Managing Understory Vegetation for Maintaining Productivity in Black Spruce Forests: A Synthesis within a Multi-Scale Research Model


 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction


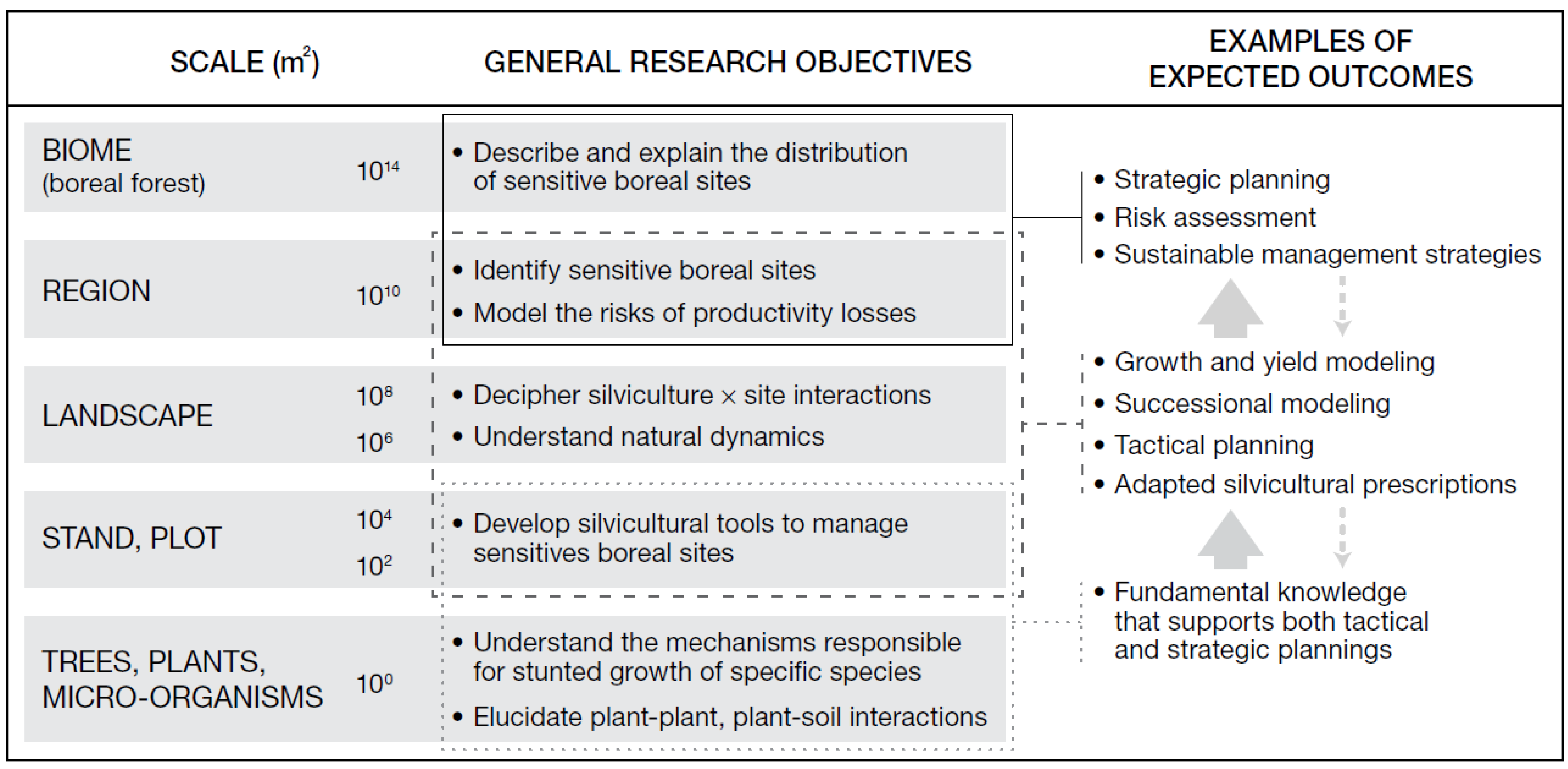
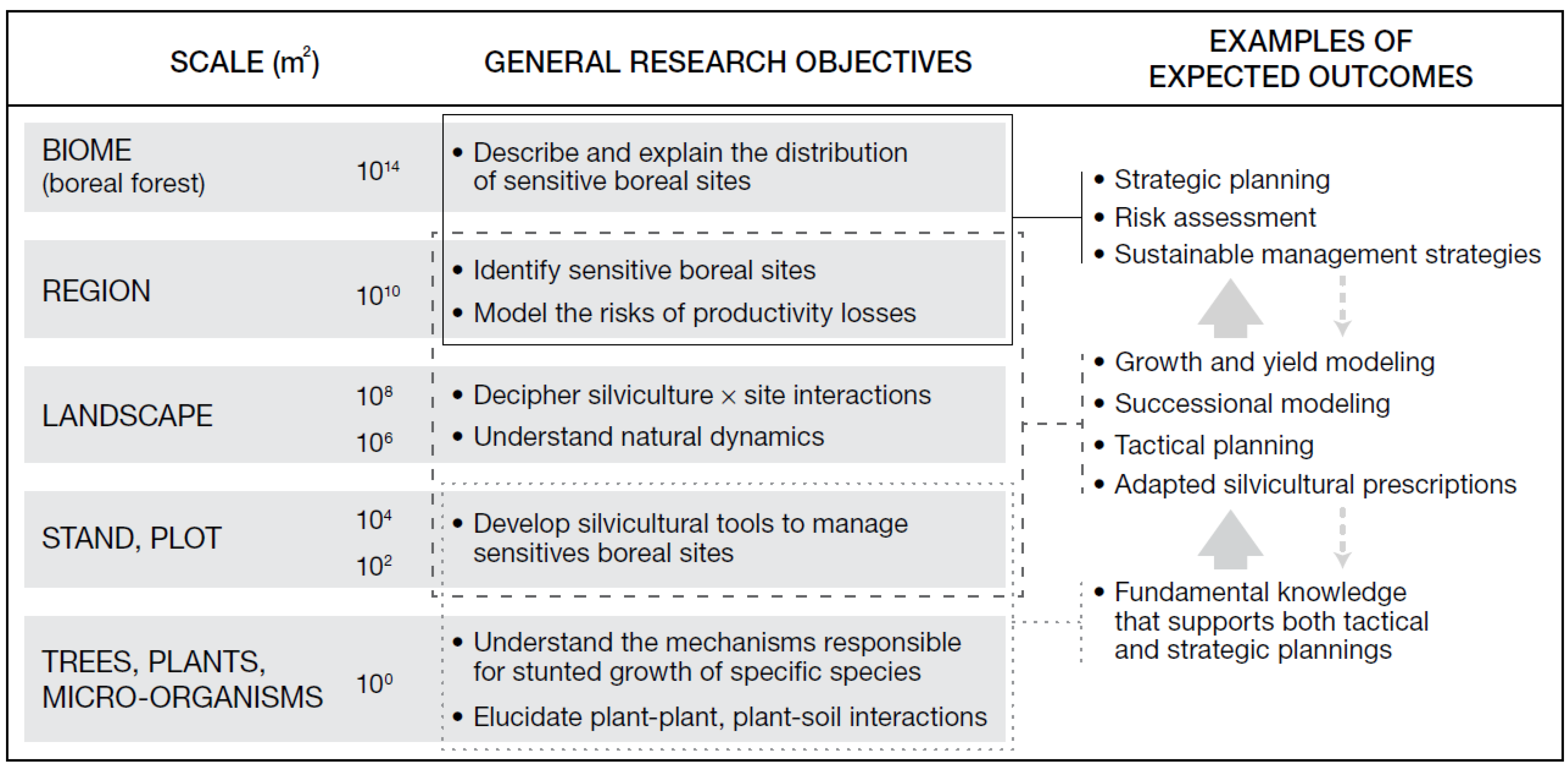
2. A Multi-Scale Research Model
3. Ericad Dominance
3.1. The Tree and Plant Level
3.2. The Stand/Plot Level
3.3. The Landscape Level
3.4. The Regional Level
3.5. The Provincial (or Biome) Level
4. Paludification
4.1. The Tree and Plant Level
4.2. The Stand/Plot Level
4.3. The Landscape Level
4.4. The Regional/Provincial (or Biome) Level
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Lieffers, V.J.; Messier, C.; Burton, P.; Ruel, J.-C.; Grover, G.E. Nature-based silviculture for sustaining a variety of boreasl forest values. In Towards Sustainable Management of the Boreal Forest; Burton, P.J., Messier, C., Smith, D.W., Adamowicz, W.L., Eds.; NRC Research Press: Ottawa, ON, USA, 2003; pp. 481–530. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F.; Lõhmus, A.; Baker, S.C.; Bauhus, J.; Beese, W.; Brodie, A.; Kiehl, B.; Kouki, J.; Pastur, G.M.; Messier, C.; Neyland, M.; Palik, B.; Sverdrup-Thygeson, A.; Volney, J.; Wayne, A.; Gustafsson, L. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012, 5, 421–431. [Google Scholar] [CrossRef]
- Gauthier, S.; Vaillancourt, M.A.; Leduc, A.; De Grandpré, L.; Kneeshaw, D.; Morin, H.; Drapeau, P.; Bergeron, Y. Ecosystem Management in the Boreal Forest; Presses de l’Université du Québec: Québec, Canada, 2009; p. 572. [Google Scholar]
- Saucier, J.P.; Robitaille, A.; Grondin, P. Cadre bioclimatique du Québec. In Écologie forestière. Manuel de foresterie, 2nd ed. Ordre des ingénieurs forestiers du Québec; Éditions Multimondes: Québec, Canada, 2009; pp. 186–205. [Google Scholar]
- Harvey, B.; Brais, S. Effects of mechanized careful logging on natural regeneration and vegetation competition in the southeastern Canadian boreal forest. Can. J. For. Res. 2002, 32, 653–666. [Google Scholar] [CrossRef]
- Raymond, P.; Bédard, S.; Roy, V.; Larouche, C.; Tremblay, S. The irregular shelterwood system: review, classification, and potential application to forests affected by partial disturbances. J. For. 2009, 108, 405–413. [Google Scholar]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Can. J. For. Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Nilsson, M.-C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- De Montigny, L.M.; Weetman, G.F. The Effects of Ericaceous Plants on Forest Productivity; N-X-271; Canadian Forest Service, Forestry Canada: Newfoundland, Canada, 1990; pp. 83–90. [Google Scholar]
- Read, D.J.; Jalal, M.A.F. The Physiological Basis of Interaction between Calluna Vulgaris, Forest Trees, and Other Plant Species, Proceedings of Weed Control in Forestry. University of Nottingham, Nottingham, UK, 1–2 April 1980; Atkinson, D., McCavish, W.J., Spencer-Jones, D.H., O’Keeffe, M.G., Allen, M.G., Makepeace, R.J., Eds.; Association of Applied Biologists—Weed Group in conjunction with the Institute of Foresters of Great Britain, University of Nottingham: Nottingham, UK, 1980; pp. 21–32.
