
Impact of a Mountain Pine Beetle Outbreak on Young Lodgepole Pine Stands in Central British Columbia

Abstract
:1. Introduction
2. Experimental Section
2.1. Stand Selection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Class (AC) | Age Range (Years) | Number of Stands | Stands by Biogeoclimatic Subzone | Number of TSP | ||
|---|---|---|---|---|---|---|
| Moist | Mesic | Dry | ||||
| 1 | 13–20 | 52 | 25 | 16 | 11 | 254 |
| 2 | 21–40 | 148 | 46 | 31 | 71 | 738 |
| 3 | 41–60 | 38 | 13 | 6 | 19 | 190 |
| 4 | 61–80 | 71 | 15 | 13 | 43 | 345 |
| Overall | 309 | 99 | 66 | 144 | 1527 | |
2.2. Sampling Protocol
2.3. Surviving Residual Secondary Structure Data Collection
2.4. Analysis
3. Results
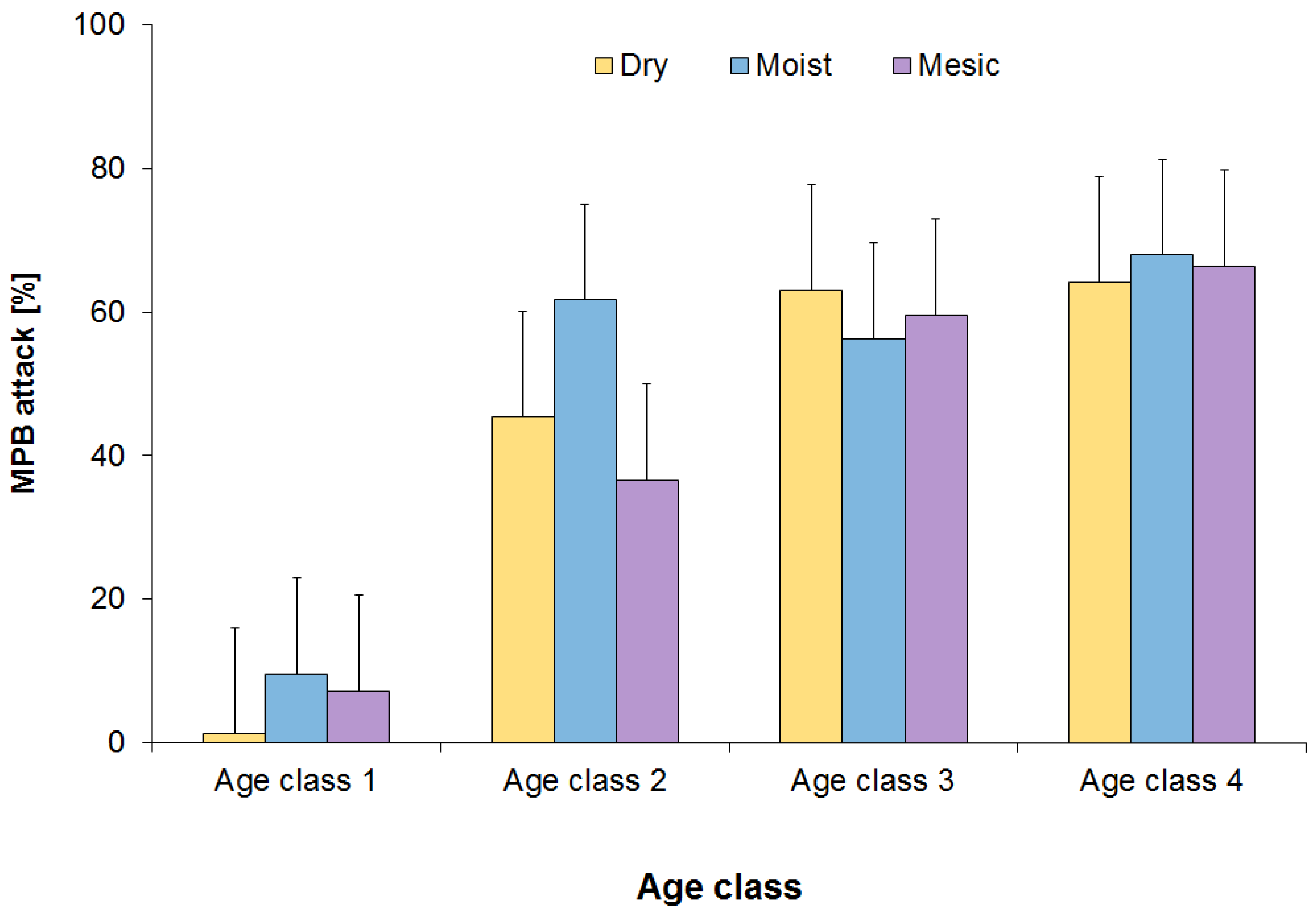
3.1. MPB Attack

| Age Class (Years) | Equation (Y = a + bX) | R2 | F | p | df |
|---|---|---|---|---|---|
| MPB attack as a function of stand mean pine DBH | |||||
| 1 (13–20) | −46.5985 + 4.4209 × X | 0.1576 | 10.5387 | 0.002 | 1.50 |
