
Applying Resilience Concepts in Forest Management: A Retrospective Simulation Approach

Abstract
:1. Introduction
2. Experimental Section
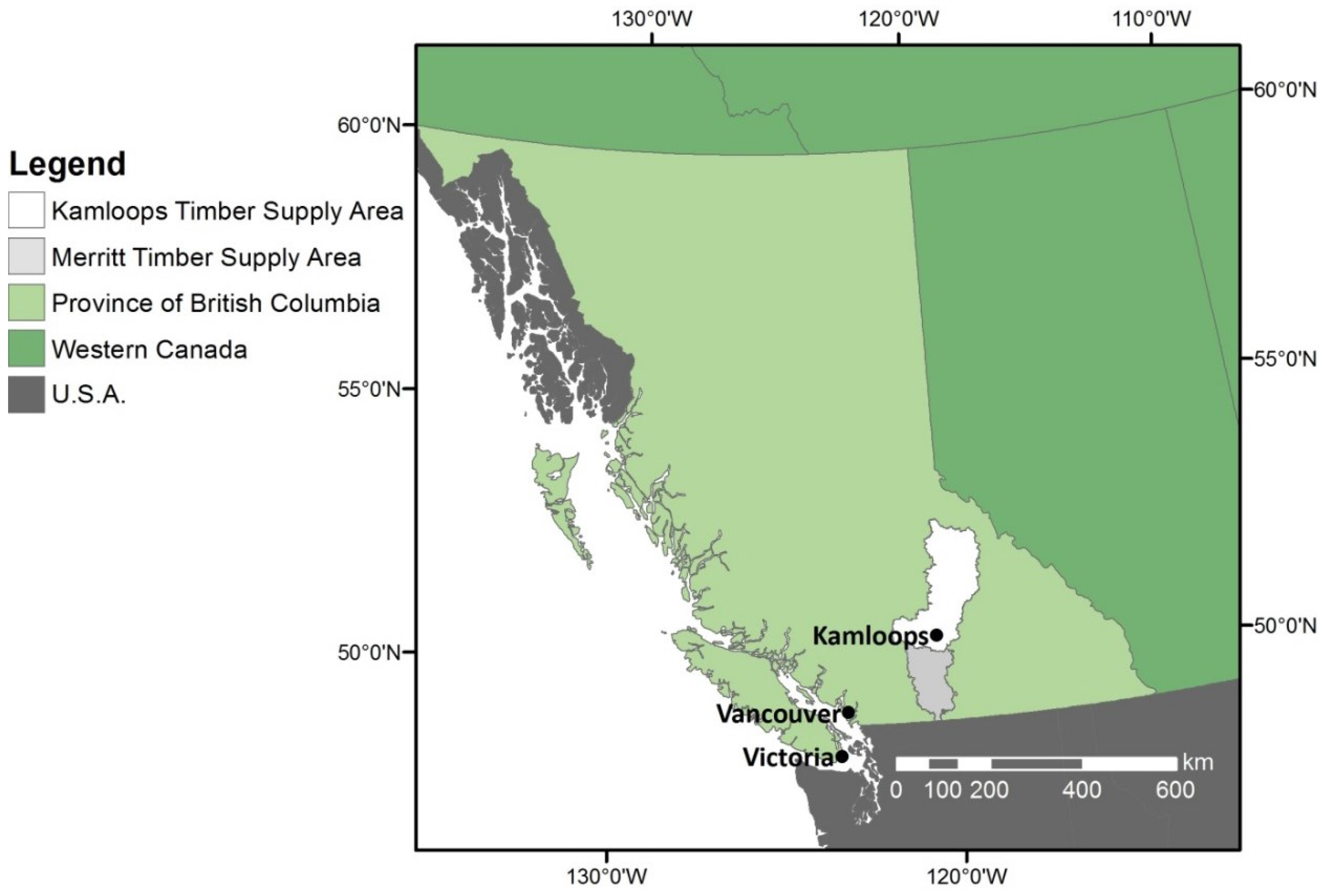
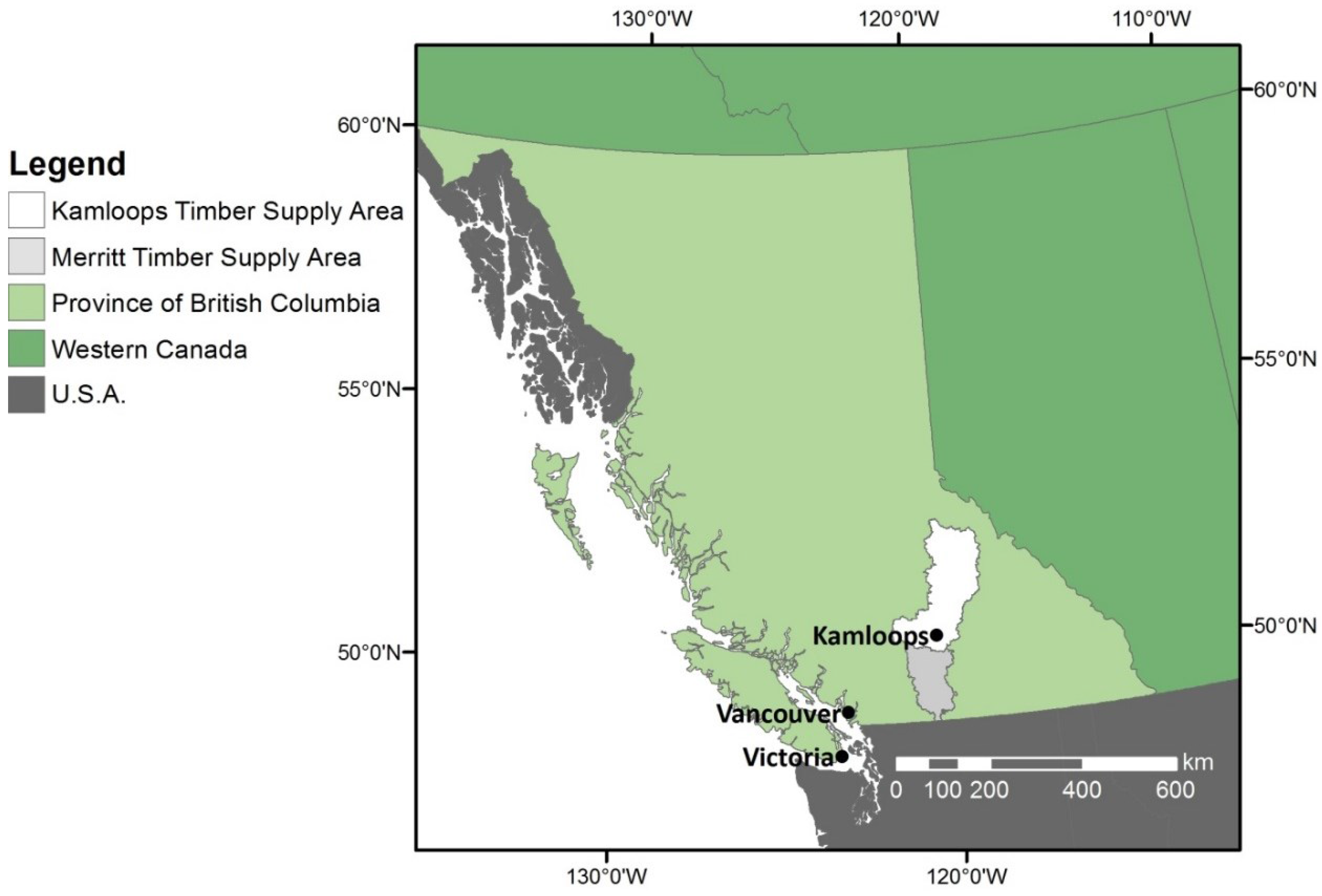
2.1. Study Area
2.2. Modelling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strategy Name | Harvesting | Regeneration (see also Figure 4) |
|---|---|---|
| Business as usual (BAU) |
|
|
| Mixed planting (MP) |
|
|
| Early pine cut, mixed planting, increased natural regeneration, (EMR) |
|
|
2.3. Resilience Indicators
3. Results
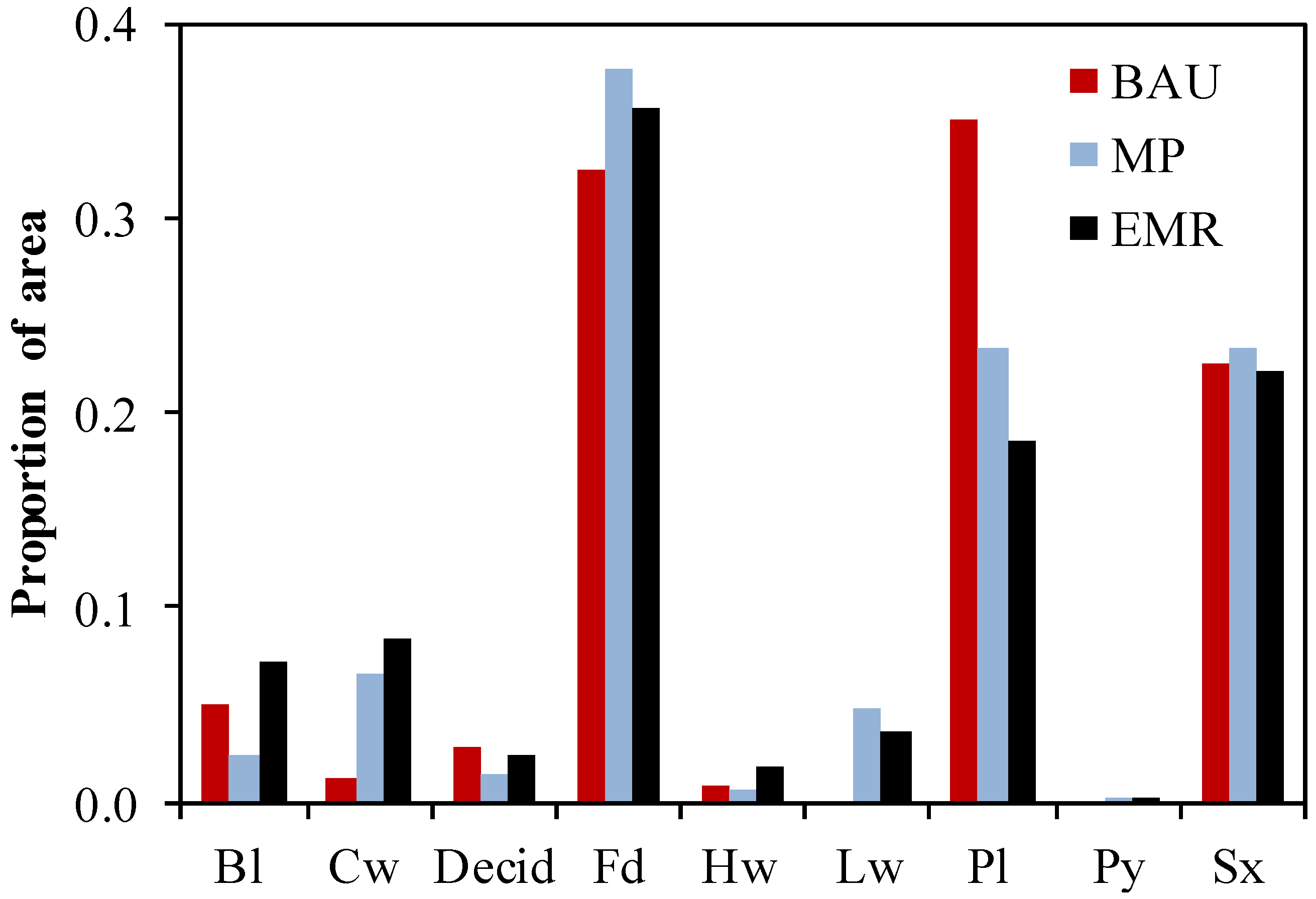
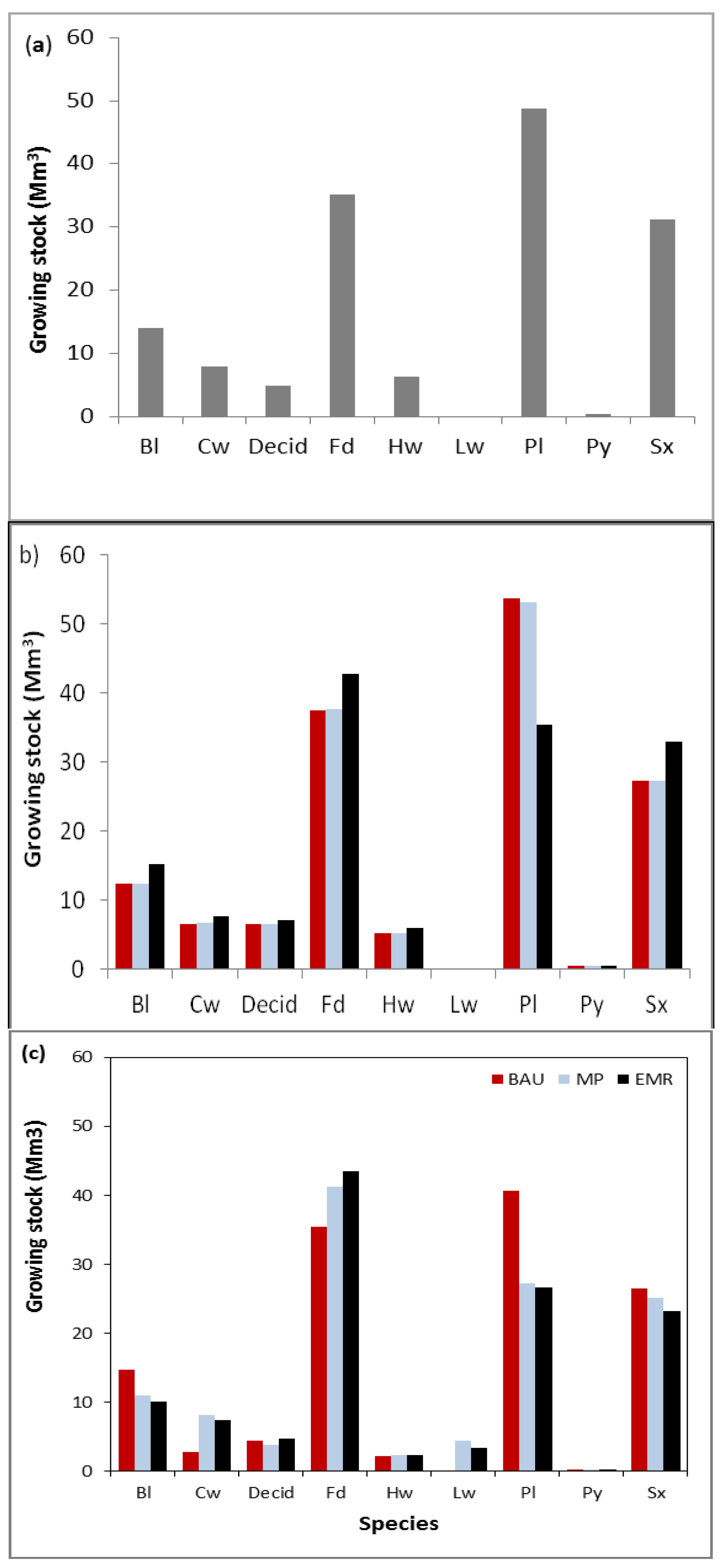
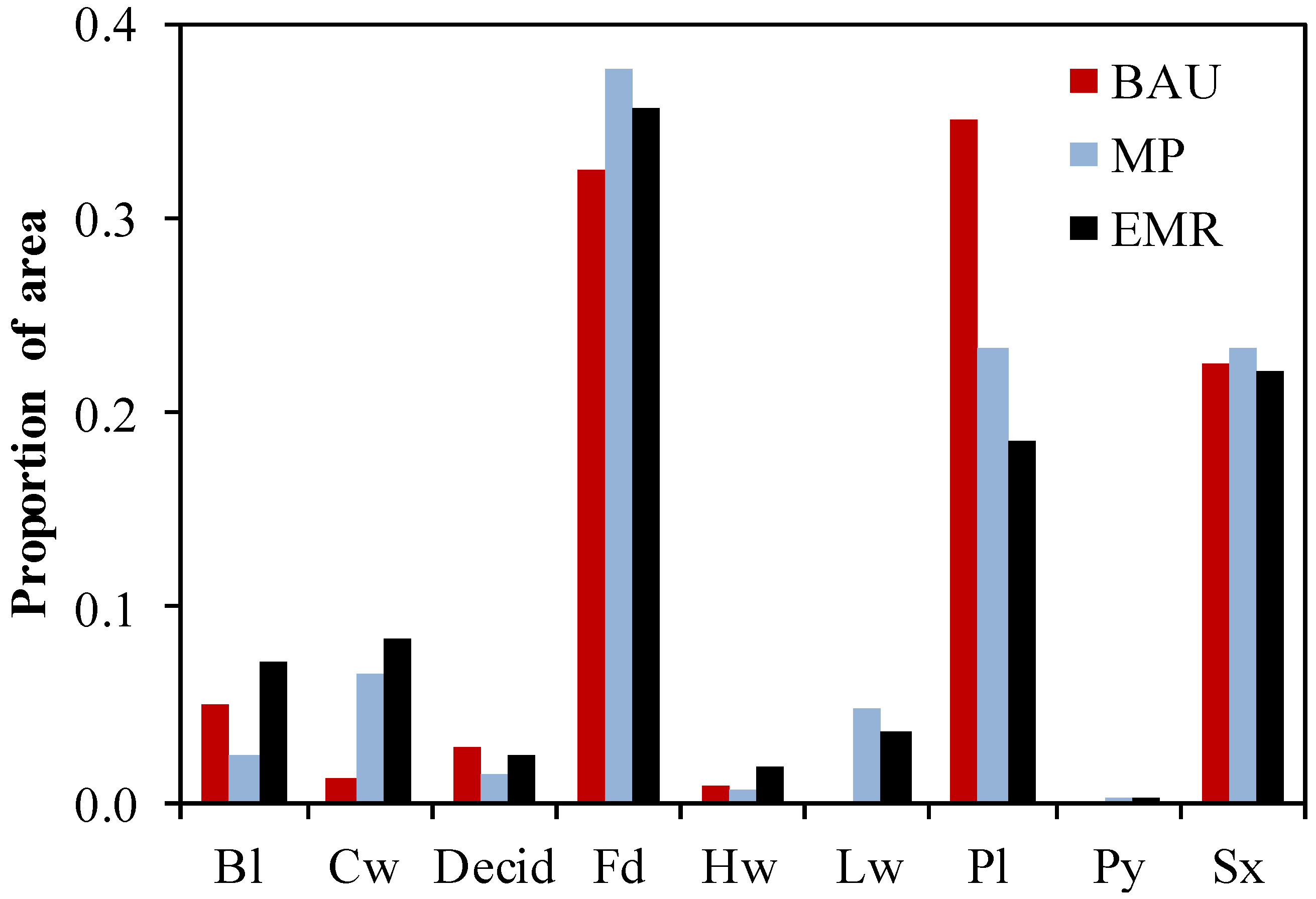
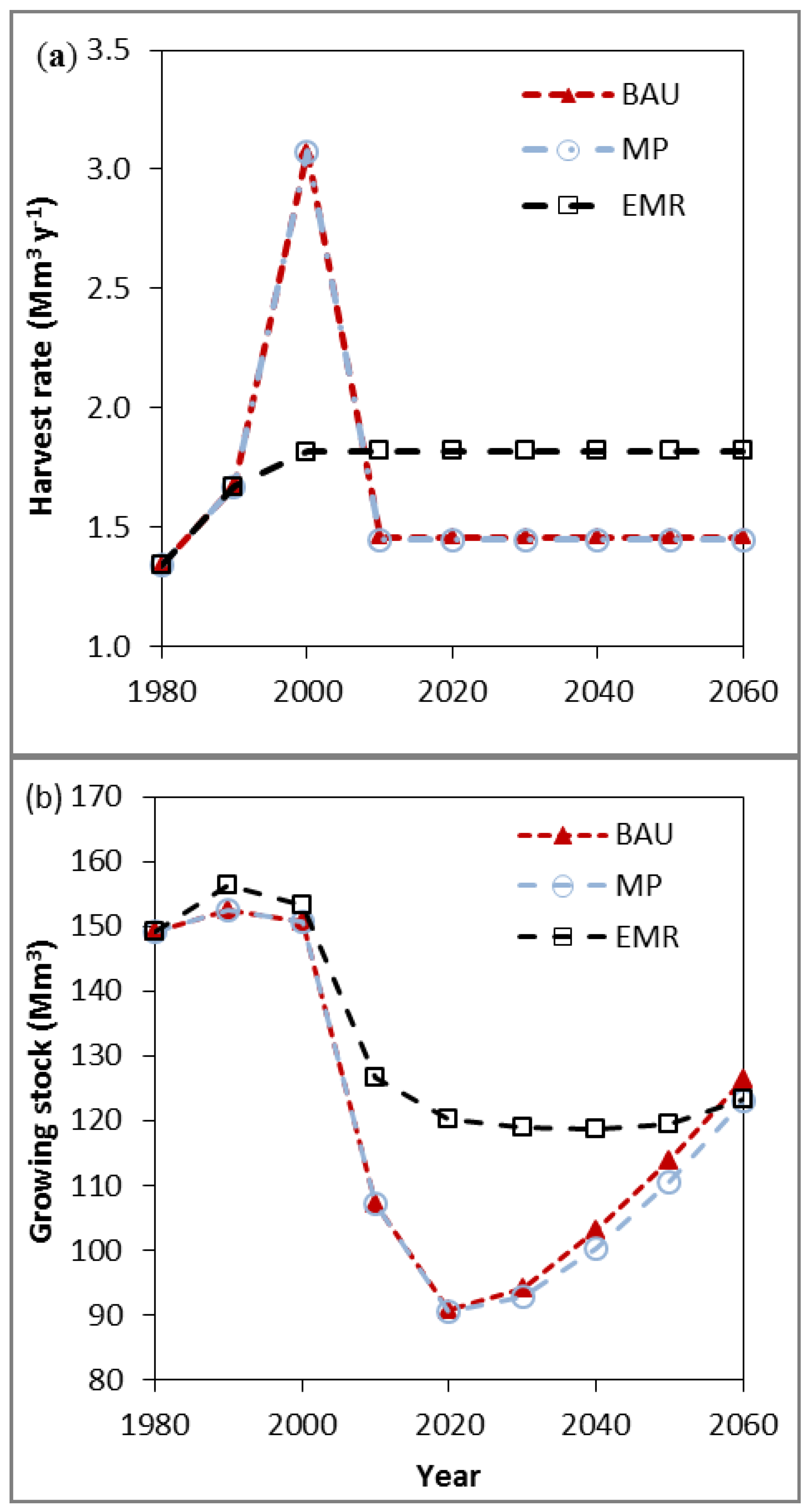
3.1. Species Composition, Growing Stock and Harvest Rates

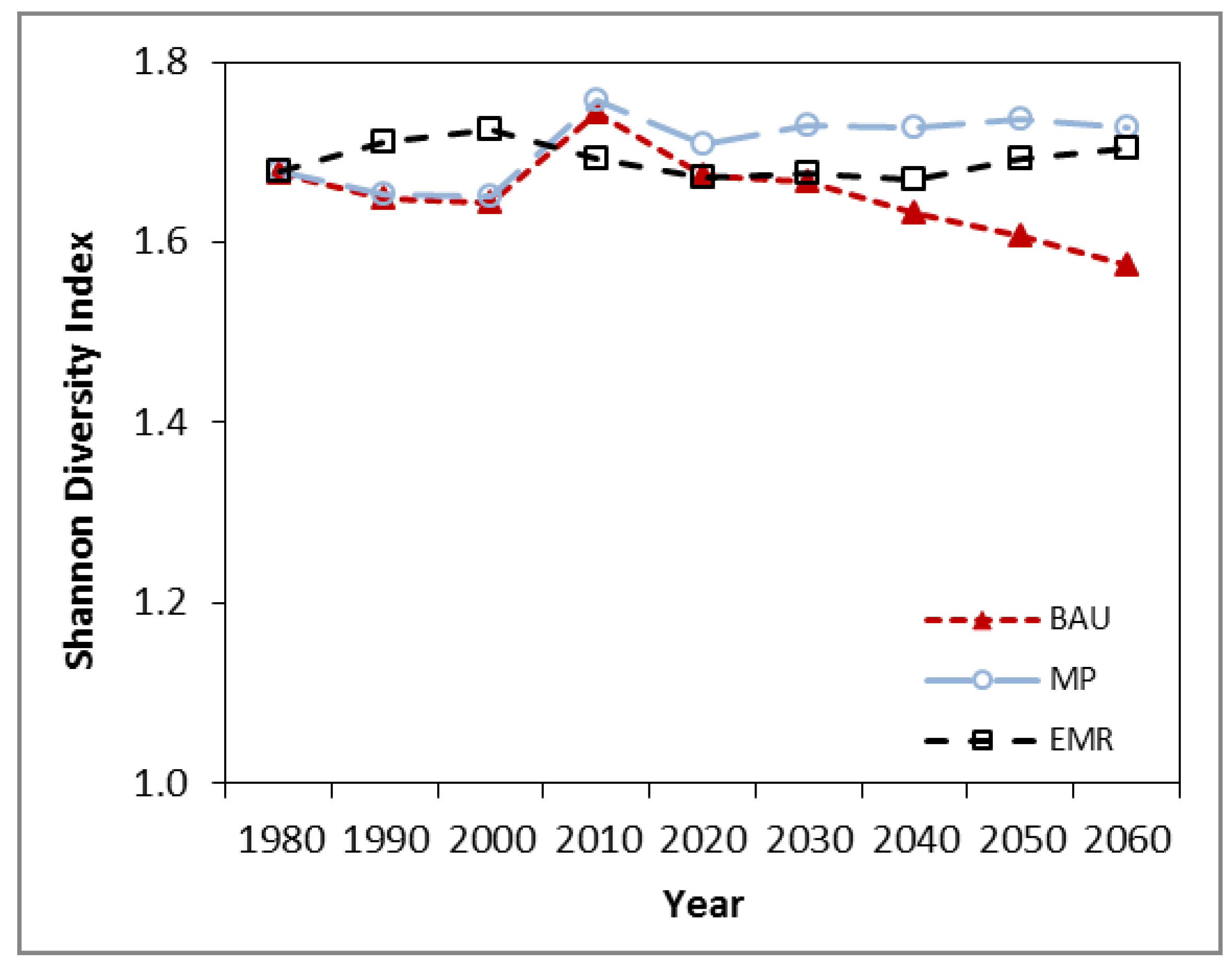
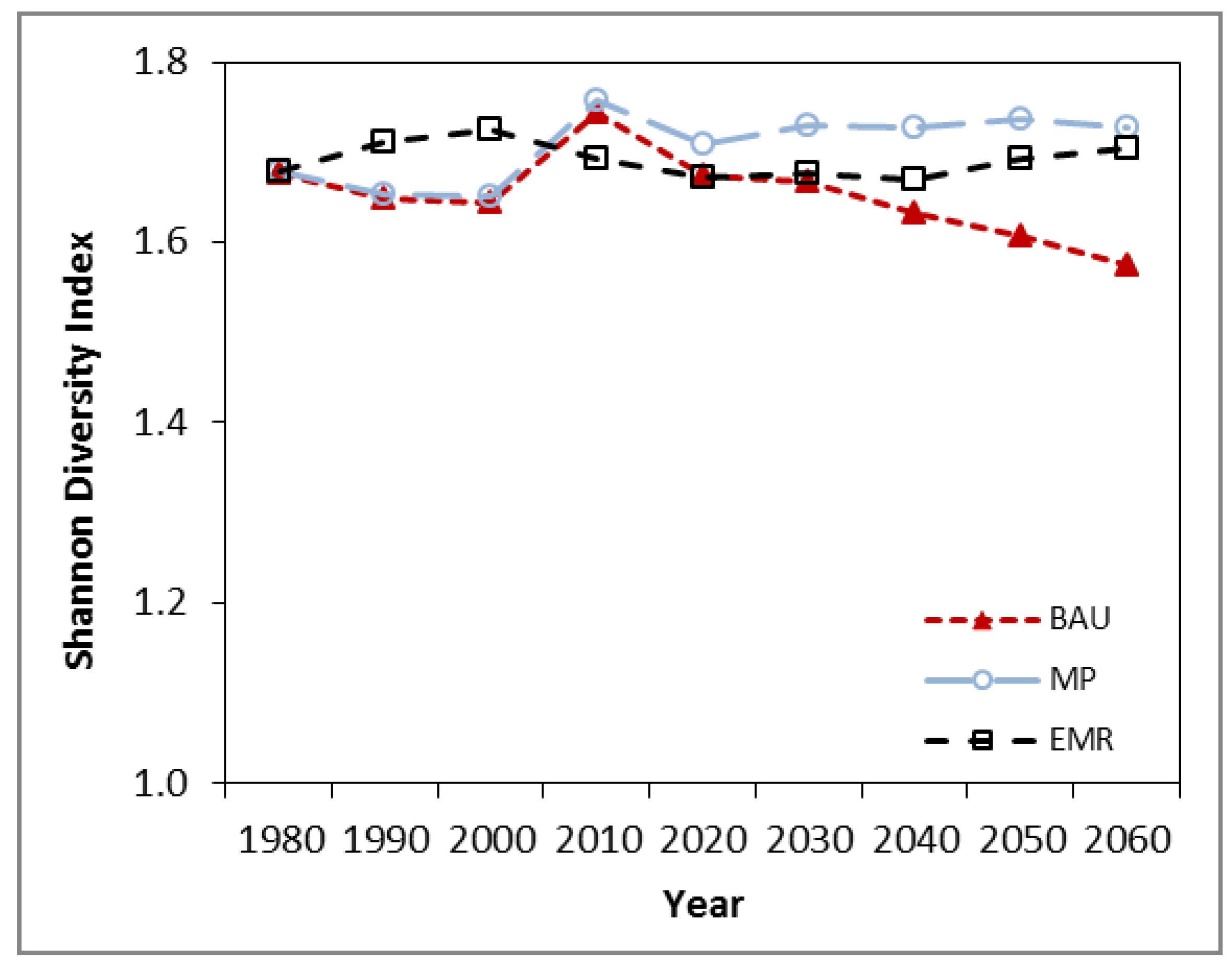
3.2. Diversity
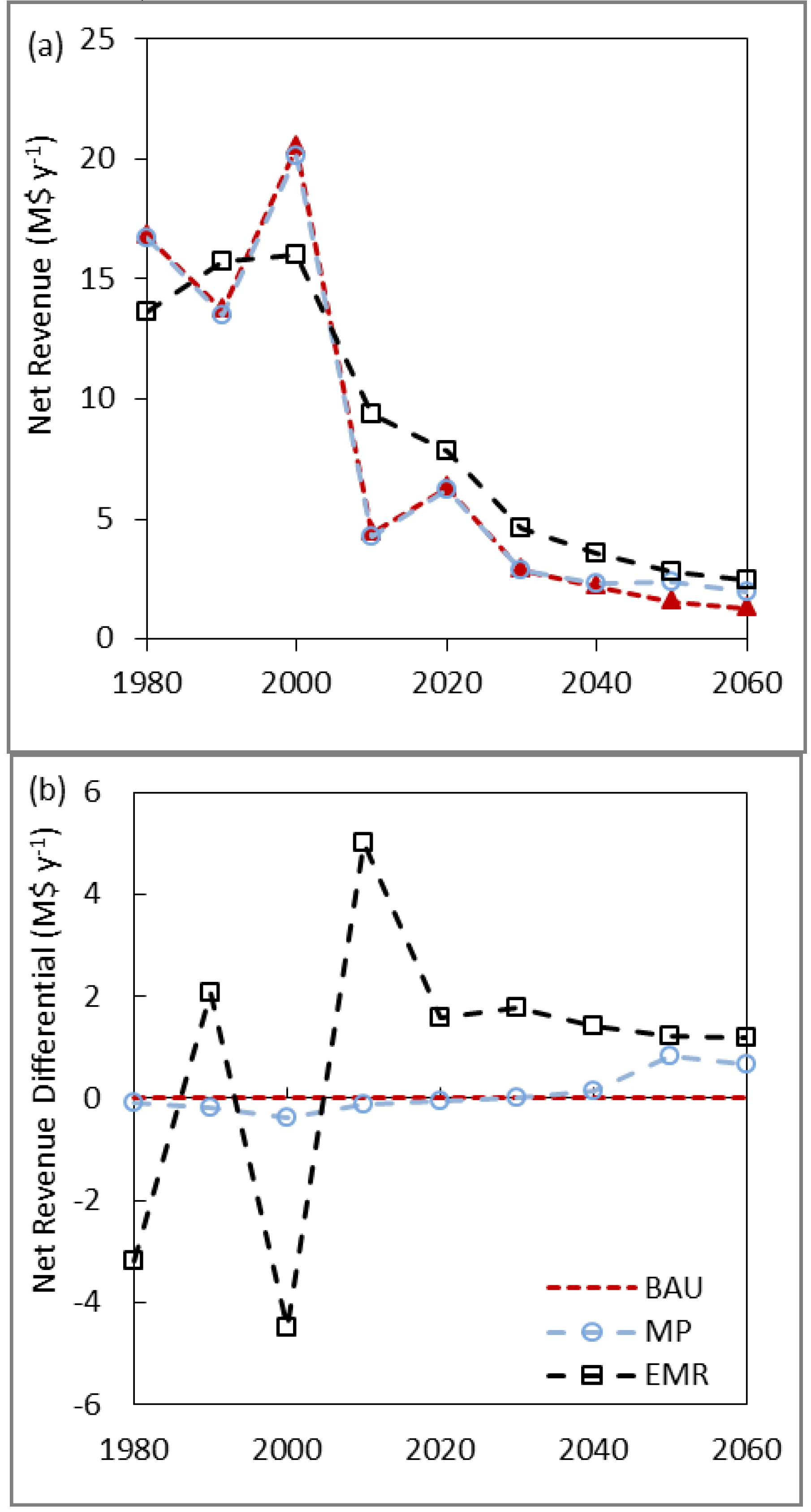
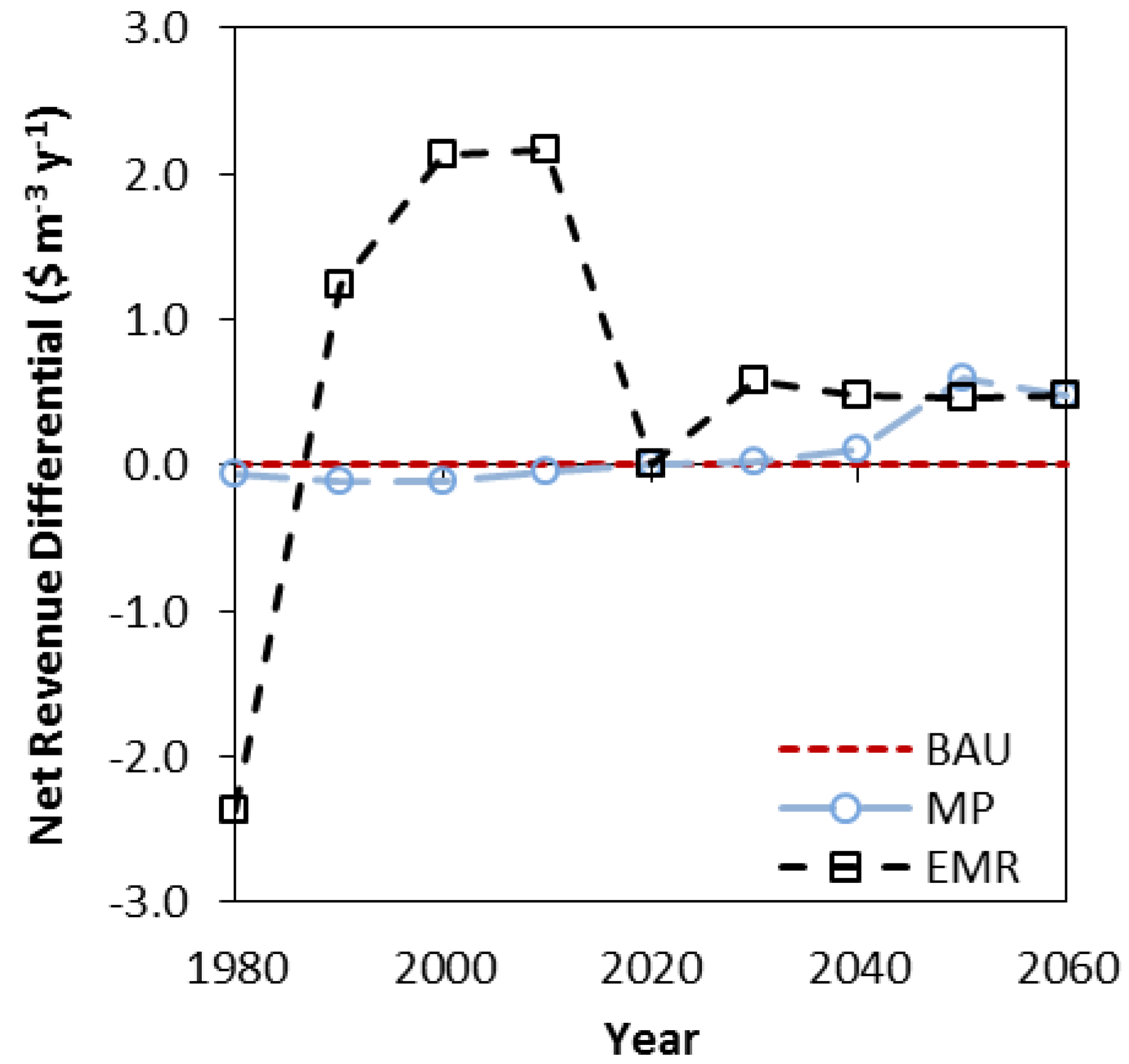
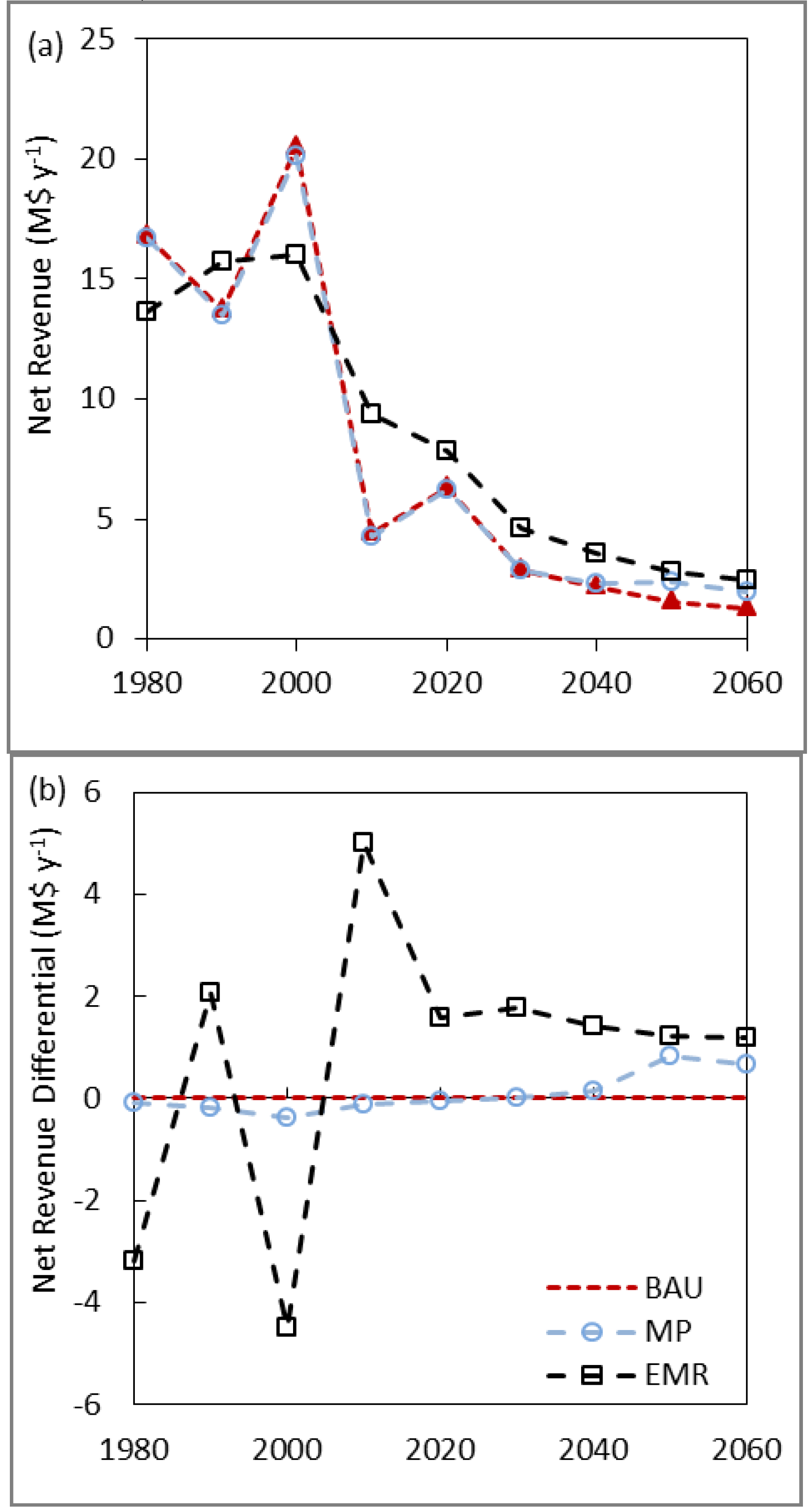
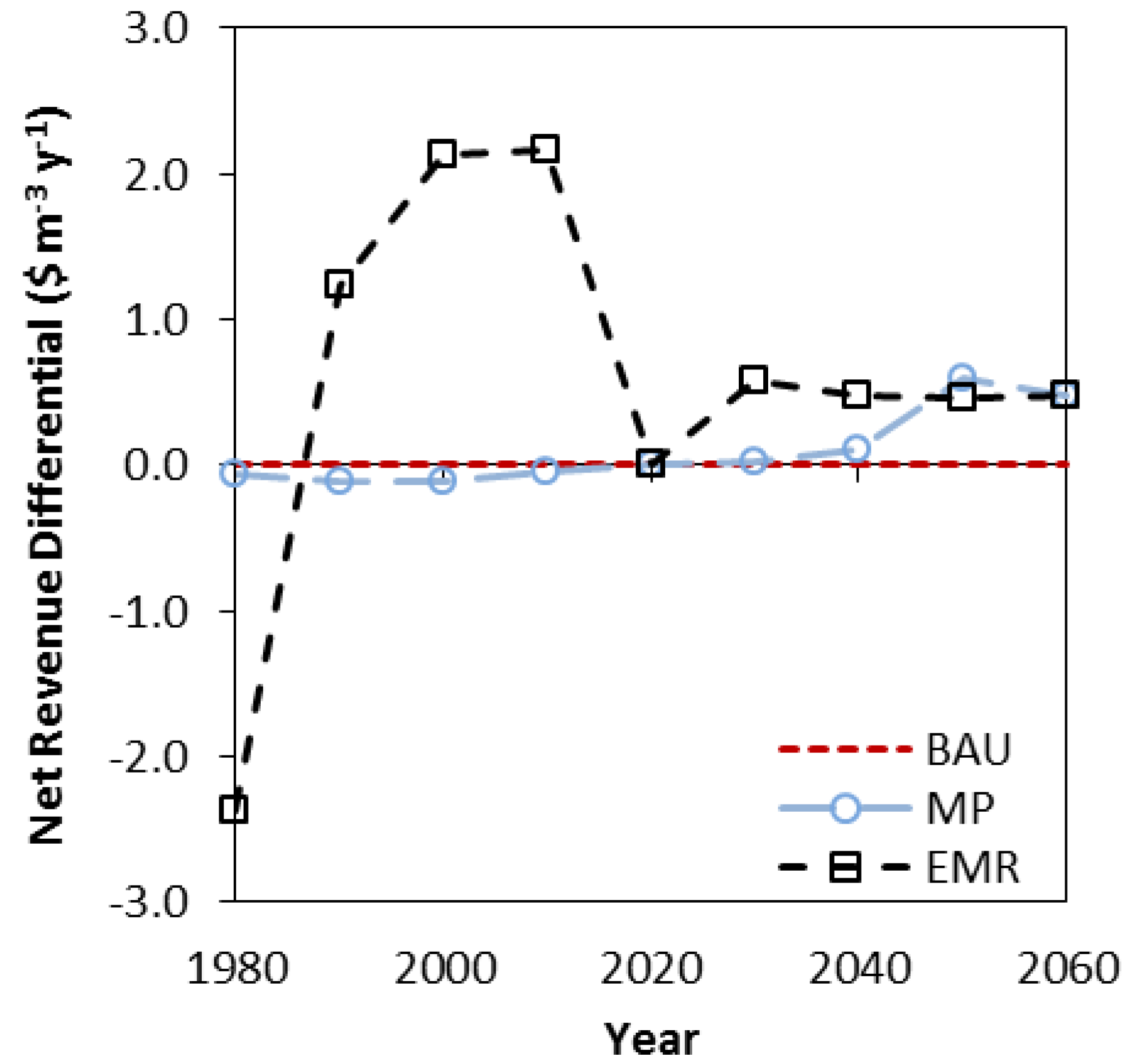
3.3. Net Present Value, Revenues and Costs
| Management Strategy | Discount Rate | |||
|---|---|---|---|---|
| 0% | 1% | 3% | 5% | |
| BAU | 1543 | 1123 | 695 | 498 |
| MP | 1678 | 1182 | 704 | 497 |
| EMR | 2015 | 1372 | 760 | 505 |
| Management Strategy | Discount Rate | |||
|---|---|---|---|---|
| 0% | 1% | 3% | 5% | |
| BAU | 10.41 | 7.57 | 4.68 | 3.36 |
| MP | 11.36 | 8.00 | 4.76 | 3.36 |
| EMR | 12.81 | 8.72 | 4.83 | 3.21 |
| Management Strategy | Decade | |||
|---|---|---|---|---|
| 1980–1989 | 1990–1999 | 2000–2009 | >2010 | |
| BAU | 44.8 | 43.8 | 42.1 | 46.01 |
| MP | 44.9 | 44.0 | 42.3 | 45.97 |