- Zackrisson, O.; Nilsson, M.C.; Dahlberg, A.; Jäderlund, A. Interference mechanisms in conifer-Ericaceae-feathermoss communities. Oikos 1997, 78, 209–220. [Google Scholar] [CrossRef]
- Prescott, C.E.; Sajedi, T. The role of salal in forest regeneration problems in coastal British Columbia: problem or symptom? For. Chron. 2008, 84, 29–36. [Google Scholar]
- Mallik, A.U. Conifer regeneration problems in boreal and temperate forests with ericaceous understory: Role of disturbance, seedbed limitation, and keystone species change. Crit. Rev. Plant. Sci. 2003, 22, 341–366. [Google Scholar] [CrossRef]
- Hébert, F.; Thiffault, N. The biology of Canadian weeds. 146. Rhododendron groenlandicum (Oeder) Kron and Judd. Can. J. Plant. Sci. 2011, 91, 725–738. [Google Scholar] [CrossRef]
- Joosten, H.; Clarke, D. Wise Use of Mires and Peatlands: Background and Principles Including A Framework for Decision-Making; International Mire Conservation Group and International Peat Society: Jyväskylä, Finland, 2002; p. 303. [Google Scholar]
- Lavoie, M.; Paré, D.; Fenton, N.; Groot, A.; Taylor, K. Paludification and management of forested peatland in Canada: a literature review. Can. J. For. Res. 2005, 13, 21–50. [Google Scholar]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.Y.; Paré, D. Forest productivity decline caused by successional paludification of boreal soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef]
- Mallik, A.U. Conversion of temperate forests into heaths: Role of ecosystem disturbance and ericaceous plants. Environ. Manag. 1995, 19, 675–684. [Google Scholar] [CrossRef]
- Ganio, L.M.; Puettmann, K.J. Designing long-term, large-scale forestry experiments with research objectives at multiple scales. J. Sust. For. 2008, 26, 1–18. [Google Scholar] [CrossRef]
- Peterson, E.B. Inhibition of black spruce primary roots by a water-soluble substance in Kalmia angustifolia. For. Sci. 1965, 11, 473–479. [Google Scholar]
- Mallik, A.U. Allelopathic potential of Kalmia angustifolia to black spruce (Picea mariana). For. Ecol. Manag. 1987, 20, 43–51. [Google Scholar]
- Thiffault, N.; Titus, B.D.; Munson, A.D. Black spruce seedlings in a Kalmia-Vaccinium association: Microsite manipulation to explore interactions in the field. Can. J. For. Res. 2004, 34, 1657–1668. [Google Scholar] [CrossRef]
- LeBel, P.; Thiffault, N.; Bradley, R.L. Kalmia removal increases nutrient supply and growth of black spruce seedlings: An effect fertilizer cannot emulate. For. Ecol. Manag. 2008, 256, 1780–1784. [Google Scholar] [CrossRef]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Munson, A.D. Soil enzyme inhibition by condensed litter tannins may drive ecosystem structure and processes: The case of Kalmia angustifolia. New. Phytol. 2007, 175, 535–546. [Google Scholar] [CrossRef]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Bending, G.D. Sequestration of soil nitrogen as tannin-protein complexes may improve the competitive ability of sheep laurel (Kalmia angustifolia) relative to black spruce (Picea mariana). New. Phytol. 2009, 181, 187–198. [Google Scholar] [CrossRef]
- Callaway, R.M.; Pennings, S.C.; Richards, C.L. Phenotypic plasticity and interactions among plants. Ecology 2003, 1115–1128. [Google Scholar]
- Hébert, F.; Thiffault, N.; Munson, A.D. Does trait plasticity of three boreal nutrient-conserving species relate to their competitive ability? Ecoscience 2011, 18, 382–393. [Google Scholar] [CrossRef]