| 2 (21–40) | −46.5292 + 6.8968 × X | 0.2133 | 40.8669 | <0.001 | 1.146 |
| 3 (41–60) | 0.1895 + 4.4400 × X | 0.3627 | 22.0534 | <0.001 | 1.36 |
| 4 (61–80) | 11.2292 + 3.3448 × X | 0.4426 | 56.5885 | <0.001 | 1.69 |
| MPB attack as a function of stand mean density (stems ha−1) | |||||
| 1 (13–20) | −1.2010 + 0.0050 × X | 0.0000 | 0.6446 | 0.4258 | 1.50 |
| 2 (21–40) | 57.7120 − 0.0104 × X | 0.0169 | 3.5194 | 0.0626 | 1.146 |
| 3 (41–60) | 79.2897 − 0.0132 × X | 0.2392 | 12.6282 | 0.0011 | 1.36 |
| 4 (61–80) | 68.9088 − 0.0095 × X | 0.0640 | 5.7830 | 0.0189 | 1.69 |
| Age Class (Years) | Attack Category | Total Number of Stands | Stands (%) | Mean MPB Pine Attack (%) | Mean Pine DBH (cm) |
|---|---|---|---|---|---|
| 1 (13–20) | A (≤25%) | 48 | 92 | 1.2 | 11.8 |
| B (25.1%–50%) | 0 | 0 | n/a | n/a | |
| C (50.1%–75%) | 3 | 6 | 63.7 | 14.8 | |
| D (>75.1%) | 1 | 2 | 84.5 | 13.6 | |
| Total | 52 | 100 | 6.4 | 12.0 | |
| 2 (21–40) | A (≤25%) | 43 | 29 | 3.1 | 12.9 |
| B (25.1%–50%) | 26 | 18 | 42.3 | 12.7 | |