| EMR | 39.5 | 40.4 | 42.3 | 43.99 |
3.4. Uncertainty Assessment
| Douglas-Fir Productivity | BAU | EMR | ||||||
|---|---|---|---|---|---|---|---|---|
| Harvest (Mm3·yr−1) | % from Base | Net Present Value (M$) | % from Base | Harvest (Mm3·yr−1) | % from Base | Net Present Value (M$) | % from Base | |
| −20% | 1.39 | −4.8% | 667 | −4.0 | 1.67 | −8.3% | 713 | −6.2 |
| Base | 1.46 | n/a | 695 | n/a | 1.8 | n/a | 760 | n/a |
| +20% | 1.55 | +6.2% | 729 | 4.9 | 1.96 | +7.7% | 803 | 5.7 |
| Log Price Assumption | THLB | BAU | EMR | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Area (‘000 ha) | % from Base | Harvest (Mm3·year−1) | % from Base | NPV (M$2005) | % from Base | Harvest (Mm3·year−1) | % from Base | NPV (M$2005) | % from Base | |
| −20% | 596 | −43% | 0.67 | −54% | 672 | −3.3 | 0.72 | −56% | 658 | −13.4 |
| Base | 1,051 | n/a | 1.46 | n/a | 695 | n/a | 1.8 | n/a | 760 | n/a |
| +20% | 1,175 | +12% | 1.64 | +13% | 708 | 1.9 | 1.76 | +7% | 828 | 8.9 |
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- IPCC. Summary for Policy Makers. In Climate Change 2014: Impacts, Adaptation and Vulnerability, Working Group II, 5th Assessment Report; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014; p. 44. [Google Scholar]
- Costanza, R.; de Groot, R.; Sutton, P.; van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Fettig, C.J.; Reid, M.L.; Bentz, B.J.; Sevanto, S.; Spittlehouse, D.L.; Wang, T. Changing climates, changing forests: A western North American perspective. J. For. 2013, 111, 214–228. [Google Scholar] [CrossRef]
