Abstract
Canopy structure affects forest function by determining light availability and distribution. Many forests throughout the upper Great Lakes region are dominated by mature, even-aged, early successional aspen and birch, which comprise 35%–40% of canopy leaf area, and which are senescing at accelerating rates. In 2008 at the University of Michigan Biological Station, we initiated the Forest Accelerated Succession ExperimenT (FASET) by stem girdling all aspen and birch in replicated stands to induce mortality. Our objective was to understand type and rate of canopy structural changes imposed by rapid but diffuse disturbance consisting of mortality of a single age-species cohort. We characterized changes in canopy structural features in 2008–2011 using ground-based Portable Canopy Lidar (PCL) in paired treated and control stands. As aspen and birch in treated plots died, gap fraction of the upper canopy increased, average leaf height decreased, total canopy height declined, and openness of the whole-canopy increased. All of these trends became more pronounced with time. Our findings suggest that as forests throughout the region pass through the impending successional transition prompted by widespread mortality of canopy-dominant early successional aspen and birch species, the canopy will undergo significant structural reorganization with consequences for forest carbon assimilation.
1. Introduction
Canopy structure influences forest carbon (C) gain by influencing the amount of light available to all but the uppermost canopy foliage, and the distribution of that light within the canopy. Numerous structural features of forest canopies directly affect light distribution and interception throughout the depth of the canopy with consequences for canopy photosynthesis. Repeated, small-scale canopy disturbances serve to increase canopy structural complexity as forests age [1,2,3,4] and partial canopy disturbance has been suggested as a mechanism for sustaining productivity in aging forests by reducing intensity of competition for light across the landscape [5,6,7]. Partial canopy disturbances are inevitable throughout succession, and in stands with clumped age distributions of canopy trees are likely to manifest as pulsed-mortality events. These less-intense canopy disturbances can reorganize leaf area in the canopy, increasing heterogeneity of foliage distribution [8,9]. Disturbances which kill some fraction of canopy trees (via defoliation, storm damage, pathogens, or age-related senescence) alter canopy structure over time [10] in ways that influence availability and distribution of productivity-controlling resources such as water, light, and nitrogen (N) [11,12,13,14].
Up to 100,000 km2 of forests throughout the upper Great Lakes region are dominated by aspen (Populus grandidentata Michx. and P. tremuloides Michx.) and birch (Betula papyrifera Marsh) [15]. In this region, these early successional species are even-aged and usually among the oldest and tallest trees in these forests due to a legacy of anthropogenic clear-cutting and burning a century ago [16,17]. At the University of Michigan Biological Station (UMBS) in northern lower Michigan, USA, aspen and birch together comprise ~39% of forest basal area and 35% of leaf area [18,19]. Aspen and birch, regeneration niches, however, tend to favor recently disturbed areas and do not recruit successfully into the understory of forests with intact canopies and litter layers [20,21,22]. Contemporary management strategies and disturbance regimes throughout the region do not support the continuation or renewal of widespread aspen-birch dominated forests [23,24,25]. Aspen and birch in these forests are senescing at an accelerating rate as they reach maturity [18] and their increased mortality and eventual elimination from the forest will trigger significant changes to canopy structure.
Our objective was to evaluate the rate, magnitude, and primary forms of canopy structural change associated with an experimentally induced forest mortality rate of ~35%–40%. We relate these structural reorganizations to functional data from other studies at this site, which indicate high resilience of C storage to moderate disturbance. These previous analysis at our site focused on functional changes in response to the treatment [19,26]. Here, we emphasize underlying structural changes supporting the functional resilience reported by these studies. These changes to canopy structure were expected to occur simultaneously with aspen and birch mortality and to produce lasting structural rearrangement of the canopy. Since aspen and birch are among the oldest and tallest trees in the canopy and by virtue of their shade intolerance do not retain leaf area within the lower levels of the canopy, we hypothesized that structural effects of widespread aspen and birch mortality should be more pronounced in the upper canopy of treated stands and most prevalent in stands with high aspen-birch density. We hypothesized that average height of leaf area throughout the canopy and average height of maximum leaf area density would both decline. Further, openness of the canopy (gap fraction of upper canopy, canopy porosity, and the amount of sky visible through the canopy) should increase. We also predicted increased spatial heterogeneity of various features of canopy structure (variability of mean leaf height, variability of leaf height variability, and variability of canopy sky fraction) across the landscape with aspen-birch mortality since these species have a patchy distribution across the landscape [26]. We used a ground-based Portable Canopy Lidar (PCL) system to characterize canopy structural changes following widespread, experimentally accelerated mortality of canopy dominant aspen and birch.
2. Experimental Section
2.1. Site Description and Experimental Design
We conducted this study from 2008 through 2011 at the University of Michigan Biological Station (UMBS) located in northern lower Michigan (45°35.5' N, 84°43' W). This site in the Upper Great Lakes region is located in the transition zone between Northern mixed hardwood and boreal forests with similar species composition and age structure as other aspen-dominated forests in the upper Midwest [15]. Early successional aspen (Populus grandidentata Michx. and P. tremuloides Michx.) and birch (Betula papyrifera Marsh) dominate these stands with substantial representation of co- or sub-dominant northern red oak (Quercus rubra L.), red maple (Acer rubrum L.), white pine (Pinus strobus L.), and American beech (Fagus grandifolia Ehrh.) [27]. Forests throughout this region were clear-cut and subject to slash-fueled wildfires in the early 1900s [15,20], producing contemporary forests that are relatively young and even aged [28]. Mean annual (1942–2003) temperature is 5.5 °C and mean annual precipitation is 817 mm. Soils in the study area are coarse grained, excessively drained mixed frigid Entic Haplorthods with minimal relief [29] and have been described in detail in previous publications [19,30].
2.2. Forest Accelerated Succession ExperimenT (FASET)
In spring 2008, prior to leaf-out, we girdled all aspen (Populous spp.) and birch (B. papyrifera) trees (>6700 stems throughout the treated area) to induce mortality and accelerate succession in a 33 ha stand as described in Nave et al. [19]. We similarly girdled aspen in three 2 ha replicate stands. Girdled species comprise an average of 35%–40% of the leaf area in both treated and control stands (range: 14%–48%) [26]. An adjacent forest serves as control for this experiment. Both sites are arranged around a central eddy covariance tower measuring exchanges of carbon, water, and energy between forest and atmosphere [31]. Each tower is surrounded by a 1.1 ha circular plot (60 m radius), and transects radiate from the towers at 15° and 20° intervals in control and treatment stands, respectively. In 0.08 ha plots (16 m radius) located at 100 m intervals along each transect, we conducted a suite of measurements quantifying landscape-level variation in forest responses to disturbance. Sixteen (8 treatment, 8 control) of these plots were paired using of a Principle Components Analysis (PCA) that formed matches based on similarity in stand-level pre-treatment production and canopy tree species composition [19].
2.3. Canopy Structure (Portable Canopy Lidar)
We used a portable canopy lidar system to quantify canopy structural changes following stem girdling of all aspen and birch. The original PCL system design and principles of operation are described in Parker et al. [2]; modifications of the system for use at our site are given in Hardiman et al. [32,33]. Additional details on processing of data and derivation of canopy structural metrics are given here.
We deployed the PCL system in PCA-paired treated and control plots. PCL data were not available for one treated plot, introducing minor imbalance (n = 7 treated, 8 control). All PCL data was collected from 40 m long transects (~2× canopy height) spanning the diameter of each 16 m radius plot present in treated and control areas. Each transect was flagged at 10 m intervals along its length to allow PCL raw data to be aligned with plot dimensions. PCL raw data consisted of ASCII strings of return height values interspersed with data markers corresponding to physical locations of marker flags along each PCL transect. Raw data was processed in Matlab (MathWorks, Natick, MA, USA) by binning raw PCL return heights into 1 m2 bins (Figure 1). Return data were divided evenly among a number of 1 m wide columns equal to the distance between marker flags. This operation assumed that the PCL operator walked a constant pace between marker flags. Heights of lidar returns from the canopy within each column were then grouped into bins based on height. All PCL returns originating within the boundaries of a bin were assigned the mean height of that bin (e.g., return heights of 9.25 m and 9.75 m were grouped into a bin spanning from 9.0 to 10.0 m and were considered to have a bin height of 9.5 m). We then calculated several canopy structural metrics describing numerous features of canopy structure based on spatial heterogeneity of density and distribution of returns along vertical and horizontal axes and within a 2D vertical plane transecting the canopy (Table 1). Lidar returns from each transect include canopy surface area from both girdled and ungirdled species. Since branch area is a relatively small fraction of total canopy surface area in these and other deciduous stands [34], we considered lidar return distributions as indicative of leaf area distribution in the canopy. We interpreted regions of the canopy that produced denser distribution of returns to have denser canopy surface area.
