
Maturation in Corymbia torelliana × C. citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations


Abstract
:1. Introduction
2. Experimental Section
2.1. Stock Plant and Cutting Production
2.2. Hormone Analyses
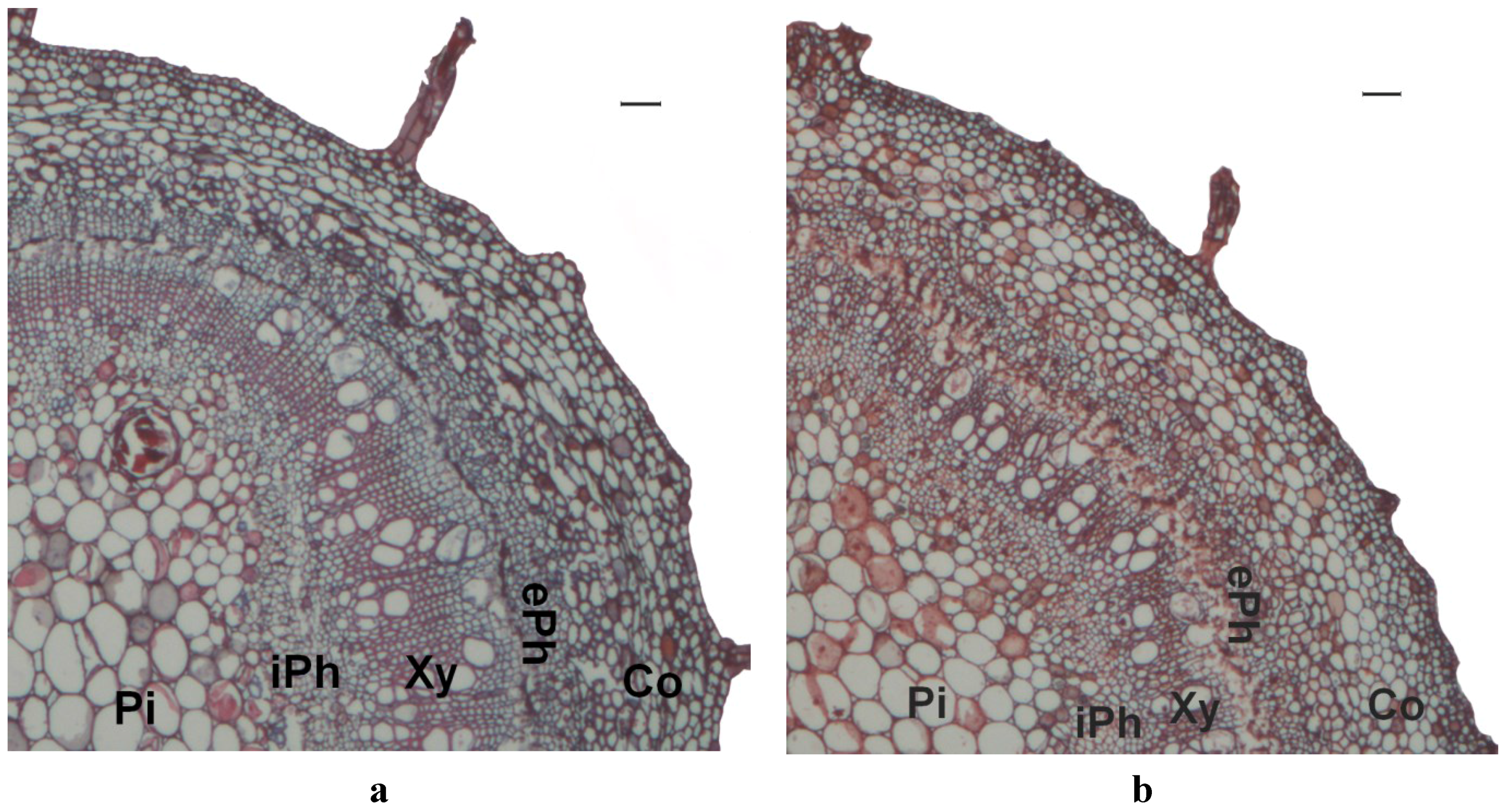
2.3. Stem Anatomy
2.4. Experimental Design and Statistical Analyses
3. Results

{kind=link}
| Clone | Pruning Height | Setting 1 | Setting 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time after Setting (days) | |||||||||||||||
| 36 | 40 | 44 | 48 | 52 | 56 | 60 | 16 | 20 | 24 | 28 | 32 | 36 | |||
| 29 | Tall | 0 a | 0 a | 0 a | 0 b | 4 b | 4 b | 6 b | 0 a | 2 a | 2 a | 4 a | 4 a | 4 b | |
| Short | 0 a | 8 a | 12 a | 18 a | 24 a | 24 a | 26 a | 2 a | 4 a | 8 a | 10 a | 16 a | 22 a | ||
| 67 | Tall | 2 a | 8 a | 10 a | 12 b | 16 b | 16 b | 16 b | 12 a | 14 a | 18 b | 26 a | 34 a | 36 a | |
| Short | 0 a | 10 a | 16 a | 26 a | 32 a | 32 a | 34 a | 20 a | 34 a | 42 a | 42 a | 42 a | 42 a | ||
| 83 | Tall | 0 a | 2 a | 4 a | 6 a | 14 a | 14 a | 14 a | 0 b | 0 b | 14 a | 16 a | 22 a | 22 a | |
| Short | 0 a | 0 a | 6 a | 12 a | 22 a | 22 a | 26 a | 20 a | 22 a | 22 a | 26 a | 26 a | 26 a | ||
| 156 | Tall | 4 a | 6 a | 22 a | 40 a | 50 a | 50 a | 54 a | 0 b | 2 b | 12 b | 16 b | 26 b | 32 b | |
| Short | 6 a | 12 a | 22 a | 30 a | 36 a | 36 a | 42 a | 30 a | 58 a | 68 a | 72 a | 74 a | 74 a | ||
| 160 | Tall | 0 a | 0 a | 0 b | 0 b | 4 b | 4 b | 4 b | 4 b | 4 b | 10 b | 10 b | 10 b | 18 b | |
| Short | 6 a | 10 a | 14 a | 22 a | 28 a | 28 a | 28 a | 20 a | 40 a | 40 a | 44 a | 50 a | 50 a | ||
| 163 | Tall | 4 a | 12 a | 26 a | 28 a | 40 a | 40 a | 40 a | 18 a | 28 b | 30 b | 34 b | 34 b | 34 a | |
| Short | 8 a | 16 a | 30 a | 40 a | 58 a | 58 a | 60 a | 34 a | 48 a | 48 a | 52 a | 54 a | 54 a | ||
| Clone | Pruning Height | Setting 1 | Setting 2 |
|---|---|---|---|
| 29 | Tall | 6 ± 2 Db | 8 ± 4 Db |
| Short | 26 ± 5 Ca | 22 ± 4 Ca | |
| 67 | Tall | 20 ± 4 ABb | 40 ± 3 ABb |
| Short | 54 ± 9 Ba | 54 ± 2 Ba | |
| 83 | Tall | 20 ± 3 BCa | 28 ± 5 BCa |
| Short | 34 ± 7 Ca | 36 ± 2 Ca | |
| 156 | Tall | 74 ± 5 Ab | 54 ± 7 Ab |
| Short | 82 ± 4 Aa | 86 ± 2 Aa | |
| 160 | Tall | 6 ± 2 CDb | 17 ± 1 CDb |
| Short | 50 ± 4 Ba | 60 ± 3 Ba | |
| 163 | Tall | 54 ± 7 Bb | 36 ± 2 Bb |
| Short | 74 ± 6 Ba | 56 ± 5 Ba | |
| Mean | Tall | 30.0 | 30.5 |
| Short | 53.3 | 52.3 |
| Clone | Pruning Height | Setting 1 | Setting 2 |
|---|---|---|---|
| 29 | Tall | 0.30 ± 0.01 Aa | 0.16 ± 0.00 Ba |
| Short | 0.24 ± 0.02 Ab | 0.15 ± 0.01 Aa | |
| 67 | Tall | 0.22 ± 0.04 Aa | 0.17 ± 0.01 Ba |
| Short | 0.21 ± 0.02 Aa | 0.20 ± 0.04 Aa | |
| 83 | Tall | 0.24 ± 0.02 Aa | 0.24 ± 0.03Aba |
| Short | 0.23 ± 0.03 Aa | 0.22 ± 0.03 Aa | |
| 156 | Tall | 0.24 ± 0.03 Aa | 0.19 ± 0.02 Ba |
