
Short-Term Response of Native Flora to the Removal of Non-Native Shrubs in Mixed-Hardwood Forests of Indiana, USA

Abstract
:1. Introduction
2. Materials and Methods
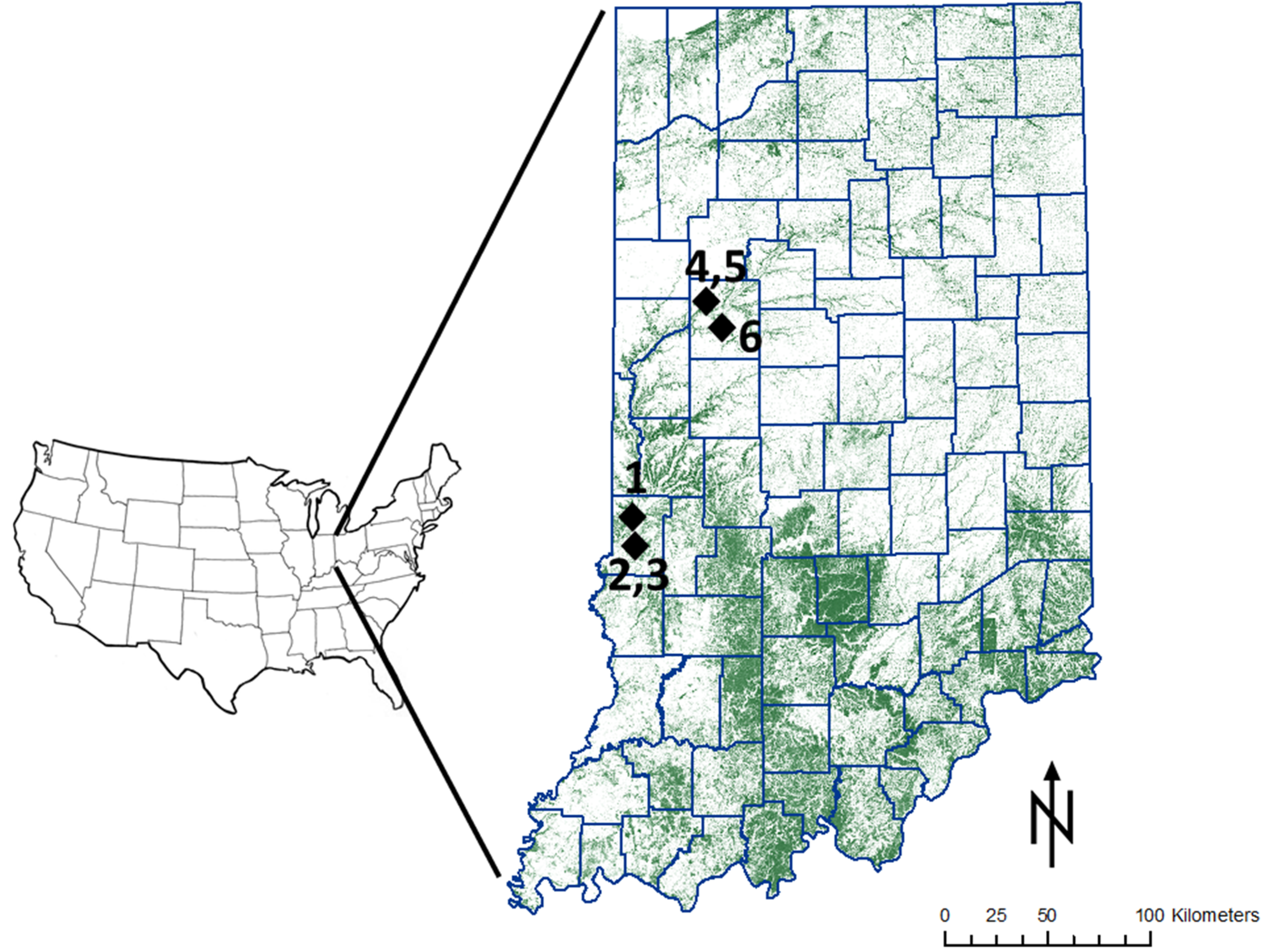
2.1. Study Area
2.2. Experimental Design

2.3. Data Collection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
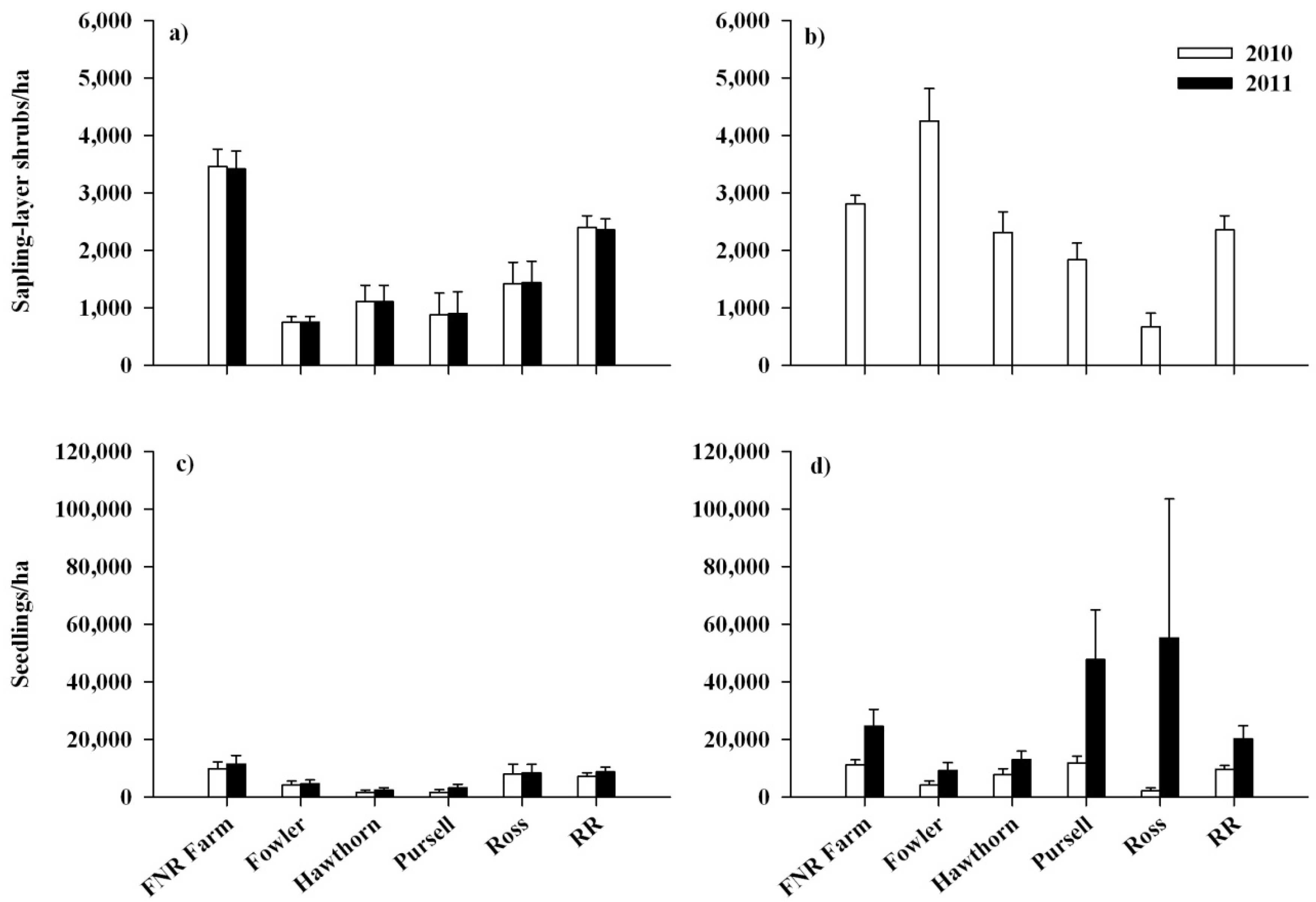
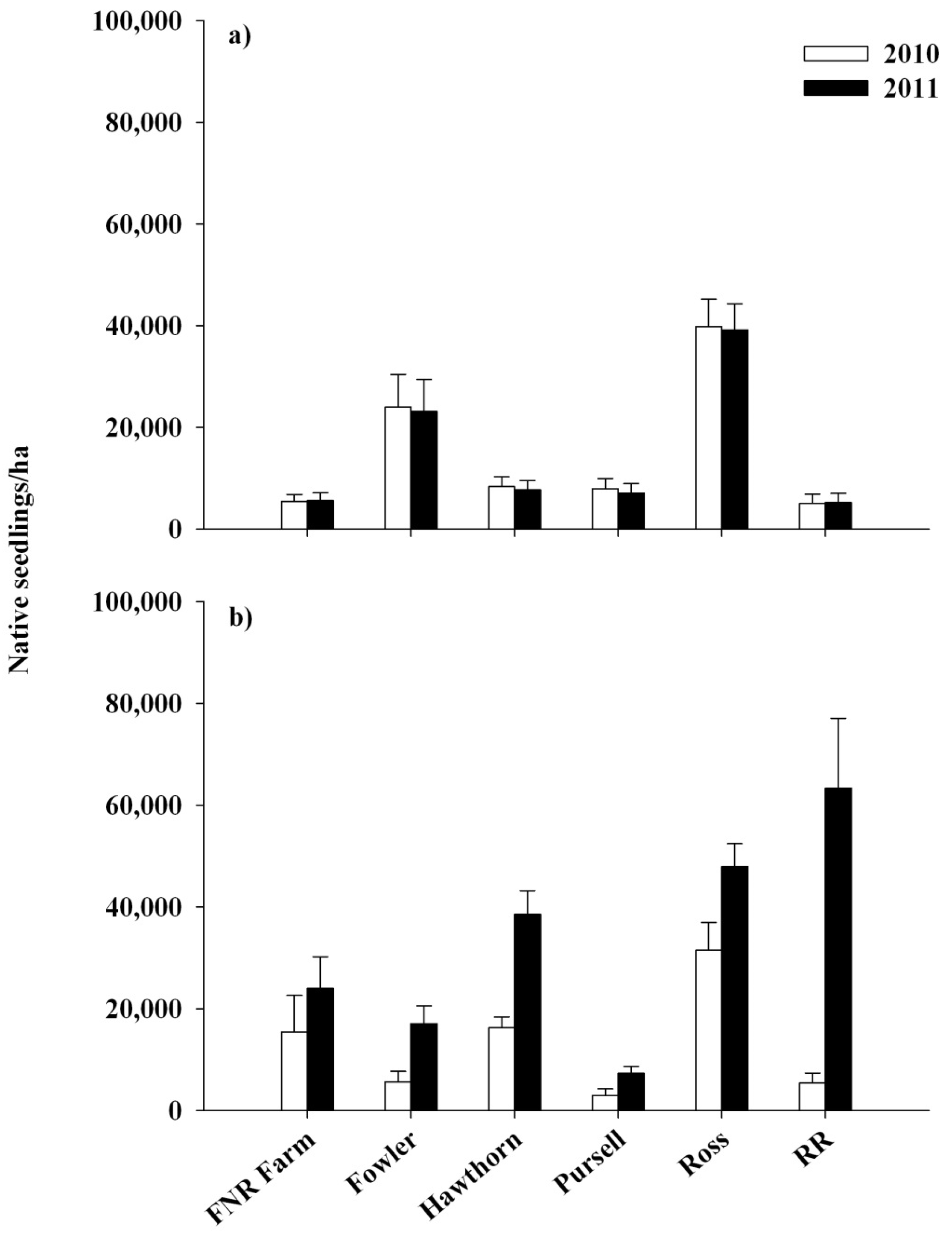{kind=link}
{kind=link}
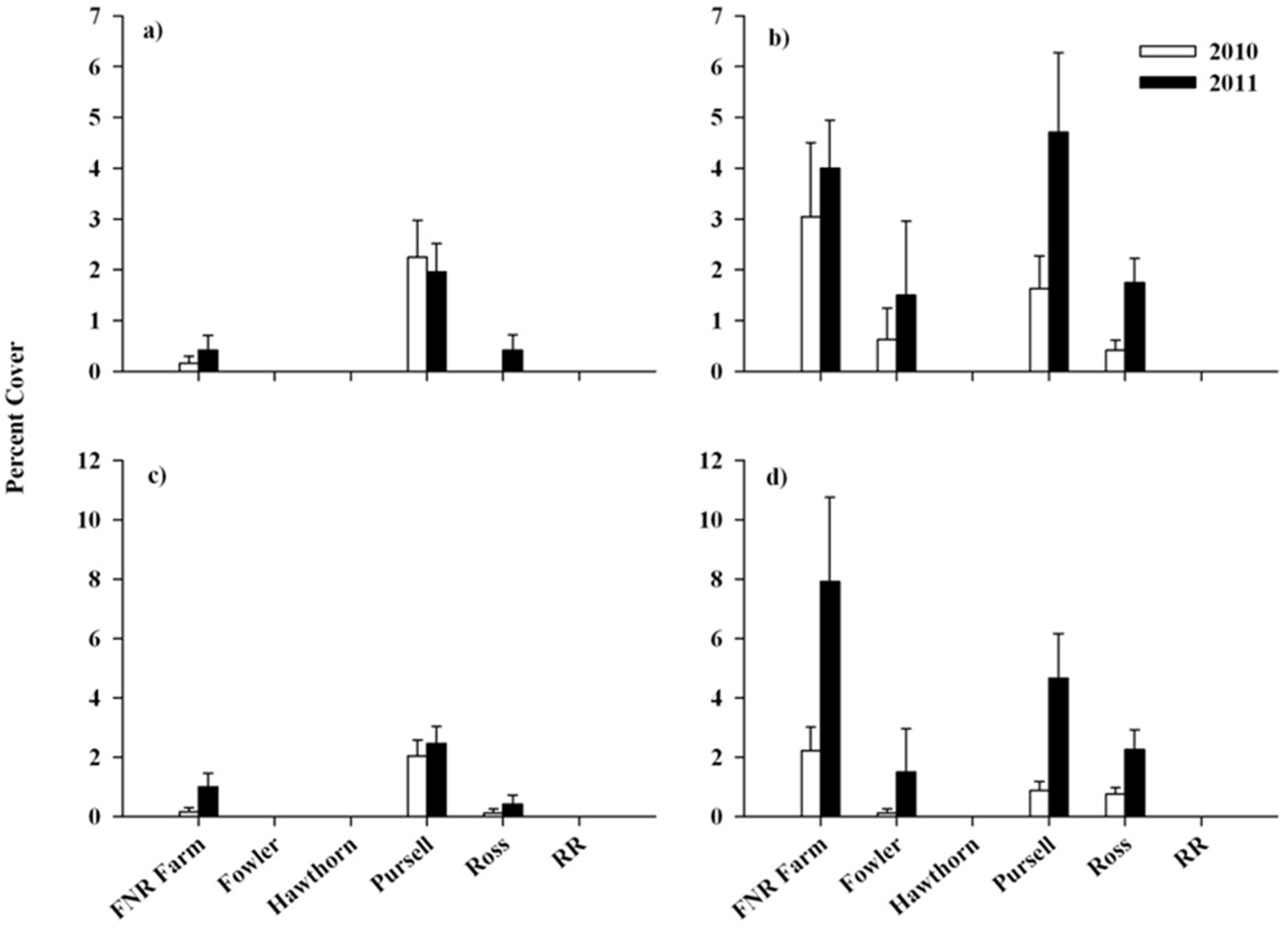{kind=link}
| Study Site | Dominant Overstory Cohort | Soil Type(s) | L. maaccii Saplings/ha |
|---|---|---|---|
| Fowler Park | Fraxinus americana Juglans nigra, Prunus serotina, Ulmus americana, Sassafras albidum | Silt loam | 2500 ± 459 |
| FNR Farm | Robinia pseudoacacia, U. americana | Silt loam | 3135 ± 176 |
| Hawthorn Park | S. albidum, Nyssa sylvatica, Liriodendron tulipifera, P. serotina, Quercus bicolor | Silt loam | 1708 ± 256 |
| Pursell | Maclura pomifera, P. serotina | Loam, loamy sand, sandy loams, silt loams | 1354 ± 255 |
| Rifle Range | L. tulipifera, P. serotina, S. albidum | Fine sandy loams | 2375 ± 158 |
2.4. Data Analyses
3. Results
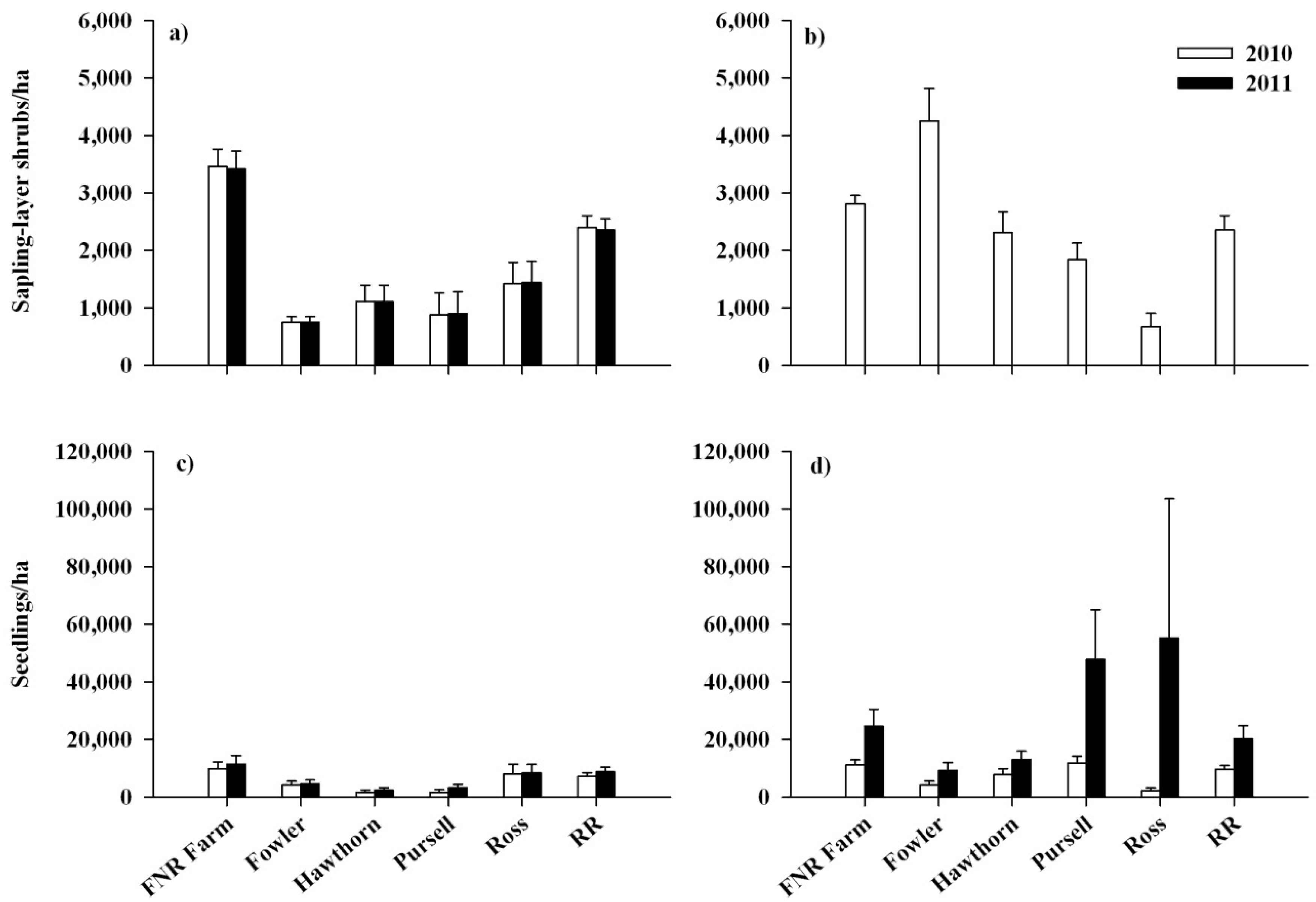
3.1. Response of Lonicera maackii
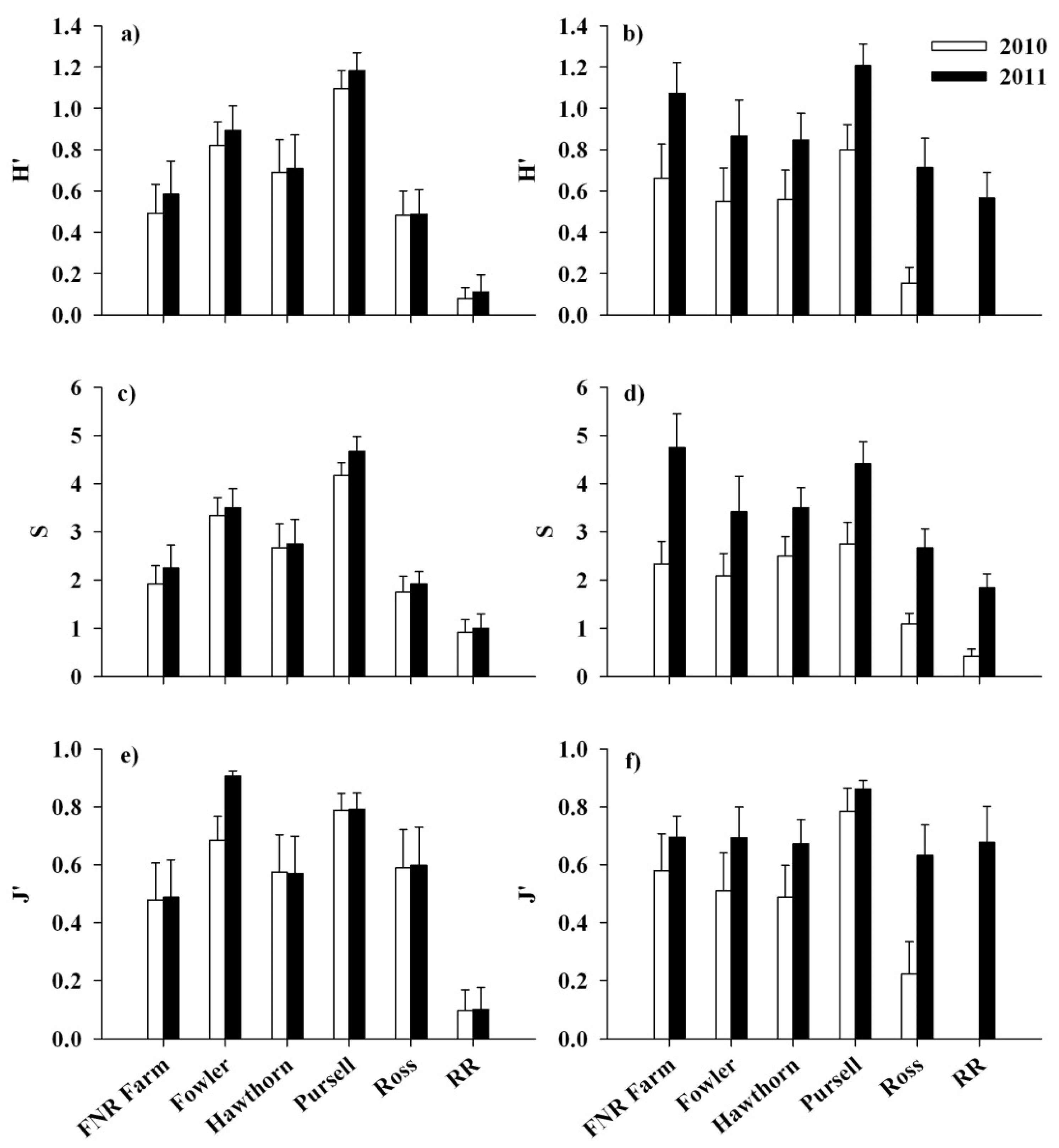
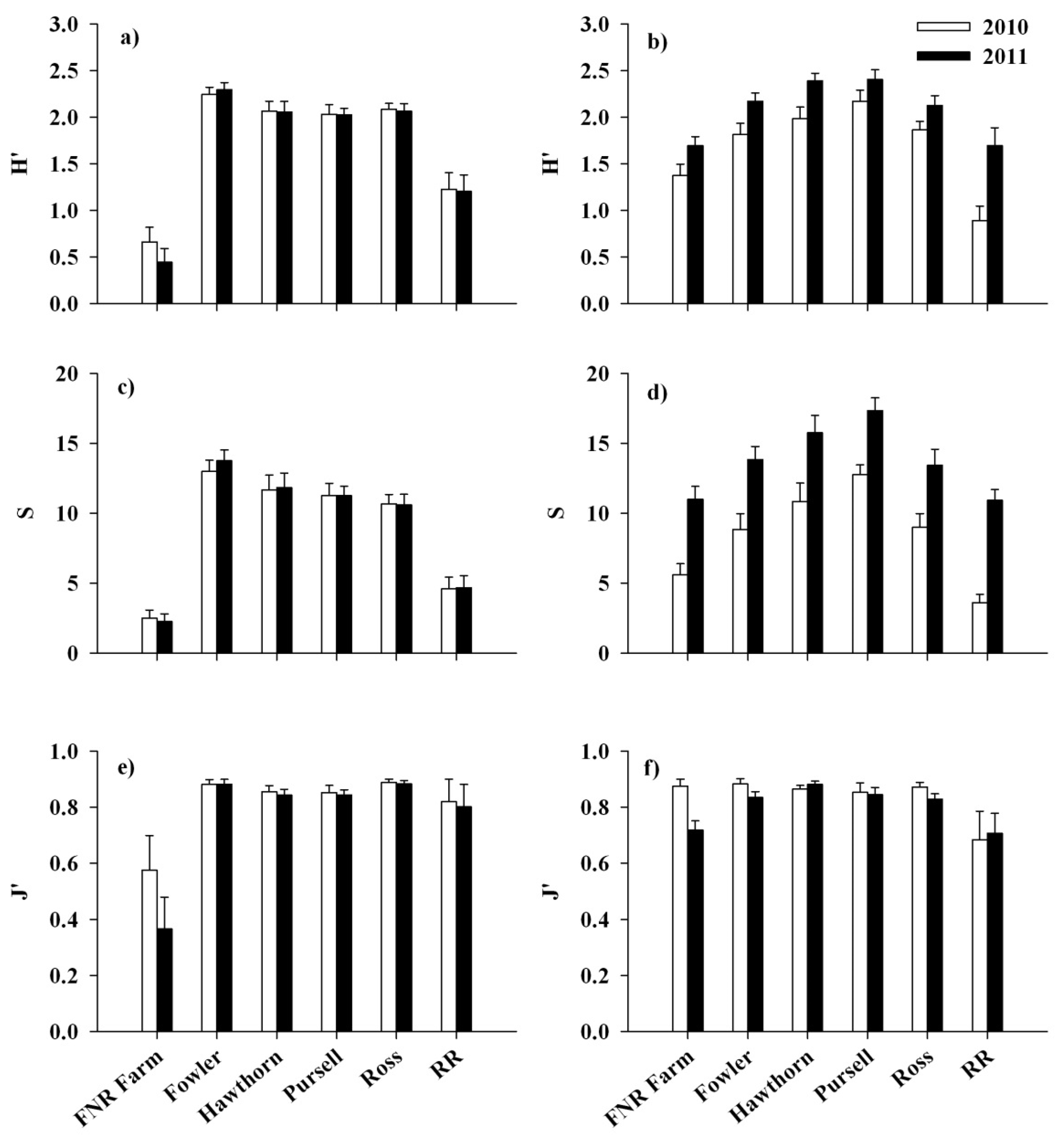
3.2. Response of Native Flora and Alliaria petiolata
| Vegetation Group | Reference | Removal | ||||
|---|---|---|---|---|---|---|
| 2010 | 2011 | p | 2010 | 2011 | p | |
| Lower stratum (≤1 m) | ||||||
| Native forbs | 20.6 ± 6.5 | 22.6 ± 6.9 | 0.19 | 17.2 ± 3.8 | 31.0 ± 6.3 | 0.04 |
