1. Introduction
Bark beetles (Coleoptera: Curculionidae: Scolytinae) exploit the subcortical tissues of woody species for sustenance and reproductive shelter. A select few of over 600 bark beetle species erupt from innocuous levels to populations capable of altering forest stand characteristics and composition. For example, the mountain pine beetle (
Dendroctonus ponderosae Hopk.) is capable of transitioning from endemic populations, to incipient-epidemic, to epidemic within a relatively short time period [
1]. Within these population phases, the behavior of the mountain pine beetle can vary depending upon population levels [
2]. For instance, as mountain pine beetle populations increase, large-diameter, heavily defended trees are attacked compared to endemic beetle populations that attack weakened and predisposed trees that maintain populations not capable of overcoming well defended trees [
2]. Eruptive mountain pine beetle-caused mortality from 1990 through 2010 was more than ten times greater than any previously recorded [
3].
Conifer defense mechanisms that occur when bark beetles attack include oleoresin produced in a network of preformed resin ducts (reviewed in [
4]). The amount of preformed oleoresin released from the ducts upon wounding by the beetle is known as a quantitative defense. The released oleoresins serve as a physical barrier to penetrating organisms by flushing and sealing the wounds (reviewed in [
5]). The constituents of the oleoresin, known as qualitative defenses, are also important for bark penetrating organisms and their symbionts (reviewed in [
6]). The oleoresins mainly comprise secondary defensive compounds such as monoterpenes, diterpenes, sesquiterpenes, and stilbene phenolics [
4,
6]. Monoterpenes are the fraction of resin most deleterious to attacking bark beetles, while diterpene and stilbene phenolics appear to possess antifungal properties and are inducible upon penetration of the bark [
4,
6]. The preformed (constitutive) monoterpene efficacy against attacking
Ips species in red pine (
Pinus resinosa) and jack pine (
Pinus bansiana) killed 60% of the beetles, while induced concentrations produced de novo killed 90% of the beetles [
7]. Once an attack from bark beetles is underway, the induced concentration of secondary monoterpene compounds is significantly greater. The induced production of monoterpenes is also elevated when phytopathogenic fungal associates of bark beetles are introduced [
7].
The ability of trees to produce a greater amount of induced defensive compounds than preformed compounds after an attack occurs in several conifer families [
8]. Particular to this study, monoterpene cyclase activity was greater upon wounding in both lodgepole pine and ponderosa pines [
8]. To overcome these induced defenses, bark beetles have evolved a suite of semiochemical communications derived from host monoterpenes (reviewed in [
9]). The monoterpenes are oxidized as the beetles mine subcortical tissues, which in turn are emitted from the entry wounds and frass as semiochemicals for conspecific attraction or aggregation pheromones [
9,
10]. This attraction results in an attack
en masse by recruiting conspecifics to individual “focal” trees, which subsequently overcome the host defenses [
9]. Furthermore,
Dendroctonus species have evolved in a close relationship with their hosts, expressing a tolerance for host species defense compounds while lacking a tolerance for non-host volatiles. Minor differences in host oleoresin chemistry has been demonstrated to affect
Dendroctonus species oviposition behavior and fecundity [
11]. This tolerance of defensive compounds, or lack thereof, may explain in part the high level of host specificity seen in many bark beetles [
12].
The deleterious effects of conifer monoterpenes on bark beetle adults, eggs, and associated fungi are well documented in laboratory and field assays [
6,
13]. The toxicity of α-pinene to mountain pine beetles exposed to ecologically relevant doses suggests low concentrations lack a strong insecticidal property, while higher concentrations had more than 90% lethality [
14]. However, α-pinene is oxidized by attacking mountain pine beetles to produce an aggregation pheromone,
trans-verbenol [
9], and has attraction properties within a range of concentrations [
15]. Thus, increased concentrations of α-pinene in one tree species over another may synergize an aggregated or mass attack of the host containing greater levels of the monoterpene, provided the attacking beetles can tolerate the monoterpene and synthesize the specific monoterpene concentrations.
Bark beetles are well known to exert selective pressure on tree chemistry. Raffa and Berryman [
16] found no differences in qualitative defenses of monoterpene compositions between trees that were attacked versus trees that resisted attack, though resistant trees had greater quantitative defenses after attack. Additionally, trees killed by mountain pine beetles in Colorado had fewer and smaller vertical resin ducts than trees not attacked [
17]. Limonene is one of the secondary host defense compounds present in ponderosa pine and lodgepole pine phloem layers and resins [
18,
19,
20]. This monoterpene has toxic properties against several species of bark beetles [
21,
22,
23], particularly at induced concentrations [
13]. Levels of induced limonene after an initial beetle attack rose to 95% more than that of preformed limonene in lodgepole pine [
13]. Ponderosa pines from areas that had survived historic outbreaks of the Western pine beetle (
D. brevicomis LeConte) had higher preformed concentrations of limonene [
24]. In another study, ponderosa pine trees with higher preformed limonene coupled with high oleoresin flow resisted attack over trees lacking these traits [
25].
In addition to limonene, another monoterpene of particular interest to beetle–host interactions is β-phellandrene. In lodgepole pine, β-phellandrene has been found to be the most abundant compound in oleoresin, but β-phellandrene is proportionally low to non-existent in ponderosa pine [
19,
20]. β-phellandrene’s role as a defensive compound against mountain pine beetle is not yet known. β-phellandrene appears to have less toxicity to bark beetles when compared with other defensive monoterpenes found in relatively small proportions in some oleoresins [
23]. However, β-phellandrene increased attractiveness of
Ips pini (Say) to its aggregation pheromone [
26], which suggests this compound may have kairomonal properties and demonstrates the variation in bark beetle adaptation by the beneficial utilization of these compounds.