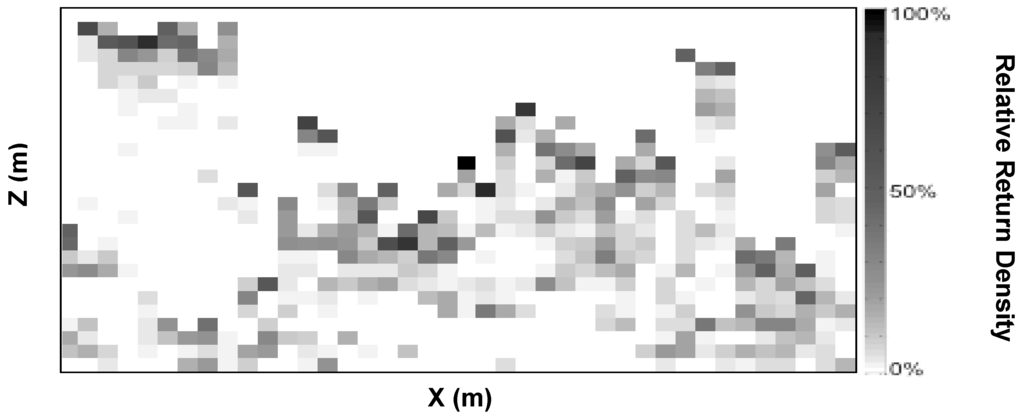
Figure 1.
Vertical canopy cross-section illustrating portable canopy lidar data. Bin (1 m2) shading is proportional to number of lidar returns. Canopy structural metrics were calculated from rows and columns of bins.

Table 1.
Description and method of calculation for 13 metrics of canopy structure derived from PCL data. Parameters were calculated by applying the indicated statistics in the following order: Parameter = transect(column[bin × weight]). For example, modeEl is calculated as the plot mean of the mode of count-weighted return heights in each of 40 columns comprising the transect spanning the plot. “Sky hits” refers to non-returned lidar pulses.
2.4. Statistical Analysis
Statistical analysis of canopy structural changes was performed on paired treatment and control plots. The plot was the level of replication within each treatment block. All structural metrics were calculated as summary statistics of the population of PCL canopy returns from the 40 m transect spanning each plot. Means and standard errors were calculated among plots within treatment blocks in each year. Standard errors thus represent within-treatment variation. Clumping index was based on a modified method for calculating clumping index for hemispherical canopy photographs [35] and was calculated as natural log of the average gap fraction at each canopy height (z) [across the length of the transect (x)] divided by the average natural log of gap fraction at each height (across the length of the transect; see Equation 3 in Gonsamo et al. [35]). We used paired t-tests assuming unequal variance (JMP 9.0; SAS Institute, Cary, NC, USA) to evaluate differences between treatments and years. We considered differences significant at p ≤ 0.1.
3. Results
3.1. Canopy Openness
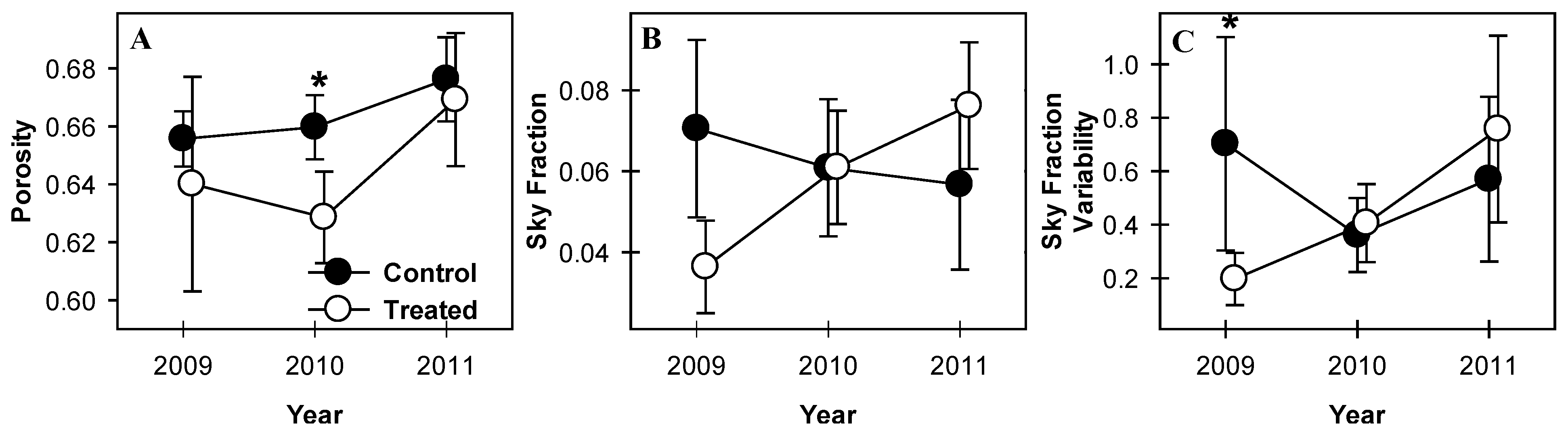
Canopies in treated plots became more open as aspen and birch senesced. The vertical distribution of canopy gap fraction in control plots was not significantly different between years (p > 0.1, Figure 2A). Treated plots, however, showed significant increases in gap fraction from 2009 and 2010 to 2011 at 19 m (p = 0.03 and 0.02 from 2009 and 2010 to 2011, respectively) and 21–22 m (p = 0.06 for both heights in both years; Figure 2B). Gap fraction decreased from 2009 to 2010 but increased from 2010 to 2011 at 15 m (p = 0.08 and 0.02, respectively; Figure 2B) such that gap fraction in 2011 was greater than in 2009. Clumping index was initially greater in control plots but declined in 2010 and 2011 and was not different from treated plots in 2011 (Figure 3C). While mean clumping index of canopies was unchanged from year to year, among-plot variability in treated stands (indicated by standard error) increased in all years following girdling, a trend not seen in control plots.
Whole-canopy porosity of treated plots did not change from 2009 to 2010 but increased from 2010 to 2011 (p = 0.09; Figure 3A). Porosity of treated plots was lower than control plots in 2010 (p = 0.07), the only year when a significant difference was observed between treated and control plots. Sky fraction and variability of sky fraction in treated plots both increased from 2009 to 2011 (p = 0.03 and 0.08 for sky fraction and sky fraction variability respectively, Figure 3B,C) and sky fraction was significantly greater in control plots in 2009 than in treated plots (p = 0.1). Sky fraction variability was not different between treated and control plots in any year (p > 0.1; Figure 3C). Sky fraction declined monotonically but not significantly in control plots from 2009 to 2011 and variability of sky fraction in control plots was similarly stable across years. The number of gaps extending from forest floor to canopy surface did not change significantly over time in either control or treated plots and did not differ between treatment blocks in any year (p > 0.1, not shown).
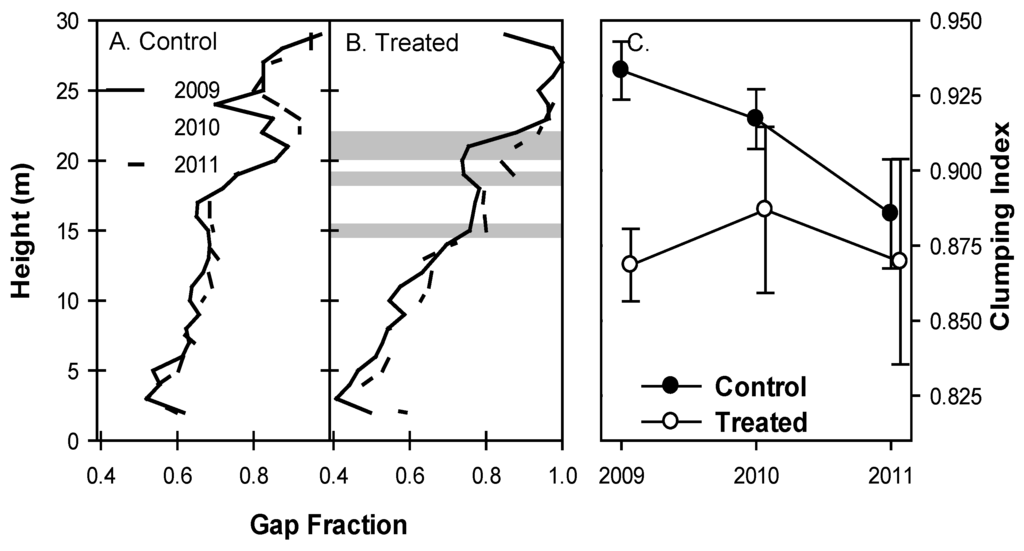
Figure 2.
