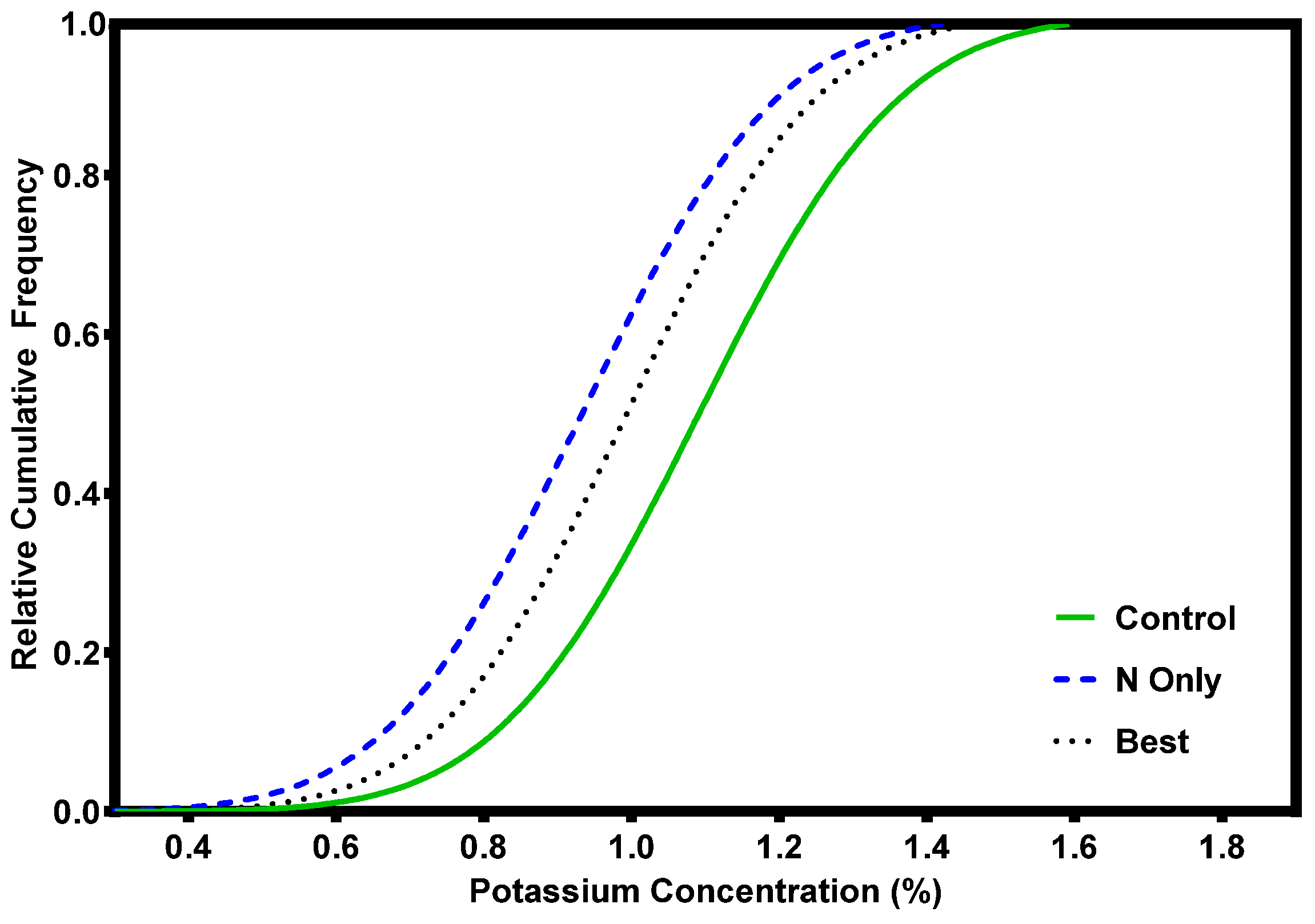
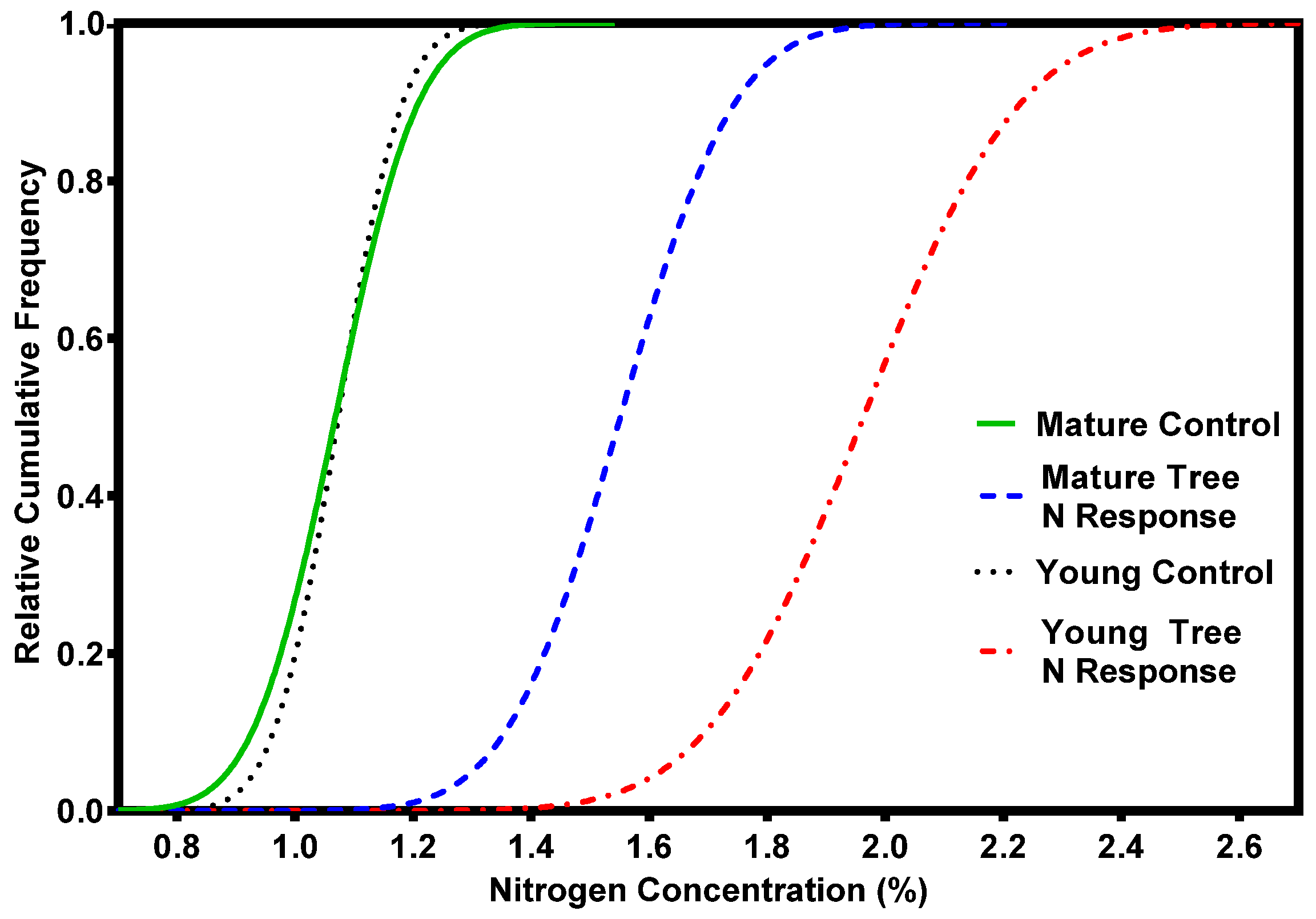
2. Nutrient Characteristics
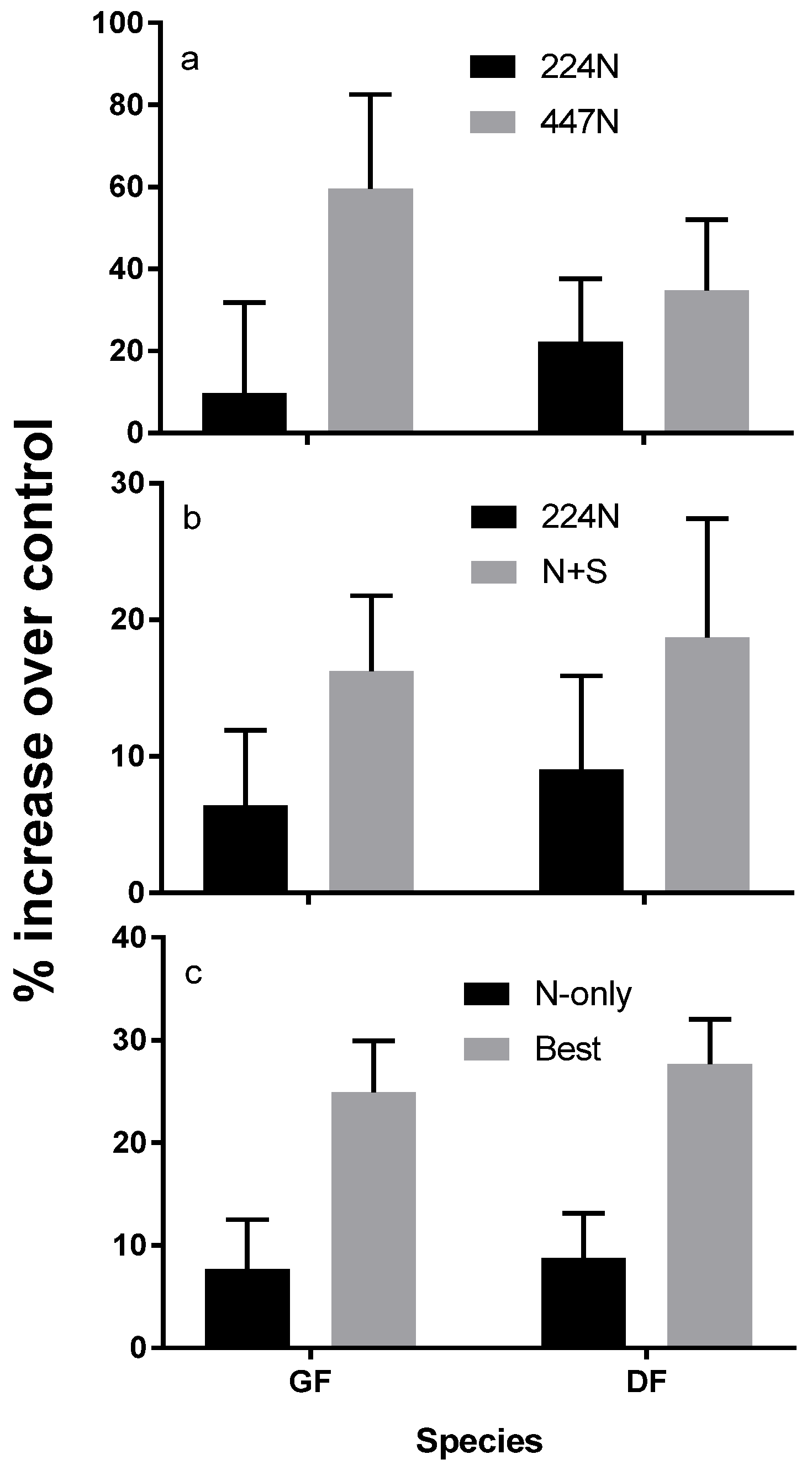
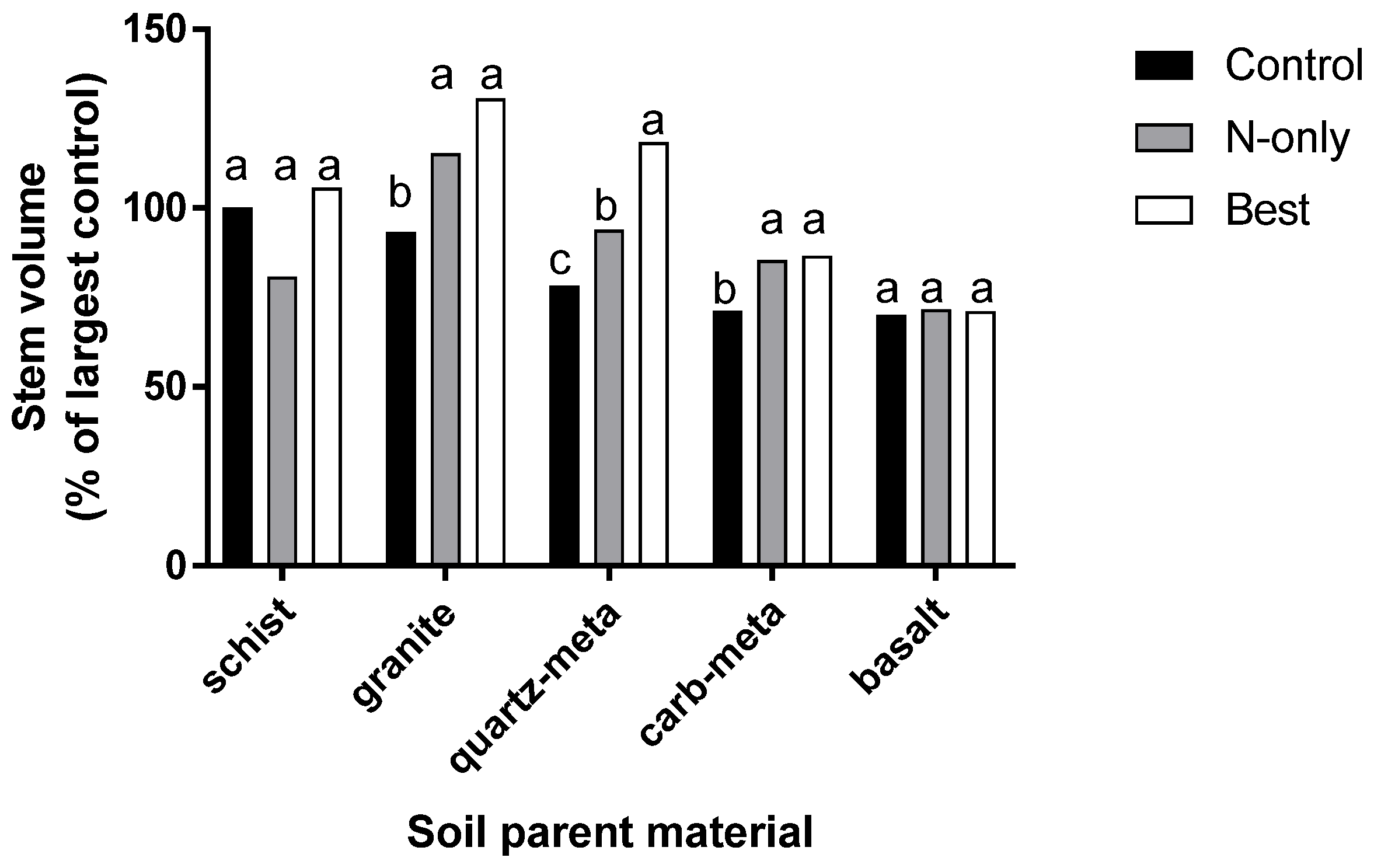
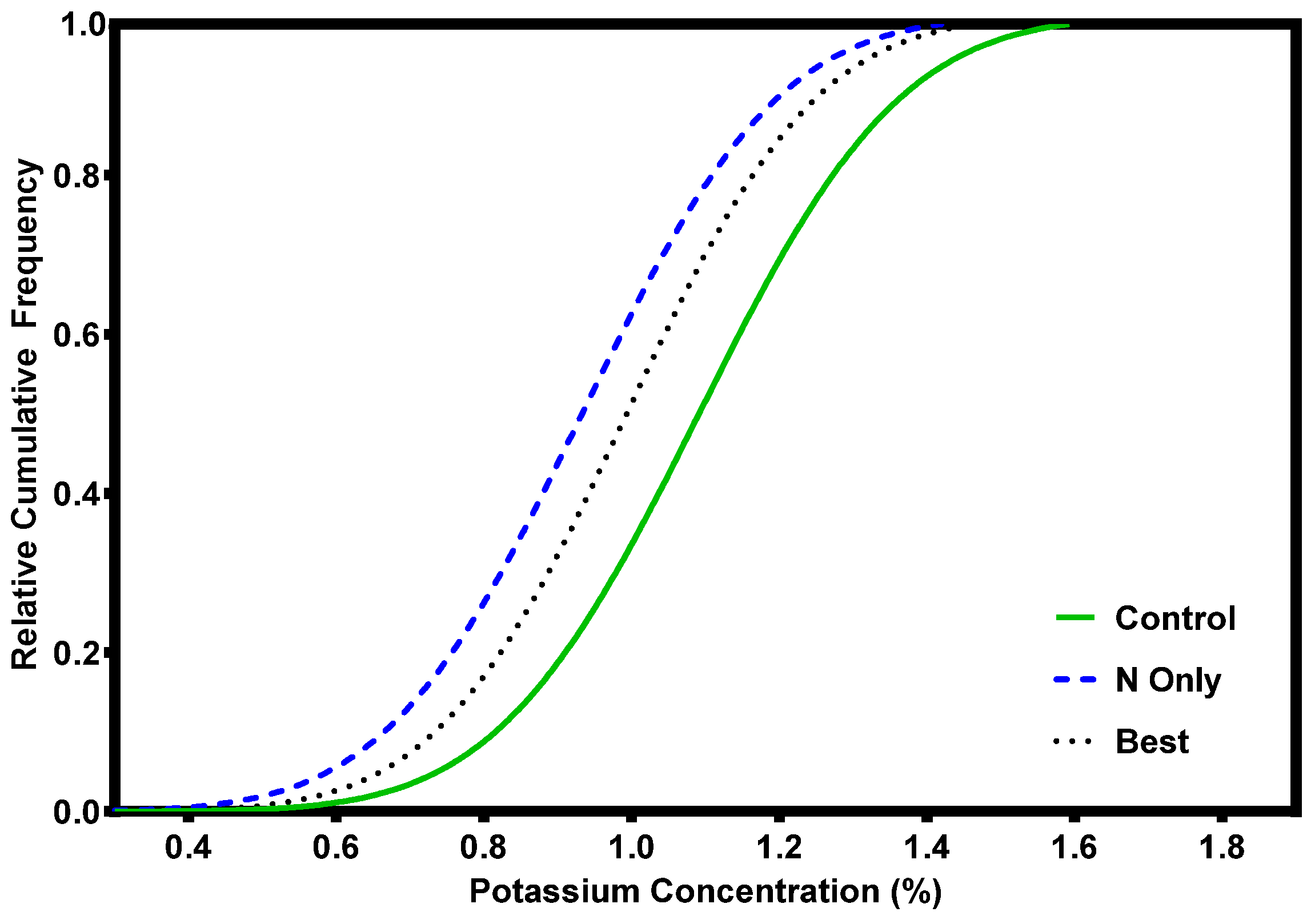
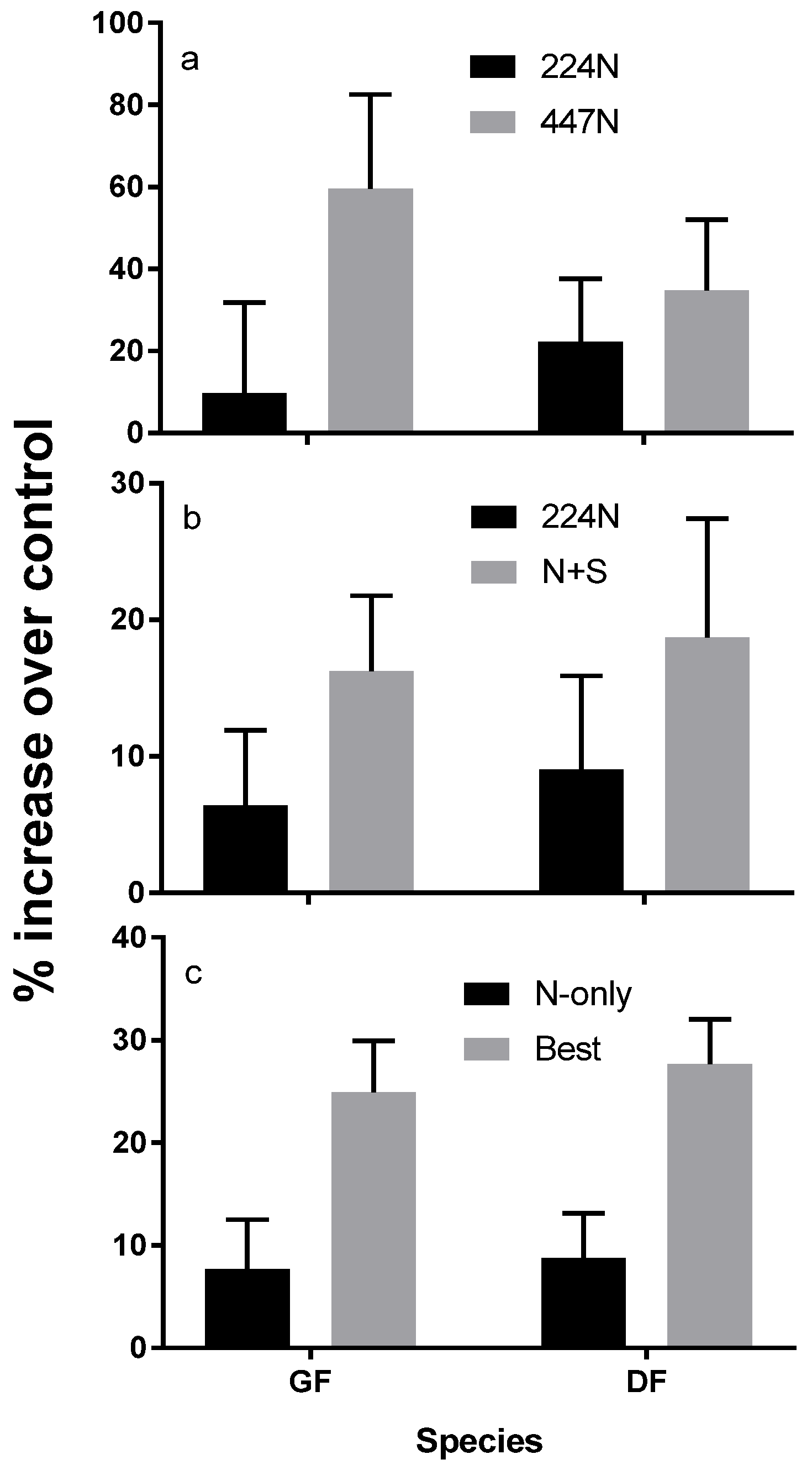
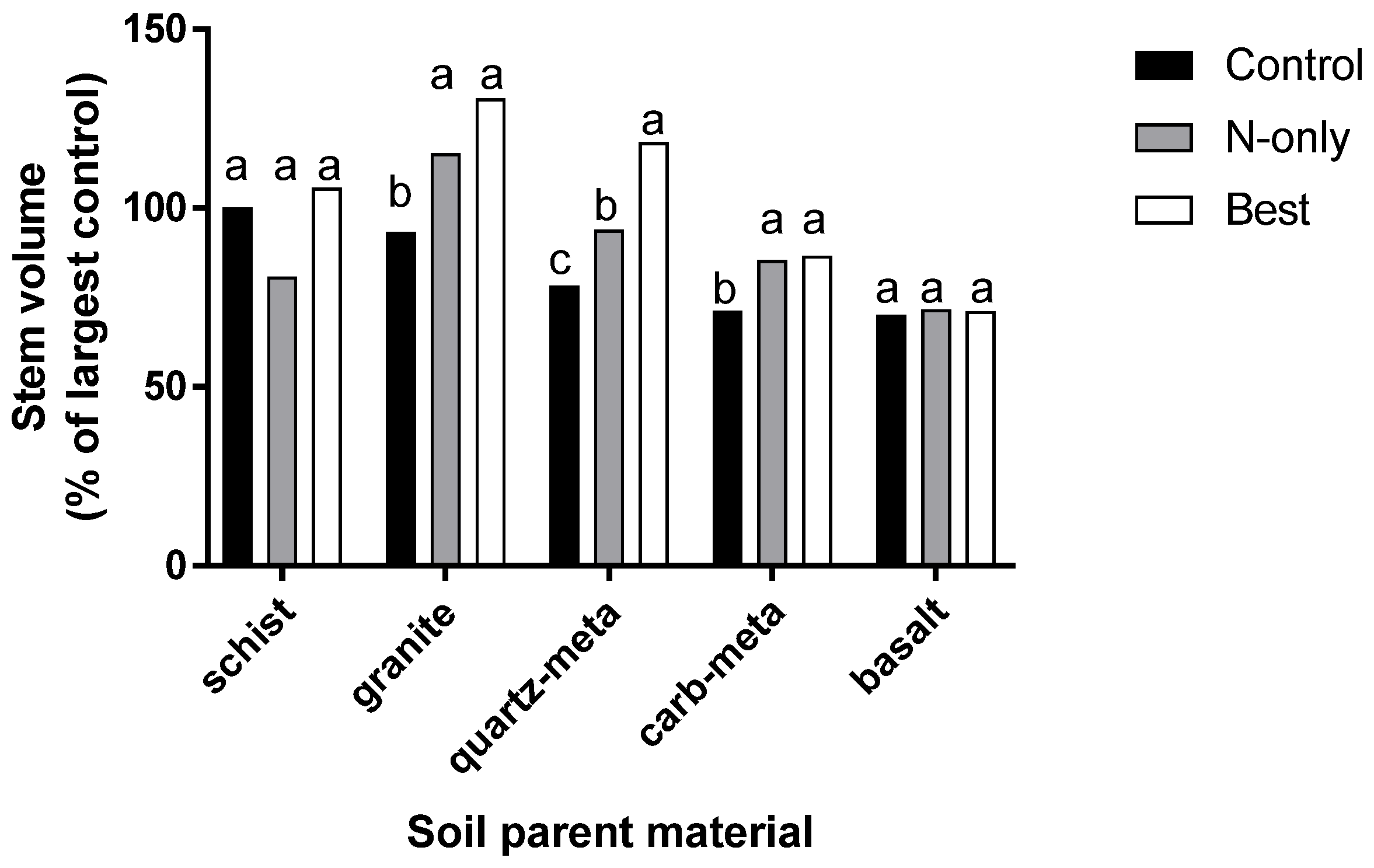
3. Response to Fertilization
