1. Introduction
Loblolly pine (
Pinus taeda L.) is one of the most important commercial tree species in the world, and is the foundation of the forest products economy of the southeastern USA, which produces 16% of the world’s timber volume, more than any other country [
1]. As of 2010, loblolly pine plantations represented one-half of the standing pine volume in the southern USA (11 million hectares), with over 1 billion seedlings planted annually [
2]. Forest management requires a long-term perspective because decisions made today dictate outcomes decades into the future. Even with application of intensive silviculture to increase growth rates, rotation lengths of loblolly pine plantations in the southeastern USA are usually longer than 20 years [
3]. Significant increases in atmospheric CO
2 concentration are occurring and associated changes in temperature and rainfall may begin to occur over this time frame [
4]. To respond to projected changes in climate, a better understanding is needed of how climate affects loblolly pine plantation growth, productivity, and resilience. This information can be used to adapt forest management to novel climate conditions and help mitigate the rise in atmospheric greenhouse gases through increased carbon sequestration.
By the end of this century, an increase of 2.5 to 4.0 °C in mean annual temperature is predicted for the southeastern USA [
4,
5]. Vapor pressure deficit increases exponentially with temperature such that higher temperatures will increase evapotranspiration and potential for tree water stress [
6]. Even though total precipitation is predicted to increase slightly across the southeastern USA [
4], an increase in rainfall intensity is projected, which will increase runoff from the soil and increase the number of dry days [
5]. The combined effects of increased evapotranspiration and reduced infiltration is predicted to decrease soil moisture [
4] and increase the frequency and intensity of drought [
4,
5]. Because of the importance of soil water availability to productivity and ecosystem function, there is strong interest in understanding the effects of altered precipitation on global ecosystems. While extensive loblolly pine plantation research has focused on maximizing productivity through the addition of nutrients and control of competing vegetation (e.g., [
3,
7]), less is known regarding the potential impact of changes in climate, specifically drought stress, on loblolly pine productivity and how reduced soil moisture availability may interact with nutrient availability.
Genetic improvement and intensive silviculture (mechanical site preparation, competition control, fertilization) have increased loblolly pine plantation total yields from 91 to 420 m
3·ha
−1 since the 1950’s [
3], with maximum annual yields approaching 30 m
3·ha
−1·year
−1 [
8]. Extensive research efforts by academia, corporate landowners, and their cooperative partnerships provided much of the technical improvement that led to current pine plantation yields. Given the size of the actively managed loblolly pine plantation resource and its responsiveness to silviculture, management can be modified to increase regionwide ecosystem carbon sequestration, especially when gains in ecosystem carbon align with increased stem growth or efficiency of fertilizer use. For instance, an additional 8 Tg CO
2 equivalents year
−1 is sequestered due to application of N and P fertilizer to loblolly pine plantations [
9].
Considerable research has focused on the effects of adding limiting resources, such as nutrients and water, on the growth and ecophysiological processes in pine plantations in the southern USA (e.g., [
7]). Fertilization often increases loblolly pine plantation development and productivity, primarily by increasing leaf area development e.g., [
7]. The typical response to a one-time, mid rotation application of 135–180 kg·ha
−1·N and 22–45 kg·ha
−1·P is a 25% growth increase over an 8–10 year period [
10]. Between 485,000 and 600,000 ha of loblolly pine plantations are fertilized annually due to the large and consistent growth response to fertilization [
10].
In productivity research studies, irrigation has had a more variable and a generally smaller effect than fertilization on pine growth in the southeastern USA. Results of irrigation studies in loblolly pine generally indicated no response to water addition [
11,
12] or modest positive responses [
13,
14]. In all cases, the response to irrigation was less than the response to fertilizer application when fertilization and irrigation were measured in the same study. The loblolly pine response to irrigation may have been relatively small because previous studies were in humid areas with annual precipitation between 1110 and 1260 mm and annual net precipitation may have exceeded the stand’s transpiration potential [
15,
16,
17]. In contrast, a lack of water,
i.e., drought, has a large negative effect on stand growth. Loblolly pine trees are very sensitive to prolonged low soil water content (e.g., [
18]) and exhibit reduced diameter growth [
19] as well as decreased tree transpiration, gross ecosystem productivity, and net carbon exchange [
13] in response to drought.
Effects of decreased water availability on stand growth and leaf physiology may be more severe for fertilized stands because of increased stand and leaf-level transpiration that results from greater leaf area development due to nutrient additions. For example, in one of the few throughfall reduction studies with loblolly pine, the positive effects of fertilization on leaf area development were muted in the throughfall reduction treatment [
20]. Therefore, understanding the interaction between fertilization and decreased water availability is essential to sustaining loblolly pine growth, especially under a changing climate scenario. Studies are needed to test whether higher leaf area index from fertilization and associated increases in stand-level water use will exacerbate the potential negative effects of drought on leaf physiology, leaf area production and duration, carbon sequestration and carbon fluxes.
The objective of this paper is to describe a novel regional research project that measures the early growth response of loblolly pine plantations to throughfall reduction and fertilization at four locations spanning the commercial range. We hypothesized that fertilization alone would increase stand growth but that the benefits of fertilization would be absent when combined with throughfall reduction. We expected that throughfall reduction alone would reduce stand growth. Growth results from the first two years are provided along with descriptive site data. This research was carried out within the context of a regionwide, integrated research, education, and outreach project, known as PINEMAP (Pine Integrated Network: Education, Mitigation and Adaptation Project), involving over 100 scientists, graduate students, and research staff from eleven universities in the southeastern USA and the USDA Forest Service. The data generated from this research will be used to parameterize and validate climate-responsive forest growth models, and to inform outreach and education efforts focused on improving forest management under future, variable climate and management conditions.