- Taylor, S.W.; Carroll, A.L.; Alfaro, R.I.; Safranyik, L.; Wilson, B. Forest, climate and mountain pine beetle outbreak dynamics in Western Canada. In The Mountain Pine Beetle: A Synthesis of Biology, Management and Impacts on Lodgepole Pine; Safranyik, L., Wilson, B., Eds.; Natural Resources Canada: Victoria, BC, Canada, 2006; pp. 67–94. [Google Scholar]
- Spittlehouse, D.L. Integrating climate change adaptation into forest management. For. Chron. 2005, 81, 691–695. [Google Scholar] [CrossRef]
- Hallegatte, S. Strategies to adapt to an uncertain climate change. Glob. Environ. Chang. 2009, 19, 240–247. [Google Scholar] [CrossRef]
- Wohlgemuth, T.; Bürgi, M.; Scheidegger, C.; Schütz, M. Dominance reduction of species through disturbance—A proposed management principle for central European forests. For. Ecol. Manag. 2002, 166, 1–15. [Google Scholar] [CrossRef]
- Campbell, E.M.; Saunders, S.C.; Coates, K.D.; Meidinger, D.V.; MacKinnon, A.; O’Neill, G.A.; MacKillop, D.J.; DeLong, S.C.; Morgan, D.G. Ecological Resilience and Complexity: A Theoretical Framework for Understanding and Managing British Columbia’s Forest Ecosystems in a Changing Climate; Technical Report 055; Government of B.C.: Victoria, BC, Canada, 2009. [Google Scholar]
- Puettmann, K.J.; Coates, D.; Messier, C. A Critique of Silviculture: Managing for Complexity; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Cameron, A.D. Building resilience into Sitka Spruce (Picea sitchensis (Bong.) Carr.) forests in Scotland in response to the threat of climate change. Forests 2015, 6, 398–414. [Google Scholar] [CrossRef]