| Native grasses | 0.6 ± 0.4 | 0.7 ± 0.5 | 0.78 | 0.5 ± 0.2 | 0.6 ± 0.3 | 0.23 |
| Native sedges | 0.2 ± 0.1 | 0.2 ± 0.1 | --- | 0.5 ± 0.3 | 0.9 ± 0.3 | 0.08 |
| Native ferns | 0.2 ± 0.1 | 0.4 ± 0.2 | 0.22 | 0.2 ± 0.1 | 0.4 ± 0.1 | 0.08 |
| Native vines | 0.7 ± 0.3 | 1.06 ± 0.31 | 0.32 | 0.8 ± 0.5 | 1.4 ± 0.7 | 0.08 |
| Native trees and shrubs | 2.8 ± 1.1 | 4.6 ± 1.6 | 0.17 | 3.8 ± 1.7 | 5.3 ± 1.5 | 0.06 |
| Non-native forbs | 0.5 ± 0.4 | 0.7 ± 0.4 | 0.19 | 1.0 ± 0.5 | 2.3 ± 0.9 | 0.06 |
| Non-native grasses | T | T | 0.78 | --- | --- | --- |
| Non-native shrubs | 12.0 ± 2.6 | 12.3 ± 2.9 | 0.71 | 18.7 ± 4.0 | 3.3 ± 0.6 | 0.04 |
| Non-native vines | 0.3 ± 0.3 | 0.3 ± 0.3 | --- | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.23 |
| Upper stratum (1.01–5 m) | ||||||
| Native trees and shrubs | 9.8 ± 3.2 | 10.9 ± 3.4 | 0.19 | 9.3 ± 2.7 | 9.7 ± 2.8 | 0.20 |
| Native vines | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.78 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.30 |
| Non-native shrubs | 35.5 ± 9.3 | 35.6 ± 10.5 | 0.71 | 40.6 ± 7.0 | 0.1 ± 0.1 | 0.04 |
| Non-native vines | --- | --- | --- | 0.1 ± 0.1 | --- | 0.49 |
| Substrate variables | ||||||
| Bare soil | 15.6 ± 6.9 | 14.7 ± 5.8 | 0.78 | 13.6 ± 4.0 | 10.1 ± 2.9 | 0.06 |
| Coarse woody debris | 5.2 ± 2.1 | 5.1 ± 2.1 | 0.78 | 3.0 ± 0.7 | 3.2 ± 0.8 | 0.24 |
| Fine woody debris | 7.9 ± 1.1 | 8.1 ± 1.0 | 0.56 | 8.6 ± 1.4 | 10.6 ± 1.3 | 0.04 |
| Dead leaves/dead herbaceous stems | 35.1 ± 8.3 | 36.7 ± 7.9 | 0.56 | 27.8 ± 8.8 | 26.7 ± 8.2 | 0.24 |