2. Experimental Section
2.1. Study Sites and Experimental Design
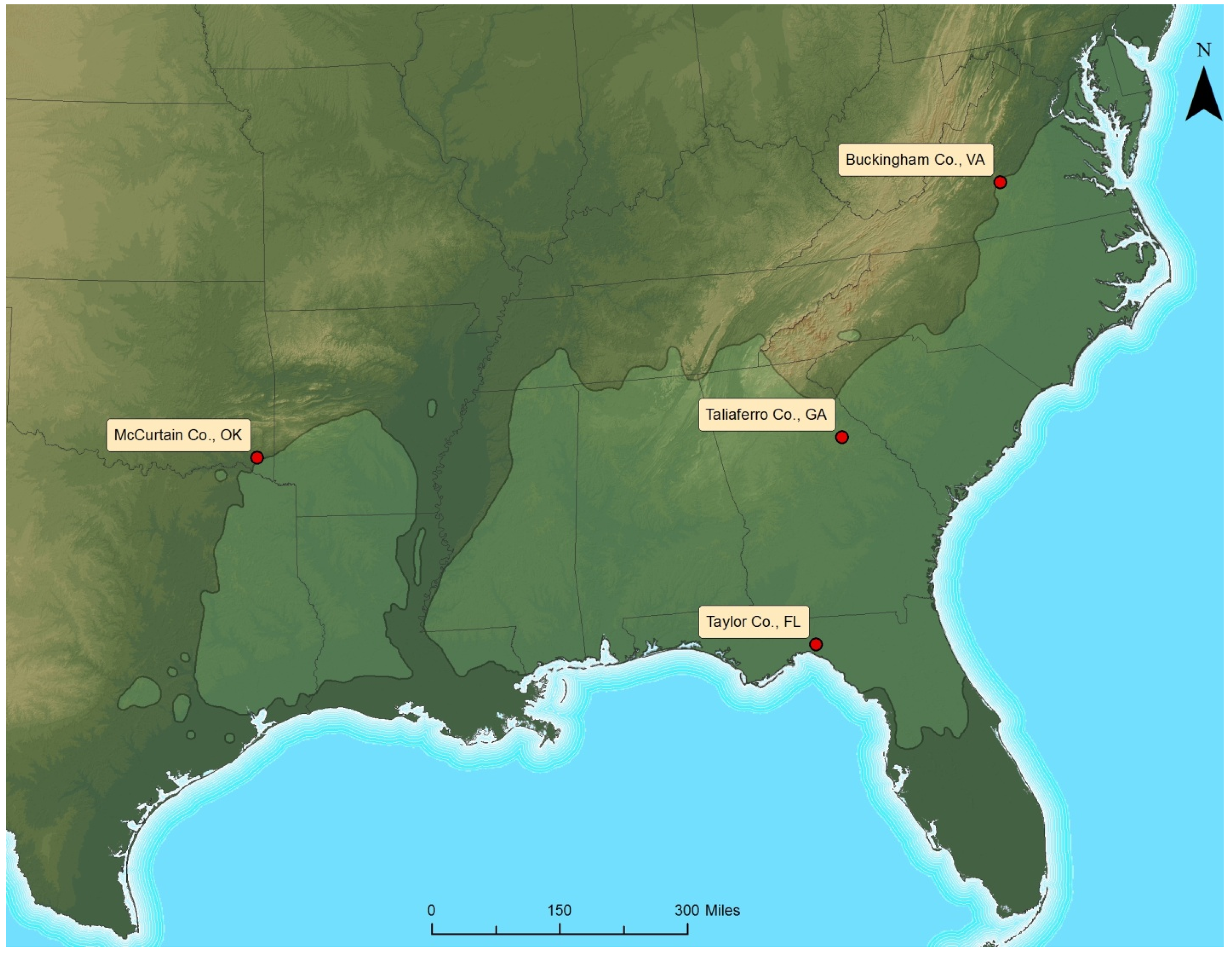
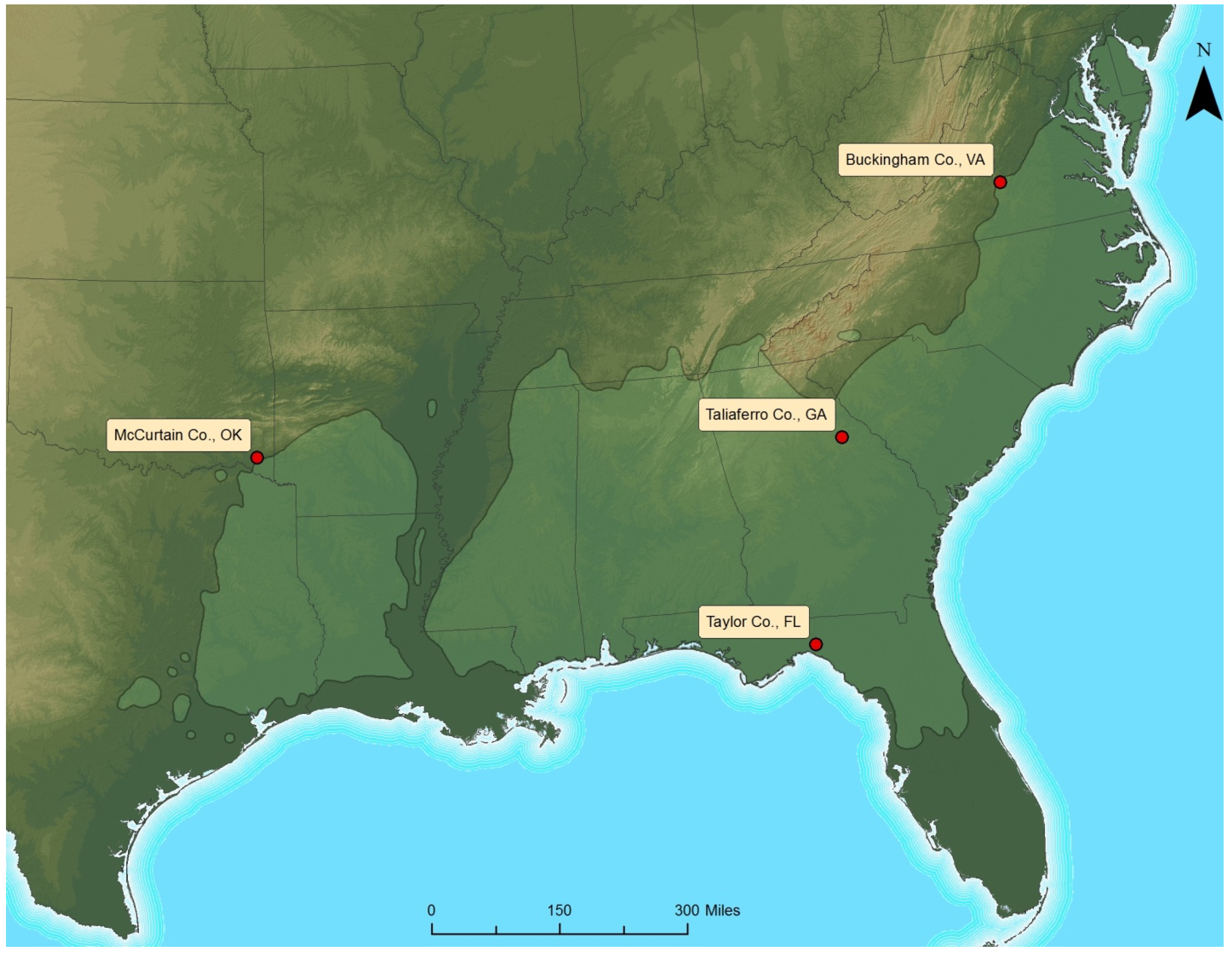
The long-term Throughfall Reduction x Fertilization (TRxF) study was installed during spring of 2012 at four locations across the commercial range of loblolly pine. The four TRxF sites were established in Virginia, Georgia, Florida, and Oklahoma to capture the range-wide variability of climate and productivity for this species (
Figure 1,
Table 1). Across the four sites, precipitation varied from 1120 cm per year at the Virginia site to 1450 mm per year at the Florida site. Likewise, annual temperature and January daily minimum temperature were lowest at the Virginia site and highest at the Florida site. However, the Virginia, Georgia, and Florida sites had similar August daily temperatures (within 1.0 °C), while the Oklahoma site was at least 2.6 °C warmer than the others. Soils ranged from well drained to somewhat poorly drained and have surface textures ranging from fine sand to clay loam (
Table 2). Across the four sites, initial soil chemistry varied, e.g., soil pH ranged from 4.22 to 5.53, soil carbon at 0–10 cm ranged from 13.3 to 37.4 (mg·g
−1), soil carbon at 10–20 cm ranged from 4.0 to 13.1 (mg·g
−1), soil nitrogen at 0–10 cm ranged from 0.73 to 1.30 (mg·g
−1), and soil nitrogen at 10–20 cm ranged from 0.40 to 0.55 (mg·g
−1) (
Table 3).
All plantations were operationally established between 2003 and 2008 at densities ranging from 1200 to 1800 trees·ha
−1. At the time of study initiation (2012), all plantations had not been thinned (densities ranged from 789 to 1720 trees·ha
−1,
Table 4), had not been fertilized, and were near or recently past crown closure. Average tree size at establishment ranged from 2.9 m tall and 3.6 cm·dbh (diameter at breast height) at the Oklahoma site to 10.5 m tall and 12.5 cm dbh at the Florida site (
Table 4). The Virginia site had the trees of larger diameter (14.5 cm) than those at the Florida site due to lower stand density. To expand inference of results to the broader regional genetic base, selected stands had been planted with a seed orchard mix of half-sib families representing local, genetically adapted sources.
Figure 1.
Map of the four throughfall reduction x fertilization study sites across the natural range of loblolly pine.
Figure 1.
Map of the four throughfall reduction x fertilization study sites across the natural range of loblolly pine.
Table 1.