| Short | 0.23 ± 0.02 Aa | 0.24 ± 0.04 Aa | |
| 160 | Tall | 0.31 ± 0.03 Aa | 0.30 ± 0.03 Aa |
| Short | 0.18 ± 0.03 Ab | 0.28 ± 0.04 Aa | |
| 163 | Tall | 0.23 ± 0.02 Aa | 0.20 ± 0.02 Ba |
| Short | 0.20 ± 0.02 Aa | 0.19 ± 0.00 Aa | |
| Mean | Tall | 0.26 | 0.21 |
| Short | 0.21 | 0.21 |
| Clone | Pruning Height | Collection | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | Mean | ||
| 29 | Tall | 243 a | 245 a | 298 a | 354 b | 658 a | 548 b | 919 a | 589 a | 897 a | 851 a | 858 a | 1,170 a | 560 |
| Short | 177 b | 226 a | 201 b | 533 a | 423 b | 699 a | 533 b | 741 a | 780 a | 824 a | 692 b | 1,160 a | 514 | |
| 67 | Tall | 259 a | 440 a | 255 a | 642 a | 580 a | 733 a | 793 a | 605 a | 917 a | 1,081 a | 1,190 a | 1,277 a | 631 |
| Short | 141 b | 209 b | 154 b | 488 b | 474 b | 499 b | 676 a | 565 a | 680 b | 648 b | 903 b | 1,070 b | 453 | |
| 83 | Tall | 141 b | 412 a | 252 a | 660 a | 557 a | 627 a | 867 a | 676 a | 728 a | 922 a | 1,248 a | 1,307 a | 584 |
| Short | 185 a | 245 b | 217 a | 631 a | 460 a | 695 a | 559 b | 741 a | 780 a | 1,010 a | 917 b | 936 b | 552 | |
| 156 | Tall | 262 a | 348 a | 293 a | 677 a | 571 a | 790 a | 787 a | 646 a | 713 a | 891 a | 925 a | 1,426 a | 598 |
| Short | 280 a | 256 a | 192 b | 568 a | 562 a | 483 b | 650 a | 566 a | 614 a | 895 a | 644 b | 1,151 b | 507 | |
| 160 | Tall | 151 b | 226 a | 142 b | 373 a | 418 a | 421 a | 426 b | 478 a | 577 a | 425 b | 686 a | 655 b | 364 |
| Short | 190 a | 213 a | 226 a | 433 a | 324 a | 494 a | 594 a | 468 a | 669 a | 770 a | 657 a | 869 a | 438 | |
| 163 | Tall | 277 a | 323 a | 396 a | 505 a | 761 a | 916 a | 923 a | 813 a | 1,073 a | 1,108 a | 1,112 a | 1,433 a | 709 |
| Short | 151 b | 153 b | 230 b | 459 a | 450b | 431 b | 713 a | 646 a | 992 a | 800 b | 1,125 a | 1,237 a | 503 | |
| Mean | Tall | 222 | 332 | 273 | 535 | 591 | 672 | 786 | 635 | 817 | 880 | 1,003 | 1,211 | 574 |
| Short | 187 | 217 | 203 | 518 | 449 | 550 | 621 | 621 | 752 | 825 | 823 | 1,070 | 494 | |
| Clone | Pruning Height | Setting 1 | Setting 2 |
|---|---|---|---|
| 29 | Tall | 14.7 b | 93.6 a |
| Short | 58.7 a | 255.3 a | |
| 67 | Tall | 88.0 a | 510.9 a |
| Short | 113.4 a | 427.9 a | |
| 83 | Tall | 82.5 a | 365.8 a |
| Short | 83.3 a | 337.0 a | |
| 156 | Tall | 257.6 a | 770.2 a |
| Short | 210.1 a | 989.4 a | |
| 160 | Tall | 13.5 b | 112.0 b |
| Short | 106.6 a | 521.5 a | |
| 163 | Tall | 174.5 a | 516.0 a |
| Short | 113.4 a | 692.6 a | |
| Mean | Tall | 105.1 | 394.7 |
| Short | 114.2 | 537.2 |
| Clone | Pruning Height | Setting 1 | Setting 2 | ||
|---|---|---|---|---|---|
| Free IAA (ng·g−1 DM) | ABA (ng·g−1 DM) | Free IAA (ng·g−1 DM) | ABA (ng·g−1 DM) | ||
| 29 | Tall | 657 ± 31 ABa | 153 ± 3 CDa | 305 ± 9 BCa | 130 ± 8 BCa |
| Short | 414 ± 11 Cb | 151 ± 5 Ba | 294 ± 9 Da | 168 ± 24 Da | |