| Vegetation Group | Reference | Removal | ||||
|---|---|---|---|---|---|---|
| 2010 | 2011 | p | 2010 | 2011 | p | |
| Lower stratum (≤1 m) | ||||||
| Native forbs | 20.9 ± 6.6 | 20.1 ± 6.3 | 0.78 | 22.7 ± 5.2 | 37.8 ± 6.8 | 0.04 |
| Native grasses | 0.9 ± 0.6 | 0.8 ± 0.5 | 0.56 | 0.7 ± 0.3 | 0.8 ± 0.4 | 0.24 |
| Native sedges | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.78 | 0.9 ± 0.3 | 1.0 ± 0.4 | 0.13 |
| Native ferns | 0.4 ± 0.2 | 0.4 ± 0.2 | 0.78 | 0.3 ± 0.1 | 0.4 ± 0.1 | 0.08 |
| Native vines | 1.4 ± 0.5 | 1.4 ± 0.4 | 0.78 | 2.1 ± 1.1 | 2.7 ± 0.8 | 0.15 |
| Native trees and shrubs | 5.7 ± 2.0 | 5.0 ± 1.6 | 0.22 | 6.4 ± 2.5 | 8.7 ± 2.1 | 0.06 |
| Non-native forbs | 0.6 ± 0.4 | 1.0 ± 0.5 | 0.19 | 1.1 ± 0.6 | 3.3 ± 1.5 | 0.06 |
| Non-native grasses | T | T | --- | --- | --- | --- |
| Non-native shrubs | 12.0 ± 2.6 | 12.7 ± 2.9 | 0.22 | 19.9 ± 3.7 | 4.7 ± 0.7 | 0.04 |
| Non-native vines | 0.3 ± 0.3 | 0.4 ± 0.4 | 0.78 | 0.2 ± 0.1 | 0.3 ± 0.2 | 0.49 |
| Upper stratum (1.01–5 m) | ||||||
| Native trees and shrubs | 13.2 ± 4.4 | 12.6 ± 4.1 | 0.32 | 12.4 ± 3.4 | 11.6 ± 3.0 | 0.24 |
| Native vines | 0.2 ± 0.1 | 0.2 ± 0.1 | --- | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.15 |
| Non-native shrubs | 37.2 ± 10.3 | 37.3 ± 10.3 | 0.56 | 42.4 ± 6.9 | 0.33 ± 0.31 | 0.04 |
| Non-native vines | --- | --- | --- | 0.1 ± 0.1 | --- | 0.49 |
| Substrate variables | ||||||
| Bare soil | 17.8 ± 7.4 | 15.0 ± 6.0 | 0.19 | 15.1 ± 4.0 | 8.4 ± 2.9 | 0.04 |
| Coarse woody debris | 5.2 ± 2.1 | 5.3 ± 2.1 | 0.78 | 3.3 ± 0.9 | 3.4 ± 1.0 | 0.23 |