4. Economics of Grand Fir Fertilization
5. Management Recommendations
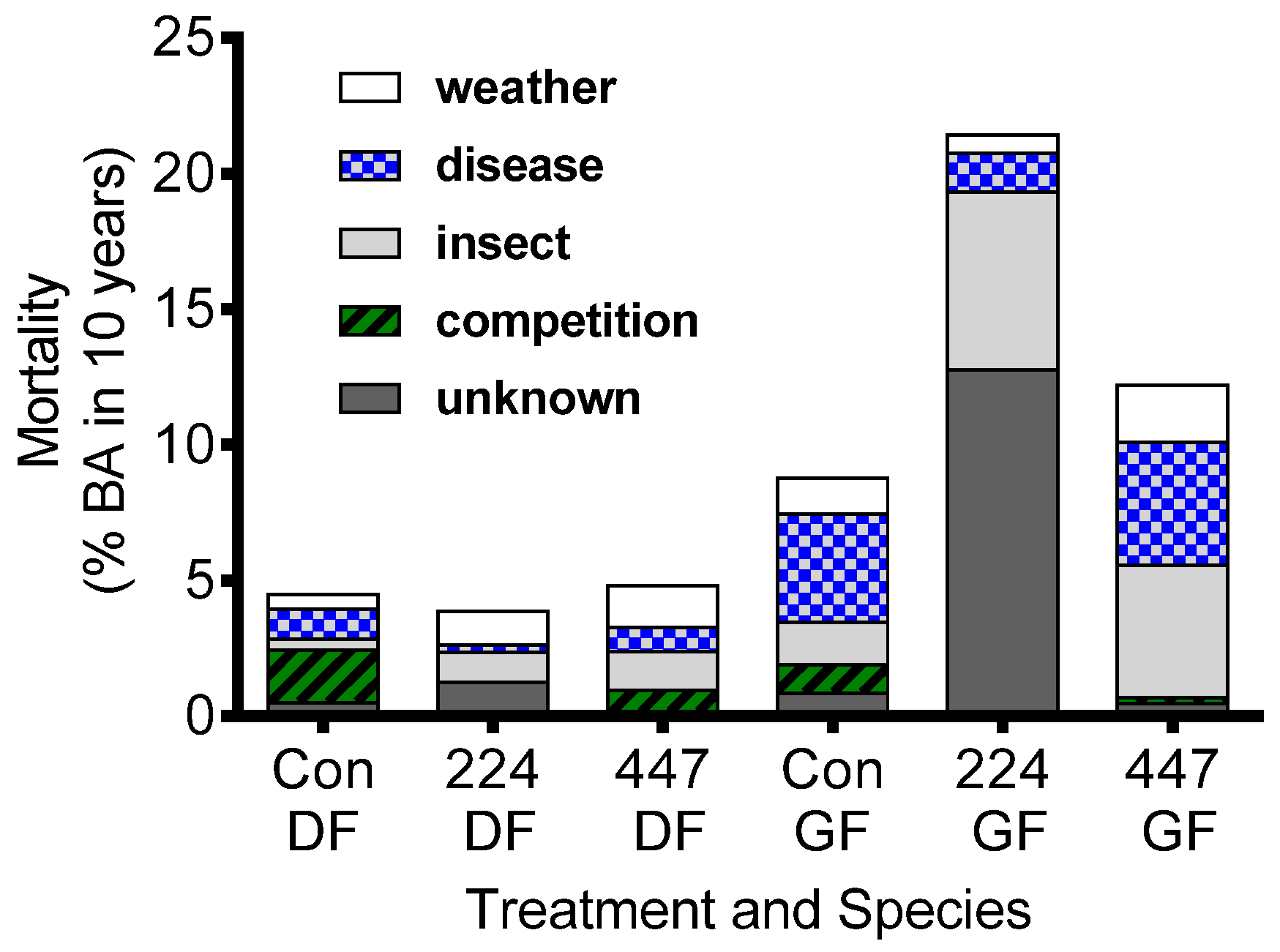
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Foiles, M.W.; Graham, R.T.; Olson, D.F. Grand fir. In Silvics of Forest Trees of the United States, Conifers; Burns, R.M., Honkala, B.H., Eds.; USFS: Washington, DC, USA, 1990; Volume 1, pp. 52–59. [Google Scholar]
- Cooper, S.V.; Neiman, K.E.; Roberts, D.W. Forest Habitat Types of Northern Idaho: A Second Approximation; USFS, Intermountain Research Station: Ogden, UT, USA, 1991.
- Garrison-Johnston, M.T.; Mika, P.G.; Miller, D.L.; Cannon, P.; Johnson, L.R. Ash cap influences on site productivity and fertilizer response in forests of the Inland Northwest. In Volcanic-ash-derived Forest Soils of the Inland Northwest: Properties and implications for management and restoration; USFS, RMRS: Coeur d’Alene, ID, USA, 2007; pp. 137–163. [Google Scholar]
- Pfister, R.D.; Kovalchik, B.L.; Arno, S.F.; Presby, R.C. Forest Habitat Types of Montana; USFS, INT Station: Ogden, UT, USA, 1977.