| C (50.1%–75%) | 35 | 23 | 65.5 | 14.0 | |
| D (>75.1%) | 44 | 30 | 85.9 | 15.6 | |
| Total | 148 | 100 | 49.4 | 13.9 | |
| 3 (41–60) | A (≤25%) | 3 | 8 | 10.8 | 10.1 |
| B (25.1%–50%) | 8 | 21 | 38.4 | 12.5 | |
| C (50.1%–75%) | 14 | 37 | 66.3 | 13.5 | |
| D (>75.1%) | 13 | 34 | 85.3 | 16.5 | |
| Total | 38 | 100 | 62.6 | 14.0 | |
| 4 (61–80) | A (≤25%) | 2 | 3 | 13.6 | 10.8 |
| B (25.1%–50%) | 13 | 18 | 41.2 | 14.1 | |
| C (50.1%–75%) | 24 | 34 | 64.6 | 16.1 | |
| D (>75.1%) | 32 | 45 | 88.1 | 20.2 | |
| Total | 71 | 100 | 69.5 | 17.4 |
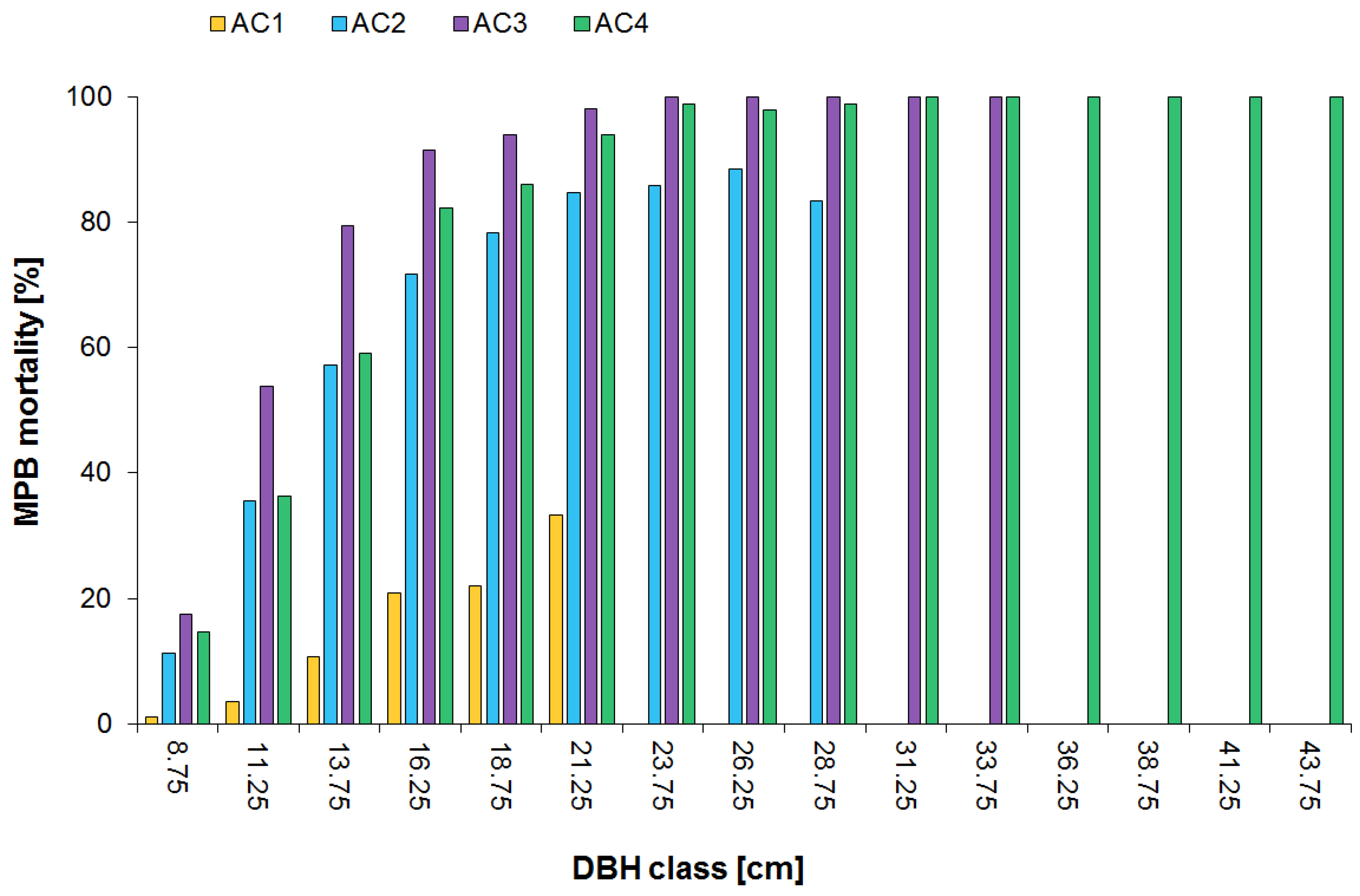
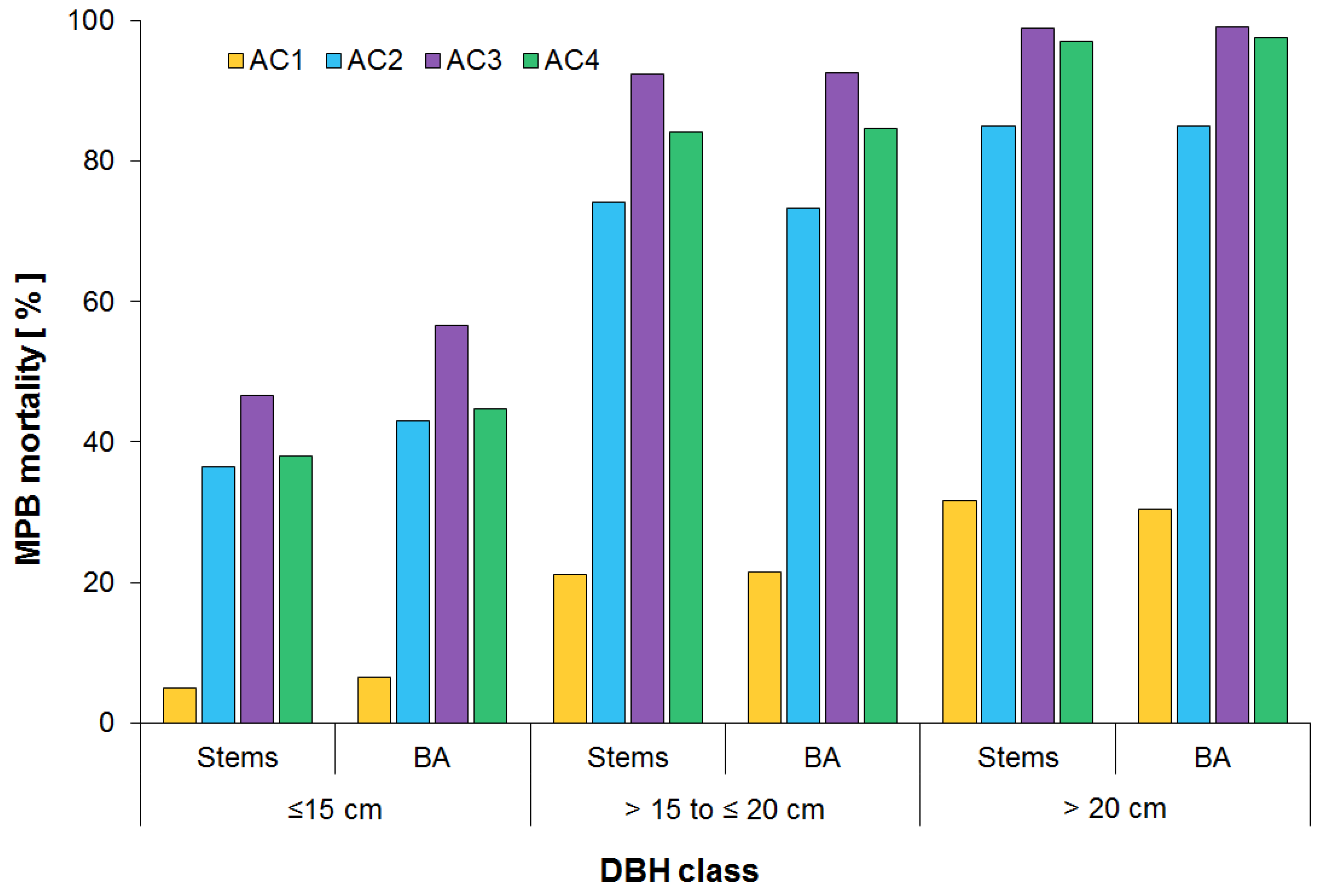
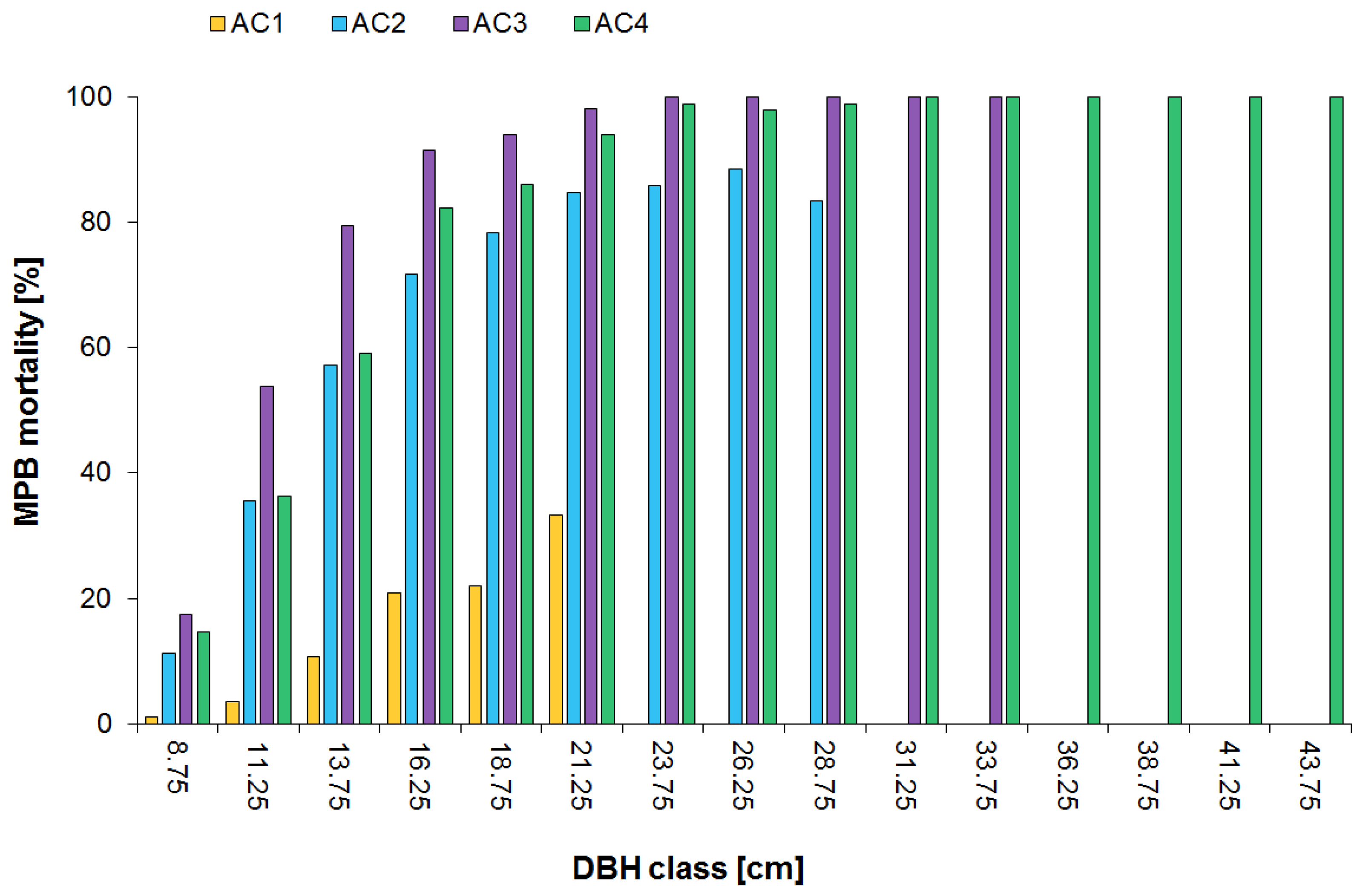
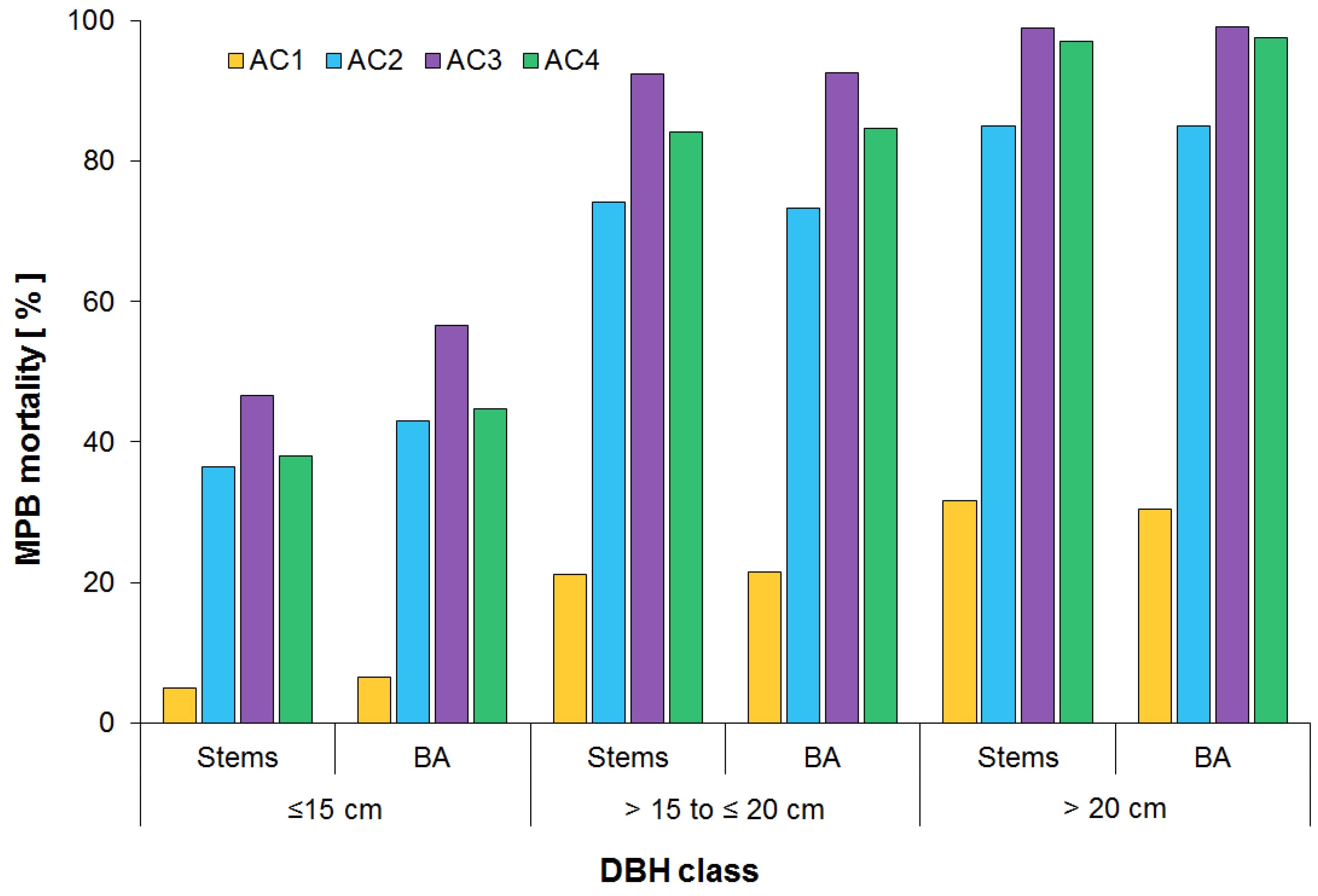
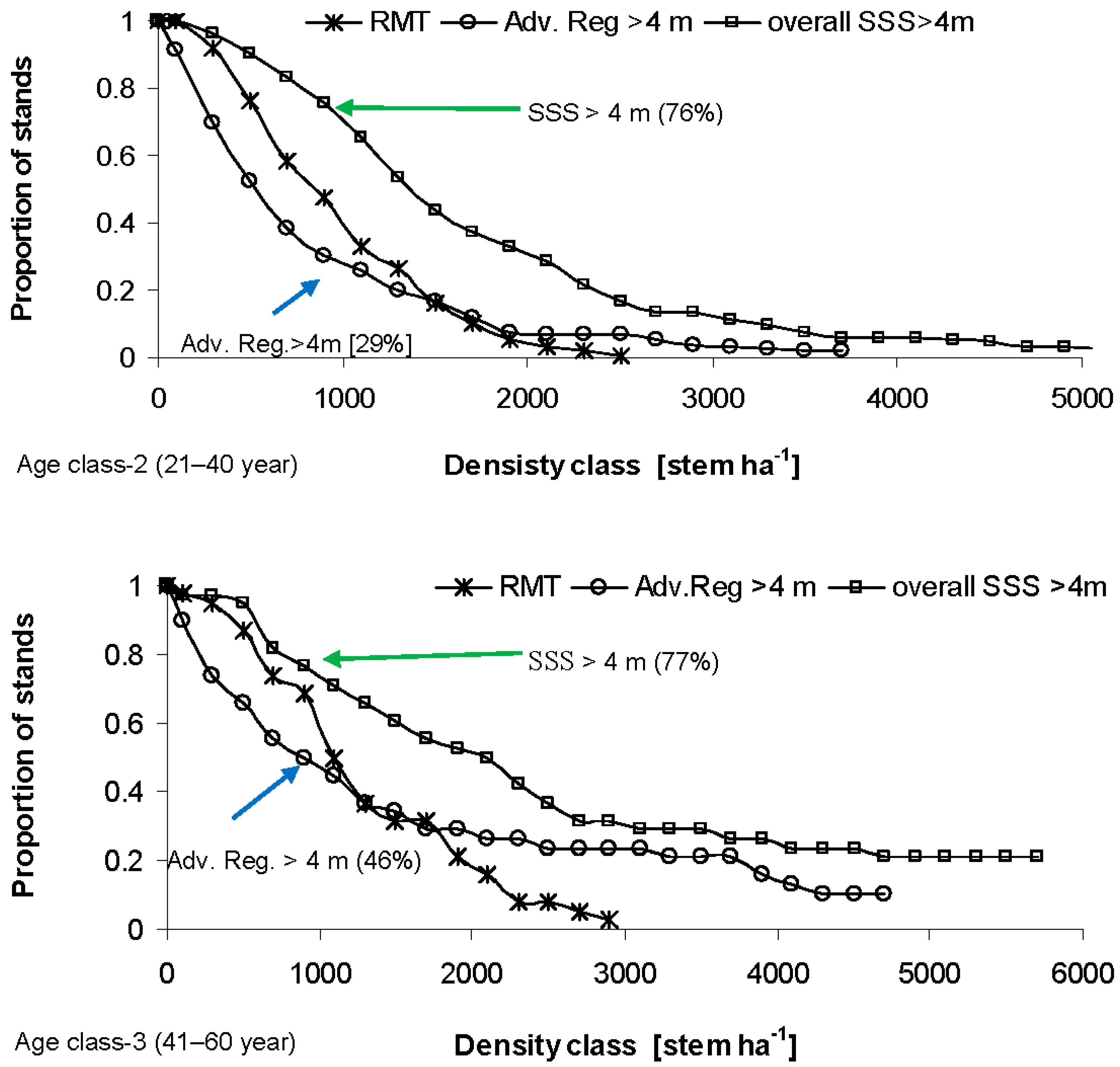
3.2. Surviving Residual Structure of Immature Stands
| Source | Mean Square | df | F | p |
|---|---|---|---|---|
| Residual mature trees (DBH ≥ 7.5 cm) | ||||
| Age class | 6,627,569.67 | 3 | 22.815 | <0.001 |
| Biogeoclimatic/Ecological subzone | 590,068.64 | 2 | 2.031 | 0.133 |