Canopy gap fraction in control plots. (A) shows no significant increases from year to year, but gap fraction in the upper canopy of treated plots. (B) increased from 2009 to 2011. Standard error bars and 95% confidence intervals overlap substantially (not presented) in unshaded areas. Among-plot variability of clumping index (C) increased in treated plots with time but was not different from control plots in 2010 or 2011. Standard error bars represent between-plot variation.
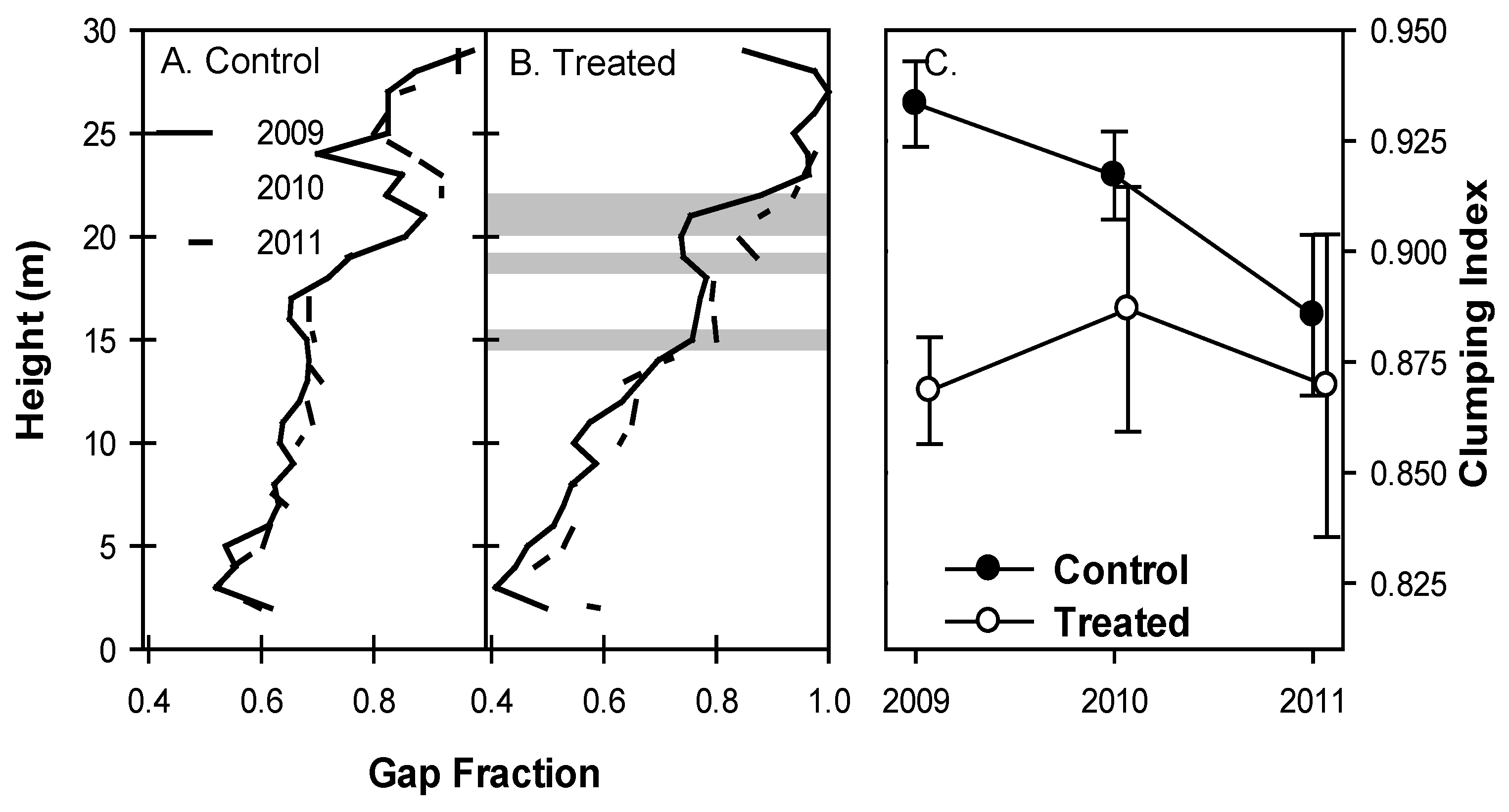
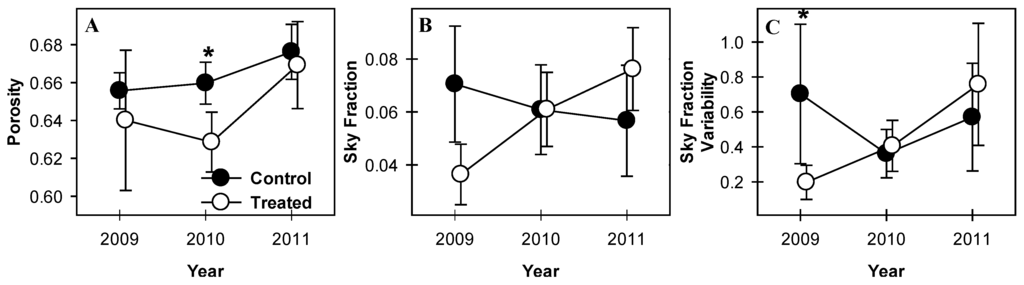
Figure 3.
Porosity and canopy sky fraction mean and variation. Porosity (A) of treated plots temporarily declined from 2009 to 2010 and was significantly lower than control stands in 2010. Sky fraction (B) and variability of sky fraction (C) increased in treated stands in all years following girdling. Values are means of paired treated and control plots with standard error bars representing between-plot variation. * indicate significant differences between treated and control plot means.
3.2. Foliage Height and Density
Metrics of height consistently documented significant reduction of canopy height accompanying mortality of canopy-dominant aspen and birch. Mean leaf height (Mean Return Height) and height of maximum leaf density (modeEl) (Figure 4A,B) declined significantly from 2008 to 2009 (p = 0.08, 0.1 for mean return height and modeEl, respectively) and from 2010 to 2011 (p = 0.09, 0.03 mean return height and modeEl, respectively) in treated but not control plots indicating a net downward shift of leaf area following girdling. Both structural metrics were significantly different in 2011, post-treatment, than in 2008 (p = 0.03, 0.01 for mean return height and modeEl, respectively; Figure 4A,B), and both were significantly lower in treated than in control plots in 2011 (p = 0.05, 0.04 for mean return height and modeEl, respectively). Mean height of maximum leaf area density (Mode2) declined in treated plots from 2008 to 2011 but was only significantly different from control plots in 2011 (p = 0.09; Figure 4C). Most of the change in treated plots occurred from 2009 to 2010 but this change was not significant (p > 0.1). Mean maximum canopy height did not change between years in control plots but did decline significantly from 2008 to 2011 in treated plots (p = 0.04; Figure 4D). Mean maximum canopy height was not different between treated and control plots in any year (p > 0.1).