- Williams, C.K.; Kelley, B.F.; Smith, B.; Lillybridge, T.R. Forested Plant Associations of the Colville National Forest; Pacific Northwest Research Station: Portland, OR, USA, 1995.
- USGS; Little, E.L., Jr. Digital representations of “Atlas of United States Trees”. Available online: http://esp.cr.usgs.gov/data/atlas/little/abiegran.pdf (accessed on 31 August 2016).
- Deitschman, G.H.; Green, A.W. Relations between Western White Pine Site Index and Tree Height of Several Associated Species; USFS: Ogden, UT, USA, 1965.
- Haig, I.T. Second-growth Yield, Stand, and Volume Tables for the Western White Pine Type; USFS, Northern Rocky Mountain Forest Experiment Station: Washington, DC, USA, 1932.
- Stage, A.R.; Renner, D.L.; Chapman, R.C. Selected Yield Tables for Plantations and Natural Stands in the Inland Northwest forests; USFS, INT Research Station: Ogden, UT, USA, 1988.
- McArdle, R.E.; Meyer, W.H.; Bruce, D. The Yield of Douglas-Fir in the Pacific Northwest; USFS: Washington, DC, USA, 1961.
- USFS. Forest Inventory Data Online (FIDO). Available online: http://apps.fs.fed.us/fia/fido/index.html (accessed on 2 Jaunary 2016).
- Simmons, E.A.; Hayes, S.W.; Morgan, T.A.; Keegan, C.E., III; Witt, C. Idaho’s Forest Products Industry and Timber Harvest 2011 with Trends through 2013; USFS: Fort Collins, CO, USA, 2013.
- Moore, J.A.; Mika, P.G.; Shaw, T.M.; Garrison-Johnston, M. Foliar nutrient characteristics of four conifer species in the interior Northwest United States. West. J. Appl. For. 2004, 19, 13–24. [Google Scholar]
- Garrison, M.T.; Moore, J.A. Nutrient Management: A Summary and Review. Available online: http://digital.lib.uidaho.edu/cdm/singleitem/collection/iftnc/id/3357/rec/40 (accessed on 29 September 2016).
- Brown, J.K. Weight and Density of Crowns of Rocky Mountain Conifers; USFS, INT Forest & Range Experiment Station: Ogden, UT, USA, 1978.
- Mika, P.G. Grand Fir: Nutrient Ecology and Response to Fertilization. Available online: http://digital.lib.uidaho.edu/cdm/singleitem/collection/iftnc/id/3446/rec/6 (accessed on 29 September 2016).
- Garrison-Johnston, M. Nutrient Cycling in a North Idaho Conifer Stand. Ph.D. Thesis, University of Idaho, Moscow, ID, USA, 2003. [Google Scholar]
- Walker, R.B.; Gessel, S.P. Mineral Deficiencies of Coastal Northwest Conifers; College of Forest Resources, University of Washington: Seattle, WA, USA, 1991. [Google Scholar]
- Ingestad, T. Mineral nutrient requirements of Pinus silvestris and Picea abies seedlings. Physiol. Plant. 1979, 45, 373–380. [Google Scholar] [CrossRef]
- Ingestad, T. A definition of optimum nutrient requirements in birch seedlings. II. Physiol. Plant. 1971, 24, 118–125. [Google Scholar] [CrossRef]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Coleman, M.D.; Shaw, T.M.; Kimsey, M.J.; Moore, J.A. Nutrition of Douglas-fir in the Inland Northwest. Soil Sci. Soc. Am. J. 2014, 78, 11–22. [Google Scholar] [CrossRef]
- Graham, R.T.; Tonn, J.R. Ten-Year Results of Fertilizing Grand Fir, Western Hemlock, Western Larch, and Douglas-Fir with Nitrogen in Northern Idaho; USFS, INT Station: Ogden, UT, USA, 1985.
- Shaw, T.M.; Kimsey, M. Screening trial data. Intermountain Forestry Cooperative, University of Idaho: Moscow, ID, USA, Unpublished data. 2016. [Google Scholar]
- Mainwaring, D.B.; Maguire, D.A.; Perakis, S.S. Three-year growth response of young Douglas-fir to nitrogen, calcium, phosphorus, and blended fertilizers in Oregon and Washington. For. Ecol. Manag. 2014, 327, 178–188. [Google Scholar] [CrossRef]
- Loewenstein, H.; Pitkin, F.H. Growth Responses and Nutrient Relations of Fertilized and Unfertilized Grand Fir; University of Idaho, Forestry, Wildlife, and Range Experiment Station: Moscow, ID, USA, 1971. [Google Scholar]
- Shaw, T.; Coleman, M.; Kimsey, M.; Mika, P. Forest Health and Nutrition Ten-year Growth, Mortality and Foliar Nutrition; Intermountain Forest Tree Nutrition Cooperative: Moscow, ID, USA, 2014. [Google Scholar]
- Garrison, M.T.; Moore, J.A.; Shaw, T.M.; Mika, P.G. Foliar nutrient and tree growth response of mixed-conifer stands to three fertilization treatments in northeast Oregon and north central Washington. For. Ecol. Manag. 2000, 132, 183–198. [Google Scholar] [CrossRef]
- Moore, J.A.; Mika, P.G.; Schwandt, J.W.; Shaw, T.M. Nutrition and forest health. In Interior Cedar-Hemlock-White Pine Forests: Ecology and Management; Washington State University: Spokane, WA, USA, 1994; pp. 173–176. [Google Scholar]
- Moore, J.A.; Mika, P.G.; Vanderploeg, J.L. Nitrogen fertilizer response of Rocky Mountain Douglas-fir by geographic area across the Inland Northwest. West. J. Appl. For. 1991, 6, 94–98. [Google Scholar]
- Garrison, M.T.; Moore, J.A.; Mika, P.G.; Shaw, T.M. Six-Year Growth Response of the Umatilla and Four-Year Growth Response of the Okanogan Mixed Conifer Stands to N, N + K and N + S Fertilization. Available online: http://digital.lib.uidaho.edu/cdm/ref/collection/iftnc/id/3358 (accessed on 3 October 2016).
- Shafii, B.; Moore, J.A.; Olson, J.R. Effects of nitrogen fertilization on growth of grand fir and Douglas-fir stands in northern Idaho. West. J. Appl. For. 1989, 4, 54–57. [Google Scholar]
- Scanlin, D.C.; Lowenstein, H. Forest Fertilization in the Intermountain Region-six Year Results; University of Idaho, Forestry, Wildlife and Range Experiment Station: Moscow, ID, USA, 1980. [Google Scholar]
- Scanlin, D.C.; Loewenstein, H.; Pitkin, F.H. Two-year Response of North Idaho Stands of Douglas-Fir and Grand Fir to Urea Fertilizer and Thinning; University of Idaho: Moscow, ID, USA, 1976. [Google Scholar]
- Kimmins, J.P. Forest Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Shaw, T.M.; Johnson, L.R. Two-Year Growth Response of Douglas-fir and Ponderosa Pine Seedlings to Boron and Multi-Nutrient Fertilization in Northeast Oregon. Available online: http://digital.lib.uidaho.edu/cdm/ref/collection/iftnc/id/4860 (accessed on 4 October 2016).