Location and climate information for the four throughfall reduction x fertilization study sites established in non-thinned, mid rotation loblolly pine plantations.
Table 1.
Location and climate information for the four throughfall reduction x fertilization study sites established in non-thinned, mid rotation loblolly pine plantations.
| Site | Latitude | Longitude | Annual Prec. | Annual Temp. | Jan. Daily Min Temp. | Aug. Daily Max Temp. |
|---|
| | | | (mm) | (°C) | (°C) | (°C) |
|---|
| Virginia | 37°27′37″ N | 78°39′50″ W | 1120 | 13.6 | −4.4 | 30.6 |
| Georgia | 33°37′35″ N | 82°47′54″ W | 1220 | 16.1 | 0.0 | 31.6 |
| Florida | 30°12′22″ N | 83°52′12″ W | 1450 | 19.4 | 7.3 | 31.1 |
| Oklahoma | 34°01′47″ N | 94°49′23″ W | 1300 | 16.6 | −1.6 | 34.2 |
Table 2.
Soil description for the four throughfall reduction x fertilization study sites established in non-thinned, mid rotation loblolly pine plantations.
Table 2.
Soil description for the four throughfall reduction x fertilization study sites established in non-thinned, mid rotation loblolly pine plantations.
| Site | Physiographic Region | Dominant Soil Series | Soil Texture Surface | Soil Texture Subsoil | Drainage Class | Slope (%) |
|---|
| Virginia | Piedmont | Little Joe | Silt loam | Silty clay loam | Well drained | 0–15 |
| Georgia | Piedmont | Lloyd | Clay loam | Clay | Well drained | 0–3 |
| Florida | South. Coastal Plain | Melvina | Fine sand | Fine sand | Somewhat poorly drained | 0–2 |
| Oklahoma | Upper Coastal Plain | Ruston | Fine sandy loam | Clay loam | Well drained | 3–8 |
Table 3.
Soil pH and carbon and nitrogen concentrations for the four throughfall reduction x fertilization study sites established in non-thinned, mid rotation loblolly pine plantations.
Table 3.
Soil pH and carbon and nitrogen concentrations for the four throughfall reduction x fertilization study sites established in non-thinned, mid rotation loblolly pine plantations.
| Site | Soil pH 0–10 cm | Carbon 0–10 cm (mg·g−1) | Carbon 10–20 cm (mg·g−1) | Nitrogen 0–10 cm (mg·g−1) | Nitrogen 10–20 cm (mg·g−1) |
|---|
| Virginia | 4.22 | 37.4 | 9.6 | 1.30 | 0.40 |
| Georgia | 4.84 | 16.3 | 7.7 | 0.82 | 0.55 |
| Florida | 5.53 | 20.5 | 13.1 | 0.73 | 0.44 |
| Oklahoma | 4.65 | 13.3 | 4.0 | 0.80 | 0.40 |
Table 4.
Plot-level average stand density, tree height, tree diameter breast height (dbh), stand basal area (BA) and site index (SI) (base age 25) at time of treatment initiation (n = 16). Numbers in parentheses are the standard deviation.
Table 4.