| 67 | Tall | 605 ± 28 Ba | 342 ± 24 Aa | 369 ± 27 ABa | 329 ± 12 ABa |
| Short | 624 ± 27 ABa | 195 ± 23 Ba | 445 ± 18 Aa | 225 ± 27 Ab | |
| 83 | Tall | 771 ± 26 Aa | 282 ± 26 ABa | 243 ± 11 CDb | 241 ± 24 CDa |
| Short | 599 ± 13 ABb | 149 ± 6 Bb | 396 ± 10 BCa | 166 ± 15 BCa | |
| 156 | Tall | 741 ± 5 ABa | 330 ± 30 Aa | 247 ± 16 CDb | 99 ± 8 CDb |
| Short | 668 ± 22 Ab | 345 ± 41 Aa | 360 ± 2 Ca | 267 ± 29 Ca | |
| 160 | Tall | 331 ± 40 Ca | 88 ± 4 Db | 400 ± 10 Aa | 169 ± 20 Aa |
| Short | 361 ± 38 Ca | 130 ± 3 Ba | 388 ± 5 BCa | 138 ± 9 BCa | |
| 163 | Tall | 448 ± 32 Ca | 197 ± 11 BCa | 224 ± 14 Db | 134 ± 2 Db |
| Short | 495 ± 45 BCa | 208 ± 5 Ba | 413 ± 9 ABa | 257 ± 4 ABa | |
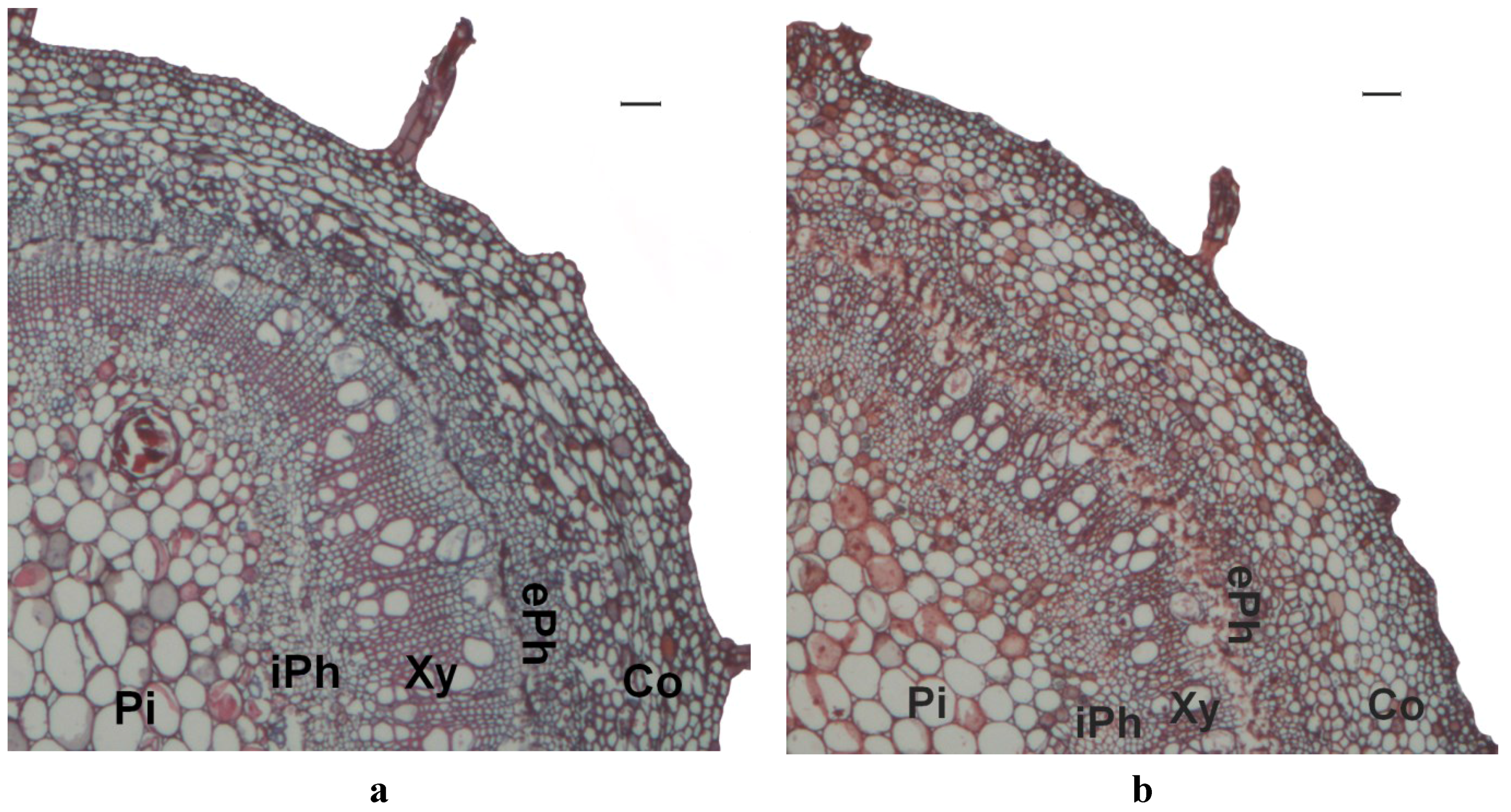
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Xavier, A.; Wendling, I.; Silva, R.L. Silvicultura Clonal—Princípios e Técnicas; Editora UFV: Viçosa, Brazil, 2013. [Google Scholar]
- Assis, T.F.; Fett-Neto, A.G.; Alfenas, A.C. Current techniques and prospects for the clonal propagation of hardwoods with emphasis on Eucalyptus. In Plantation Forest Biotechnology for the 21st Century; Walter, C., Carson, M., Eds.; Research Signpost: Kerala, India, 2004; pp. 303–333. [Google Scholar]
- Bettinger, P.; Clutter, M.; Siry, J.; Kane, M.; Pait, J. Broad implications of southern United States pine clonal forestry on planning and management of forests. Int. For. Rev. 2009, 11, 331–345. [Google Scholar] [CrossRef]
- Greenwood, M.S.; Hutchinson, K.W. Maturation as a developmental process. In Clonal Forestry I: Genetics and Biotechnology; Ahuja, M.R., Libby, W.J., Eds.; Springer-Verlag: Berlin, Germany, 1993; pp. 14–33. [Google Scholar]
- Mitchell, R.G.; Zwolinski, J.; Jones, N.B. A review on the effects of donor maturation on rooting and field performance of conifer cuttings. South. Afr. For. J. 2004, 201, 51–63. [Google Scholar] [CrossRef]
- McMahon, T.V.; Hung, C.D.; Trueman, S.J. In vitro storage delays the maturation of African mahogany (Khaya senegalensis) clones. J. Plant Sci. 2013, 8, 31–38. [Google Scholar]
- Pijut, P.M.; Woeste, K.E.; Michler, C.H. Promotion of adventitious root formation of difficult-to-root hardwood tree species. Hortic. Rev. 2011, 38, 213–251. [Google Scholar]
- Husen, A. Changes of soluble sugars and enzymatic activities during adventitious rooting in cuttings of Grewia optiva as affected by age of donor plants and auxin treatments. Am. J. Plant Physiol. 2012, 7, 1–16. [Google Scholar] [CrossRef]
- Wendling, I.; Trueman, S.J.; Xavier, A. Maturation and related aspects in clonal forestry—Part I: Concepts, regulation and consequences of phase change. New For. 2014, 45, 449–471. [Google Scholar] [CrossRef]