In Colorado, the major hosts of the native-mountain pine beetle are lodgepole pine (
Pinus contorta Dougl.) and ponderosa pine (
Pinus ponderosa Lawson), while less abundant limber pine (
Pinus flexilis James) and bristlecone pine (
Pinus aristata Englm.) are also host species. Historical outbreaks of the mountain pine beetle failed to spread into adjacent alternate host stands from the host species for which they developed, though current outbreaks of this beetle have successfully caused adjacent alternate host mortality as populations spread from higher elevation lodgepole pine stands into lower elevation ponderosa pine-dominated stands [
27]. In these lodgepole-ponderosa pine ecotone stands, ponderosa pine was attacked as frequently as lodgepole pine [
27]. Additionally, recent studies of mountain pine beetle host selection between lodgepole pine and ponderosa pine favored ponderosa pine 2:1 in field-choice experiments and laboratory bioassays [
28]. Those studies were conducted with cut-log and bark/phloem units that offered no quantitative host defenses (resin flow) against attack. Therefore, our objective was to assess the preformed host defenses from both pine species growing together in ecotone stands where the mountain pine beetle could choose either host. We asked two specific questions: (1) does the preformed quantitative host defense (amount of resin) and (2) the preformed qualitative host defenses (monoterpene constituents) differ between lodgepole and ponderosa pines? We addressed these questions by comparing the exuded preformed oleoresins from the two hosts.
2. Materials and Methods
2.1. Study Sites