Plot-level average stand density, tree height, tree diameter breast height (dbh), stand basal area (BA) and site index (SI) (base age 25) at time of treatment initiation (n = 16). Numbers in parentheses are the standard deviation.
| Site | Year Est. | Stand Density 2011 | Mean Height 2011 | Mean dbh 2011 | Mean BA 2011 | Mean SI 2011 |
|---|
| | | (trees·ha−1) | (m) | (cm) | (m2·ha−1) | (m) |
| Virginia | 2003 | 789 (68) | 8.79 (0.19) | 14.45 (0.52) | 13.20 (0.71) | 21.0 (0.3) |
| Georgia | 2006 | 1383 (63) | 6.34 (0.58) | 8.85 (0.93) | 8.79 (1.58) | 23.1 (1.9) |
| Florida | 2004 | 1720 (87) | 10.48 (0.52) | 12.53 (0.68) | 22.42 (2.19) | 25.9 (0.1) |
| Oklahoma | 2008 | 1610 (102) | 2.85 (0.14) | 3.55 (2.86) | 1.80 (0.30) | 20.2 (0.4) |
Treatments were established at each site consisting of a factorial combination of throughfall reduction (30% reduction) and fertilization with a complete suite of essential nutrients, replicated four times (16 plots per study site). Treatment plots were approximately square with a minimum 0.08 ha treatment area and an internal measurement plot size of at least 0.03 ha. Beginning at the time of treatment establishment, all competing vegetation was eliminated by manual clearing and directed spraying of glyphosate applied at labelled rates.
The throughfall reduction treatment, established during spring and summer 2012, consisted of covering approximately 30% of the plot area with troughs to capture and funnel throughfall away from the plots. Constructed below the tree canopies, the troughs were elevated above the soil surface between 0.5 and 1.5 m, depending on topography and the need to gravimetrically move water from the plots. Frames were constructed using pressure treated lumber attached to wooden stakes or metal T-posts. Each frame ran the length of the plot and was 1.5 to 1.3 m wide (depending on row spacing at the different sites). Along each frame were two 0.5 to 0.6 m wide troughs separated by 0.3 to 0.5 m wide open space (
Figure 2). Troughs were covered with clear 12 mil, UV stabilized polyethylene with an internal grid of high strength cord (polyscrim 12, Americover Inc., Escondido, CA, USA). The split-trough configuration, as well as elevation of the troughs above the ground, was employed to minimize banding in soil moisture. Collected throughfall was deposited on the ground a minimum of 3 m beyond the treatment plot. When topography and plot configuration might cause water movement among plots, collected throughfall was piped far enough away and down slope to prevent adding water to adjacent plots.
The elemental fertilization treatment consisted of 224 kg·ha−1 nitrogen and 27 kg·ha−1 of phosphorus and was representative of operational applications used with loblolly pine plantations (10). Nitrogen and phosphorus was applied as 432 kg·ha−1 urea and 140 kg·ha−1 diammonium phosphate. Agrotain Ultra (Koch Agronomic Services, LLC, Wichita, KS, USA) was included at a rate of 0.43 mL·kg−1 of urea to reduce nitrogen volatilization. In addition to nitrogen and phosphorus, elemental potassium was applied at a rate of 56 kg·ha−1 supplied as potash (112 kg·ha−1). To prevent micronutrient deficiency, granular oxysulfate micronutrient mix (Southeast Mix, Cameron Chemicals, Inc., Virginia Beach, VA, USA) was applied at a rate of 22.4 kg·ha−1. The mix contained 6% sulfur, 5% boron, 2% copper, 6% manganese, and 5% zinc. All fertilizers were broadcast applied by hand in March or April 2012.
Figure 2.
Recently installed (2012) throughfall excluders at the site in Taliaferro, Co., GA (a), McCurtain Co, OK, (b), Taylor Co., FL (c), and Buckingham Co., VA (d).
Figure 2.
Recently installed (2012) throughfall excluders at the site in Taliaferro, Co., GA (a), McCurtain Co, OK, (b), Taylor Co., FL (c), and Buckingham Co., VA (d).
2.2. Measurements
At all sites, dormant season measurements of tree heights and diameter at breast height (dbh) were taken before the 2012 growing season and then again after the 2012 and 2013 growing season. Stem volume outside-bark was calculated using the equation reported by Van Deausen
et al. [
22]. Foliar nitrogen and phosphorus concentrations were measured following the 2012 growing season to determine the effects of fertilizer additions. Samples were taken in mid-winter from the upper canopy on the south side from a minimum of five trees per plot and pooled.
Weather and environmental variables were monitored at all sites on a continuous basis. Variables included air temperature, relative humidity, photosynthetically active radiation, soil moisture, soil temperature, and precipitation. Volumetric soil water content was monitored between 0 and 12 cm in all replications and at ~30, 60, and 90 cm in the center of each plot in one or two blocks per site using time domain reflectometers (TDR) (CWS655 soil water reflectometer, Campbell Scientific, Logan, UT, USA).
The main effects (site, fertilization, throughfall reduction) and their interactions on periodic annual increment (first two years of treatment) were tested using the mixed procedure in SAS 9.3 (Proc Mixed, SAS Inc., Cary, NC, USA) with block as a random variable. All variables were normally distributed with equal variances and no transformations were needed.