- Windmuller-Campione, M.A.; Long, J.N. If long-term resistance to a spruce beetle epidemic is futile, can silvicultural treatments increase resilience in spruce-fir forests in the Central Rocky mountains. Forests 2015, 6, 1157–1178. [Google Scholar] [CrossRef]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A. Resilience, adaptability and transformability in social-ecological systems. Ecol. Soc. 2004, 9, 9. [Google Scholar]
- Folke, C. Resilience: the emergence of a perspective for social-ecological systems analyses. Glob. Environ. Chang. 2006, 16, 253–267. [Google Scholar] [CrossRef]
- Rist, L.; Moen, J. Sustainability in forest management and a new role for resilience thinking. For. Ecol. Manag. 2013, 310, 416–427. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.A. Resistance and resilience: A conceptual framework for silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar] [CrossRef]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2015. [Google Scholar] [CrossRef]
- Duveneck, M.J.; Scheller, R.M. Measuring and managing resistance and resilience under climate change in northern Great Lake forests (USA). Land. Ecol. 2015, 30, 1–18. [Google Scholar] [CrossRef]
- Dymond, C.C.; Tedder, S.; Spittlehouse, D.L.; Raymer, B.; Hopkins, K.; McCallion, K.; Sandland, J. Diversifying managed forests to increase resilience and adapt to climate change. Can. J. For. Res. 2014, 44, 1196–1205. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Ann. Rev. Ecol. Evol. System. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- Ives, A.R.; Carpenter, S.R. Stability and diversity of ecosystems. Science 2007, 317, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Nat. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Dymond, C.C.; Neilson, E.T.; Stinson, G.; Porter, K.; MacLean, D.A.; Gray, D.R.; Campagna, M.; Kurz, W.A. Future spruce budworm outbreak may create a carbon source in eastern Canadian forests. Ecosystems 2010, 13, 917–931. [Google Scholar] [CrossRef]