- Taylor, C.M.A.; Tabbush, P.M. Nitrogen Deficiency in Sitka Spruce Plantations; Forestry Commission (UK): Farnham, UK, 1990; p. 20. [Google Scholar]
- Bennett, J.N.; Blevins, L.L.; Barker, J.E.; Blevins, D.P.; Prescott, C.E. Increases in tree growth and nutrient supply still apparent 10 to 13 years following fertilization and vegetation control of salal-dominated cedar-hemlock stands on Vancouver Island. Can. J. For. Res. 2003, 33, 1516–1524. [Google Scholar] [CrossRef]
- Thiffault, N.; Cyr, G.; Pregent, G.; Jobidon, R.; Charette, L. Artificial regeneration of an ericacees black spruce stand—Effects of scarification, fertilization and plant type 10 years later. For. Chron. 2004, 80, 141–149. [Google Scholar]
- Thiffault, N.; Hébert, F.; Jobidon, R. Planted Picea mariana growth and nutrition as influenced by silviculture × nursery interactions on an ericaceous-dominated site. Silva. Fenn. 2012, 46, 667–682. [Google Scholar]
- Thiffault, N.; Jobidon, R. How to shift unproductive Kalmia angustifolia–Rhododendron groenlandicum heath to productive conifer plantation. Can. J. For. Res. 2006, 36, 2364–2376. [Google Scholar] [CrossRef]
- Thiffault, N.; Titus, B.D.; Munson, A.D. Silvicultural options to promote seedling establishment on Kalmia-Vaccinium-dominated sites. Scand. J. For. Res. 2005, 20, 110–121. [Google Scholar] [CrossRef]
- Bradley, R.L.; Titus, B.D.; Preston, C.P.; Bennett, J. Improvement of nutritional site quality 13 years after single application of fertiliser N and P on regenerating cedar-hemlock cutovers on northern Vancouver Island, B.C. Plant. Soil 2000, 223, 195–206. [Google Scholar]
- Moroni, M.T.; Thiffault, N.; Titus, B.D.; Mante, C.; Makeschin, F. Controlling Kalmia and reestablishing conifer dominance enhances soil fertility indicators in central Newfoundland, Canada. Can. J. For. Res. 2009, 39, 1270–1279. [Google Scholar]
- Prévost, M. Effets du Scarifiage sur les propriÉtés du sol, la Croissance des semis et la compétition: Revue des Connaissances Actuelles et Perspectives de Recherches au Québec. Ann. Sci. For. 1992, 49, 277–296. [Google Scholar]
- Titus, B.D.; Sidhu, S.S.; Mallik, A.U. A Summary of some Studies on Kalmia Angustifolia L.: A Problem Species in Newfoundland Forestry; N-X-296; Canadian Forest Service: St. John’s, Canada, 1995; p. 68. [Google Scholar]
- Thiffault, N.; Picher, G.; Auger, I. Initial distance to Kalmia angustifolia as a predictor of planted conifer growth. New For. 2012, 43, 849–868. [Google Scholar] [CrossRef]
- Thiffault, N.; Titus, B.D.; Moroni, M.T. Silviculture and planted species interact to influence reforestation success on a Kalmia-Dominated Site—A 15-Year Study. For. Chron. 2010, 86, 234–242. [Google Scholar]
- Hawkins, C.B.D.; Steele, T.W.; Letchford, T. The economics of site preparation and the impacts of current forest policy: Evidence from central British Columbia. Can. J. For. Res. 2006, 36, 482–494. [Google Scholar] [CrossRef]
- Lorente, M.; Parsons, W.F.J.; Bradley, R.L.; Munson, A.D. Soil and plant legacies associated with harvest trails in boreal black spruce forests. For. Ecol. Manag. 2012, 269, 168–176. [Google Scholar] [CrossRef]
- Thiffault, N.; Roy, V. Living without herbicides in Québec (Canada): Historical context, current strategy, research and challenges in forest vegetation management. Eur. J. For. Res. 2011, 130, 117–133. [Google Scholar] [CrossRef]