Three sites were selected in 2011 within the lodgepole-ponderosa pine ecotone between 2568–2648 m along the Front Range of the Rocky Mountains, CO, USA. Two sites were located on the Arapaho-Roosevelt National Forest (Molly Lake area, UTM NAD-83 Easting Northing: 13N 449861 4512902; Pennock Pass Area, UTM 13N 458090 4492379), and one site was in Rocky Mountain National Park (Little Horseshoe Park, UTM 13N 448546 4471386). To reduce potential site condition and tree size influences on resin amounts, we selected paired-trees of lodgepole and ponderosa pines from each site that were growing in similar site conditions in the same stands that appeared healthy and free of bark beetle attack and that were within 2.5 cm DBH from each other (diameter at breast height; 1.4 m from forest floor). We sampled a total of 56 pairs of trees with 28 pairs within 36.3 ha at the Molly Lake area, 17 pairs in 24.2 ha at the Pennock Pass area, and 11 pairs within 25.3 ha at the Little Horseshoe Park area; n = 112 trees).
2.2. Quantitative Host Defense Mechanisms
In 2011, preformed oleoresin was collected from tree stems at 1.4 m from the forest floor for 24 h (±1 h). Disks of outer bark and phloem down to the xylem were removed on the north and south aspects of each tree using an 11-mm-diameter punch (#14; C.S. Osbourne & Company, Harrison, NJ, USA). Both north and south sides of trees were sampled to test for variances between oleoresin flow by aspect, since mountain pine beetle attack densities are commonly greatest on north aspects [
29]. Glass vials were placed into wounds and held in place with a band of packing tape around the tree. Oleoresin that flowed into the vials was left in vials, weighed, and stored frozen (−22 °C) prior to chemical analyses. Empty vial weights were subtracted from the total weights of vial and resin for overall oleoresin