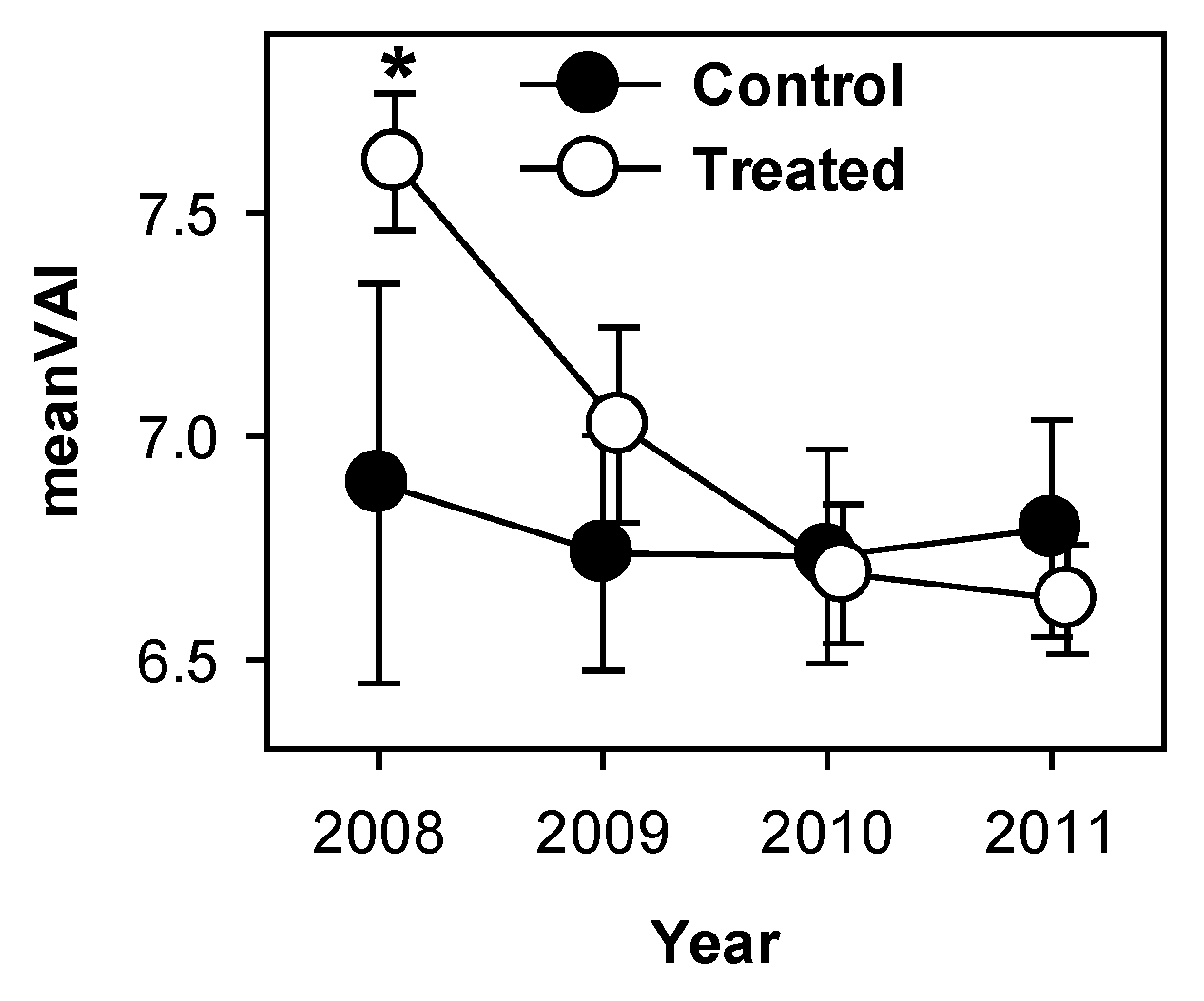
PCL-derived meanVAI (vegetation area index), an indicator of total canopy surface area, declined significantly in treated plots following treatment (p < 0.001) but remained static in control plots over the same period. MeanVAI differed between treated and control plots in 2008 (p = 0.09) but not in any other year (Figure 5).
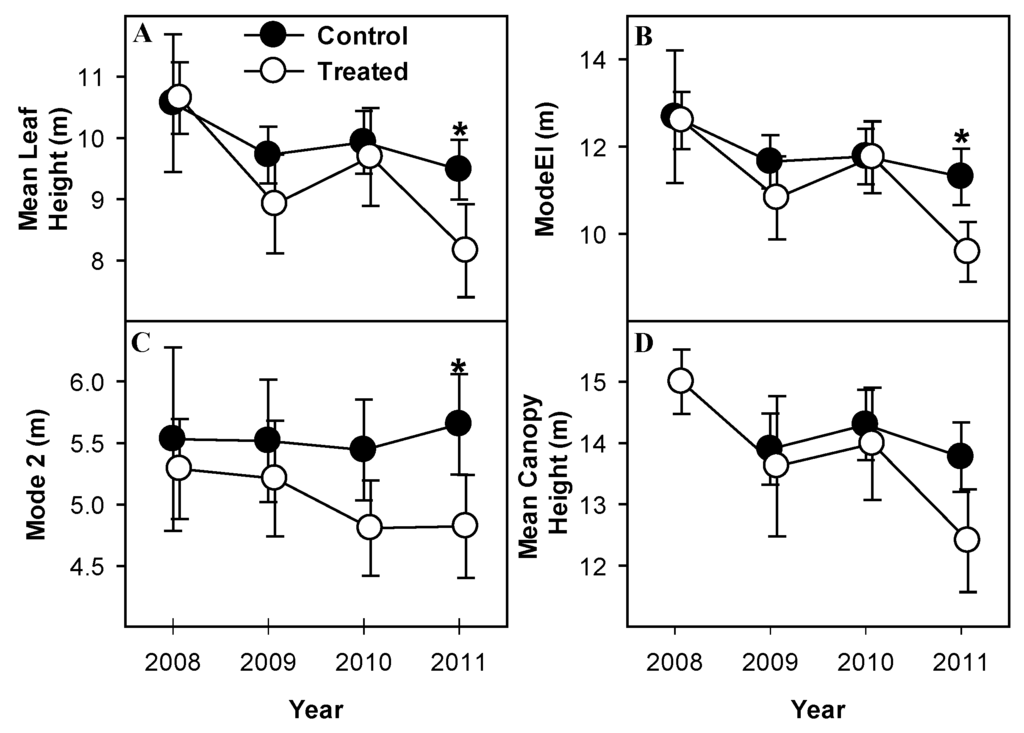
Figure 4.
Metrics of canopy height demonstrate reductions in height of girdled stands. Mean leaf height (A) height of maximum leaf density (modeEl) (B) mean height of maximum leaf area density (Mode2) (C) and mean maximum canopy height, (D), all indicate greater declines in height in treated stands than control stands. Values are means of paired treated and control plots with standard error bars representing between-plot variation. * indicate significant differences between treated and control plot means.