| Error | 290,491.55 | 303 | ||
| Advance regeneration (>4 m height and DBH < 7.5 cm) | ||||
| Age class | 13,174,804.15 | 3 | 10.326 | <0.001 |
| Biogeoclimatic/Ecological subzone | 5,184,251.71 | 2 | 4.063 | 0.018 |
| Error | 1,275,857.19 | 288 | ||
| Age Class (Year) | RMT DBH ≥ 7.5 cm (stem ha−1) | Adv. Reg. Height >4 m DBH < 7.5 cm (stem ha−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Dry | Moist | Mesic | Mean | Dry | Moist | Mesic | Mean | |
| 1 (13–20) | 1309 | 1403 | 1301 | 1338 c | 789 | 592 | 850 | 744 b |
| 2 (21–40) | 847 | 733 | 989 | 856 a | 714 | 1031 | 460 | 735 b |
| 3 (41–60) | 1097 | 1296 | 1098 | 1164 b | 2000 | 1846 | 527 | 1458 c |
| 4 (61–80) | 969 | 821 | 626 | 805 a | 688 | 773 | 203 | 555 a |
| Eco. Subzone (mean) | 1056 x | 1063 x | 1004 x | 1048 y | 1061 y | 510 x | ||
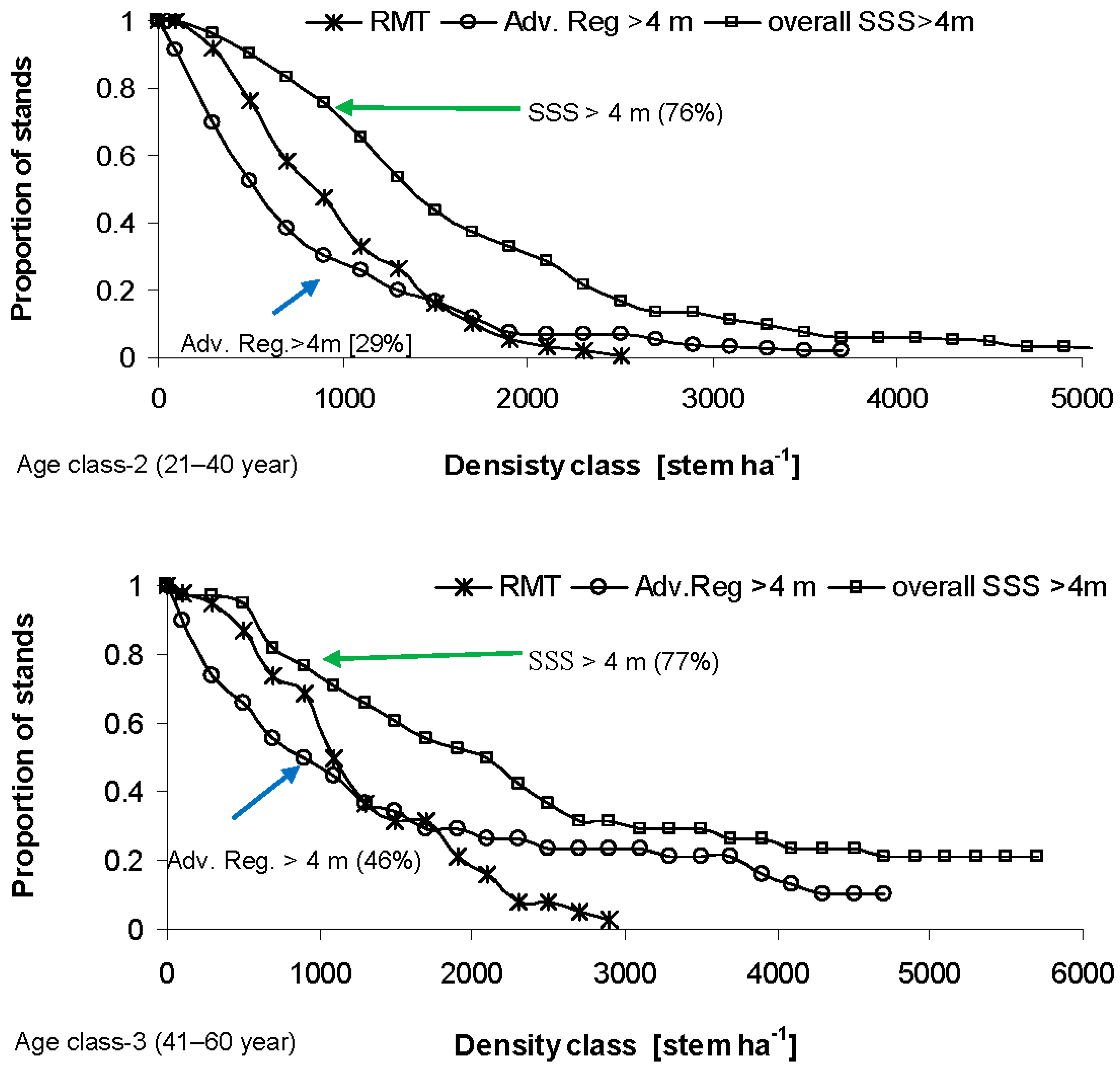
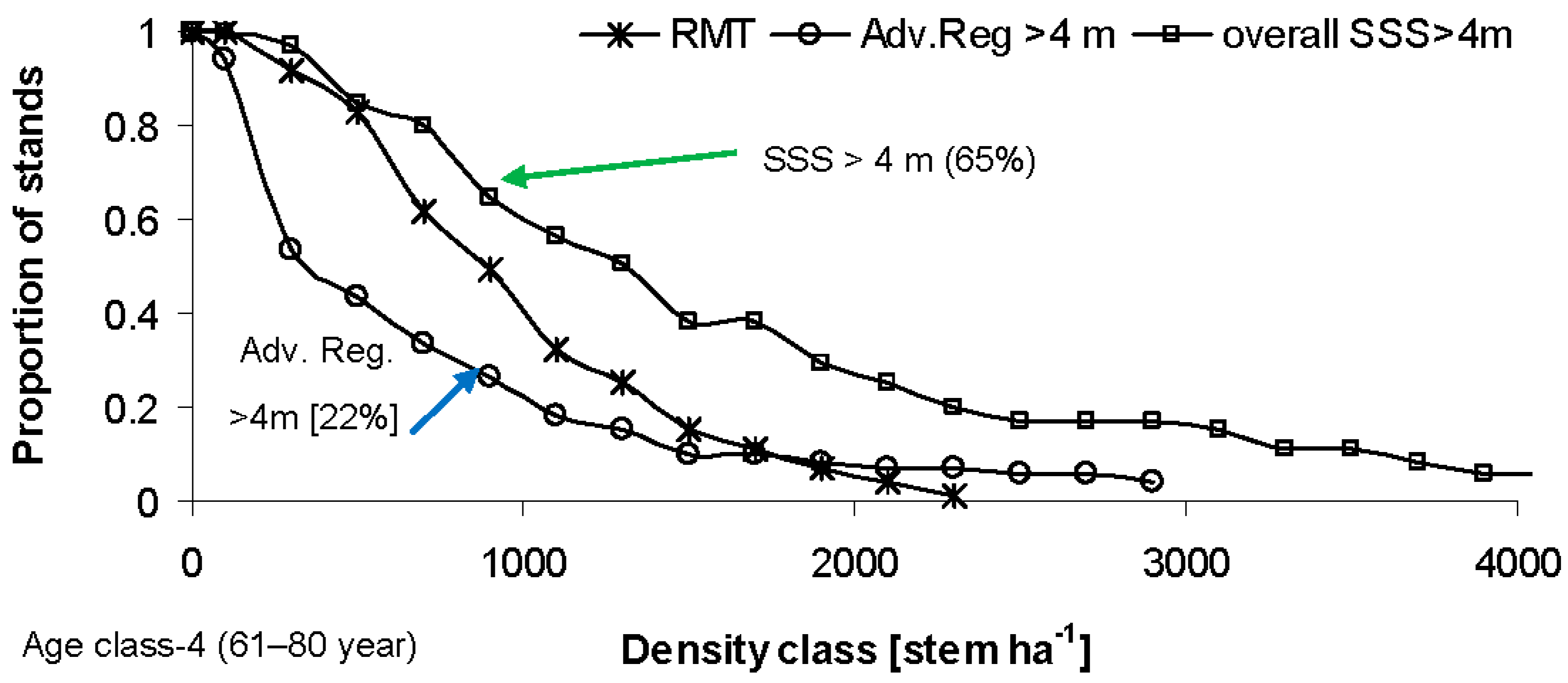
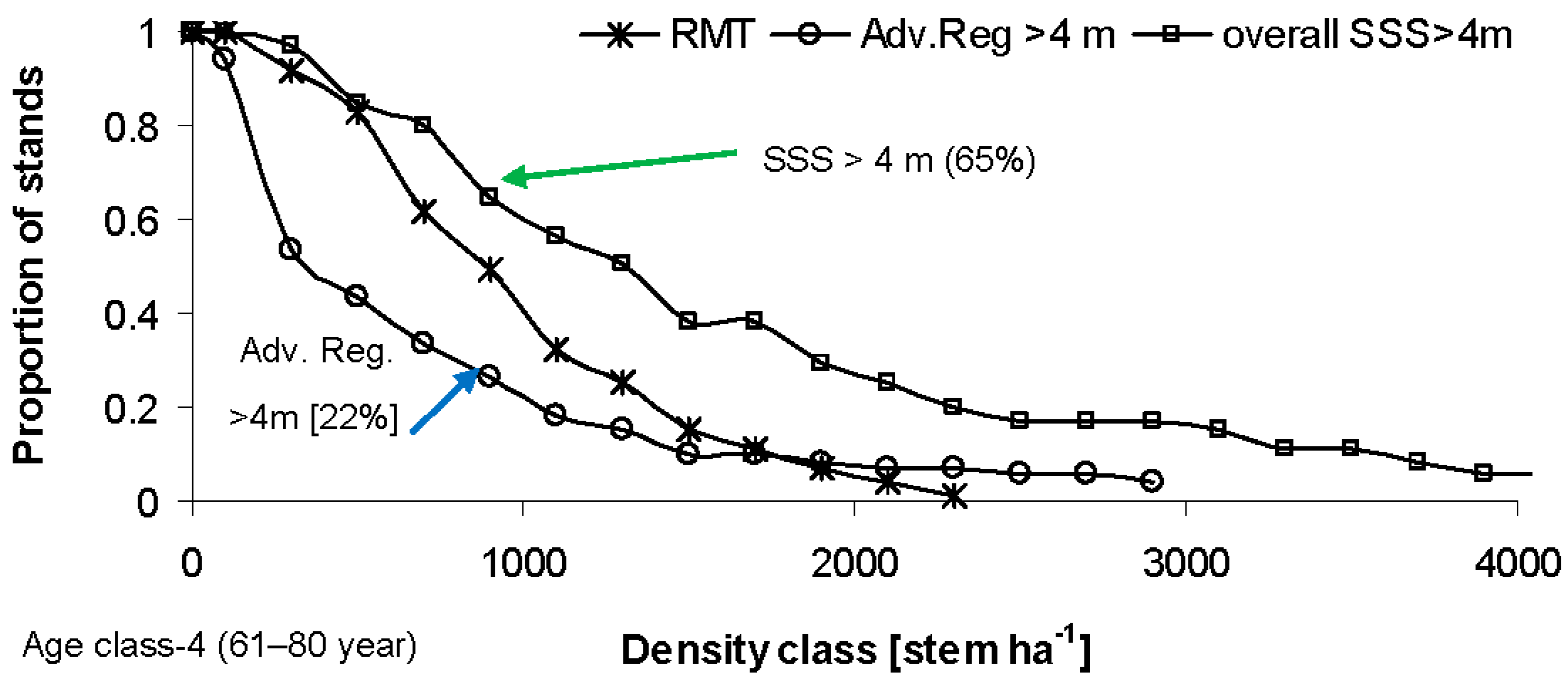
4. Discussion
4.1. MPB Attack
4.2. Post Beetle Stand Dynamics of Immature Stands