weights. Log10-transformations of oleoresin weights were used to reduce high leverage points. Differences in oleoresin flow amounts were tested in response to north and south aspects of both host species by using a two-way ANOVA (Proc GLM; SAS v9.2, SAS Institute Inc., Cary, NC, USA) and calculating half least significant difference (LSD) values for confidence bounds (Proc GLM; SAS 9.2). We found that oleoresin flows were not different between north and south aspects for either tree species (lodgepole: 0.87 mg (0.73, 1.05); mean (Lower half LSD, Upper half LSD); ponderosa: 0.99 mg (0.83, 1.2);
F3,183 = 0.52,
p = 0.47). Additionally, no differences in tree diameters were detected between the paired alternate host pine species used in the analyses (lodgepole pines 24.9 cm (24.1, 25.7); mean (lower half LSD, upper half LSD); ponderosa pines 25.5 cm (24.7, 26.2);
F1,102 = 0.63,
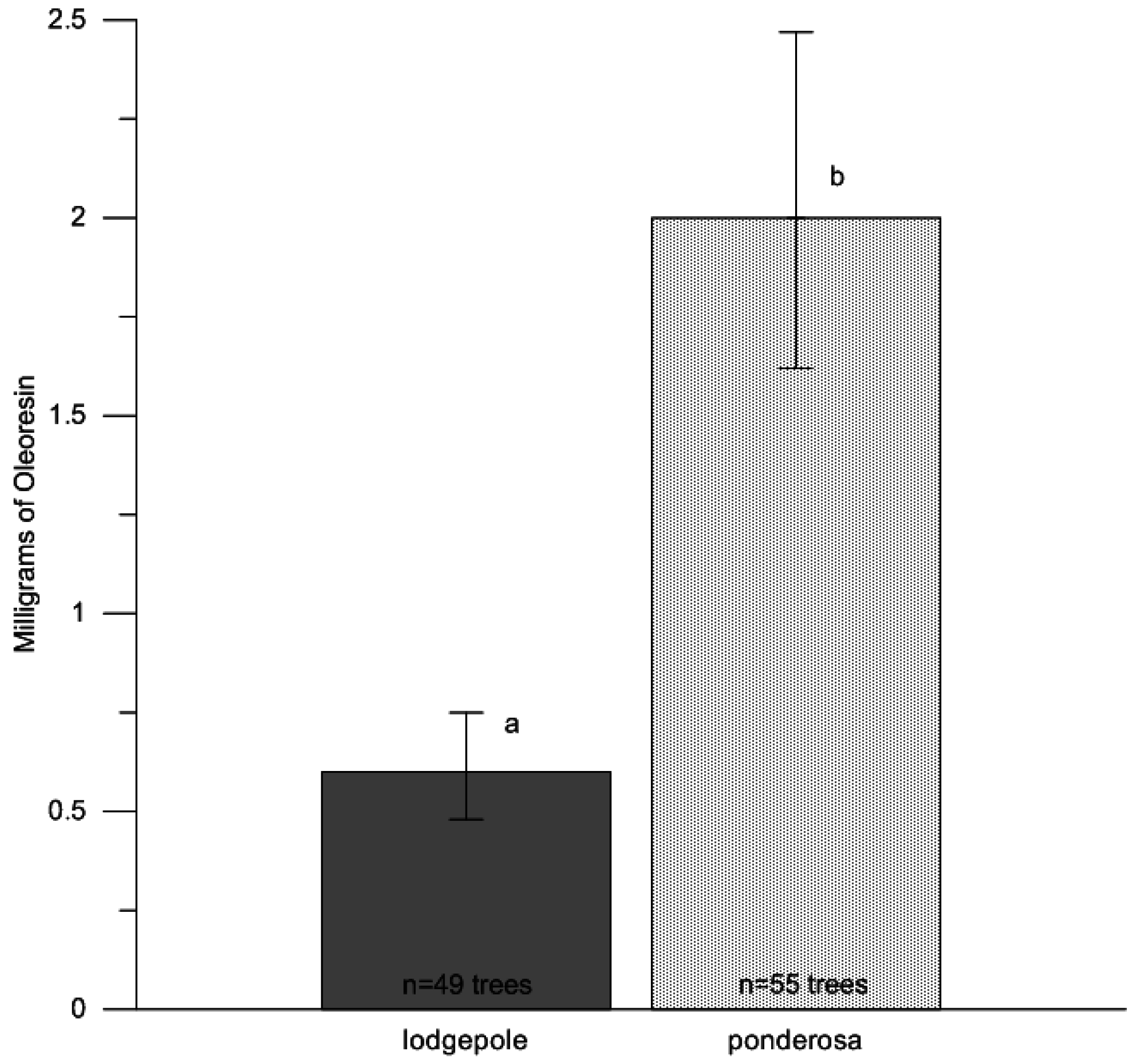
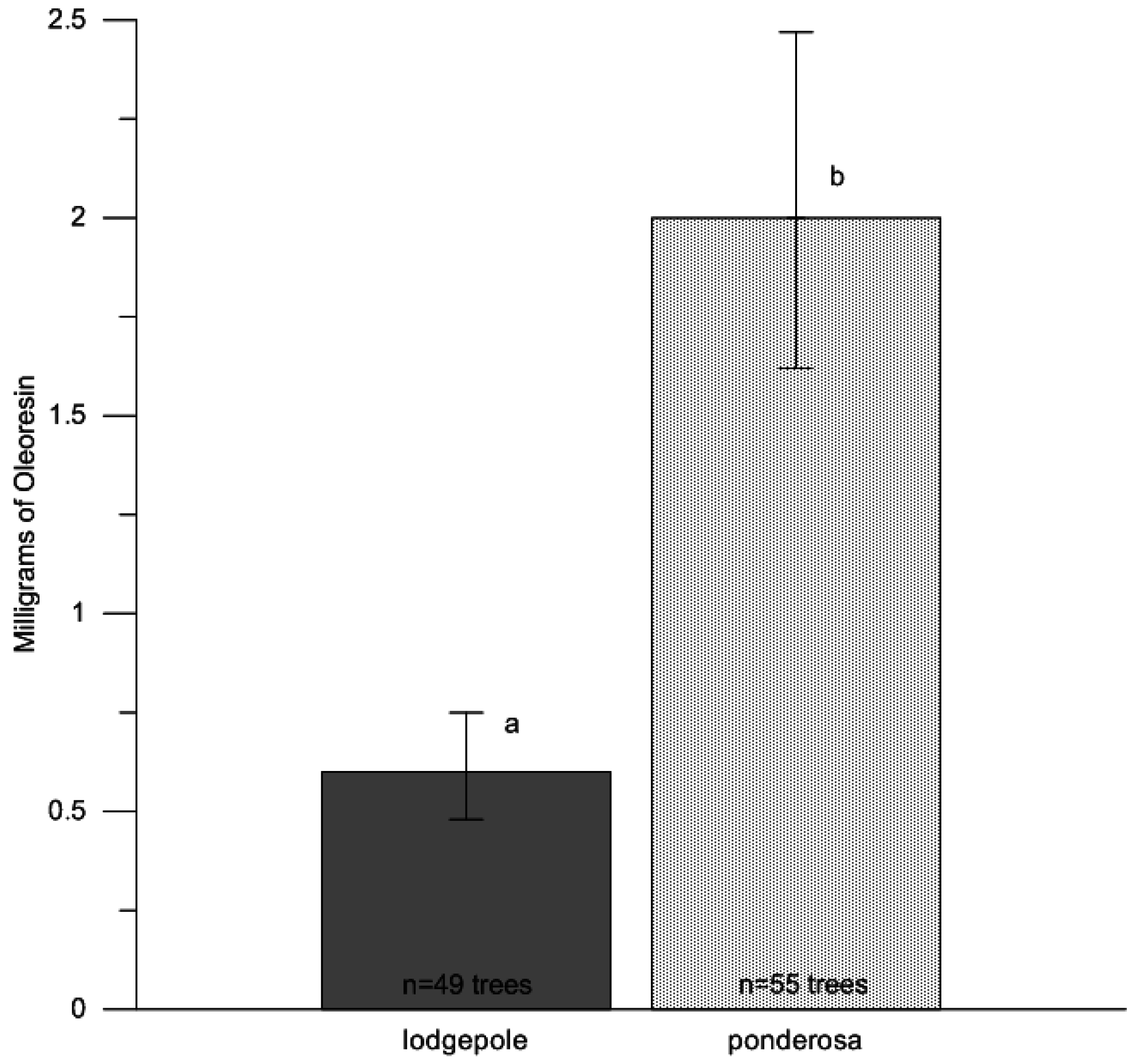
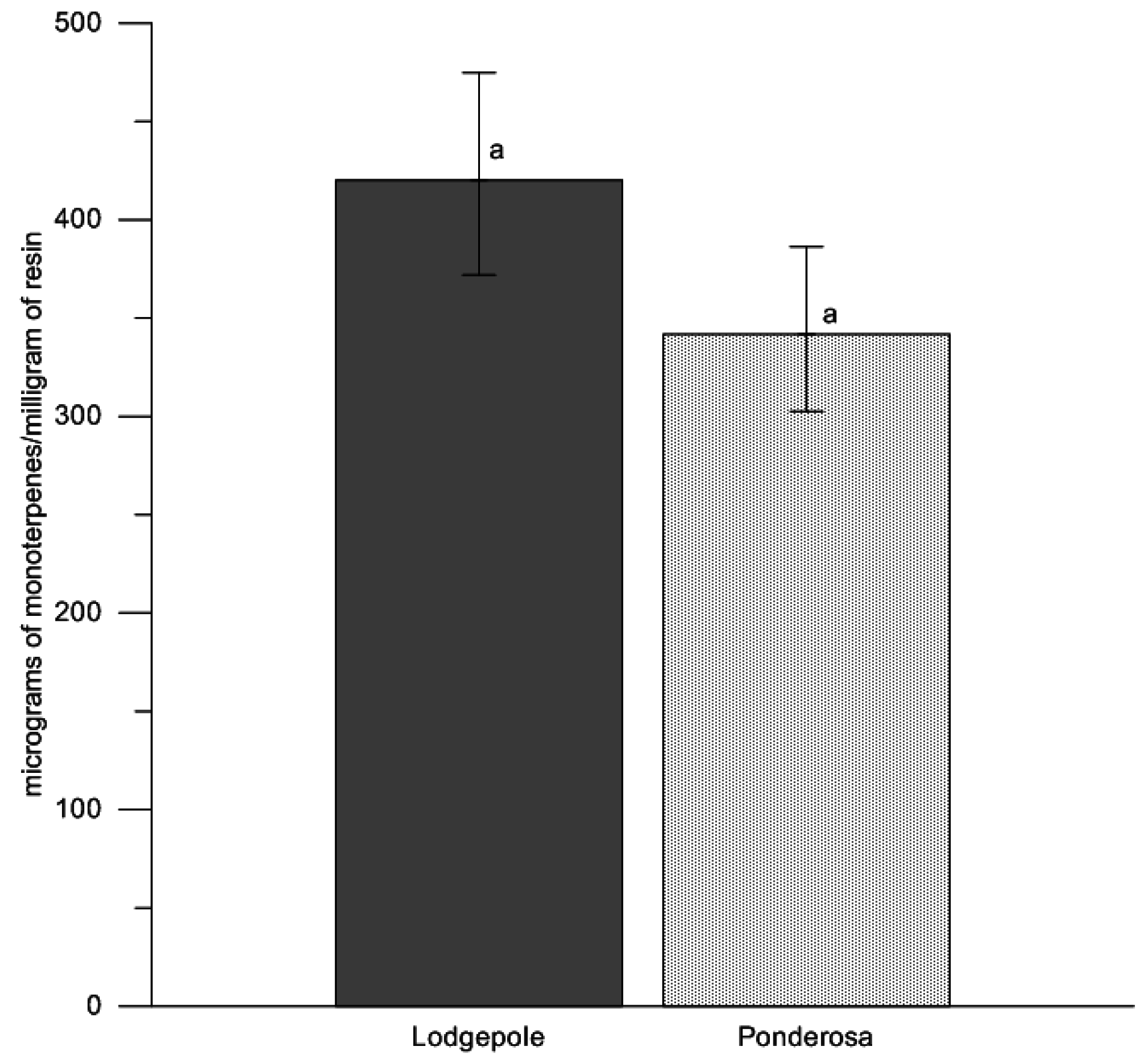
p = 0.42). To test for differences in oleoresin flow amount between host species, we averaged the oleoresin from the north and south aspects of each tree, using an analysis of variance treating paired resin amounts in response to tree species (Proc GLM; SAS v9.2, SAS Institute Inc., Cary, NC, USA). We further calculated half LSD values for confidence bounds. Mean estimates were back-transformed from log
10 values to improve interpretation of the results in
Figure 1. Upper and lower confidence bounds were calculated through log
10 back-transformed values (mean ±1/2 LSD). Eight samples leaked during collection and were not used in the analyses (7 collections from lodgepole pine trees, 1 from ponderosa pine), leaving a total of 104 samples.