- Fan, Z.; Moore, J.A.; Osborne, H.L. Three-year response of ponderosa pine seedlings to controlled-release fertilizer applied at planting. West. J. Appl. For. 2002, 17, 154–164. [Google Scholar]
- Oliver, C.D.; O’Hara, K.L.; McFadden, G.; Nagame, L. Concepts of thinning regimes. In Douglas-Fir: Stand Management for the Future; Oliver, C.D., Hanley, D.P., Johnson, J.A., Eds.; Institute of Forest Resources: Seattle, WA, USA, 1986; pp. 246–257. [Google Scholar]
- Weiskittel, A.; Gould, P.; Temesgen, H. Sources of variation in the self-thinning boundary line for three species with varying levels of shade tolerance. For. Sci. 2009, 55, 84–93. [Google Scholar]
- Loewenstein, H.; Pitkin, F.H. Response of grand fir and western white pine to fertilizer applications. Northwest Sci. 1963, 37, 23–30. [Google Scholar]
- Graham, R.T.; Tonn, J.R. Response of Grand Fir, Western Hemlock, Western White Pine, Western Larch, and Douglas-Fir to Nitrogen Fertilizer in Northern Idaho; USFS, INT: Ogden, UT, USA, 1979.
- Powers, R.F. Response of California true fir to fertilization. In Forest Fertilization Conference; Gessel, S.P., Ed.; University of Washington: Union, WA, USA, 1979; pp. 95–101. [Google Scholar]
- Olson, J. Four-Year Fertilization and Thinning Response Analysis of Douglas-fir and Grand Fir Stands in Northern Idaho; Potlatch Corporation: Lewiston, ID, USA, 1979. [Google Scholar]
- Olson, J. Response of Intermountain Grand Fir and Douglas-Fir Stand Types to Nitrogen Fertilization and Thinning; Potlatch Corporation: Lewiston, ID, USA, 1981. [Google Scholar]
- Olson, J.; Hatch, C.R. Volume response of intermountain grand fir stand types to nitrogen fertilizer and thinning treatments. In The Biology and Management of True Fir in the Pacific Northwest; University of Washington: Seattle, WA, USA, 1981; pp. 247–250. [Google Scholar]
- Chase, C.W.; Kimsey, M.J.; Shaw, T.M.; Coleman, M.D. The response of light, water, and nutrient availability to pre-commercial thinning in dry inland Douglas-fir forests. For. Ecol. Manag. 2016, 363, 98–109. [Google Scholar] [CrossRef]
- Scanlin, D.C.; Loewenstein, H. Response of inland Douglas-fir and grand-fir to thinning and nitrogen fertilization in northern Idaho. In Forest Fertilization; Gessel, S.P., Kenady, R.M., Atkinson, W.A., Eds.; University of Washington: Alderbrook Inn, Union, WA, USA, 1979; pp. 82–88. [Google Scholar]
- Cochran, P.H. Response of Thinned White Fir Stands to Fertilization with Nitrogen Plus Sulfur; USFS, PNW Research Station: Portland, OR, USA, 1991.
- Shen, G.; Moore, J.A.; Hatch, C.R. The effect of nitrogen fertilization, rock type, and habitat type on individual tree mortality. For. Sci. 2001, 47, 203–213. [Google Scholar]
- Herms, D.A. Effects of fertilization on insect resistance of woody ornamental plants: Reassessing an entrenched paradigm. Environ. Entomol. 2002, 31, 923–933. [Google Scholar] [CrossRef]
- Stoszek, K.J.; Mika, P.J. Douglas-fir tussock moth outbreaks related to stand conditions and pest management. In 1980 SAF Convention: Land Use Allocation; Society of American Foresters: Bethesda, MD, USA, 1981; pp. 134–137. [Google Scholar]
- Mika, P.G.; Moore, J.A. Foliar potassium status explains Douglas-fir response to nitrogen fertilization in the inland Northwest, USA. Water Air Soil Pollut. 1990, 54, 477–491. [Google Scholar] [CrossRef]
- Schwandt, J.W. Fertilization effects on root disease in two Douglas-fir stands in northern Idaho. In Proceedings of the Fiftieth Western International Forest Disease Work Conference; USFS: Powell River, BC, Canada, 2002; pp. 1–6. [Google Scholar]
- Shaw, T.M.; Moore, J.A.; Marshall, J.D. Root chemistry of Douglas-fir seedlings grown under different nitrogen and potassium regimes. Can. Jour. For. Res. 1998, 28, 1566–1573. [Google Scholar] [CrossRef]
- Mandzak, J.M.; Moore, J.A. The role of nutrition in the health of the inland western forest. J. Sustain. For. 1994, 2, 191–210. [Google Scholar] [CrossRef]
- Miller, D.L.; Gravelle, P.J. Species Selection Guidelines for Planting, Natural Regeneration and Crop Tree Selection on Potlatch Land in Northern Idaho; Potlatch Corporation: Lewiston, ID, USA, 2009. [Google Scholar]
- Will, R.E.; Munger, G.T.; Zhang, Y.; Borders, B.E. Effects of annual fertilization and complete competition control on current annual increment, foliar development, and growth efficiency of different aged Pinus taeda stands. Can. J. For. Res. 2002, 32, 1728–1740. [Google Scholar] [CrossRef]
- Miller, R.E.; Webster, S.R. Fertilizer response in mature stands of Douglas-fir. In Forest Fertilization; University of Washington: Union, WA, USA, 1979; pp. 126–132. [Google Scholar]
- Miller, R.E.; Fight, R.D. Fertilizing Douglas-fir Forests; USFS, PNW Research Station: Portland, OR, USA, 1979.
- Olson, J.R. Experience with nitrogen fertilization in northern Idaho. In Forest Fertilization: Sustaining and Improving Nutrition and Growth in Western Forests; Chappell, H.N., Weetman, G.F., Miller, R.E., Eds.; University of Washington: Seattle, WA, USA, 1992; pp. 232–236. [Google Scholar]
- National Timber Tax Website. Timber Fertilization. Available online: http://www.timbertax.org/research/revenuerulings/bda/04-62/ (accessed on 14 January 2015).