| Fine woody debris | 8.3 ± 1.1 | 8.3 ± 1.2 | --- | 8.9 ± 1.6 | 12.5 ± 3.0 | 0.04 |
| Dead leaves/dead herbaceous stems | 30.0 ± 8.8 | 31.6 ± 7.1 | 0.56 | 22.2 ± 8.4 | 19.4 ± 7.7 | 0.15 |
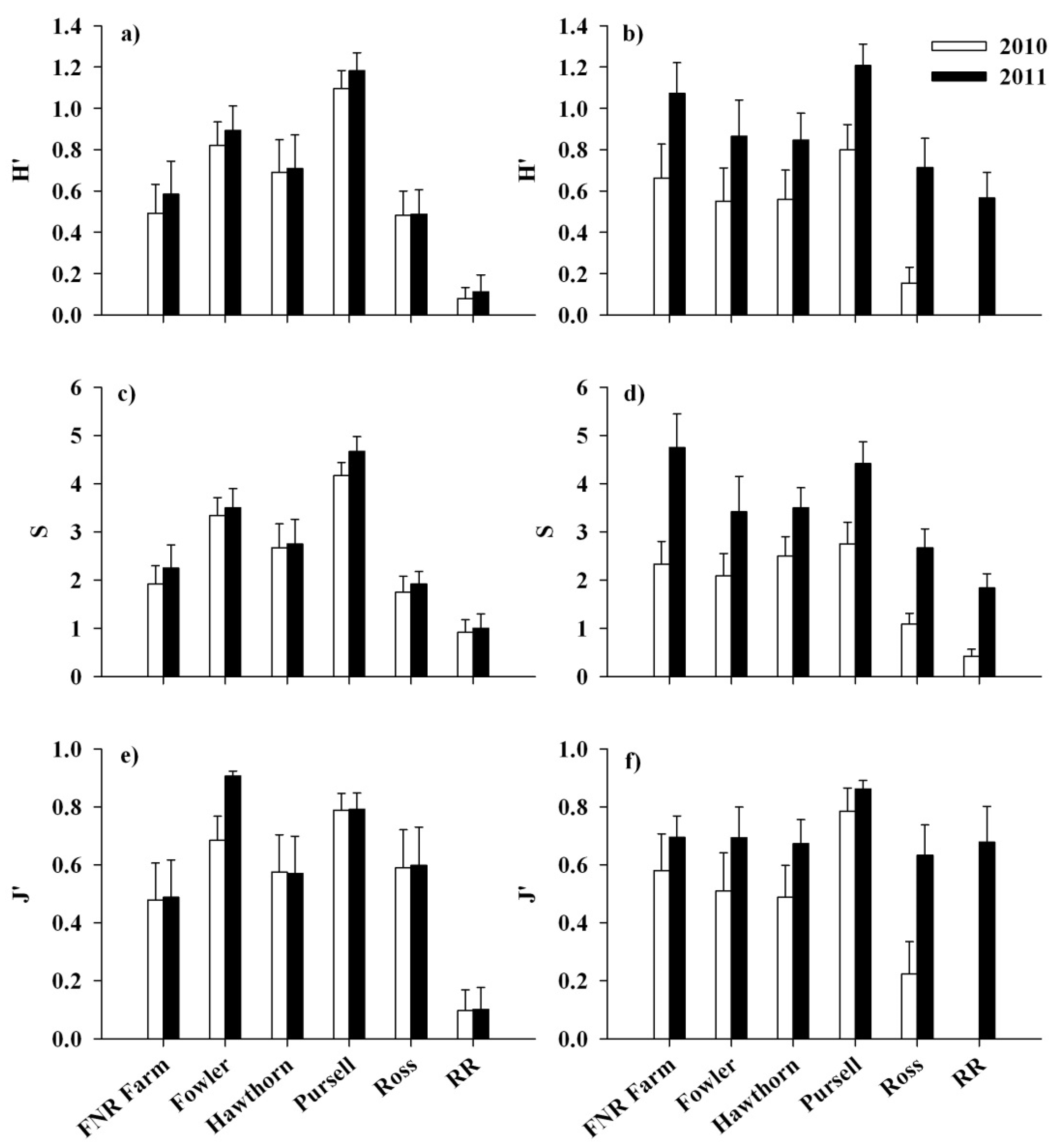
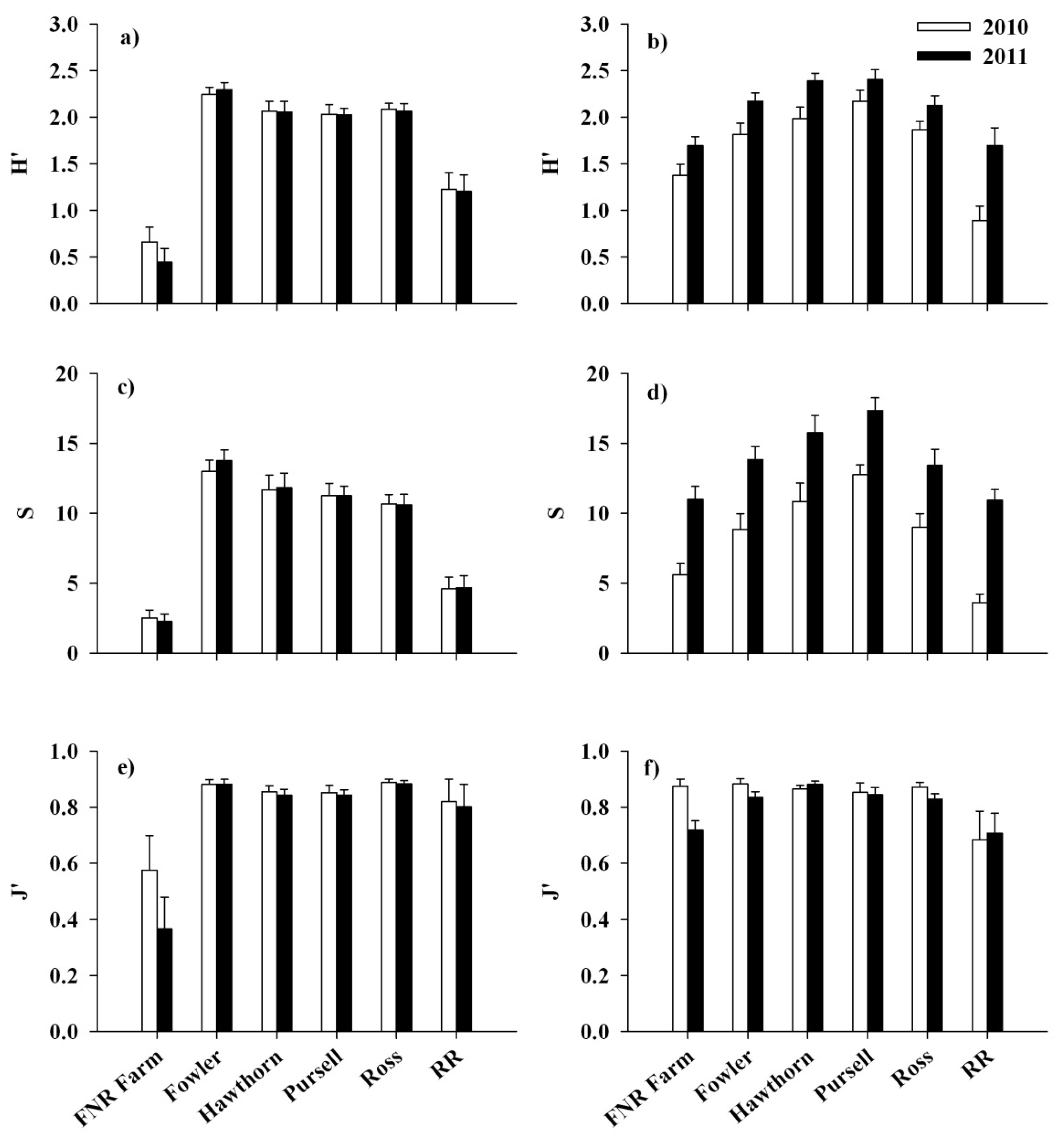


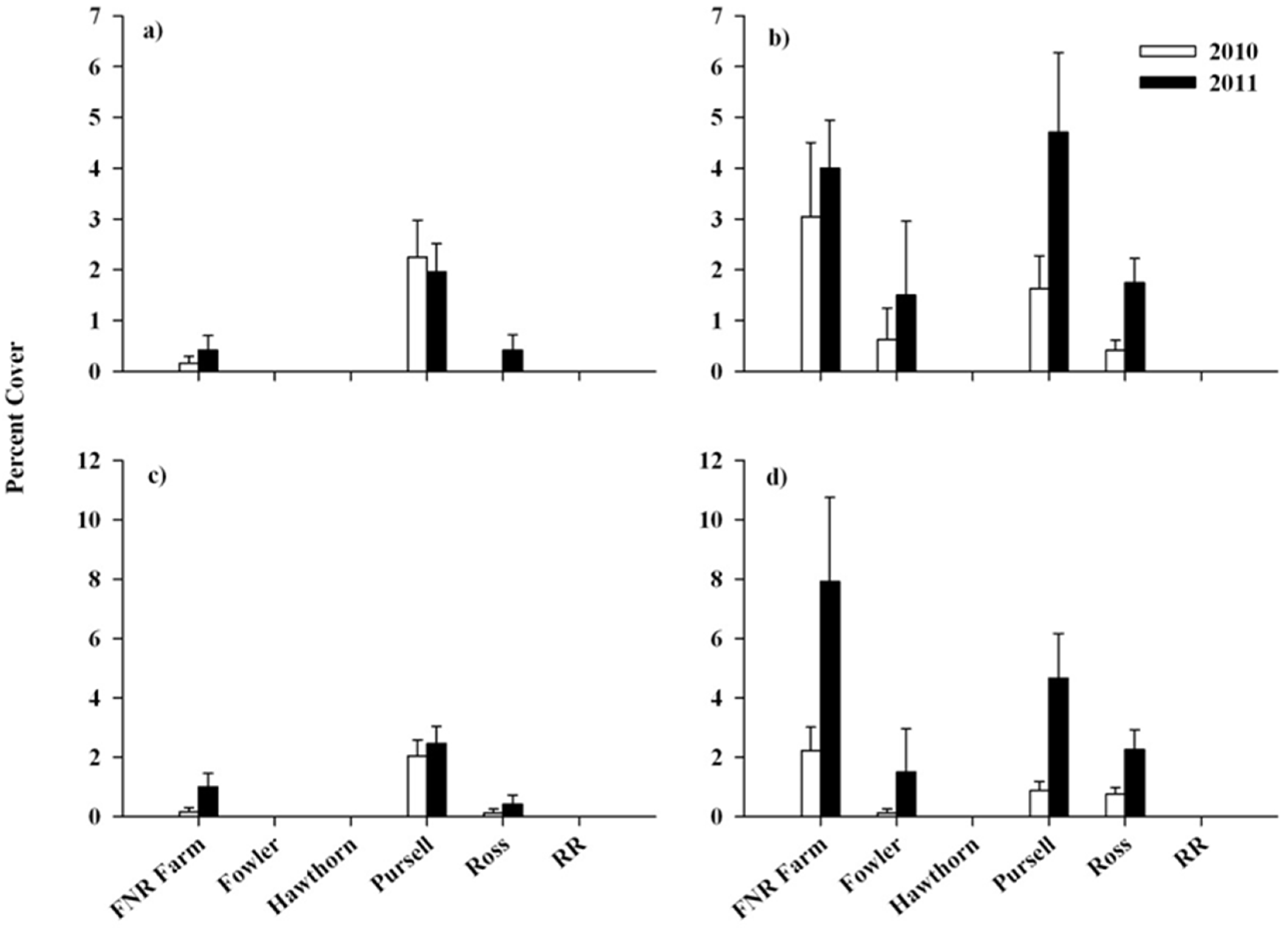
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Supporting Information
Conflicts of Interest
References
- Blossey, B. Before, during and after: The need for long-term monitoring in invasive plant species management. Biol. Invasions 1999, 1, 301–311. [Google Scholar] [CrossRef]
- Reid, A.M.; Morin, L.; Downey, P.O.; French, K.; Virtue, J.G. Does invasive plant management aid the restoration of natural ecosystems? Biol. Conserv. 2009, 142, 2342–2349. [Google Scholar] [CrossRef]
- Webster, C.R.; Jenkins, M.A.; Jose, S. Woody invaders and the challenges they pose to forest ecosystems in the eastern United States. J. For. 2006, 104, 366–374. [Google Scholar]
- Uddin, M.B.; Steinbauer, M.J.; Jentsch, A.; Mukul, S.A.; Beierkuhnlein, C. Do environmental attributes, disturbances, and protection regimes determine the distribution of exotic plant species in Bangladesh forest ecosystem? For. Ecol. Manag. 2013, 303, 72–80. [Google Scholar] [CrossRef]
- Luken, J.O.; Thieret, J.W. Amur honeysuckle, its fall from grace: Lessons from the introduction and spread of a shrub species may guide future plant introductions. BioScience 1996, 46, 18–24. [Google Scholar] [CrossRef]