2.3. Qualitative Defense Mechanisms
Terpenoid profiles from a random sample of oleoresin from 20 study trees (ten per species) were analyzed via gas chromatography coupled with flame ionization detection (GC-FID). We prepared a solution of 1 g of oleoresin/mL hexane. A 10 μL solution was transferred into a clean vial, and eicosane (100 μg in dichloromethane) was added as an internal standard. The sample was concentrated under a gentle stream of nitrogen until dry, and then re-dissolved in 1 mL of hexane. A 0.5 μL sample was injected onto a FAMEWAX column (30 m, 0.25 mm ID, 0.25 μm df, Cat. No. 12497, Restek, Bellefonte, PA, USA) on a HP 5890 gas chromatograph with a FID detector with temperature programming from 40 °C (held for 1 min) then increased by 5 °C per min to 200 °C (held for 2 min). Results were analyzed with Clarity® software (Clarity, 2008, DataApex Ltd., Prague, The Czech Republic) and quantified using a calibration curve created for synthetic standards of all compounds (Sigma-Aldrich, Inc., St. Paul, MN, USA) with the internal standard. β-phellandrene was identified on a HP 5890 Series II gas chromatograph coupled with a HP 5971 mass selective detector with temperature programming from 40 °C (held for 1 min) to 200 °C at a rate of 5 °C/min. The resulting mass spectra were analyzed visually and with a probability-based computer search of the NIST/EPA/NIH 75K spectral library (Hewlett-Packard, Palo Alto, CA, USA). Total and individual terpenoid concentrations were standardized based on per milligram of oleoresin.
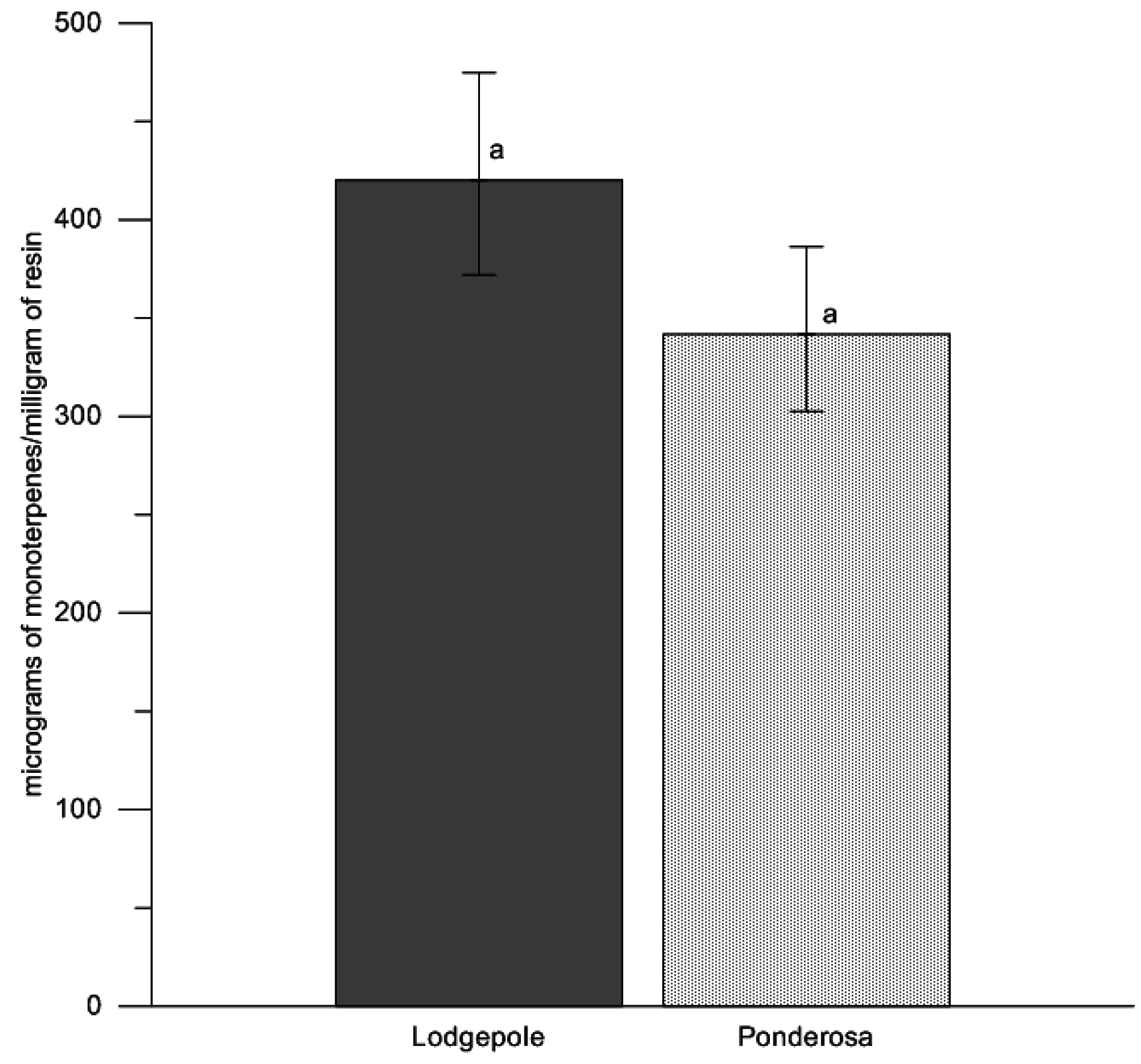
To investigate potential differences in the total cumulative concentrations of terpenoids within the oleoresin, we used a nonparametric, one factor model (host species: lodgepole pine, ponderosa pine; Proc NPAR1WAY, SAS v9.2, SAS Institute Inc., Cary, NC, USA) using Wilcoxon Exact tests, treating total monoterpene concentrations in response to tree species. The nonparametric tests were used to account for any skewness of any single terpenoid concentration.