- Day, J.K.; Perez, D.M. Reducing uncertainty and risk through forest management planning in British Columbia. For. Ecol. Manag. 2013, 300, 117–124. [Google Scholar] [CrossRef]
- Temperli, C.; Bugmann, H.; Elkin, C. Adaptive management for competing forest goods and services under climate change. Ecolog. Appl. 2012, 22, 2065–2077. [Google Scholar] [CrossRef]
- Steenberg, J.W.N.; Duinker, P.N.; Bush, P.G. Exploring adaptation to climate change in the forests of central Nova Scotia, Canada. For. Ecol. Manag. 2011, 262, 2316–2327. [Google Scholar] [CrossRef]
- Schou, E.; Jacobsen, J.B.; Kristensen, K.L. An economic evaluation of strategies for transforming even-aged into near-natural forestry in a conifer-dominated forest in Denmark. For. Policy Econ. 2012, 20, 89–98. [Google Scholar] [CrossRef]
- Lloyd, D.A.; Angove, K.; Hope, G.D.; Thompson, C. A guide to site identification and interpretation for the Kamloops forest region. In Land Management Handbook 23nd; Government of British Columbia: Victoria, BC, Canada, 1990. [Google Scholar]
- PCIC. Climate Summary of the Thompson-Okanagan Region; Pacific Climate Impacts Consortium: Victoria, BC, Canada, 2014; p. 4. [Google Scholar]
- Sillmann, J.; Kharin, V.V.; Zwiers, F.W.; Zhang, X.; Bronaugh, D. Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J. Geophys. Res.: Atmospheres 2013, 118, 2473–2493. [Google Scholar] [CrossRef]