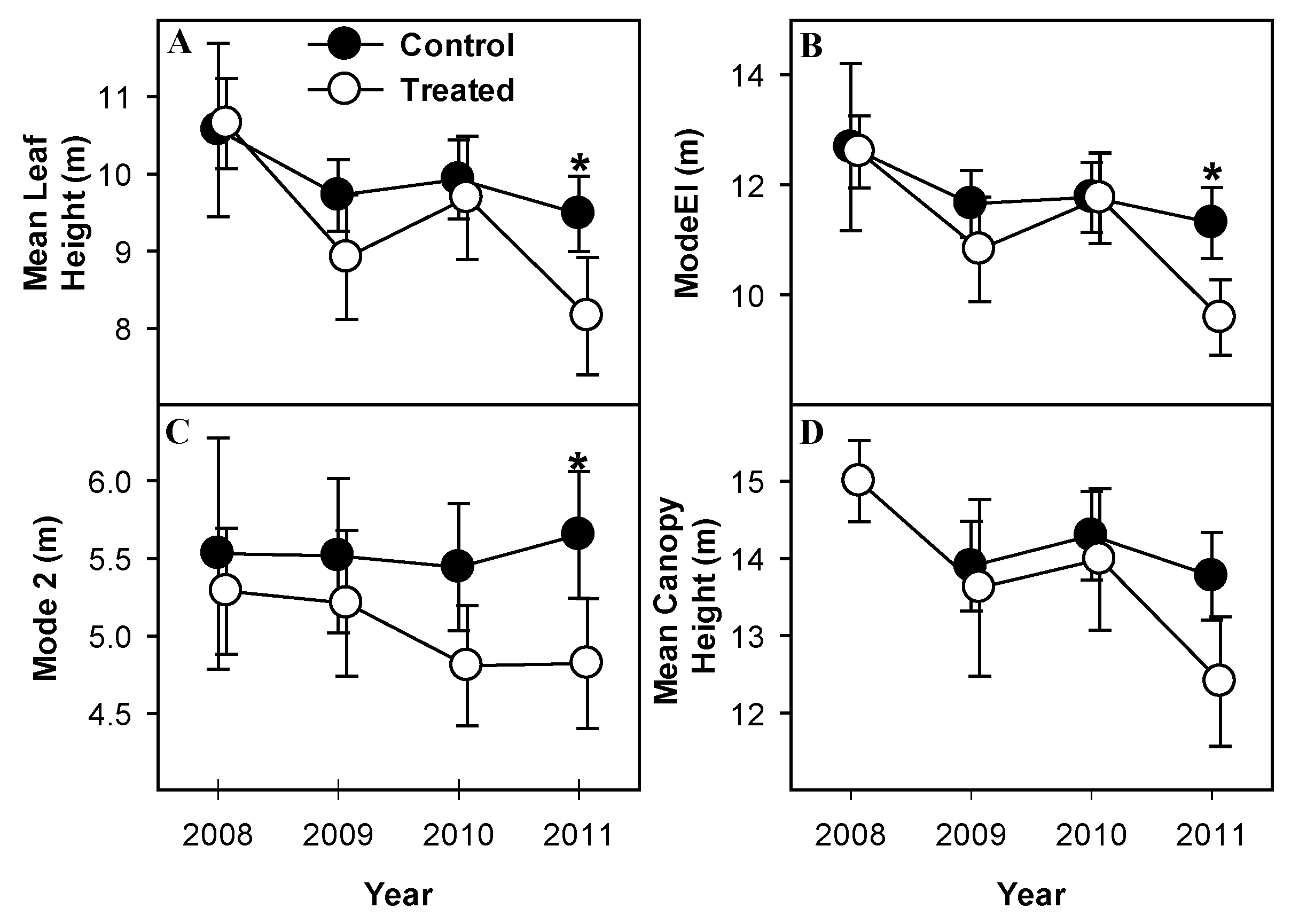
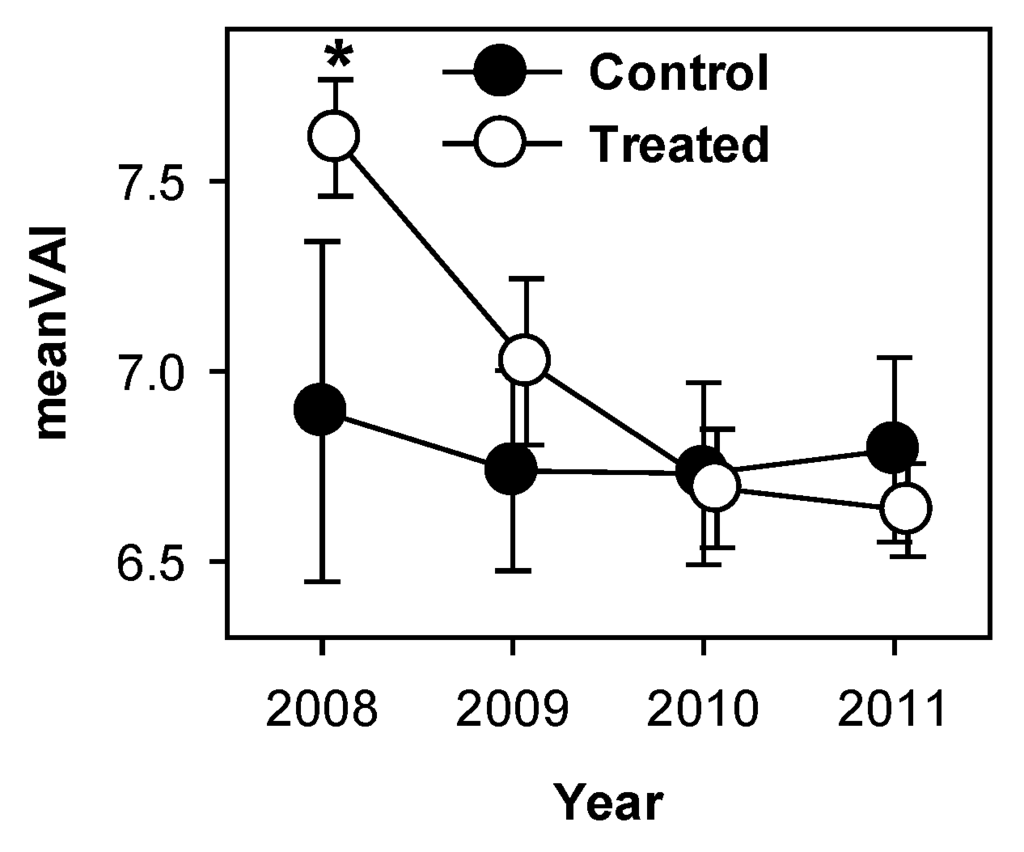
Figure 5.
MeanVAI (vegetation area index) declined in treated, but not in control plots following girdling. * indicate significant differences between treated and control plot means.
3.3. Canopy Structural Heterogeneity
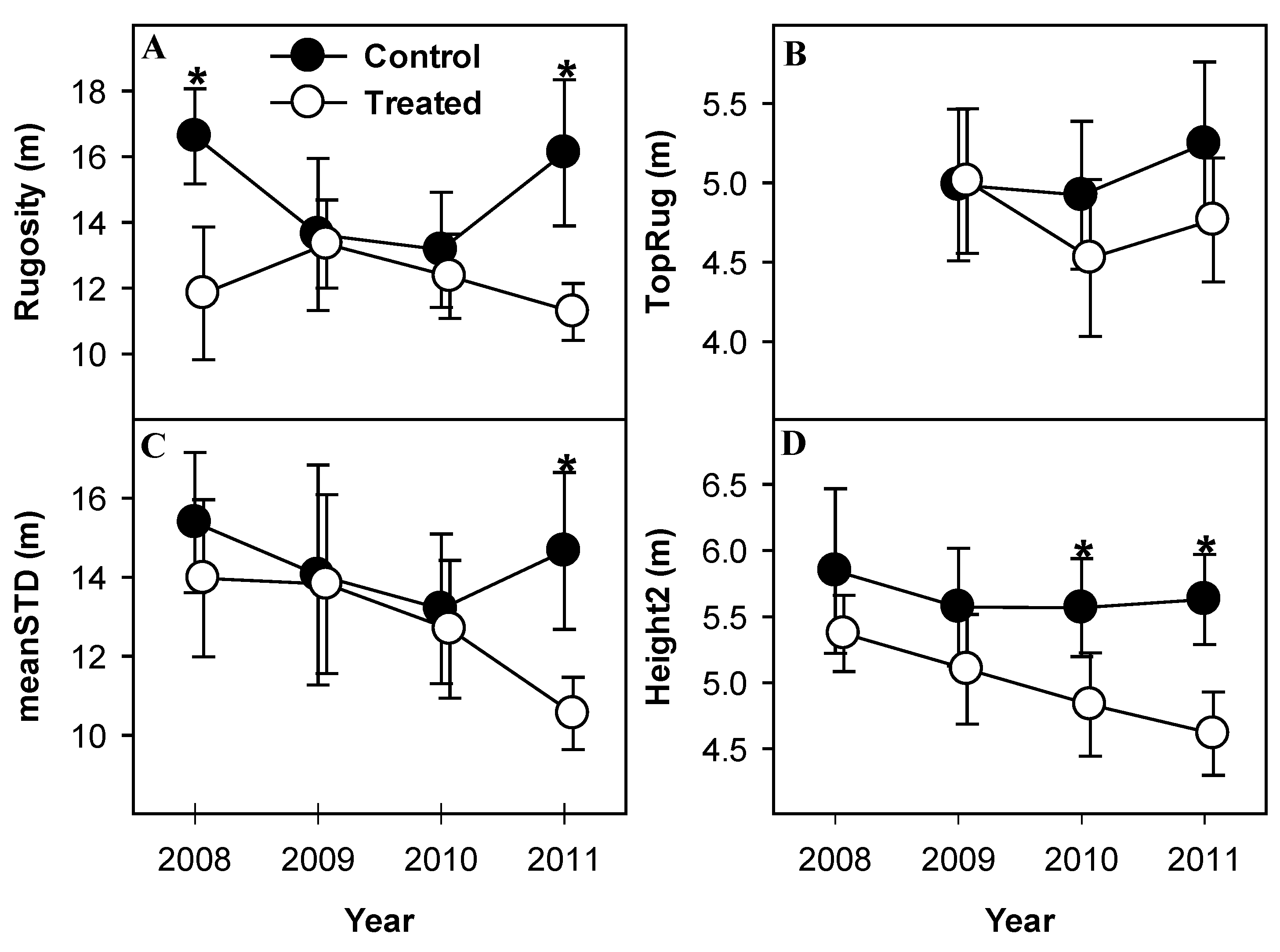
Rugosity, an index of canopy structural heterogeneity, was significantly lower in treated than control plots in 2008 and 2011 (p = 0.04, both years), but not 2009 and 2010 (p > 0.1, both years; Figure 6A), and showed no net change over four years in treated or control plots. Rugosity of control plots showed greater magnitude of inter-annual variability than treated plots, despite not differing significantly between years. Height variability of the upper canopy surface (TopRug) changed between years but showed no significant response to treatment in any year, or between any years within either treated or control plots (p > 0.1, Figure 6B). Mean variability of leaf height (meanSTD) declined monotonically from 2008 to 2011 in treated plots. Control plots mirrored this pattern from 2008 to 2010, but reversed in 2010 resulting in significant divergence from treated plots in 2011 (p = 0.05, Figure 6C). MeanSTD in treated plots was lower in 2011 than 2008 (p = 0.08), indicating that mean leaf height in treated plots was less variable in 2011 than in 2008, but control plots did not differ between any years. Variability of mean leaf height (Height2) in treated plots declined gradually from 2008 to 2011, diverging significantly from control plots in 2010 and 2011 (p = 0.1, 0.02 in 2010 and 2011, respectively; Figure 6D). Height2 in control plots exhibited no net change over the observation period.