3. Results and Discussion
The throughfall diverter systems performed well. During rainstorm events the troughs transferred all captured water off plots with minimal to no leakage, and the diversion structures were robust to heavy rain, solar radiation, and a 30 cm snowfall (in Oklahoma). Based on these observations, total plot throughfall volume reduction was presumed equal to trough coverage (
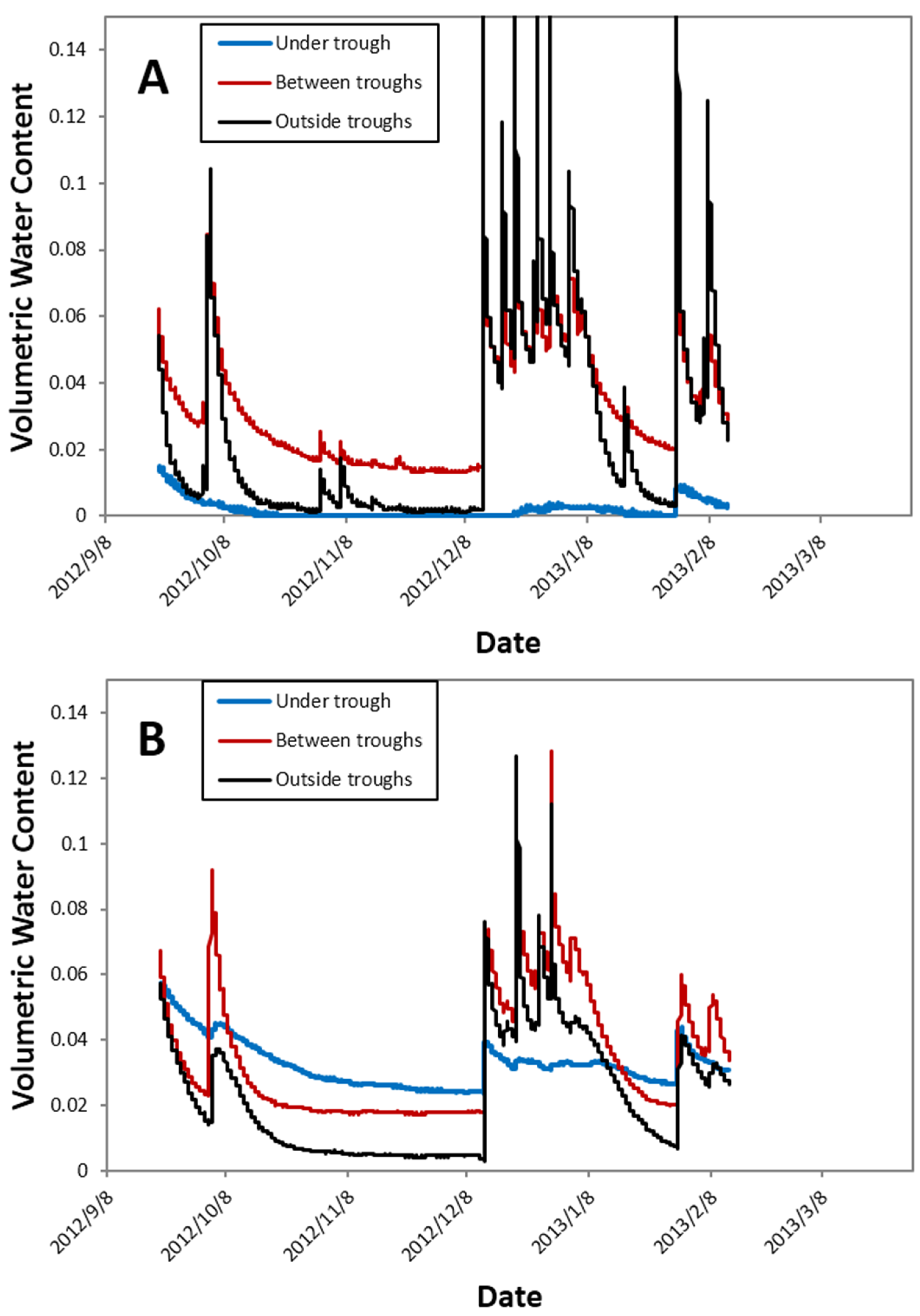
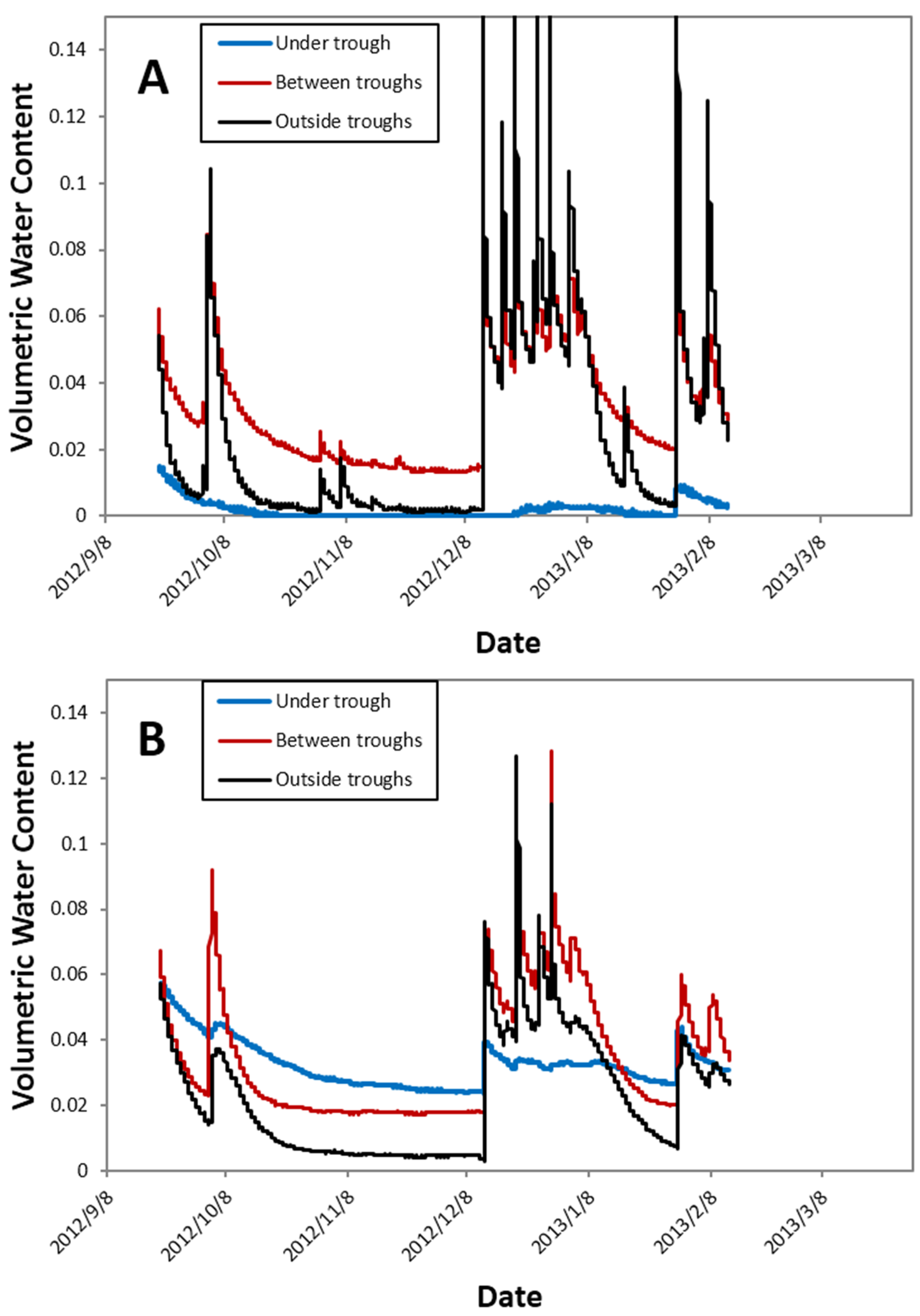
i.e., approximately 30%). When periodic measurements from the 2013 growing season were averaged, the throughfall reduction treatments reduced soil volumetric water content in the 0–12 cm layer from 13.3 to 9.4% at Virginia, 24.2 to 19.8% at Georgia, 6.6 to 4.9% at Florida, and 10.8 to 8.5% at Oklahoma. However, the throughfall diversion structures did introduce some spatial variability in soil moisture, especially in the surface 0–12 cm soil depth measured in Florida (
Figure 3A), where the soil directly under the troughs remained quite dry, while regions underneath the central gap between troughs and areas just outside the trough had wet-up and dry-down patterns more similar to ambient soil (data not shown). However, at deeper levels, these patterns were muted, and there was less pronounced variation relative to position below the troughs (
Figure 3B).