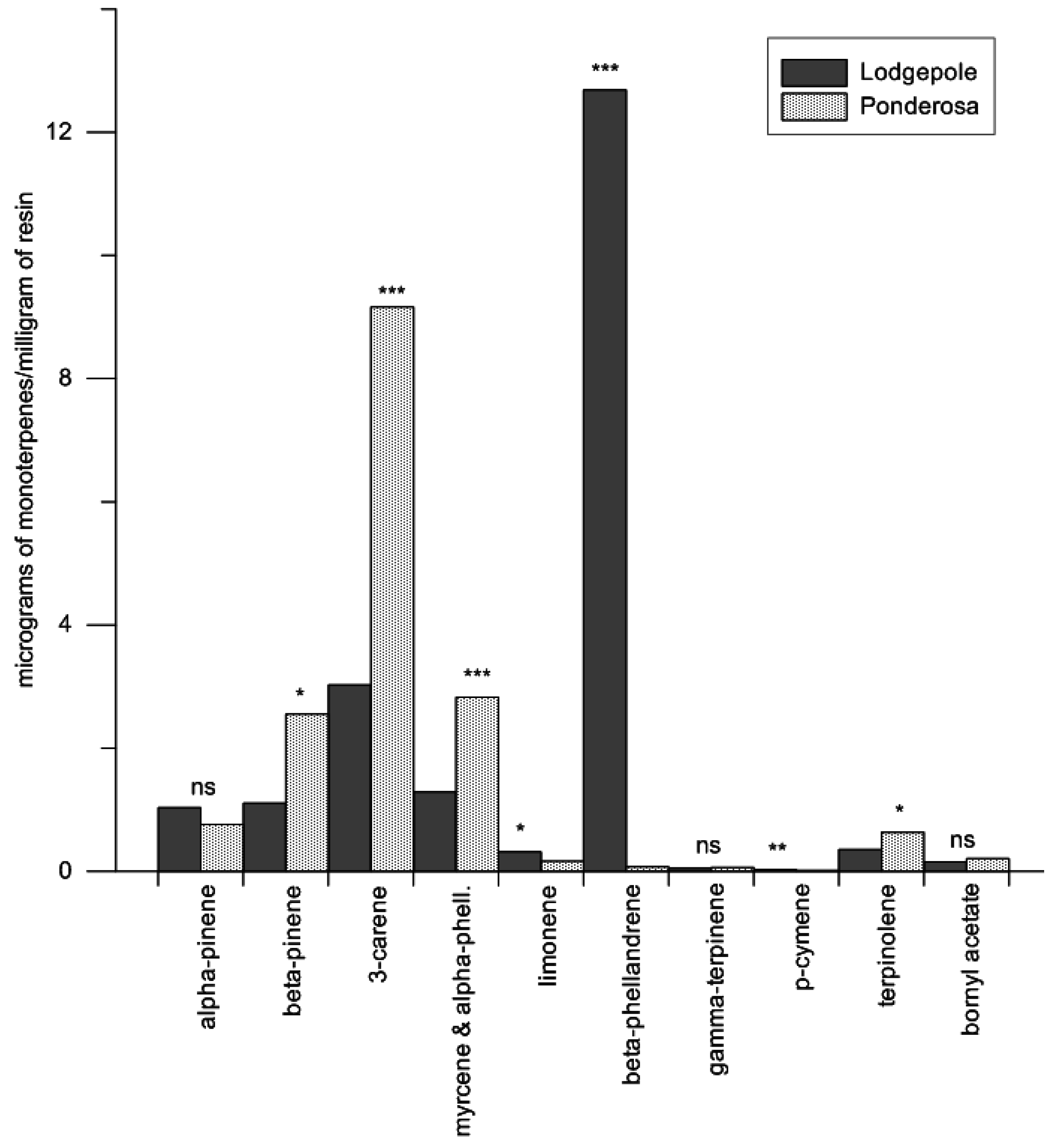
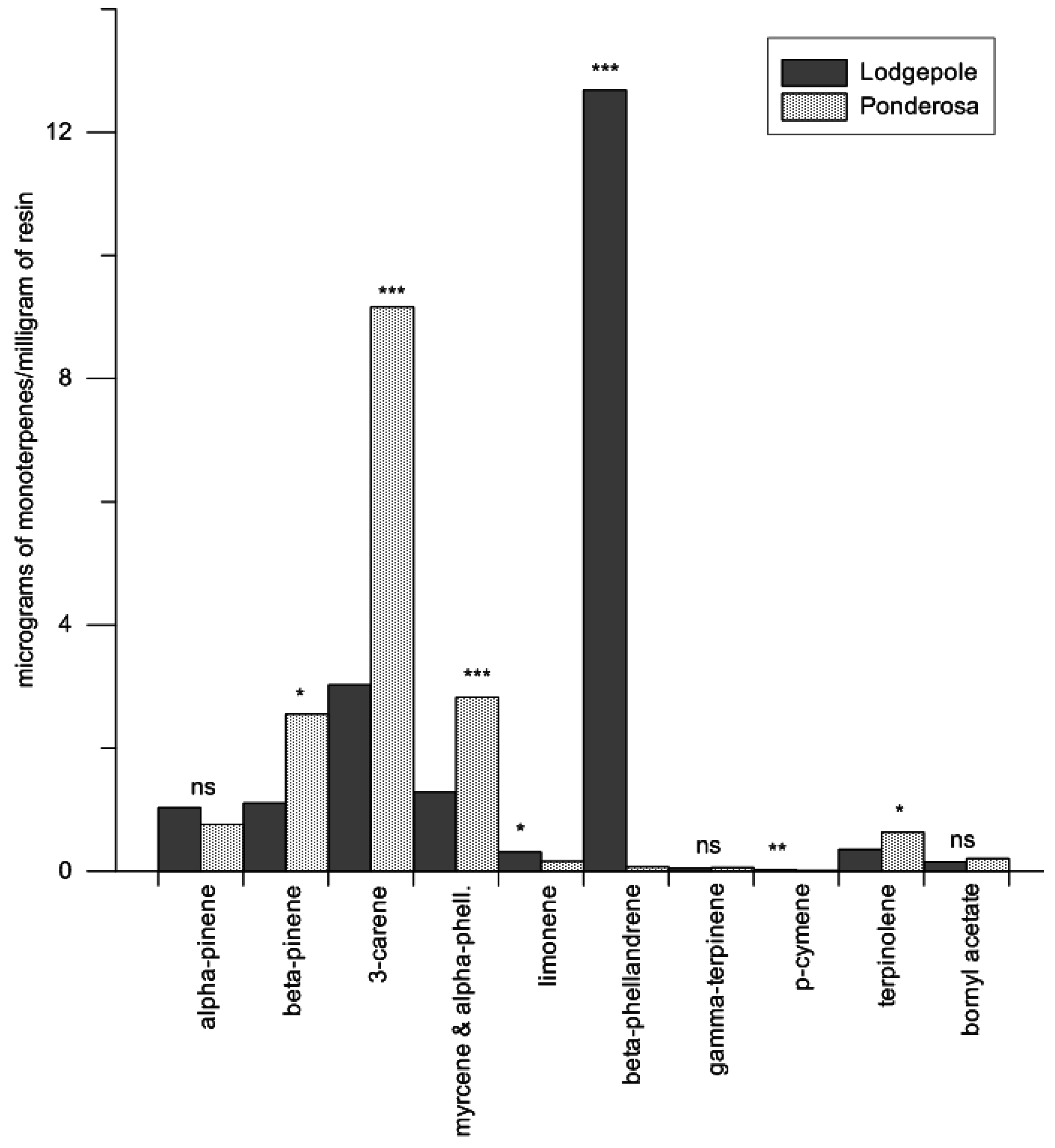
Specific terpenoid concentrations were analyzed between species using a multivariate ANOVA (Proc GLM; SAS v9.2, SAS Institute Inc., Cary, NC, USA ), treating eleven terpenoid concentrations in response to host species (α-pinene, β-pinene, myrcene, 3-carene, limonene, α-phellandrene, β-phellandrene, p-cymene, γ-terpinene, terpinolene, and bornyl acetate). Log10-transformations were performed on terpenoid concentrations for the multivariate ANOVA. Overall differences of terpenoid concentrations between species were detected in the model under Hotellings–Lawley trace tests; therefore, Tukey’s HSD adjustment for multiple comparisons was used to determine differences between the specific terpenoid mean concentrations between the species. Mean estimates were back-transformed to improve interpretation of the results, and confidence bounds were calculated as previously described (mean ± 1/2 LSD).
2.4. Chemicals
The following known-standard compounds and reagents used in the monoterpenoid analysis were purchased from Sigma-Aldrich, Inc., St. Paul, MN, USA: p-Cymene (99%, Cat. No. C121452), (−)-bornyl acetate (97%, Cat. No. B55203), γ-terpinene (97%, Cat. No. 223190), 3-carene (95%, Cat. No. 115576), (−)-α-pinene (99%, Cat. No. 305715), R(−)-β-pinene (99%, Cat. No. 112089), myrcene (tech. grade, Cat. No. M100005), and R(−)-α-phellandrene (>95%, Cat. No. 77429). Terpinolene was purchased from TCI America (>85%, Cat. No. T0817).
4. Discussion
The results from this study suggest that the quantitative defense of ponderosa pine is greater than that of lodgepole pine. The qualitative analyses also suggest that preformed oleoresin compositions vary markedly between the two pine species from the same ecotone stands, particularly those compounds known to be toxic to bark beetles (e.g., limonene) [