- Gould, A.M.; Gorchov, D.L. Effects of the exotic invasive shrub Lonicera maackii on the survival and fecundity of three species of native annuals. Am. Midl. Nat. 2000, 144, 36–50. [Google Scholar] [CrossRef]
- Collier, M.H.; Vankat, J.L.; Hughes, M.R. Diminished plant richness and abundance below Lonicera maackii, an invasive shrub. Am. Midl. Nat. 2002, 147, 60–71. [Google Scholar] [CrossRef]
- Gorchov, D.L.; Trisel, D.E. Competitive effects of the invasive shrub Lonicera maackii. Plant Ecol. 2003, 166, 13–24. [Google Scholar] [CrossRef]
- Miller, K.E.; Gorchov, D.L. The invasive shrub, Lonicera maackii, reduces growth and fecundity of perennial forest herbs. Oecologia 2004, 139, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Hartman, K.M.; McCarthy, B.C. Changes in forest structure and species composition following invasion by a non-indigenous shrub, Amur honeysuckle (Lonicera maackii). J. Torrey Bot. Soc. 2008, 135, 245–259. [Google Scholar] [CrossRef]
- Meiners, S.J. Apparent competition: An impact of exotic shrub invasion on tree regeneration. Biol. Invasions 2007, 9, 849–855. [Google Scholar] [CrossRef]
- Hutchinson, T.F.; Vankat, J.L. Invasibility and effects of Amur honeysuckle in Southwestern Ohio forests. Conserv. Biol. 1997, 11, 1117–1124. [Google Scholar] [CrossRef]
- Luken, J.O.; Kuddes, L.M.; Tholemeier, T.C. Response of understory species to gap formation and soil disturbance in Lonicera maackii thickets. Restor. Ecol. 1997, 5, 229–235. [Google Scholar] [CrossRef]