- Wendling, I.; Trueman, S.J.; Xavier, A. Maturation and related aspects in clonal forestry—Part II: Reinvigoration, rejuvenation and juvenility maintenance. New For. 2014, 45, 473–486. [Google Scholar] [CrossRef]
- Aimers-Halliday, J.; Menzies, M.I.; Faulds, T.; Holden, D.G.; Low, C.B.; Dibley, M.J. Nursery systems to control maturation in Pinus radiata cuttings, comparing hedging and serial propagation. N. Z. J. For. Sci. 2003, 33, 135–155. [Google Scholar]
- Trueman, S.J. Clonal propagation and storage of subtropical pines in Queensland, Australia. South. Afr. For. J. 2006, 208, 49–52. [Google Scholar] [CrossRef]
- Abu-Abied, M.; Szwerdszarf, D.; Mordehaev, I.; Levy, A.; Rogovoy, O.; Belausov, E.; Yaniv, Y.; Uliel, S.; Katzenellenbogen, M.; Riov, J.; et al. Microarray analysis revealed upregulation of nitrate reductase in juvenile cuttings of Eucalyptus grandis, which correlated with increased nitric oxide production and adventitious root formation. Plant J. 2012, 71, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.D.; Trueman, S.J. Topophysic effects differ between node and organogenic cultures of the eucalypt Corymbia torelliana × C. citriodora. Plant Cell Tissue Organ Cult. 2011, 104, 69–77. [Google Scholar] [CrossRef]
- Davies, P.J. The plant hormones: Their nature, occurrence, and functions. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar]
- Costa, C.T.; de Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.S.; Fett-Neto, A.G. When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.W.; Yang, Y.W.; Lur, H.S.; Tsai, Y.G.; Chang, M.C. A novel function of abscisic acid in the regulation of rice (Oryza sativa) root growth and development. Plant Cell Physiol. 2006, 47, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mwange, W.N.; Hou, H.W.; Cui, K.M. Relationship between endogenous indole-3-acetic acid and abscisic acid changes and bark recovery in Eucommia ulmoides Oliv. after girding. J. Exp. Bot. 2003, 54, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Materán, M.E.; Fernández, M.; Valenzuela, S.; Sáez, K.; Seemann, P.; Sánchez-Olate, M.; Ríos, D. Abscisic acid and 3-indolacetic acid levels during the reinvigoration process of Pinus radiata D. Don adult material. Plant Growth Regul. 2009, 59, 171–177. [Google Scholar] [CrossRef]
- Wendling, I.; Brooks, P.R.; Trueman, S.J. Topophysis in Corymbia torelliana × C. citriodora seedlings: Adventitious rooting capacity, stem anatomy, and auxin and abscisic acid concentrations. New For. 2015, 46, 107–120. [Google Scholar] [CrossRef]
- Trueman, S.J.; Adkins, M.F. Effect of aminoethoxyvinylglycine and 1-methylcyclopropene on leaf abscission and root formation in Corymbia and Eucalyptus cuttings. Sci. Hortic. 2013, 161, 1–7. [Google Scholar] [CrossRef]