- Yamasaki, S.H.; Fyles, J.W.; Titus, B.D. Interactions among Kalmia Angustifolia, soil characteristics, and the growth and nutrition of black spruce seedlings in two boreal newfoundland plantations of contrasting fertility. Can. J. For. Res. 2002, 32, 2215–2224. [Google Scholar] [CrossRef]
- Hébert, F.; Thiffault, N.; Ruel, J.-C.; Munson, A.D. Ericaceous shrubs affect black spruce physiology independently from inherent site fertility. For. Ecol. Manag. 2010, 260, 219–228. [Google Scholar] [CrossRef]
- Burton, P.J. Some limitations inherent to static indices of plant competition. Can. J. For. Res. 1993, 23, 2141–2152. [Google Scholar] [CrossRef]
- Rheault, H. Contrer l’envahissement par les éricacées. Fascicule 4.10. In Manuel de détermination des possibilités forestières 2013–2018; Bureau du forestier en chef, Gouvernement du Québec: Roberval, Canada, 2013; in press. [Google Scholar]
- Franklin, S.E.; Gillespie, R.T.; Titus, B.D.; McCaffrey, T.M. Discrimination of Kalmia angustifolia using Compact Airborne Spectrographic Imager (CASI) data. Can. J. Remote Sens. 1997, 23, 71–75. [Google Scholar]
- Franklin, S.E.; Gillespie, R.T.; Titus, B.D.; Pike, D.B. Aerial and satellite sensor detection of Kalmia angustifolia at forest regeneration sites in central Newfoundland. Int. J. Remote Sens. 1994, 15, 2533–2557. [Google Scholar]
- van Lier, O.; Fournier, R.A.; Bradley, R.L.; Thiffault, N. A multi-resolution satellite imagery approach for large area mapping of ericaceous shrubs in Northern Quebec, Canada. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 334–343. [Google Scholar] [CrossRef]
- Boucher, Y.; Arseneault, D.; Sirois, L. Logging-induced change (1930–2002) of a preindustrial landscape at the northern range limit of northern hardwoods, eastern Canada. Can. J. For. Res. 2006, 36, 505–517. [Google Scholar] [CrossRef]
- Bureau du forestier en chef, Bilan d’aménagement forestier durable (2000–2008); Gouvernement du Québec: Roberval, Canada, 2010; p. 290.
- Silvola, J. Moisture dependance of CO2 exchange and its recovery after drying in certain boreal forest peat mosses. Lindbergia 1991, 17, 5–10. [Google Scholar]
- Groot, A.; Adams, M.J. Direct seeding black spruce on peatlands: Fifth-year results. For. Chron. 1994, 70, 585–592. [Google Scholar]
- Lavoie, M.; Paré, D.; Bergeron, Y. Relationships between microsite type and the growth and nutrition of young black spruce on post-disturbed lowland black spruce sites in eastern Canada. Can. J. For. Res. 2007, 37, 62–73. [Google Scholar] [CrossRef]
- Lafleur, B.; Paré, D.; Fenton, N.J.; Bergeron, Y. Growth and nutrition of black spruce seedlings in response to disruption of Pleurozium and Sphagnum moss carpets in boreal forested peatlands. Plant. Soil 2011, 345, 141–153. [Google Scholar] [CrossRef]
- Lavoie, M.; Paré, D.; Bergeron, Y. Quality of growth substrates of post-disturbed lowland black spruce sites for black spruce (Picea mariana) seedling growth. New For. 2007, 33, 207–216. [Google Scholar] [CrossRef]
- DeLuca, T.; Zackrisson, O.; Nilsson, M.; Sellstedt, A. Quantifying nitrogen-fixation in feather moss carpets of boreal forests. Nature 2002, 419, 917–920. [Google Scholar] [CrossRef]
- Näsholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y.; Paré, D. Decomposition rates of bryophytes in managed boreal forests: Influence of bryophyte species and forest harvesting. Plant. Soil 2010, 336, 499–508. [Google Scholar] [CrossRef]