- Hartman, K.M.; McCarthy, B.C. Restoration of a forest understory after the removal of an invasive shrub, Amur honeysuckle (Lonicera maackii). Restor. Ecol. 2004, 12, 154–165. [Google Scholar] [CrossRef]
- Runkle, J.R.; diSalvo, A.; Graham-Gibson, Y.; Dorning, M. Vegetation release eight years after removal of Lonicera maackii in west-central Ohio. Ohio J. Sci. 2007, 107, 125–129. [Google Scholar]
- Cipollini, K.; Ames, E.; Cipollini, D. Amur honeysuckle (Lonicera maackii) management method impacts restoration of understory plants in the presence of white-tailed deer (Odocoileus virginiana). Invasive Plant Sci. Manag. 2009, 2, 45–54. [Google Scholar] [CrossRef]
- Swab, R.M.; Zhang, L.; Mitsch, W.J. Effect of hydrologic restoration and Lonicera maackii removal on herbaceous understory vegetation in a bottomland hardwood forest. Restor. Ecol. 2008, 16, 453–463. [Google Scholar] [CrossRef]
- Shields, J.M.; Jenkins, M.A.; Saunders, M.R.; Zhang, H.; Jenkins, L.H.; Parks, A.M. Age distribution and spatial patterning of an invasive shrub in secondary hardwood forests. For. Sci. 2014, 60, 830–840. [Google Scholar] [CrossRef]
- Dutra, H.P.; Barnett, K.; Reinhardt, J.R.; Marquis, R.J.; Orrock, J.L. Invasive plant species alters consumer behavior by providing refuge from predation. Oecologia 2011, 166, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.M.; Jenkins, M.A.; Zollner, P.A.; Saunders, M.R. Effects of Amur honeysuckle invasion and removal on white-footed mice. J. Wildlife Manag. 2014, 78, 867–880. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Manson, R.H.; Canham, C.D. Effects of rodents on survival of tree seeds and seedlings invading old fields. Ecology 1997, 78, 1531–1542. [Google Scholar] [CrossRef]