10] or those that have been shown to elicit mountain pine beetle behavior (e.g., β-phellandrene, myrcene, and terpinolene). West et al. [
27] reported that mountain pine beetle attacked ponderosa and lodgepole pines at equal rates in stands containing both species in the Southern Rockies. However, in both field-based choice experiments and laboratory-based bioassays where quantitative defenses were largely eliminated, the mountain pine beetle preferred ponderosa pine to lodgepole pine 2:1 [
28]. The results from this study found a greater amount of oleoresin exuded by ponderosa pine over lodgepole pine within 24 h of wounding, which may in part explain mountain pine beetle preference for ponderosa pine over lodgepole pine in manipulated choice experiments where oleoresin flow was excluded [
28,
30]. The results from this study are similar to other studies comparing defensive strategies in loblolly pine and shortleaf pine in the Southern US [
31]. Loblolly pines (
Pinus taeda L.) appear to rely more on quantitative defenses against southern pine beetle attack (
D. frontalis Zimmermann), while shortleaf pines may rely more heavily on qualitative defenses [
31]. Oleoresin viscosity may also play a role in
Pinus species defense strategy against attacking bark beetles [
32]. However, oleoresin viscosity was not examined in this study since the comparison of monoterpene concentration per milligram did not differ between ponderosa and lodgepole pine, which in theory governs the viscosity through chemical composition of monoterpenes within the oleoresins [
32].