- Haughian, S.R.; Burton, P.J.; Taylor, S.W.; Curry, C. Expected effects of climate change on forest disturbance regimes in British Columbia. B.C. J. Ecosys. Manag. 2012, 13, 1–24. [Google Scholar]
- Griesbauer, H.P.; Green, D.S.; O’Neill, G.A. Using a spatiotemporal climate model to assess population-level Douglas-fir growth sensitivity to climate change across large climatic gradients in British Columbia, Canada. For. Ecol. Manag. 2011, 261, 589–600. [Google Scholar] [CrossRef]
- Carson, D.M. Timber supply analysis: An industrial model from British Columbia. For. Chron. 1995, 71, 735–738. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Jacquish, B.C. Ecological impacts and management strategies for western larch in the face of climate-change. Mitigat. Adapt. Strat. Glob. Chang. 2010, 15, 283–306. [Google Scholar] [CrossRef]
- Adger, W.N.; Desai, S.; Goulden, M.; Hulme, M.; Lorenzoni, I.; Nelson, D.R.; Naess, L.O.; Wolf, J.; Wreford, A. Are there social limits to adaptation to climate change? Clim. Chang. 2009, 93, 335–354. [Google Scholar] [CrossRef]
- Miller, L.T.; Park, C.S. Decision making under uncertainty—Real options to the rescue? Eng. Econ. 2002, 47, 105–150. [Google Scholar] [CrossRef]
- Roumboutsos, A.B. Sustainability, social discount dates and the selection of project procurement method. Int. Adv. Econ. Res. 2010, 16, 165–174. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- E-Flora B.C.: Electronic Atlas of the Flora of British Columbia; Klinkenberg, B. (Ed.) University of British Columbia: Vancouver, BC, Canada, 2013.
- Schneider, R.R.; Latham, M.C.; Stelfox, B.; Farr, D.; Boutin, S. Effects of a severe mountain pine beetle epidemic in western Alberta, Canada under two forest management strategies. Int. J. For. Res. 2010. [Google Scholar] [CrossRef]