- Shaw, T.M.; Kimsey, M.; Parent, D.R. Growth Response of an Operationally Fertilized Stand at Brickel Creek, North Idaho; IFTNC, University of Idaho: Moscow, ID, USA, 2010. [Google Scholar]
- Olson, J. The fertilization decision: Using what we have learned to evaluate a nitrogen fertilization program. In IFTNC Fertilization Workshop; IFTNC: Moscow, ID, USA, 1988; pp. 1–15. [Google Scholar]
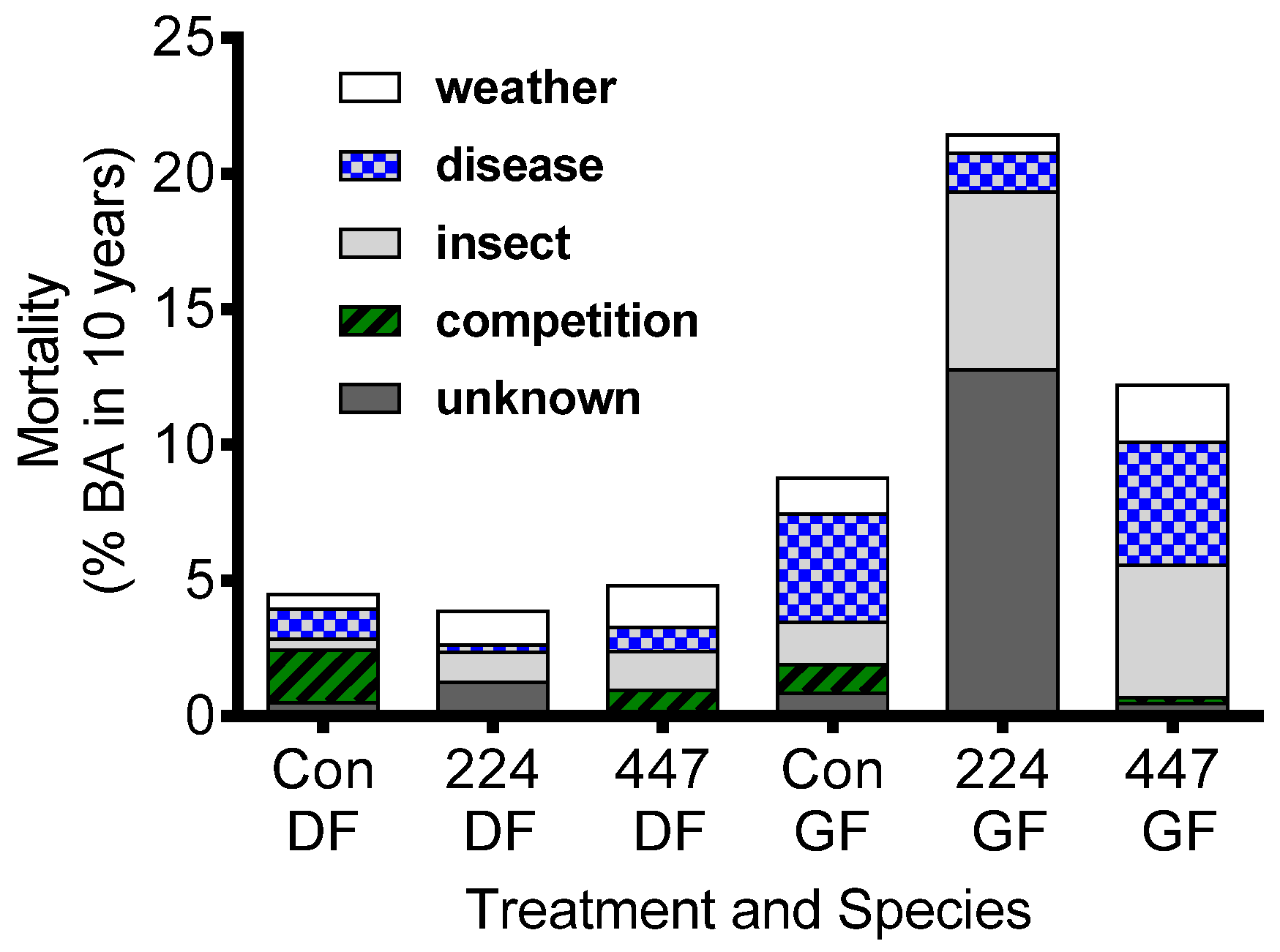

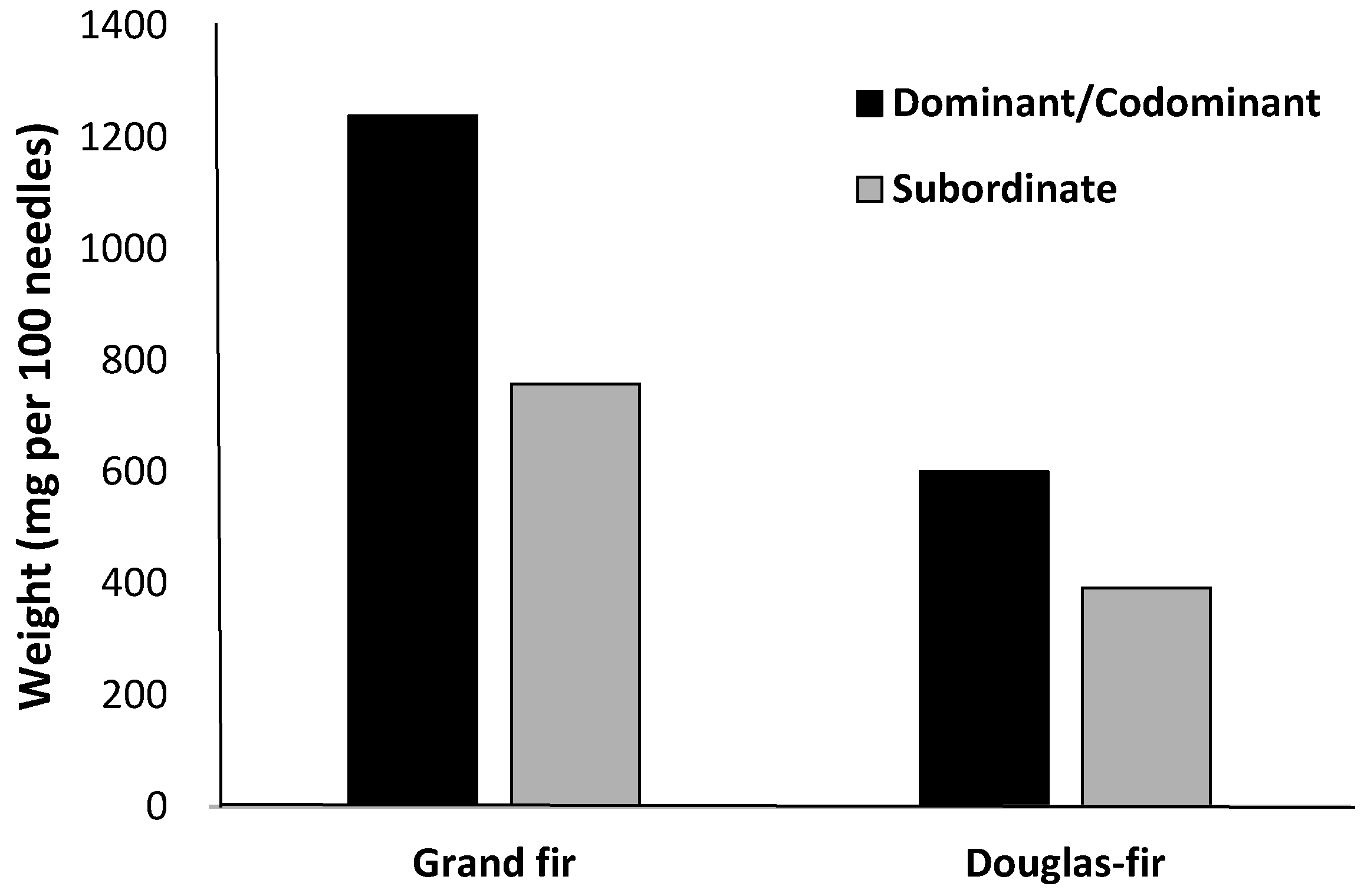
{kind=link}
{kind=link}
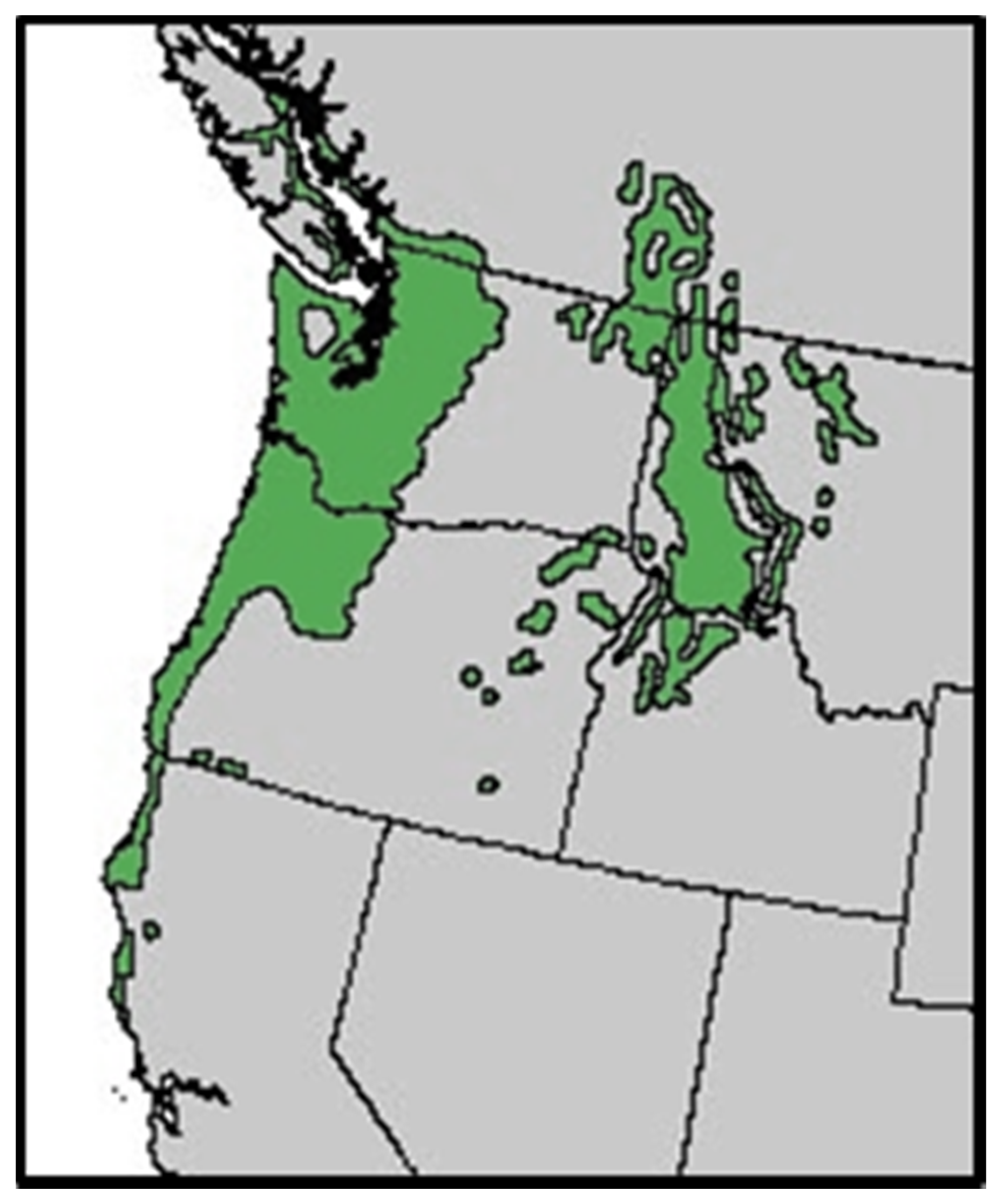
{kind=link}
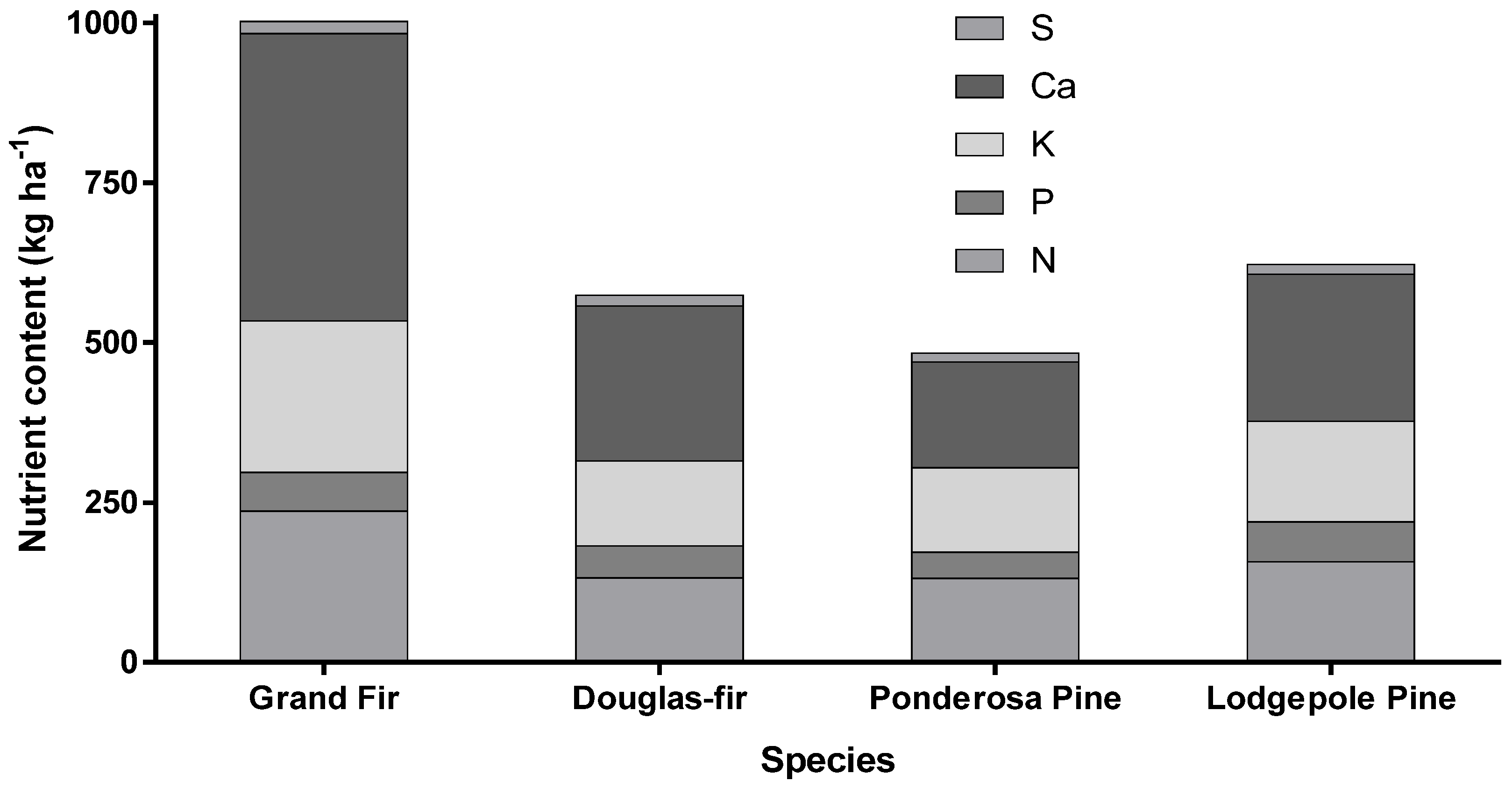
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age | Grand Fir | Western White Pine | Western Larch | Inland Douglas-Fir | Coastal Douglas-Fir 1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Height (m) 2 | Volume (m3) 3 | Height (m) 2 | Volume (m3) 4 | Height (m) 2 | Volume (m3) 5 | Height (m) 2 | Volume (m3) 6 | Height (m) 7 | Volume (m3) 8 | |
| 50 | 20.6 | 74.1 | 21.5 | 140.3 | 22.8 | 37.9 | 20.0 | 73.4 | 23.7 | 110.0 |
| 100 | 36.3 | 367.1 | 42.4 | 393.3 | 37.8 | 141.2 | 35.4 | 206.6 | 33.8 | 260.1 |