- Trueman, S.J.; McMahon, T.V.; Bristow, M. Production of cuttings in response to stock plant temperature in the subtropical eucalypts, Corymbia citriodora and Eucalyptus dunnii. New For. 2013, 44, 265–279. [Google Scholar] [CrossRef]
- Trueman, S.J.; McMahon, T.V.; Bristow, M. Production of Eucalyptus cloeziana cuttings in response to stock plant temperature. J. Trop. For. Sci. 2013, 25, 60–69. [Google Scholar]
- Bryant, P.H.; Trueman, S.J. Stem anatomy and adventitious root formation in cuttings of Angophora, Corymbia and Eucalyptus. Forests 2015, 6, 1227–1238. [Google Scholar] [CrossRef]
- Titon, M.; Xavier, A.; Otoni, W.C. Clonal propagation of Eucalyptus grandis using the mini-cutting and micro-cutting techniques. Sci. For. 2006, 71, 109–117. [Google Scholar]
- Rosa, L.S.; Wendling, I.; Grossi, F.; Reissmann, C.B. Effect of the nitrogen dose and substrate formulations on the minicutting technique of Eucalyptus dunnii Maiden. Rev. Árv. 2009, 33, 1025–1035. [Google Scholar]
- Brondani, G.E.; Grossi, F.; Wendling, I.; Dutra, L.F.; Araujo, M.A. Eucalyptus benthamii × Eucalyptus dunnii mini-cutting technique: (II) mini-cutting survival and rooting in relation to collection and seasons. Ciência Flor. 2010, 20, 453–465. [Google Scholar]
- Brondani, G.E.; Wendling, I.; Grossi, F.; Dutra, L.F.; de Araujo, M.A. Miniestaquia de Eucalyptus benthamii × Eucalyptus dunnii: (I) sobrevivência de minicepas e produção de miniestacas em função das coletas e estações do ano. Ciência Flor. 2012, 22, 11–21. [Google Scholar] [CrossRef]
- Hung, C.D.; Trueman, S.J. Nutrient responses differ between node and organogenic cultures of Corymbia torelliana × C. citriodora (Myrtaceae). Aust. J. Bot. 2010, 58, 410–419. [Google Scholar] [CrossRef]
- McMahon, T.V.; Hung, C.D.; Trueman, S.J. Clonal maturation of Corymbia torelliana × C. citriodora is delayed by minimal-growth storage. Aust. For. 2014, 77, 9–14. [Google Scholar] [CrossRef]
- Kelen, M.; Demiralay, E.C.; Sen, S.; Ozkan, G. Separation of abscisic acid, indole-3-acetic acid, gibberellic acid in 99 R (Vitis berlandieri × Vitis rupestris) and rose oil (Rosa damascena Mill.) by reversed phase liquid chromatography. Turk. J. Chem. 2004, 28, 603–610. [Google Scholar]
- Kilkenny, A.J.; Wallace, H.M.; Walton, D.A.; Adkins, M.F.; Trueman, S.J. Improved root formation in eucalypt cuttings following combined auxin and anti-ethylene treatments. J. Plant Sci. 2012, 7, 138–153. [Google Scholar]
- Pohio, K.E.; Wallace, H.M.; Peters, R.F.; Smith, T.E.; Trueman, S.J. Cuttings of Wollemi pine tolerate moderate photoinhibition and remain highly capable of root formation. Trees 2005, 19, 587–595. [Google Scholar] [CrossRef]
- Trueman, S.J.; Richardson, D.M. Relationships between indole-3-butyric acid, photoinhibition and adventitious rooting of Corymbia torelliana, C. citriodora and F1 hybrid cuttings. Tree For. Sci. Biotechnol. 2008, 2, 26–33. [Google Scholar]