The fertilization treatment increased nutrient availability as assessed by foliar nutrient concentrations. When measured during the dormant season following fertilization, foliar nitrogen concentrations were higher at all four sites. Foliar phosphorus was higher due to fertilization at the Virginia, Georgia, and Oklahoma site, but not at the Florida site (
Table 5). Critical levels of foliar N and P in loblolly pine are generally considered to be 1.2 and 0.11% respectively [
23]. In regards to foliar N, all sites except the Virginia site had initial concentrations exceeding the critical level. While the critical level for nitrogen is useful as an initial assessment, fertilization response to N is associated with leaf area development and stands with projected LAI <3.5 respond to fertilization [
10] such that the fertilization response measured across the sites should contribute to accelerated growth. Fertilization with P increased foliar levels above the critical concentration at Virginia and Georgia. At the Florida site, the initial levels were at the critical concentration. The Oklahoma site remained slightly below the critical level one year after fertilization indicating growth potential might be limited by nutrition even in the fertilized treatment one year after application.
Figure 3.
Temporal variation in volumetric water content at 0–12 cm (A) and 12–20 cm (B) depths at three locations relative to a throughfall diversion structure at the Taylor County, Florida site.
Figure 3.
Temporal variation in volumetric water content at 0–12 cm (A) and 12–20 cm (B) depths at three locations relative to a throughfall diversion structure at the Taylor County, Florida site.
Table 5.
Dormant season measurements of foliar nitrogen (N) and phosphorus (P) concentrations one year after treatment initiation. TR = throughfall reduction treatment, F = Fertilization treatment. Numbers in parentheses are standard errors.
Table 5.
Dormant season measurements of foliar nitrogen (N) and phosphorus (P) concentrations one year after treatment initiation. TR = throughfall reduction treatment, F = Fertilization treatment. Numbers in parentheses are standard errors.
| | | 2012 Foliar N | 2012 Foliar P |
|---|
| Site | Treatment | (%) | (%) |