- Hildebrandt, P.; Knoke, T. Investment decisions under uncertainty—A methodological review on forest science studies. For. Policy Econ. 2011, 13, 1–15. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Brooks, J.E.; Hodge, J.C. Analysis of historic western spruce budworm defoliation in south central British Columbia. For. Ecol. Manag. 2006, 226, 351–356. [Google Scholar] [CrossRef]
- Maclauchlan, L.; Brooks, J.E. Influence of past forestry practices on western spruce budworm defoliation and associated impacts in southern British Columbia. B.C. J. Ecosyst. Manag. 2009, 10, 37–49. [Google Scholar]
- Wang, T.; Hamman, A.; Spittlehouse, D.L.; Murdock, T.Q. ClimateWNA—High-resolution spatial climate data for western North America. J. Appl. Meteorol. Climatol. 2012, 51, 16–29. [Google Scholar]
- Crookston, N.L.; Rehfeldt, G.E.; Dixon, G.E.; Weiskittel, A.R. Addressing climate change in the forest vegetation simulator to assess impacts on landscape forest dynamics. For. Ecol. Manag. 2010, 260, 1198–1211. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Smith, A.L.; Hewitt, N.; Klenk, N.; Bazely, D.R.; Yan, N.; Wood, S.; Henriques, I.; MacLellan, J.I.; Lipsig-Mummé, C. Effects of climate change on the distribution of invasive alien species in Canada: A knowledge synthesis of range change projections in a warming world. Environ. Rev. 2012, 20, 1–16. [Google Scholar] [CrossRef]
- Pearson, M.; Healey, M.C. Species at Risk and Local Government: A Primer for B.C. Stewardship Centre of British Columbia: Courtenay, Canada, 2012. Available online: http://www.speciesatrisk.bc.ca/ (accessed on 1 October 2013).