- Whigham, D.F. Ecology of woodland herbs in temperate deciduous forests. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 583–621. [Google Scholar] [CrossRef]
- Owen, H.R.; McDonnell, A.L.; Mounteer, A.M.; Todd, B.L. Influence of stem cutting and glyphosate treatment of Lonicera maackii, an exotic and invasive species, on stem regrowth and native species richness. Trans. Illinois State Acad. Sci. 2005, 98, 1–17. [Google Scholar]
- Harrington, R.A.; Brown, B.J.; Reich, P.B. Ecophysiology of exotic and native shrubs in southern Wisconsin. I. Relationship of leaf characteristics, resource availability, and phenology to seasonal patterns of carbon gain. Oecologia 1989, 80, 356–367. [Google Scholar] [CrossRef]
- Harrington, R.A.; Brown, B.J.; Reich, P.B.; Fownes, J.H. Ecophysiology of exotic and native shrubs in southern Wisconsin. II. Annual growth and carbon gain. Oecologia 1989, 80, 368–373. [Google Scholar] [CrossRef]
- Homer, C.; Dewitz, J.; Fry, J.; Coan, M.; Hossain, N.; Larson, C.; Herold, N.; McKerrow, A.; VanDriel, J.N.; Wickham, J. Completion of the 2001 National Land Cover Database for the conterminous United States. Photogramm. Eng. Remote Sens. 2007, 73, 337–341. [Google Scholar]
- Soil Survey Staff; Natural Resources Conservation Service; United States Department of Agriculture. Web Soil Survey. Available online: http://websoilsurvey.nrcs.usda.gov/ (accessed on 20 March 2013).