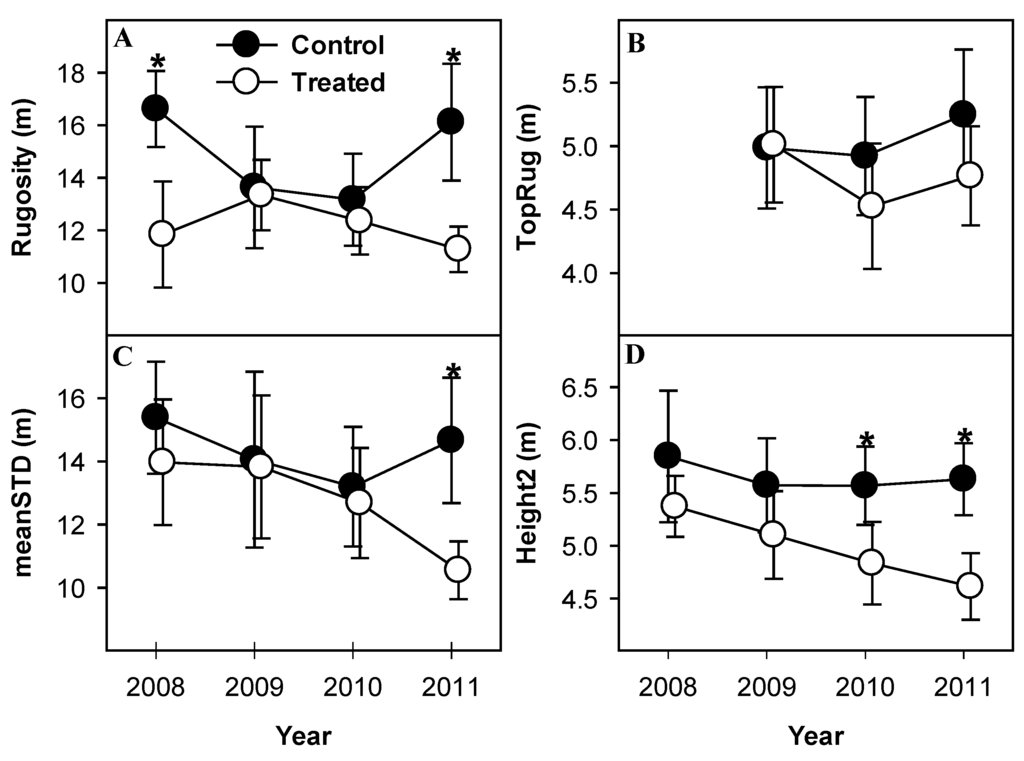
Figure 6.
Canopy heterogeneity following girdling as indicated by canopy structural complexity (rugosity) (A), variability of outer canopy surface height (TopRug) (B), mean variability of leaf height (meanSTD) (C), and variability of mean leaf height (height2) (D). Treated canopies became less structurally heterogeneous according to all metrics except TopRug. Values are means of paired treated and control plots with standard error bars representing between-plot variation. * indicate significant differences between treated and control plot means.
4. Discussion
Most studies that measure forest or canopy structure define stand structural metrics such as stem diameter size classes, stem density, crown diameter, and canopy height [36,37]. While these are certainly valid descriptors of stand structure, they are more often consequences of edaphic site factors or longer-term forest demographic trends than of canopy responses to recent disturbance. In this study we define several features of the 3-dimentional canopy structure based on spatial distribution of leaves as measured by lidar returns from the canopy [2,38]. These features relate more directly to forest function through their influence on light distribution and interception. Canopy structural metrics described here proved capable of detecting canopy structural shifts occurring from one year to the next as a result of diffuse mortality in a mixed deciduous forest canopy. Importantly, these metrics were defined a priori using simple summary statistics of density and distribution of canopy surface area to test our hypotheses regarding structural changes associated with aspen and birch mortality. Structural metrics in both treated and control plots illustrated interannual variability not attributable to treatment effects alone. Insect defoliation, stochastic individual mortality, and crown damage are likely leading causes of this variation. Overall, control plots were less variable from year-to-year than treated plots. Interannual variability observed in control plots underscores the importance and value of multi-year observations of canopy structure. We observed several structural changes likely to influence forest ecosystem C dynamics throughout the upper Great Lakes region.
4.1. Type, Rate, and Magnitude of Canopy Structural Changes
Canopies throughout the treated area became more open, especially at the top of the canopy, following aspen and birch mortality (Figure 2A,B). Gap fraction of the upper canopy increased significantly in treated plots, providing support for our hypothesis that the most pronounced effects of aspen and birch mortality would occur in the upper canopy where these shade-intolerant species allocate the majority of their leaf area. Control plots did not show any systematic or significant change in gap fraction at any height in the same period. We defined gap fraction very strictly: gaps were only counted when a 1 m2 bin was completely devoid of any lidar returns. Thus gap fraction itself is highly conservative, and probably an underestimate of canopy openness following treatment. This definition of a gap may permit less inference on the cause of any change in gap fraction but is preferable to less conservative definitions because its interpretation is unambiguous: increases in gap fraction in treated plots following girdling indicate an absence of biomass in canopy regions previously occupied.
Among-stand variability of clumping index of treated stands increased while the mean was unchanged (Figure 2C), indicating that intensity of disturbance posed by aspen and birch mortality was not uniform across the treated area. Given the patchy distribution of clonal aspen trees in the study area [20], this is not surprising. The canopy of treated stands also became more porous as foliage clumping became more variable (Figure 3A) and PCL returns indicated increasing numbers of non-returned pulses (“sky-hits”; Figure 3B,C). These increases in openness of the upper canopy corresponded with temporary reductions in LAI of all species observed from litter traps in 2010 in both treated and control stands [26] and likely increased light availability to previously light-limited foliage lower in the canopy. Indeed, other forest disturbances resulting in partial canopy mortality (e.g., due to spruce budworm defoliation of a balsam spruce forest) produced similar increases in canopy openness and number of canopy openings, increasing light availability to understory vegetation [36,39]. VAI, as calculated from lidar returns, showed sustained declines in treated plots (Figure 5). However, litter trap data from the same plots did not reveal similar trends [26] suggesting that this metric may not be a reliable indicator of true leaf area index. Patterns observed in other lidar metrics suggest that while the decline in surface area demonstrated by lidar-derived meanVAI is indeed real, it is probably linked to increased canopy porosity, foliage clumping, and sky fraction, suggesting that meanVAI is a better indicator of total canopy surface area than LAI per se. Patterns from lidar and litter baskets indicate that while remaining tree species increased production of foliage, this new leaf area did not fill the canopy spaces left by defoliated aspen and birch within the first four years following treatment. Increased light availability corresponding to a more open upper canopy will boost canopy carbon assimilation, but canopy openness will likely decline as remaining trees produce additional leaf area.
Several aspects of canopy height indicated a general downward shift of canopy surface area concomitant with aspen and birch senescence, supporting our hypothesis that canopy heights would decline with aspen and birch mortality. Both mean leaf height, indicative of average height of leaf area, and mean canopy height, indicative of height of the upper canopy surface, declined over time in treated stands (Figure 5A,D). ModeEl and Mode2 (Figure 4B,C), both indicators of relative skewness of vertical canopy foliage distribution, declined suggesting that in addition to development of a shorter canopy overall, distribution of leaf area in treated stands became more “bottom-heavy”. Canopies in nearby, much older stands are much taller than those observed in either treated or control stands of this study, suggesting that canopy height will recover to and eventually exceed pretreatment heights. Foliage distributions in those older stands, however, tends to be similarly “bottom-heavy” [33], suggesting this trend may continue as treated stands age.