- Husen, A.; Pal, M. Effect of branch position and auxin treatment on clonal propagation of Tectona grandis Linn. f. New For. 2007, 34, 223–233. [Google Scholar] [CrossRef]
- Tchoundjeu, Z.; Leakey, R.R.B. Vegetative propagation of African mahogany: Effects of auxin, node position, leaf area and cutting length. New For. 1996, 11, 125–136. [Google Scholar] [CrossRef]
- Husen, A.; Pal, M. Effect of serial bud grafting and etiolation on rejuvenation and rooting of mature trees of Tectona grandis Linn. f. Silvae Genet. 2003, 52, 84–88. [Google Scholar]
- Husen, A.; Pal, M. Variation in shoot anatomy and rooting behaviour of stem cuttings in relation to age of donor plants in teak (Tectona grandis Linn. f.). New For. 2006, 31, 57–73. [Google Scholar] [CrossRef]
- Haines, R.; Walker, S. Maturation questions relating to clonal forestry. In Clonal Forestry Workshop; QFRI: Gympie, Australia, 1993; p. 13. [Google Scholar]
- Bon, M.C.; Riccardi, F.; Monteuuis, O. Influence of phase change within a 90-year-old Sequoia sempervirens on its in vitro organogenic capacity and protein patterns. Trees 1994, 8, 283–287. [Google Scholar] [CrossRef]
- Bitencourt, J.; Ribas, K.C.Z.; Wendling, I.; Koeler, H. Enraizamento de estacas de erva-mate (Ilex paraguariensis A. St.-Hill.) provenientes de brotações rejuvenescidas. Rev. Bras. Plant. Med. 2009, 11, 277–281. [Google Scholar]
- Hackett, W.P.; Murray, J.R. Maturation and rejuvenation in woody species. In Micropropagation of Woody Plants; Ahuja, M.R., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 93–105. [Google Scholar]
- Von Aderkas, P.; Bonga, J.M. Influencing micropropagation and somatic embryogenesis in mature trees by manipulation of phase change, stress and culture environment. Tree Physiol. 2000, 20, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T., Jr.; Geneve, R.L. Plant Propagation: Principles and Practices; Prentice-Hall: Upper Saddle River, NJ, USA, 2011. [Google Scholar]
- Osterc, G.; Štefančič, M.; Štampar, F. Juvenile stockplant material enhances root development through higher endogenous auxin level. Acta Physiol. Plant. 2009, 31, 899–903. [Google Scholar] [CrossRef]
- Osterc, G.; Štampar, F. Differences in endo/exogenous auxin profile in cuttings of different physiological ages. J. Plant Physiol. 2011, 168, 2088–2092. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.R.; Bastiani, D.; Gaeta, M.L.; Mariath, J.E.A.; Costa, F.; Retallick, J.; Nolan, L.; Tai, H.H.; Strömvik, M.V.; Fett-Neto, A.G. Comparative transcriptional analysis provides new insights into the molecular basis of adventitious rooting recalcitrance in Eucalyptus. Plant. Sci. 2015, 239, 155–165. [Google Scholar] [PubMed]