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Safranyik, L.; Carroll, A.L. The biology and epidemiology of the mountain pine beetle in Lodgepole pine forests. In The Mountain Pine Beetle: A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Safranyik, L., Wilson, B., Eds.; Natural Resources Canada: Victoria, BC, Canada, 2006; pp. 3–66. [Google Scholar]
- Roe, A.L.; Amman, G.D. The Mountain Pine Beetle in Lodgepole Pine Forests; Research Paper INT-71; USDA Forest Service Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1970; Available online: http://www.usu.edu/beetle/documents2/197Roe_Amman_MPBin_LPP_Forests.pdf (accessed on 18 May 2015).
- Taylor, S.; Carroll, A. Disturbance, forest age, and mountain pine beetle outbreak dynamics in BC: A historical perspective. In Mountain Pine Beetle Symposium: Challenges and Solutions; Information Report BC-X-399; Shore, T., Brooks, J., Stone, J., Eds.; Canadian Forest Service: Victoria, BC, Canada, 2004; pp. 44–51. [Google Scholar]
- Walton, A. Provincial-Level Projection of the Current Mountain Pine Beetle Outbreak: Update of the Infestation Projection Based on the Provincial Aerial Overview Surveys of Forest Health Conducted from 1999 through 2012 and the BCMPB Model (Year 10); BC Ministry of Forests, Lands and Natural Resource Operations: Victoria, BC, Canada, 2013. Available online: http://www.for.gov.bc.ca/ftp/hre/external/!publish/web/bcmpb/year10/BCMPB.v10.BeetleProjection.Update.pdf (accessed on 15 May 2015).
- Alfaro, R.; Axelson, J.; Hawkes, B. Mountain pine beetle increases the complexity of fire-origin lodgepole pine stands in British Columbia, Canada. J. Ecosyst. Manag. 2008, 9, 1–5. [Google Scholar]
- Westfall, J.; Ebata, T. Summary of Forest Health Conditions in British Columbia; Pest Management Report Number 15; BC Forest Service: Victoria, BC, Canada, 2008. Available online: http://www.for.gov.bc.ca/ftp/HFP/external/!publish/Aerial_Overview/2008/Aerial%20Overview%202008.pdf (accessed on 8 May 2015).