We hypothesized that increases in canopy structural complexity with forest age might be driven by the moderate disturbances posed by diffuse mortality of canopy-dominant, early-successional cohorts. In contrast, the patterns reported here, while not all statistically significant, suggest net simplification of some aspects of canopy structure in response to aspen and birch mortality. Heterogeneity of canopy surface area, both throughout the canopy depth, and at its surface (Rugosity and TopRug, Figure 6A,B, respectively) was lower after girdling than pretreatment, albeit not significantly. While rugosity has proven a reliable indicator of changes in canopy structural complexity over decades to centuries of forest succession [32,33], it was a poor predictor of short-term canopy responses to widespread, diffuse mortality occurring over very short periods. Mean variability of leaf height (meanSTD, Figure 6C) and variability of mean leaf height (height2, Figure 6D) in each plot trended downward following girdling, both eventually diverging significantly from control plots, indicating that vertical distribution of canopy surface area became less variable in treated stands as aspen and birch died. The reduction in canopy structural heterogeneity may be due to the downward shift of canopy surface area accompanying overstory mortality which temporarily compressed leaf area into a narrower band of canopy height. Additional observation is necessary to determine if the reduction of canopy structural complexity reported here would continue, but it is noteworthy that rugosity of much older stands (in which aspen and birch senesced long ago [16]) is significantly higher than any stand reported here, including those in which the aspen/birch overstory component either recently died, or soon will.
4.2. Functional Consequences
Previous studies report qualitative patterns of canopy reorganization at successional timescales, while results presented here quantify changes over a much shorter period. Results of previous studies at the University of Michigan Biological Station employing a chronosequence approach indicate that canopies become more structurally complex (rugosity increases), that distribution of leaf area becomes more heterogeneous as forests age, and that increasing canopy structural complexity drives increases in light use efficiency [32,33]. In response to a pulse of aspen and birch mortality in the overstory, lidar-derived metrics of canopy structure indicate that the canopies of treated plots become more open, shorter, and more structurally uniform. All of these changes are likely only temporary, however. Other studies have described canopy structural change in terms of expansion in young stands and erosion/elaboration in mature and old stands [8,40]; we suggest that the changes to canopy structure documented here in response to elimination of an even-aged, early successional cohort may mark a transition point between expansion and erosion phases. Further study will determine if the reductions in canopy structural complexity seen here continue or are reversed.
Despite mortality of even-aged canopy dominant aspen and birch species in this mixed deciduous forest, and concomitant reductions in canopy structural complexity, declines in stand C assimilation were smaller than expected due to resilient coupling of C and N cycles which quickly redistributed resources from dead and dying aspen/birch to rapidly growing successor species [19]. Further, LAI in treated stands declined significantly relative to pretreatment values three years post-girdling (a trend not seen in control stands), but recovered quickly and were not significantly different from control stands or pretreatment values by the fourth year post-girdling [26]. This decline and subsequent recovery of LAI was accompanied by structural rearrangement of the canopy such that the loss of LAI had minimal effect on the magnitude of light intercepted by canopies in treated stands and no discernable effect on net ecosystem exchange in any year [26]. This finding underscores previous observations of the partial functional independence of quantity of leaf area and its arrangement in the canopy [32,33]. Changes in canopy structure which rearrange leaf area within the canopy have implications for canopy light interception and thus carbon storage rates [8,9,40,41,42,43]. Such patterns were observed in a hemlock-dominated forest in southern Appalachia, where growth of successor species increased due to increased light availability in the first two years following mortality of Eastern hemlock [37] and similar results in numerous stand thinning experiments document increased stand production rates [12,13,18,32,33,36,37,39,44,45,46,47]. These studies document ecosystem functional resilience to partial stand disturbance despite mortality rates of canopy dominants approaching 40%, and attribute such resilience to redistribution of light in the canopy and rearrangement of foliage such that light interception and canopy carbon uptake rates do not decline in proportion to disturbance.
5. Conclusions
Results presented here indicate numerous rearrangements of canopy structure in the first four years following an experiment that accelerated forest succession, resulting in widespread aspen and birch mortality in the upper forest canopy. This disturbance caused rapid reorganization of canopy structure with lasting effects on ecosystem function. Gap fraction of the upper canopy increased and canopies in treated stands become shorter with foliage distributed nearer the bottom than pretreatment or control stands. Patchy distribution of aspen and birch across the landscape increased variability of foliage clumping observed in treated stands. Widespread senescence of canopy-dominant aspen and birch prompted significant shifts in canopy structural complexity, affecting light distribution and interception, with consequences for canopy-scale C assimilation. Despite increases in gap fraction of the upper canopy, there was sufficient leaf area remaining to maintain canopy functional competence. These changes indicate that reorganization of canopy structure can contribute to resilience of ecosystem function and that widespread mortality does not necessarily result in declines in forest productivity. As forests throughout the upper Great Lakes region proceed through this same successional transition, extensive tracts of forest canopy will undergo reorganizations resembling those observed in this accelerated succession experiment.
Acknowledgments
We acknowledge Geoffrey Parker at Smithsonian Environmental Research Center for generous assistance with the Portable Canopy lidar system and tips on data processing. Jennifer Nietz (née Goedhart) and Chelsea Rapp assisted with fieldwork. This research was supported by the U.S. Department of Energy’s Office of Science (BER) through the Midwestern Regional Center of the National Institute for Global Environmental Change under Cooperative Agreements No. DE-FC03-90ER610100, and DE-SC0006708, the Midwestern Regional Center of the National Institute for Climatic Change Research at Michigan technological University, under Award No. DE-FC02-06ER64158 and National Science Foundation grant #DEB-0911461. We acknowledge the University of Michigan Biological Station and its staff for facilities and material support. BSH was funded in part by an NSF IGERT Fellowship #DGE-0504552 awarded through the University of Michigan Biosphere-Atmosphere Research Training (BART) program. GB and PSC were funded in part by National Oceanic and Atmospheric Administration grant # NA11OAR4310190. Any opinions, findings, and conclusions expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
Conflict of Interest
The authors declare no conflict of interest.
References
- Valverde, T.; Silvertown, J. Canopy closure rate and forest structure. Ecology 1997, 78, 1555–1562. [Google Scholar] [CrossRef]
- Parker, G.G.; Harding, D.J.; Berger, M.L. A portable LIDAR system for rapid determination of forest canopy structure. J. Appl. Ecol. 2004, 41, 755–767. [Google Scholar] [CrossRef]
- Brown, M.J.; Parker, G.G. Canopy light transmittance in a chronosequence of mixed-species deciduous forests. Can. J. For. Res. 1994, 24, 1694–1703. [Google Scholar] [CrossRef]
- Vanderwel, M.C.; Coomes, D.A.; Purves, D.W. Quantifying variation in forest disturbance, and its effects on aboveground biomass dynamics, across the eastern United States. Glob. Change Biol. 2013, 19, 1504–1517. [Google Scholar] [CrossRef]
- Canham, C.D. Growth and canopy architecture of shade-tolerant trees—Response to canopy gaps. Ecology 1988, 69, 786–795. [Google Scholar] [CrossRef]
- Duursma, R.A.; Mäkelä, A. Summary models for light interception and light-use efficiency of non-homogeneous canopies. Tree Physiol. 2007, 27, 859–870. [Google Scholar] [CrossRef]
- North, M.; Chen, J.Q.; Oakley, B.; Song, B.; Rudnicki, M.; Gray, A.; Innes, J. Forest stand structure and pattern of old-growth western hemlock/douglas-fir and mixed-conifer forests. For. Sci. 2004, 50, 299–311. [Google Scholar]
- Ishii, H.T.; Tanabe, S.; Hiura, T. Exploring the relationships among canopy structure, stand productivity, and biodiversity of temperature forest ecosystems. For. Sci. 2004, 50, 342–355. [Google Scholar]
- Ishii, H.; Asano, S. The role of crown architecture, leaf phenology and photosynthetic activity in promoting complementary use of light among coexisting species in temperate forests. Ecol. Res. 2010, 25, 715–722. [Google Scholar] [CrossRef]
- Bradshaw, G.A.; Spies, T.A. Characterizing canopy gap structure in forests using wavelet analysis. J. Ecol. 1992, 80, 205–215. [Google Scholar] [CrossRef]
- Parker, W.C.; Dey, D.C. Influence of overstory density on ecophysiology of red oak (Quercus rubra) and sugar maple (Acer saccharum) seedlings in central Ontario shelterwoods. Tree Physiol. 2008, 28, 797–804. [Google Scholar] [CrossRef]
- Chiang, J.-M.; Brown, K.J. The effects of thinning and burning treatments on within-canopy variation of leaf traits in hardwood forests of southern Ohio. For. Ecol. Manag. 2010, 260, 1065–1075. [Google Scholar] [CrossRef]
- Goudiaby, V.; Brais, S.; Grenier, Y.; Berninger, F. Thinning effects on jack pine and black spruce photosynthesis in eastern boreal forests of Canada. Silva Fenn. 2011, 45, 595–609. [Google Scholar]
- Moreno-Gutierrez, C.; Barbera, G.G.; Nicolas, E.; de Luis, M.; Castillo, V.M.; Martinez-Fernandez, F.; Querejeta, J.I. Leaf δ18O of remaining trees is affected by thinning intensity in a semiarid pine forest. Plant Cell Environ. 2011, 34, 1009–1019. [Google Scholar] [CrossRef]
- Forest Inventory and Analysis (FIA) Forest Inventory Data Online (FIDO). Available online: http://apps.fs.fed.us/fido/ (accessed on 3 July 2013).