| Foliar Concentration | GF | DF | LP | PP | Ingestad Ratios * |
|---|---|---|---|---|---|
| N (mg·g−1) | 11.5 | 14.0 | 12.0 | 11.0 | 100 |
| P (mg·g−1) | 1.5 | 1.2 | 1.2 | 0.8 | 16 |
| K (mg·g−1) | 5.8 | 6.0 | 5.0 | 4.8 | 50 |
| S (mg·g−1)) | 0.8 | 1.1 | 0.9 | 0.8 | 7 |
| Cu (µg·g−1) | 3 | 2 | 3 | 3 | 0.03 |
| B (µg·g−1) | 10 | 10 | 4.3 | 20 | 0.2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parent, D.R.; Coleman, M.D. Grand Fir Nutrient Management in the Inland Northwestern USA. Forests 2016, 7, 261. https://doi.org/10.3390/f7110261
Parent DR, Coleman MD. Grand Fir Nutrient Management in the Inland Northwestern USA. Forests. 2016; 7(11):261. https://doi.org/10.3390/f7110261
Chicago/Turabian StyleParent, Dennis R., and Mark D. Coleman. 2016. "Grand Fir Nutrient Management in the Inland Northwestern USA" Forests 7, no. 11: 261. https://doi.org/10.3390/f7110261
APA StyleParent, D. R., & Coleman, M. D. (2016). Grand Fir Nutrient Management in the Inland Northwestern USA. Forests, 7(11), 261. https://doi.org/10.3390/f7110261