- Amman, G.D.; McGregor, M.D.; Cahill, D.B.; Klein, W.H. Guidelines for Reducing Losses of Lodgepole Pine to the Mountain Pine Beetle in the Rocky Mountains; UT. Technical Report INT-36; USDA Forest Service; Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1977; p. 28. [Google Scholar]
- Eng, M.; Fall, A.; Hughes, J.; Shore, T.; Riel, B.; Hall, P.; Walton, A. Provincial Level Projection of the Current Mountain Pine Beetle Outbreak: An Overview of the Model (BCMPB v2) and Results of Year 2 of the Project; BC Forest Service: Victoria, BC, Canada, 2005; Available online: http://www.llbc.leg.bc.ca/public/pubdocs/bcdocs/376681/bcmpb_mainreport_2004.pdf (accessed on 12 May 2015).
- BC Ministry of Forests and Range. Timber Supply and the Mountain Pine Beetle Infestation in British Columbia: 2007 Update; Forest Analysis and Inventory Branch; BC Ministry of Forest and Range: Victoria, BC, Canada, 2007; p. 32. [Google Scholar]
- MacLauchlan, L. Status of Mountain Pine Beetle Attack in Young Lodgepole Pine Stands in Central British Columbia; Report for the Chief Forester of British Columbia; BC Ministry of Forests and Range: Victoria, BC, Canada, 2006. Available online: http://www.for.gov.bc.ca/rsi/ForestHealth/PDF/Young%20Pine%20Report_CF_final.pdf (accessed on 14 May 2012).
- Runzer, K.; Hawkins, C.D.B. Success Rate of MPB (Mountain Pine Beetle) Attack in Young Stands. Executive Summary; FIA FSP Project M065002; BC Ministry of Forests and Range: Victoria, BC, Canada, 2006. Available online: http://www.for.gov.bc.ca/hfd/library/FIA/HTML/FIA2008MR194.htm (accessed on 7 May 2015).
- MacLauchlan, L.E.; Brooks, J. Determining Susceptibility of Young Pine Stands to the Mountain Pine Beetle, Dendroctonus Ponderosae, and Manipulating Future Stands to Mitigate Losses; Final Technical Report; FIA FSP Project Y072003; BC Ministry of Forests and Range: Victoria, BC, Canada, 2007. Available online: http://www.for.gov.bc.ca/hfd/library/FIA/HTML/FIA2008MR194.htm (accessed on 15 April 2015).
- Nussbaum, A. Forecasting the effects of species choices on long-term harvest levels: Sub-component of the provincial mountain pine beetle analysis project. In Proceedings of the Northern Silviculture Committee 2006 Winter Workshop; Victoria, BC, Canada, 2006. Available online: http://www.for.gov.bc.ca/hts/tsa/tsr3/rationale.pdf (accessed on 15 April 2015). [Google Scholar]
- Association of British Columbia Forest Professionals (ABCFP). Mid-Term Timber Supply Advocacy Report; Association of British Columbia Forest Professionals: Vancouver, BC, Canada, 2011. [Google Scholar]
- Bogdanski, B.; Sun, L.; Peter, B.; Stennes, B. Markets for Forest Products Following a Large Disturbance: Opportunities and Challenges from the Mountain Pine Beetle Outbreak in Western Canada; Report BC-X-429; Natural Resource Canada, Canada Forest Services: Victoria, BC, Canada, 2011; Available online: http://cfs.nrcan.gc.ca/pubwarehouse/pdfs/32226.pdf (accessed on 18 April 2015).
- Pousette, J.; Hawkins, C. An assessment of critical assumptions supporting the timber supply modelling for mountains-pine-beetle-induced allowable annual cut uplift in the Prince George Timber Supply Area. BC J. Ecosyst. Manag. 2006, 7, 93–104. [Google Scholar]
- Hawkins, C.D.B.; Dhar, A.; Balliet, N.; Runzer, K.D. Residual mature trees and secondary stand structure after mountain pine beetle attack in central British Columbia. For. Ecol. Manag. 2012, 277, 107–115. [Google Scholar] [CrossRef]
- Meidinger, D.; Pojar, J.; Harper, W.L. Sub-boreal spruce zone. In Ecosystems of British Columbia; Meidinger, D., Pojar, J., Eds.; BC Ministry of Forests: Victoria, BC, Canada, 1991; pp. 195–207. [Google Scholar]
- BC Ministry of Forest Range. Explanation of the Forest Planning and Practices Regulation Amendments to Protect Secondary Structure; B.C. Forest Service: Victoria, BC, Cananda, 2008. Available online: http://www.for.gov.bc.ca/hfp/silviculture/secondary_structure/secondary_structure_reg.pdf (accessed on 14 April 2015).
- Hawkins, C.D.B.; Dhar, A.; Balliet, N. Radial growth of residual overstory trees and understory saplings after mountain pine beetle attack in central British Columbia. For. Ecol. Manag. 2013, 310, 348–356. [Google Scholar] [CrossRef]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MD, USA, 2014; p. 291. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.M.; Walker, S. Fitting linear mixed-effects models using lme4; Journal of Statistical Software. Available online: http://arxiv.org/abs/1406.5823 (accessed on 16 September 2015).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: http://www.R-project.org/ (accessed on 15 March 2014).
- BC Ministry of Forests and Range. FFT Walk through Ground Reconnaissance Survey Procedures for Stands Impacted by MPB; BC Forest Service: Victoria, BC, Canada, 2006. Available online: http://www.for.gov.bc.ca/ftp/hfp/external/!publish/FIA%20Documents/Standards/FFT_Survey/MPB_Survey_Standard_PGTSA.pdf (accessed on 18 April 2015).
- Kane, J.M.; Kolb, T.E. Importance of resin ducts in reducing ponderosa pine mortality from bark beetle attack. Oecologia 2010, 164, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Kane, J.M.; Mitton, J.B. Resin duct characteristics associated with tree resistance to bark beetles across lodgepole and limber pines. Oecologia 2014, 174, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Shore, T.L.; Safranyik, L.; Lemieux, J.P. Susceptibility of lodgepole pine stands to the mountain pine beetle: Testing of a rating system. Can. J. For. Res. 2000, 30, 44–49. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negrón, J.F.; Castello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caisse, R. Stand characteristics and downed woody debris accumulations associated with a mountain pine beetle (Dendroctonus ponderosae Hopkins) outbreak in Colorado. For. Ecol. Manag. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Björklund, N.; Lindgren, B.S.; Shore, T.L.; Cudmore, T. Can predicted mountain pine beetle net production be used to improve stand prioritization for management? For. Ecol. Manag. 2009, 257, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Progar, R.A.; Blackford, D.C.; Cluck, D.R.; Costello, S.; Dunning, L.B.; Eager, T.; Jorgensen, C.L.; Munson, A.S.; Steed, B.; Rinella, M.J. Population densities and tree diameter effects associated with verbenone treatments to reduce mountain pine beetle caused mortality of lodgepole pine. J. Econ. Entomol. 2013, 106, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Coates, K.D.; Glover, T.; Henderson, B. Abundance of Secondary Structure in Lodgepole Pine Stands Affected by the Mountain Pine Beetle in the Cariboo-Chilcotin; Mountain Pine Beetle Working Paper 2009–20; Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2009; Available online: http://cfs.nrcan.gc.ca/pubwarehouse/pdfs/31195.pdf (accessed on 12 May 2015).