| | Control | 1.18 (0.01) | 0.092 (0.005) |
| Virginia | TR | 1.18 (0.02) | 0.094 (0.006) |
| | F | 1.68 (0.02) | 0.127 (0.006) |
| | TR + F | 1.71 (0.06) | 0.107 (0.002) |
| | Control | 1.45 (0.10) | 0.095 (0.004) |
| Georgia | TR | 1.31 (0.06) | 0.098 (0.004) |
| | F | 1.70 (0.03) | 0.123 (0.006) |
| | TR + F | 1.61 (0.01) | 0.104 (0.003) |
| | Control | 1.35 (0.01) | 0.110 (0.000) |
| Florida | TR | 1.40 (0.05) | 0.113 (0.005) |
| | F | 1.64 (0.02) | 0.113 (0.003) |
| | TR + F | 1.52 (0.01) | 0.105 (0.003) |
| | Control | 1.29 (0.03) | 0.085 (0.004) |
| Oklahoma | TR | 1.28 (0.01) | 0.082 (0.002) |
| | F | 1.48 (0.03) | 0.096 (0.003) |
| | TR + F | 1.46 (0.03) | 0.094 (0.005) |
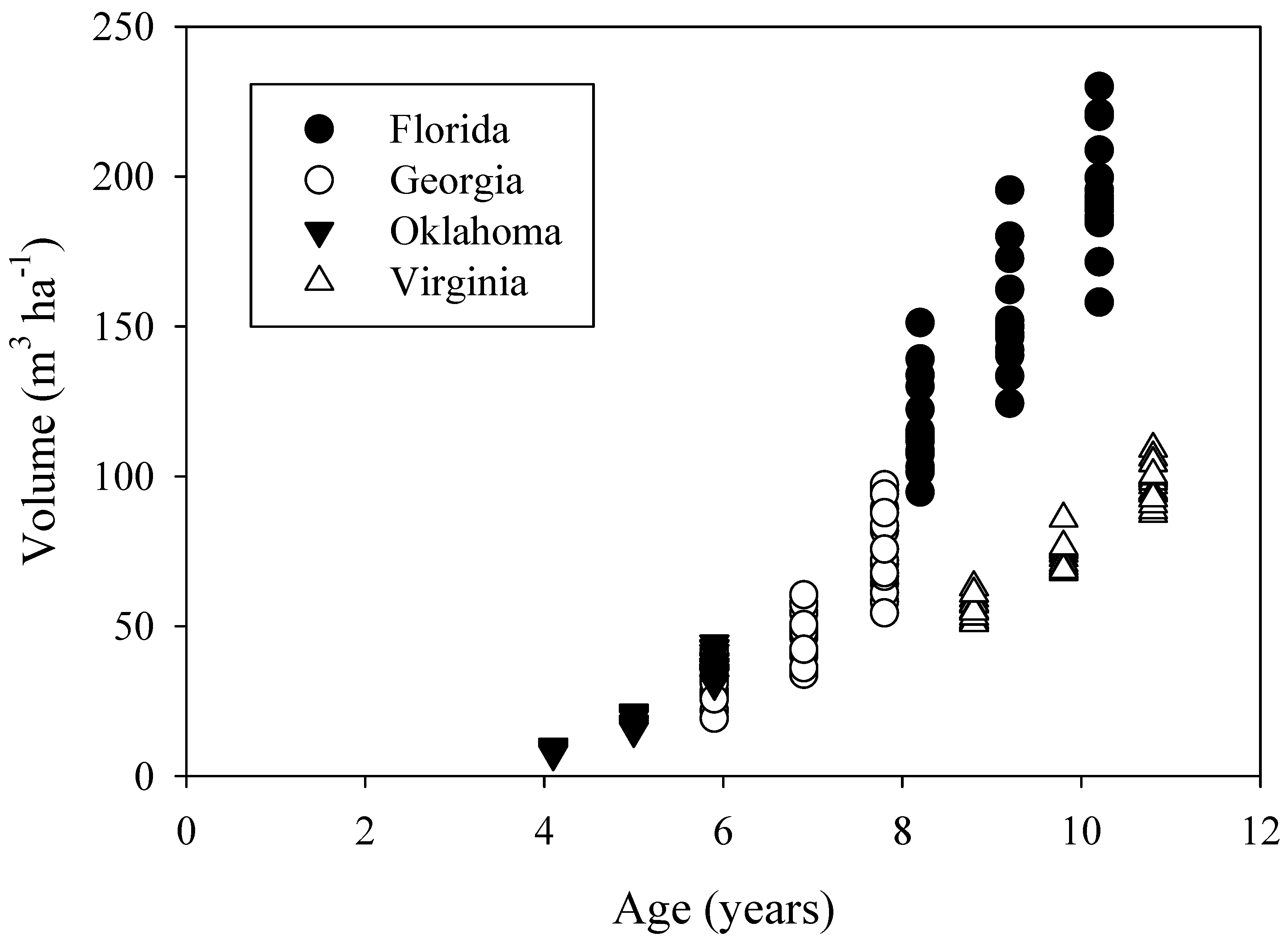
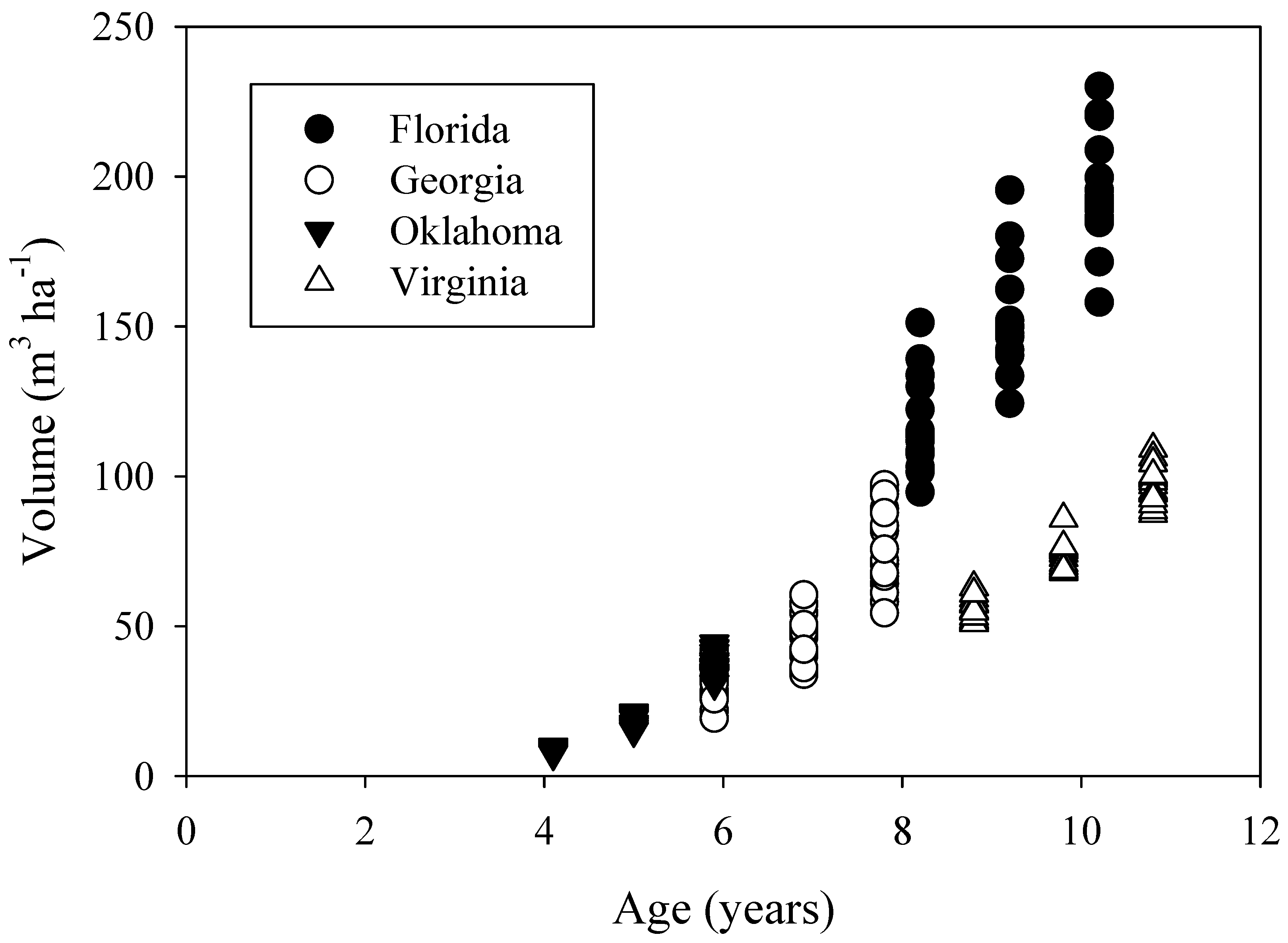
Stands were at different stages of stem volume development (
Figure 4) which is expected given the variation in age, soils, and climate among the study sites. However, the Oklahoma, Georgia, and Florida study sites, which span the loblolly pine commercial range from west to east, had similar growth trajectories. The Virginia site had a lower volume trajectory perhaps because the site was the coolest and driest (
Table 3). While standing volume at the Virginia site may be lower in part due to lower density, tree heights at the time of treatment were 1.7 m shorter at Virginia than the Florida site trees, even though the Virginia site was planted a year earlier. Accordingly, site index of the Virginia site was lower than the Georgia and Florida site, but similar to the Oklahoma site (
Table 4). A higher volume trajectory at the Oklahoma site than the Virginia site may be related to higher planting density, the use of genetic seed sources from the Western Gulf region for the Oklahoma planting, more advanced genetics (5 year time gap between plantings), or more intensive silviculture. A regionwide simulation of loblolly pine net canopy assimilation found that carbon gain was positively correlated with growing season precipitation and negatively correlated with vapor pressure deficit [
24]. However, leaf area index had the largest influence. Comparing the similarly aged Florida and Virginia plantations, leaf area index was approximately 45% lower at the Virginia site (data not shown).