- Lang, S.I.; Cornelissen, J.H.C.; Klahn, T.; van Logtestijn, R.S.P.; Broekman, R.; Schweikert, W.; Aerts, R. An experimental comparison of chemical traits and litter decomposition rates in a diverse range of subarctic bryophyte, lichen and vascular plant species. J. Ecol. 2009, 97, 886–900. [Google Scholar] [CrossRef]
- Davey, M.L.; Currah, R.S. Interactions between mosses (Bryophyta) and fungi. Can. J. Bot. 2006, 84, 1509–1519. [Google Scholar] [CrossRef]
- Fenton, N.; Lecomte, N.; Légaré, S.; Bergeron, Y. Paludification in black spruce (Picea mariana) forests of eastern Canada: Potential factors and management implications. For. Ecol. Manag. 2005, 213, 151–159. [Google Scholar] [CrossRef]
- Lecomte, N.; Bergeron, Y. Successional pathways on different surficial deposits in the coniferous boreal forest of the Quebec Clay Belt. Can. J. For. Res. 2005, 35, 1984–1995. [Google Scholar] [CrossRef]
- Légaré, S.; Paré, D.; Bergeron, Y. Influence of aspen on forest floor properties in black spruce-dominated stands. Plant Soil 2005, 275, 207–220. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bradley, R.L. Linking the abundance of aspen with soil faunal communities and rates of belowground processes within single stands of mixed aspen-black spruce. Appl. Soil Ecol. 2009, 41, 19–28. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y. Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. J. Veg. Sci. 2006, 17, 65–76. [Google Scholar] [CrossRef]
- Williams, T.; Flanagan, L. Measuring and modelling environmental influences on photosynthetic gas exchange in Sphagnum and Pleurozium. Plant. Cell. Environ. 1998, 21, 555–564. [Google Scholar] [CrossRef]
- Bisbee, K.; Gower, S.; Norman, J.; Nordheim, E. Environmental controls on ground cover species composition and productivity in a boreal black spruce forest. Oecologia 2001, 120, 261–270. [Google Scholar]
- Lecomte, N.; Simard, M.; Bergeron, Y. Effects of fire severity and initial tree composition on stand structural development in the coniferous boreal forest of northwestern Quebec, Canada. Ecoscience. 2006, 13, 152–163. [Google Scholar] [CrossRef]
- Belleau, A.; Leduc, A.; Lecomte, N.; Bergeron, Y. Forest succession rate and pathways on different surface deposit types in the boreal forest of northwestern Quebec. Ecoscience 2011, 18, 329–340. [Google Scholar] [CrossRef]
- Fenton, N.J.; Simard, M.; Bergeron, Y. Emulating natural disturbances: The role of silviculture in creating even-aged and complex structures in the black spruce boreal forest of eastern North America. J. For. Res. 2009, 14, 258–267. [Google Scholar] [CrossRef]
- Greene, D.F.; Macdonald, S.E.; Haeussler, S.; Domenicano, S.; Noel, J.; Jayen, K.; Charron, I.; Gauthier, S.; Hunt, S.; Gielau, E.T.; Bergeron, Y.; Swift, L. The reduction of organic-layer depth by wildfire in the North American boreal forest and its effect on tree recruitment by seed. Can. J. For. Res. 2007, 37, 1012–1023. [Google Scholar] [CrossRef]
- Lafleur, B.; Fenton, N.J.; Paré, D.; Simard, M.; Bergeron, Y. Contrasting effects of season and method of harvest on soil properties and the growth of black spruce regeneration in the boreal forested peatlands of eastern Canada. Silva. Fenn. 2010, 44, 799–813. [Google Scholar]
- Renard, S. Impact du brûlage dirigé comme préparation de terrain sur l'état de paludification et la croissance de l'épinette noire dans les pessières à mousses paludifiées de la ceinture d'argile. Master’s Thesis, Université du Québec en Abitibi-Témiscamingue, Rouyn-Noranda, Canada, 2009. [Google Scholar]