When exogenous factors of temperature and precipitation cause drought stress in pines, the susceptibility to bark beetle attack of such trees may be enhanced due to reduced quantitative host defenses. Previous studies have found a close positive relationship between attack susceptibility and high drought stress [
33,
34,
35]. The density of attack necessary to overcome quantitative host defenses may be reduced when drought stress is considered [
7]. When quantitative host defenses were debilitated under experimental conditions, such as cut-log experiments and bioassays, the avoidance of defenses is likely less of a behavioral strategy of bark beetles [
28]. Further investigation of drought stress and beetle-caused tree mortality is warranted between lodgepole pine and ponderosa pine in Northern Colorado to fully understand host selection behavior of bark beetles and drought stress conditions; however, the effects of drought may be greater in ponderosa pine due to the greater amount of oleoresin produced 24 h after wounding compared with lodgepole pine. In this study, ponderosa pine produced approximately four times more oleoresins than lodgepole pine, which suggests terpene concentrations approximately four times greater (micrograms of monoterpenes/milligram of resin) for attacking beetles to contend with.
Total concentration of all monoterpenes (micrograms of monoterpenes/milligram of resin) were not different between the preformed oleoresins of lodgepole pine and ponderosa pine. This finding led to further investigations of the qualitative constituents within the oleoresins. The quantity of β-phellandrene from lodgepole pine was clearly a significant proportion of the overall monoterpene composition (61%). However, β-phellandrene has been shown to cause a positive flight response in the mountain pine beetle when coupled with an aggregation pheromone blend, though it did not significantly increase trap catches across a multitude of release rates [
36]. Since this compound elicits attraction of
Ips pini, and antennal responses in the mountain pine beetle [
37,
38], further investigation may be warranted to determine the importance of β-phellandrene to lodgepole pines relative to bark beetle defense.
We found preformed lodgepole pine oleoresins contained significantly more limonene (1.5% of total monoterpenes) than ponderosa pine (1.3%) oleoresins. It is unclear if these differences translate into greater chemical defense against attack in lodgepole pine. However, when comparing lodgepole with ponderosa pine, nearly four times more resin is exuded from ponderosa over lodgepole pine, yielding not only significantly more resin for attacking beetles to contend with, but also significantly more terpenes to contend with. Since limonene is highly toxic to bark beetles, we suspect the incremental difference may have an ecological effect on attacking beetles per milligram of resin [
13,
21,
23].
We did not find any differences in concentrations of the common beetle attractant α-pinene between lodgepole and ponderosa pine. α-pinene in a blend containing other monoterpenes can synergize mountain pine beetle attack. Thus, there is a need for further investigations to determine and clarify α-pinene’s role in conjunction with differences found between myrcene, α-phellandrene, and terpinolene regarding primary host selection behavior for these two pine species. This is of particular interest since other bark beetle species have a behavioral connection to specific concentrations of α-pinene [
39]. For example, spruce beetle (
Dendroctonus rufipennis Kirby) populations avoided high concentrations of α-pinene, the monoterpene of greatest concentration in spruce, though moderate concentrations of α-pinene signaled host acceptance and gallery initiation [
39]. Similar relationships have been reported with
Ips species and host acceptance when monoterpene concentrations fell within a range, while concentrations outside of the range for a given monoterpene elicited a lack of host acceptance or decrease in gallery elongation [
40]. Not only may individual host monoterpene presence and titer exert negative pressures on host selection, pine bark beetle aggregation pheromones are synergized by the presence of some host monoterpenoids. Seybold et al. [
9] reviewed the synergisms between pheromones and kairomones, and highlighted a need for further investigations in their complexity with regard to beetle host selection. Our study has identified an ecologically relevant range of α-pinene in both lodgepole pine and ponderosa pine, setting the stage for further investigations in behavioral correlations with the mountain pine beetle and other pine bark beetles.
The success of a bark beetle attack is linked to preformed host defenses [
29,
30] and those defenses induced by the attack [
12]. Gaylord et al. [
41] found ponderosa pine may have a weak to nonexistent inducible defense to bark beetle attack, while the findings of Raffa and Berryman [
13,
16] in lodgepole pine suggest a strong response to wounding. One shortcoming of this study is that we did not attempt to quantify the induced defenses, nor did we think that a comparative study of the response to wounding between the two hosts would be of interest to this or future epidemic situations.
Although we did not focus on myrcene and terpinolene in particular at the outset of this investigation, these monoterpenes can be attractive or elicit antennal responses in the mountain pine beetle [
37,
38,
42]. We found differences in terpinolene between the two host species. Myrcene and α-phellandrene co-eluted in our gas chromatograms, and no attempts to further separate these compounds were made, as our focus was on total monoterpene load, limonene, α-pinene, and β-phellandrene.


{kind=link}
{kind=link}
{kind=link}