- Ford, Y.Y.; Bonham, E.C.; Cameron, R.W.F.; Blake, P.S.; Judd, H.L.; Harrison-Murray, R.S. Adventitious rooting: Examining the role of auxin in an easy-and a difficult-to-root plant. Plant Growth Regul. 2002, 36, 149–159. [Google Scholar] [CrossRef]
- Tsipouridis, C.; Thomidis, T.; Bladenopoulou, S. Rhizogenesis of GF677, Early Crest, May Crest and Arm King stem cuttings during the year in relation to carbohydrate and natural hormone content. Sci. Hortic. 2006, 108, 200–204. [Google Scholar] [CrossRef]
- Negishi, N.; Nakahama, K.; Urata, N.; Kojima, M.; Sakakibara, H.; Kawaoka, A. Hormone level analysis on adventitious root formation in Eucalyptus globulus. New For. 2014, 45, 577–587. [Google Scholar] [CrossRef]
- Stewart, J.L.; Nemhauser, J.L. Do trees grow on money? Auxin as the currency of the cellular economy. Cold Spring Harb. Perspect. Biol. 2010, 2, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Maynard, B.K.; Bassuk, N.L. Effects of stock plant etiolation, shading, banding, and shoot development on histology and cutting propagation of Carpinus betulus L. fastigiata. J. Am. Soc. Hortic. Sci. 1996, 121, 853–860. [Google Scholar]
- Pacholczak, A.; Szydło, W.; Łukaszewska, A. The effect of shading of stock plants on rhizogenesis in stem cuttings of Berberis thunbergii “Red Rocket”. Acta Physiol. Plant. 2006, 28, 567–575. [Google Scholar] [CrossRef]
- Amissah, J.N.; Paolillo, D.J., Jr.; Bassuk, N. Adventitious root formation in stem cuttings of Quercus bicolor and Quercus macrocarpa and its relationship to stem anatomy. J. Am. Soc. Hortic. Sci. 2008, 133, 479–486. [Google Scholar]
- Bonga, J.M.; von Aderkas, P. Rejuvenation of tissues from mature conifers and its implications for propagation in vitro. In Clonal Forestry I, Genetics and Biotechnology; Ahuja, M.R., Libby, W.J., Eds.; Springer-Verlag: Berlin, Germany, 1993; pp. 182–199. [Google Scholar]
- Hamann, A. Adventitious root formation in cuttings of loblolly pine (Pinus taeda L.): Developmental sequence and effects of maturation. Trees 1998, 12, 175–180. [Google Scholar] [CrossRef]
- Russel, J.H. Clonal forestry with yellow cedar. In Clonal Forestry II, Conservation and Application; Ahuja, M.R., Libby, W.J., Eds.; Springer-Verlag: Berlin, Germany, 1993; pp. 188–201. [Google Scholar]
- Ritchie, G.A. The commercial use of conifer rooted cuttings in forestry: A world overview. New For. 1991, 5, 247–275. [Google Scholar] [CrossRef]
- McGranahan, M.F.; Boralho, N.M.G.; Greaves, B.L. Genetic control of propagation effects and the importance of stock plant age and source on early growth in cuttings of Pinus radiata. Silvae Genet. 1999, 48, 267–272. [Google Scholar]
- Mitchell, R.G.; Zwolinski, J.; Jones, N.B. The effects of ontogenetic maturation in Pinus patula—Part I: Nursery performance. South. Afr. For. J. 2004, 202, 29–36. [Google Scholar]