- Laamrani, A.; Valeria, O.; Cheng, L.-Z.; Bergeron, Y.; Camerlynck, C. The use of ground penetrating radar for remote sensing the organic layer-mineral soil interface in paludified boreal forests. Can. J. Remote Sens. 2013, 39, 1–15. [Google Scholar] [CrossRef]
- Simard, M.; Bernier, P.Y.; Bergeron, Y.; Paré, D.; Guérine, L. Paludification dynamics in the boreal forest of the James Bay Lowlands: Effect of time since fire and topography. Can. J. For. Res. 2009, 39, 546–552. [Google Scholar] [CrossRef]
- Lavoie, M.; Harper, K.; Paré, D.; Bergeron, Y. Spatial pattern in the organic layer and tree growth: A case study from regenerating Picea mariana stands prone to paludification. J. Veg. Sci. 2007, 18, 213–222. [Google Scholar]
- Shetler, G.; Turetsky, M.R.; Kane, E.; Kasischke, E. Sphagnum mosses limit total carbon consumption during fire in Alaskan black spruce forests. Can. J. For. Res. 2008, 38, 2328–2336. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y. Stochastic processes dominate during boreal bryophyte community assembly. Ecology 2013, in press. [Google Scholar]
- Lavoie, M.; Paré, D.; Bergeron, Y. Impact of global change and forest management on carbon sequestration in northern forested peatlands. Environ. Rev. 2005, 13, 199–240. [Google Scholar] [CrossRef]
- Drobyshev, I.; Simard, M.; Bergeron, Y.; Hofgaard, A. Does soil organic layer thickness affect climate-growth relationships in the black spruce boreal ecosystem? Ecosystems 2010, 13, 556–574. [Google Scholar] [CrossRef]
- Nappi, A. Paludification. Fascicule 4.9. In Manuel de détermination des possibilités forestières 2013–2018; Bureau du forestier en chef, Gouvernement du Québec: Roberval, Canada, 2013. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F. Conserving Forest Biodiversity. A Comprehensive Multiscale Approach; Island Press: Washington, DC, USA, 2002; p. 351. [Google Scholar]
- Swanson, F.J.; Sparks, R.E. Long-term ecological research and the invisible place. Bioscience 1990, 40, 502–508. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Thiffault, N.; Fenton, N.J.; Munson, A.D.; Hébert, F.; Fournier, R.A.; Valeria, O.; Bradley, R.L.; Bergeron, Y.; Grondin, P.; Paré, D.; et al. Managing Understory Vegetation for Maintaining Productivity in Black Spruce Forests: A Synthesis within a Multi-Scale Research Model. Forests 2013, 4, 613-631. https://doi.org/10.3390/f4030613
Thiffault N, Fenton NJ, Munson AD, Hébert F, Fournier RA, Valeria O, Bradley RL, Bergeron Y, Grondin P, Paré D, et al. Managing Understory Vegetation for Maintaining Productivity in Black Spruce Forests: A Synthesis within a Multi-Scale Research Model. Forests. 2013; 4(3):613-631. https://doi.org/10.3390/f4030613
Chicago/Turabian StyleThiffault, Nelson, Nicole J. Fenton, Alison D. Munson, François Hébert, Richard A. Fournier, Osvaldo Valeria, Robert L. Bradley, Yves Bergeron, Pierre Grondin, David Paré, and et al. 2013. "Managing Understory Vegetation for Maintaining Productivity in Black Spruce Forests: A Synthesis within a Multi-Scale Research Model" Forests 4, no. 3: 613-631. https://doi.org/10.3390/f4030613
APA StyleThiffault, N., Fenton, N. J., Munson, A. D., Hébert, F., Fournier, R. A., Valeria, O., Bradley, R. L., Bergeron, Y., Grondin, P., Paré, D., & Joanisse, G. (2013). Managing Understory Vegetation for Maintaining Productivity in Black Spruce Forests: A Synthesis within a Multi-Scale Research Model. Forests, 4(3), 613-631. https://doi.org/10.3390/f4030613