- Vyse, A.; Ferguson, C.; Haggard, D.J.; Roach, J.; Zimonick, B. Regeneration beneath lodgepole pine dominated stands attacked or threatened by the mountain pine beetle in the south central Interior, British Columbia. For. Ecol. Manag. 2009, 258, S36–S43. [Google Scholar] [CrossRef]
- Axelson, J.N.; Alfaro, R.I.; Hawkes, B.C. Changes in stand structure in uneven-aged lodgepole pine stands impacted by mountain pine beetle epidemics and fires in central British Columbia. For. Chron. 2010, 86, 87–99. [Google Scholar] [CrossRef]
- Veblen, T.T.; Hadley, K.S.; Reid, M.; Rebertus, A.J. The response of subalpine forests to spruce beetle outbreak in Colorado. Ecology 1991, 72, 213–231. [Google Scholar] [CrossRef]
- Murphy, T.E.L.; Adams, D.L.; Ferguson, D.E. Response of advance lodgepole pine regeneration to overstory removal in eastern Idaho. For. Ecol. Manag. 1999, 120, 234–244. [Google Scholar]
- Amoroso, M.M.; Coates, K.D.; Astrup, R. Stand recovery and self-organization following large-scale mountain pine beetle induced canopy mortality in northern forests. For. Ecol. Manag. 2013, 310, 300–311. [Google Scholar] [CrossRef]
- Dhar, A.; Coates, K.D.; Rogers, B.; Hardy, K. Impact of mountain pine beetle on mid-term timber supply in sub boreal spruce zone of British Columbia. In Proceedings of the 16th International Boreal Forest Research Association (IBFRA) Conference, Edmonton, AB, Canada, 7–10 October 2013; p. 28.
- Astrup, R.; Coates, K.D.; Hall, E. Recruitment limitations in forests: Lessons from an unprecedented mountain pine beetle epidemic. For. Ecol. Manag. 2008, 256, 1743–1750. [Google Scholar] [CrossRef]
- Dhar, A.; Hawkins, C.D.B. Regeneration and growth following mountain pine beetle attack: A synthesis of knowledge. J. Ecosyst. Manag. 2011, 12, 1–16. [Google Scholar]
- Agee, J.K. Fire ecology of Pacific Northwest Forests; Island Press: Washington, DC, USA, 1993; p. 505. [Google Scholar]
- Lindenmayer, D.B.; Burton, P.J.; Franklin, F.J. Salvage Logging and its Ecological Consequences; Island Press: Washington, DC, USA, 2008; p. 246. [Google Scholar]
- Fedrowitz, K.; Koricheva, J.; Baker, S.C.; Lindenmayer, D.B.; Palik, B.; Rosenvald, R.; Beese, W.; Franklin, J.F.; Kouki, J.; Macdonald, E.; et al. REVIEW: Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 2014, 51, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.; Parrott, L. Salvage logging after mountain pine beetle outbreaks reduces the social-ecological resilience of forest landscapes. In Proceedings of the Mountain Pine Beetle Information Exchange Forum, Edmonton, AB, Canada, 22–23 April 2015; p. 17.
- Drever, R.C.; Peterson, G.; Messier, C.; Bergeron, Y.; Flannigan, M. Can forest management based on natural disturbances maintain ecological resilience? Can. J. For. Res. 2006, 36, 2285–2299. [Google Scholar] [CrossRef]
- Park, A.; Puettmann, K.; Wilson, E.; Messier, C.; Kames, S.; Dhar, A. Can boreal and temperate forest management be adapted to the uncertainties of 21st century climate change? Crit. Rev. Plant Sci. 2014, 33, 251–285. [Google Scholar] [CrossRef]
- Faccoli, M.; Bernardinelli, I. Composition and elevation of spruce forests affect susceptibility to bark beetle attacks: Implications for forest management. Forests 2014, 5, 88–102. [Google Scholar] [CrossRef]
- Nelson, T.A.; Boots, B.; Wulder, M.A.; Carroll, A.L. Environmental characteristics of mountain pine beetle infestation hot spots. J. Ecosyst. Manag. 2008, 8, 91–108. [Google Scholar]
- Dhar, A.; Parrott, L.; Heckbert, S. Consequences of mountain pine beetle outbreaks on forest ecosystem services in western Canada. In Proceedings of the Mountain Pine Beetle Information Exchange Forum, Edmonton, AB, Canada, 22–23 April 2015; p. 17.
- Dhar, A.; Parrott, L.; Heckbert, S. Mapping the impact of mountain pine beetle outbreaks on forest ecosystem services in British Columbia, Canada. In Proceedings of the IUFRO Landscape Ecology Conference Sustaining Ecosystem Services in Forest Landscapes Concepts, Research, and Applications, Tartu, Estonia, 23–30 August 2015; Available online: http://iufrole2015.to.ee/download/m55d4ea9be5748#iufrole_2015_abstracts_pdf (accessed on 02 September 2015).
- Pec, G.J.; Karst, J.; Sywenky, A.N.; Cigan, P.W.; Erbilgin, N.; Simard, S.W.; Cahill, J.F., Jr. Rapid increases in forest understory diversity and productivity following a mountain pine beetle (Dendroctonus ponderosae) outbreak in pine forests. PLoS ONE 2015, 10, e0124691. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.E.; Wolfe, M.L. Response of understorey vegetation to variable tree mortality following a mountain pine beetle epidemic in lodgepole pine stands in northern Utah. Vegetatio 1996, 122, 1–12. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhar, A.; Balliet, N.A.; Runzer, K.D.; Hawkins, C.D.B. Impact of a Mountain Pine Beetle Outbreak on Young Lodgepole Pine Stands in Central British Columbia. Forests 2015, 6, 3483-3500. https://doi.org/10.3390/f6103483
Dhar A, Balliet NA, Runzer KD, Hawkins CDB. Impact of a Mountain Pine Beetle Outbreak on Young Lodgepole Pine Stands in Central British Columbia. Forests. 2015; 6(10):3483-3500. https://doi.org/10.3390/f6103483
Chicago/Turabian StyleDhar, Amalesh, Nicole A. Balliet, Kyle D. Runzer, and Christopher D. B. Hawkins. 2015. "Impact of a Mountain Pine Beetle Outbreak on Young Lodgepole Pine Stands in Central British Columbia" Forests 6, no. 10: 3483-3500. https://doi.org/10.3390/f6103483
APA StyleDhar, A., Balliet, N. A., Runzer, K. D., & Hawkins, C. D. B. (2015). Impact of a Mountain Pine Beetle Outbreak on Young Lodgepole Pine Stands in Central British Columbia. Forests, 6(10), 3483-3500. https://doi.org/10.3390/f6103483