- Wendling, I.; Xavier, A.; Paiva, H.D. Influência da miniestaquia seriada no vigor de minicepas de clones de Eucalyptus grandis. Rev. Árv. 2003, 27, 611–618. [Google Scholar] [CrossRef]
- Wendling, I.; Dutra, L.F.; Grossi, F. Produção e sobrevivência de miniestacas e minicepas de erva-mate cultivadas em sistema semi-hidropônico. Pesq. Agropec. Bras. 2007, 42, 289–292. [Google Scholar] [CrossRef]
- Da Cunha, A.C.M.C.M.; Wendling, I.; Júnior, L.S. Produtividade e sobrevivência de minicepas de Eucalyptus benthamii Maiden et Cambage em sistema de hidroponia e em tubete. Ciência Flor. 2005, 15, 307–310. [Google Scholar]
- Majada, J.; Martínez-Alonso, C.; Feito, I.; Kidelman, A.; Aranda, I.; Alía, R. Mini-cuttings: An effective technique for the propagation of Pinus pinaster Ait. New For. 2011, 41, 399–412. [Google Scholar] [CrossRef]
- Da Cunha, A.C.M.C.M.; Paiva, H.N.; Leite, H.G.; Barros, N.F.; Leite, F.P. Relações entre variáveis climáticas com produção e enraizamento de miniestacias de eucalipto. Rev. Árv. 2009, 33, 195–203. [Google Scholar]
- Trueman, S.J.; McMahon, T.V.; Bristow, M. Biomass partitioning in Corymbia citriodora, Eucalyptus cloeziana and E. dunnii stock plants in response to temperature. J. Trop. For. Sci. 2013, 5, 504–509. [Google Scholar]
- Hunt, M.A.; Trueman, S.J.; Rasmussen, A. Indole-3-butyric acid accelerates adventitious root formation and impedes shoot growth of Pinus elliottii var. elliottii × P. caribaea var. hondurensis cuttings. New For. 2011, 41, 349–360. [Google Scholar]
- Hung, C.D.; Trueman, S.J. Alginate encapsulation of shoot tips and nodal segments for short-term storage and distribution of the eucalypt Corymbia torelliana × C. citriodora. Acta Physiol. Plant. 2012, 34, 117–128. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wendling, I.; Warburton, P.M.; Trueman, S.J. Maturation in Corymbia torelliana × C. citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations. Forests 2015, 6, 3763-3778. https://doi.org/10.3390/f6103763
Wendling I, Warburton PM, Trueman SJ. Maturation in Corymbia torelliana × C. citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations. Forests. 2015; 6(10):3763-3778. https://doi.org/10.3390/f6103763
Chicago/Turabian StyleWendling, Ivar, Paul M. Warburton, and Stephen J. Trueman. 2015. "Maturation in Corymbia torelliana × C. citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations" Forests 6, no. 10: 3763-3778. https://doi.org/10.3390/f6103763
APA StyleWendling, I., Warburton, P. M., & Trueman, S. J. (2015). Maturation in Corymbia torelliana × C. citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations. Forests, 6(10), 3763-3778. https://doi.org/10.3390/f6103763