- Peet, R.K.; Wentworth, T.R.; White, P.S. A flexible, multipurpose method for recording vegetation composition and structure. Castanea 1998, 63, 262–274. [Google Scholar]
- US Department of Agriculture. The PLANTS Database USDA. 2013. Available online: http://plants.usda.gov (accessed on 29 October 2013). [Google Scholar]
- Yatskievych, K. Field Guide to Indiana Wildflowers; Indiana University Press: Bloomington, IN, USA, 2000. [Google Scholar]
- Hollander, M.; Wolf, D.A. Nonparametric Statistical Methods, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Carter, D.L.; Blair, J.M. Recovery of native plant community characteristics on a chronosequence of restored prairies seeded into pastures in west-central Iowa. Restor. Ecol. 2012, 20, 170–179. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.R-project.org (accessed on 8 July 2013).
- Oksanen, J.; Blanchet, G.F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; Wagner, H. Vegan: Community Ecology. Available online: http://CRAN.R-project.org/package=vegan (accessed on 20 February 2013).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. Royal. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. On the adaptive control of the false discovery rate in multiple testing with independent statistics. J. Educ. Behav. Stat. 2000, 25, 60–83. [Google Scholar] [CrossRef]
- Pike, N. Using false discovery rates for multiple comparisons in ecology and evolution. Meth. Ecol. Evol. 2011, 2, 278–282. [Google Scholar] [CrossRef]
- Verhoeven, K.J.F.; Simonsen, K.L.; McIntyre, L.M. Implementing false discovery rate control: Increasing your power. Oikos 2005, 108, 643–647. [Google Scholar] [CrossRef]
- Schemske, D.W.; Willson, M.F.; Melampy, M.N.; Miller, L.J.; Verner, L.; Schemske, K.M.; Best, L.B. Flowering ecology of some spring woodland herbs. Ecology 1978, 59, 351–366. [Google Scholar] [CrossRef]
- Bierzychudek, P. Life histories and demography of shade-tolerant temperate forest herbs: A review. New Phytol. 1982, 90, 757–776. [Google Scholar] [CrossRef]
- Bartuszevige, A.M.; Gorchov, D.L.; Raab, L. The relative importance of landscape and community features in the invasion of an exotic shrub in a fragmented landscape. Ecography 2006, 29, 213–222. [Google Scholar] [CrossRef]
- Castellano, S.M.; Gorchov, D.L. White-tailed deer (Odocoileus virginianus) disperse seeds of the invasive shrub, Amur honeysuckle (Lonicera maackii). Nat. Areas J. 2013, 33, 78–80. [Google Scholar] [CrossRef]
- Williams, C.E.; Ralley, J.J.; Taylor, D.H. Consumption of seeds of the invasive Amur honeysuckle, Lonicera maackii (Rupr.) Maxim. by small mammals. Nat. Areas J. 1992, 12, 86–89. [Google Scholar]
- Bartuszevige, A.M.; Hrenko, R.L.; Gorchov, D.L. Effects of leaf litter on establishment, growth and survival of invasive plant seedlings in a deciduous forest. Am. Midl. Nat. 2007, 158, 472–477. [Google Scholar] [CrossRef]
- Rooney, T.P. Deer impacts on forest ecosystems: A North American perspective. Forestry 2001, 74, 201–208. [Google Scholar] [CrossRef]
- Webster, C.R.; Rock, J.R.; Froese, R.E.; Jenkins, M.A. Drought-herbivory interaction disrupts competitive displacement of native plants by Microstegium vimineum, 10 year results. Oecologia 2008, 157, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Knight, T.M.; Dunn, J.L.; Smith, L.A.; Davis, J.; Kalisz, S. Deer facilitate invasive plant success in a Pennsylvania forest understory. Nat. Areas J. 2009, 29, 110–116. [Google Scholar] [CrossRef]
- Dornbush, M.E.; Hahn, P.G. Consumer and establishment limitation contribute more than competitive interactions in sustaining dominance of the exotic herb garlic mustard in a Wisconsin, USA forest. Biol. Invasions 2013, 15, 2691–2705. [Google Scholar] [CrossRef]
- Waller, D.M.; Maas, L.I. Do white-tailed deer and the exotic plant garlic mustard interact to affect the growth and persistence of native forest plants? For. Ecol. Manag. 2013, 304, 296–302. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shields, J.M.; Saunders, M.R.; Gibson, K.D.; Zollner, P.A.; Dunning, J.B., Jr.; Jenkins, M.A. Short-Term Response of Native Flora to the Removal of Non-Native Shrubs in Mixed-Hardwood Forests of Indiana, USA. Forests 2015, 6, 1878-1896. https://doi.org/10.3390/f6061878
Shields JM, Saunders MR, Gibson KD, Zollner PA, Dunning JB Jr., Jenkins MA. Short-Term Response of Native Flora to the Removal of Non-Native Shrubs in Mixed-Hardwood Forests of Indiana, USA. Forests. 2015; 6(6):1878-1896. https://doi.org/10.3390/f6061878
Chicago/Turabian StyleShields, Joshua M., Michael R. Saunders, Kevin D. Gibson, Patrick A. Zollner, John B. Dunning, Jr., and Michael A. Jenkins. 2015. "Short-Term Response of Native Flora to the Removal of Non-Native Shrubs in Mixed-Hardwood Forests of Indiana, USA" Forests 6, no. 6: 1878-1896. https://doi.org/10.3390/f6061878
APA StyleShields, J. M., Saunders, M. R., Gibson, K. D., Zollner, P. A., Dunning, J. B., Jr., & Jenkins, M. A. (2015). Short-Term Response of Native Flora to the Removal of Non-Native Shrubs in Mixed-Hardwood Forests of Indiana, USA. Forests, 6(6), 1878-1896. https://doi.org/10.3390/f6061878