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Hicke, J.A.; Allen, C.D.; Desai, A.R.; Dietze, M.C.; Hall, R.J.; Kashian, D.M.; Moore, D.; Raffa, K.F.; Sturrock, R.N.; Vogelmann, J. Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Glob. Chang. Biol. 2012, 18, 7–34. [Google Scholar] [CrossRef]
- Metsaranta, J.; Dymond, C.C.; Kurz, W.A.; Spittlehouse, D.L. Uncertainty of 21st century growing stocks and GHG balance of forests in British Columbia, Canada resulting from potential climate change impacts on ecosystem processes. For. Ecol. Manag. 2011, 262, 827–837. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dymond, C.C.; Spittlehouse, D.L.; Tedder, S.; Hopkins, K.; McCallion, K.; Sandland, J. Applying Resilience Concepts in Forest Management: A Retrospective Simulation Approach. Forests 2015, 6, 4421-4438. https://doi.org/10.3390/f6124377
Dymond CC, Spittlehouse DL, Tedder S, Hopkins K, McCallion K, Sandland J. Applying Resilience Concepts in Forest Management: A Retrospective Simulation Approach. Forests. 2015; 6(12):4421-4438. https://doi.org/10.3390/f6124377
Chicago/Turabian StyleDymond, Caren C., David L. Spittlehouse, Sinclair Tedder, Katherine Hopkins, Katharine McCallion, and James Sandland. 2015. "Applying Resilience Concepts in Forest Management: A Retrospective Simulation Approach" Forests 6, no. 12: 4421-4438. https://doi.org/10.3390/f6124377
APA StyleDymond, C. C., Spittlehouse, D. L., Tedder, S., Hopkins, K., McCallion, K., & Sandland, J. (2015). Applying Resilience Concepts in Forest Management: A Retrospective Simulation Approach. Forests, 6(12), 4421-4438. https://doi.org/10.3390/f6124377