The response of periodic annual increment (2-year stand volume) to fertilization and throughfall reduction varied by site (site x fertilization,
p = 0.01; site x throughfall reduction,
p = 0.02). Fertilization (F and TR+F treatments) significantly increased volume growth compared to nonfertilized treatments (C and TR) at all sites (
p < 0.01), but the size of the increase varied (increase of 1.6, 2.6, 4.8, and 6.7 m
3·ha
−1·year
−1 for Oklahoma, Virginia, Florida, and Georgia sites, respectively) (
Table 6). Throughfall exclusion (TR and TR+F treatments) reduced volume increment compared to the nonreduced plots (C and F) at the Oklahoma site by 2.6 m
3 ha
−1·year
−1 (
p = 0.0001) and at the Georgia site by 3.0 m
3 ha
−1·year
−1 (
p = 0.09), but did not significantly affect volume increment at the Virginia (
p = 0.61) or Florida sites (
p = 0.57) (
Table 6). The effects of throughfall reduction and fertilization did not interact when assessed across all sites (site x fertilization x throughfall reduction,
p = 0.33).
Figure 4.
Stand volume measured before treatment initiation and following two growing seasons.
Figure 4.
Stand volume measured before treatment initiation and following two growing seasons.
Table 6.
Periodic annual increment in stem volume outside-bark after 2 years (m3·ha−1·year−1) for throughfall reduction (TR) and fertilization (F) treatment (n = 4). Numbers in parentheses are standard errors.
Table 6.
Periodic annual increment in stem volume outside-bark after 2 years (m3·ha−1·year−1) for throughfall reduction (TR) and fertilization (F) treatment (n = 4). Numbers in parentheses are standard errors.
| Treatment | Virginia | Georgia | Florida | Oklahoma |
|---|
| C | 19.2 (0.5) | 21.8 (1.7) | 36.0 (1.7) | 15.4 (0.7) |
| TR | 19.4 (0.4) | 18.9 (1.5) | 37.0 (1.4) | 14.2 (0.6) |
| F | 21.6 (1.1) | 28.6 (2.2) | 42.4 (0.5) | 18.5 (0.4) |
| TR + F | 22.2 (1.1) | 25.5 (1.0) | 40.1 (0.3) | 14.4 (0.3) |
Fertilization with nitrogen and phosphorus at the amounts applied in this study has been previously shown to increase loblolly pine growth of intermediate aged stands by an average of 3.8 m
3·ha
−1·year
−1 for the first 8 years after treatment [
10]. Averaged across the four sites, the increase over the first two years in this regional experiment averaged 3.9 m
3·ha
−1·year
−1. The relatively smaller response at the Oklahoma site could be due to the younger age of the stands. The smaller response at the Virginia site could be due to a lag in growth acceleration for the slower growing stands at this site. The Virginia site had the lowest initial foliar N concentration and the largest positive response in foliar nitrogen concentration to fertilization, which implies that the Virginia site was strongly nutrient limited.
An approximate 30% reduction in throughfall did not affect volume growth at the Florida site, perhaps because of ample precipitation (1069 and 1008 mm for 2012 and 2013) or a higher water table and slower soil drainage class (somewhat poorly drained), which may have increased water availability for all treatments. At the Virginia site, the lack of response to throughfall reduction could also be due to ample precipitation, (940 and 1331 mm in 2012 and 2013, respectively, compared to an annual average of 1120 mm). Even though the Oklahoma site received 1031 and 1450 mm of precipitation in 2012 and 2013, respectively, the large response to throughfall reduction could be related to summer temperatures that were >1.6 °C warmer than the other sites (average high temperature for June–September was 33.8 and 32.1 °C in 2012 and 2013) and associated higher vapor pressure deficits. In addition, western seed sources of loblolly pine may exhibit a greater reduction in transpiration when grown under drought conditions [
25], which could have exacerbated the growth reduction in the throughfall reduction treatments. Even though precipitation was above average in 2013 at the Georgia site (1410 mm), the site’s relatively large decrease in volume growth associated with throughfall reduction could have been due to the drought conditions during 2012 (828 mm of precipitation compared to annual average of 1220 mm) which likely exacerbated the effects of throughfall reduction. The effects of fertilization and throughfall reduction will be monitored for at least three additional growing seasons.
4. Conclusions
At two of the four sites throughfall reduction reduced growth, and at the other two sites, the effect was not significant. This result indicates that the effects of a drier climate on loblolly pine plantation growth may be modest in some regions, with effect size varying with location, regional weather patterns, edaphic features, and water table depth. As we gather additional years of data, our measurements will include responses to a greater range and variability in annual precipitation and weather conditions across the four sites. Combined with the ecophysiological measurements, we hope to expand our inference regarding a drier climate more broadly across the commercial range of loblolly pine and understand under which conditions growth will be most affected.
The additive effects of treatments indicate that when throughfall reduction does reduce growth, the positive effects of fertilization are not eliminated. The net effect on growth depended on the difference between the two responses and determined whether volume growth decreased or increased when the two were combined. At the Oklahoma site, the response to the throughfall reduction treatment was greater than the fertilization effect, such that drier conditions had a larger negative effect than the positive response to increased nutrient availability. In contrast, the fertilization effects were greater than throughfall reduction effects at the Georgia site. The biological and edaphic processes underlying these responses are being measured as part of this larger research effort and will be used to parameterize and refine models to predict growth responses under future climate and management scenarios.
Climate change may result in novel environmental conditions across the southeastern USA. This unique study, replicated across the native range of loblolly pine, ultimately will provide a region-wide mechanistic understanding of loblolly pine plantation response to rainfall reduction and its interaction with nutrient availability, which will greatly improve our ability to predict future impacts of reduced soil moisture caused by climate change. The data from the TRxF study increase our understanding of how loblolly pine plantations grow and function under a drier climate scenario so that we can better predict growth, assess risk, and ultimately alter management to sustain or increase productivity.




 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}