- Albert, D.A.; Minc, L.D. The Natural Ecology and Cultural History of the Colonial Point Red Oak Stands; University of Michigan Biological Station: Pellston, MI, USA, 1987. [Google Scholar]
- Karamanski, T.J. Deep Wood Frontier—A History of Logging in Northern Michigan; Wayne State University Press: Detroit, MI, USA, 1989. [Google Scholar]
- Gough, C.M.; Vogel, C.S.; Hardiman, B.; Curtis, P.S. Wood net primary production resilience in an unmanaged forest transitioning from early to middle succession. For. Ecol. Manag. 2010, 260, 36–41. [Google Scholar] [CrossRef]
- Nave, L.E.; Gough, C.M.; Maurer, K.D.; Bohrer, G.; Hardiman, B.S.; Le Moine, J.; Munoz, A.B.; Nadelhoffer, K.J.; Sparks, J.P.; Strahm, B.D.; et al. Disturbance and the resilience of coupled carbon and nitrogen cycling in a north temperate forest. J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef]
- Frelich, L.E.; Reich, P.B. Spatial patterns and succession in a Minnesota southern-boreal forest. Ecol. Monogr. 1995, 65, 325–346. [Google Scholar] [CrossRef]
- Edenius, L.; Ericsson, G.; Kempe, G.; Bergström, R.; Danell, K. The effects of changing land use and browsing on aspen abundance and regeneration: A 50-year perspective from Sweden. J. Appl. Ecol. 2011, 48, 301–309. [Google Scholar] [CrossRef]
- Korb, J.E.; Fule, P.Z.; Stoddard, M.T. Forest restoration in a surface fire-dependent ecosystem: An example from a mixed conifer forest, southwestern Colorado, USA. For. Ecol. Manag. 2012, 269, 10–18. [Google Scholar] [CrossRef]
- Caspersen, J.P.; Pacala, S.W.; Jenkins, J.C.; Hurtt, G.C.; Moorcroft, P.R.; Birdsey, R.A. Contributions of land-use history to carbon accumulation in U.S. forests. Science 2000, 290, 1148–1151. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Change Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Coates, K.D.; Messier, C. A Critique of Silviculture: Managing for Complexity; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Gough, C.M.; Hardiman, B.S.; Nave, L.E.; Bohrer, G.; Maurer, K.D.; Vogel, C.S.; Nadelhoffer, K.J.; Curtis, P.S. Sustained carbon uptake and storage following moderate disturbance in a great lakes forest. Ecol. Appl. 2013, 23. in press. [Google Scholar]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Su, H.B.; Curtis, P.S. Multi-year convergence of biometric and meteorological estimates of forest carbon storage. Agric. For. Meteorol. 2008, 148, 158–170. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, J.M.; Birdsey, R.; McCullough, K.; He, L.; Deng, F. Age structure and disturbance legacy of North American forests. Biogeosciences 2011, 8, 715–732. [Google Scholar] [CrossRef]
- Tardy, S.W. Soil Survey of Cheboygan County, Michigan; General Books: New York, NY, USA, 1991. [Google Scholar]
- Nave, L.E.; Vogel, C.S.; Gough, C.M.; Curtis, P.S. The contribution of atmospheric nitrogen deposition to net primary productivity in a northern hardwood forest. Can. J. For. Res. 2009, 39, 1108–1118. [Google Scholar] [CrossRef]
- Schmid, H.P.; Su, H.B.; Vogel, C.S.; Curtis, P.S. Ecosystem-atmosphere exchange of carbon dioxide over a mixed hardwood forest in northern lower Michigan. J. Geophys. Res. 2003, 108. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Bohrer, G.; Gough, C.M.; Vogel, C.S.; Curtis, P.S. The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 2011, 92, 1818–1827. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Gough, C.M.; Halperin, A.; Hofmeister, K.L.; Nave, L.E.; Bohrer, G.; Curtis, P.S. Maintaining high rates of carbon storage in old forests: A mechanism linking canopy structure to forest function. For. Ecol. Manag. 2013, 298, 111–119. [Google Scholar] [CrossRef]
- Kucharik, C.J.; Norman, J.M.; Gower, S.T. Measurements of branch area and adjusting leaf area index indirect measurements. Agric. For. Meteorol. 1998, 91, 69–88. [Google Scholar] [CrossRef]
- Gonsamo, A.; Walter, J.-M.N.; Pellikka, P. Sampling gap fraction and size for estimating leaf area and clumping indices from hemispherical photographs. Can. J. For. Res. 2010, 40, 1588–1603. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Harvey, B.D.; Reyes, G.P.; Caron, M.N.; Barlow, S. Spruce budworm, windthrow and partial cutting: Do different partial disturbances produce different forest structures? For. Ecol. Manag. 2011, 262, 482–490. [Google Scholar] [CrossRef]
- Ford, C.R.; Elliott, K.J.; Clinton, B.D.; Kloeppel, B.D.; Vose, J.M. Forest dynamics following eastern hemlock mortality in the southern Appalachians. Oikos 2012, 121, 523–536. [Google Scholar] [CrossRef]
- Parker, G.G. Structure and Microclimate of Forest Canopies. In Forest Canopies: A Review of Research on a Biological Frontier; Lowman, M., Nadkarni, N., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 73–106. [Google Scholar]
- Kneeshaw, D.D.; Bergeron, Y. Canopy gap characteristics and tree replacement in the southeastern boreal forest. Ecology 1998, 79, 783–794. [Google Scholar] [CrossRef]
- Parker, G.G.; Russ, M.E. The canopy surface and stand development: Assessing forest canopy structure and complexity with near-surface altimetry. For. Ecol. Manag. 2004, 189, 307–315. [Google Scholar] [CrossRef]
- Walcroft, A.S.; Brown, K.J.; Schuster, W.S.F.; Tissue, D.T.; Turnbull, M.H.; Griffin, K.L.; Whitehead, D. Radiative transfer and carbon assimilation in relation to canopy architecture, foliage area distribution and clumping in a mature temperate rainforest canopy in New Zealand. Agric. For. Meteorol. 2005, 135, 326–339. [Google Scholar] [CrossRef]
- Niinemets, Ü. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef]
- Mori, A.; Niinemets, Ü. Plant responses to heterogeneous environments: Scaling from shoot modules and whole-plant functions to ecosystem processes. Ecol. Res. 2010, 25, 691–692. [Google Scholar]
- Lemoine, D.; Jacquemin, S.; Granier, A. Beech (Fagus sylvatica L.) branches show acclimation of xylem anatomy and hydraulic properties to increased light after thinning. Ann. For. Sci. 2002, 59, 761–766. [Google Scholar]
- Quentin, A.G.; Beadle, C.L.; O’Grady, A.P.; Pinkard, E.A. Effects of partial defoliation on closed canopy Eucalyptus globulus Labilladiere: Growth, biomass allocation and carbohydrates. For. Ecol. Manag. 2011, 261, 695–702. [Google Scholar] [CrossRef]
- Ogunjemiyo, S.; Parker, G.; Roberts, D. Reflections in bumpy terrain: Implications of canopy surface variations for the radiation balance of vegetation. IEEE Geosci. Remote Sens. Lett. 2005, 2, 90–93. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Boerner, A.; Knohl, A.; Hessenmoeller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).