Afforestation of Boreal Open Woodlands: Early Performance and Ecophysiology of Planted Black Spruce Seedlings

Abstract
:1. Introduction
2. Methods
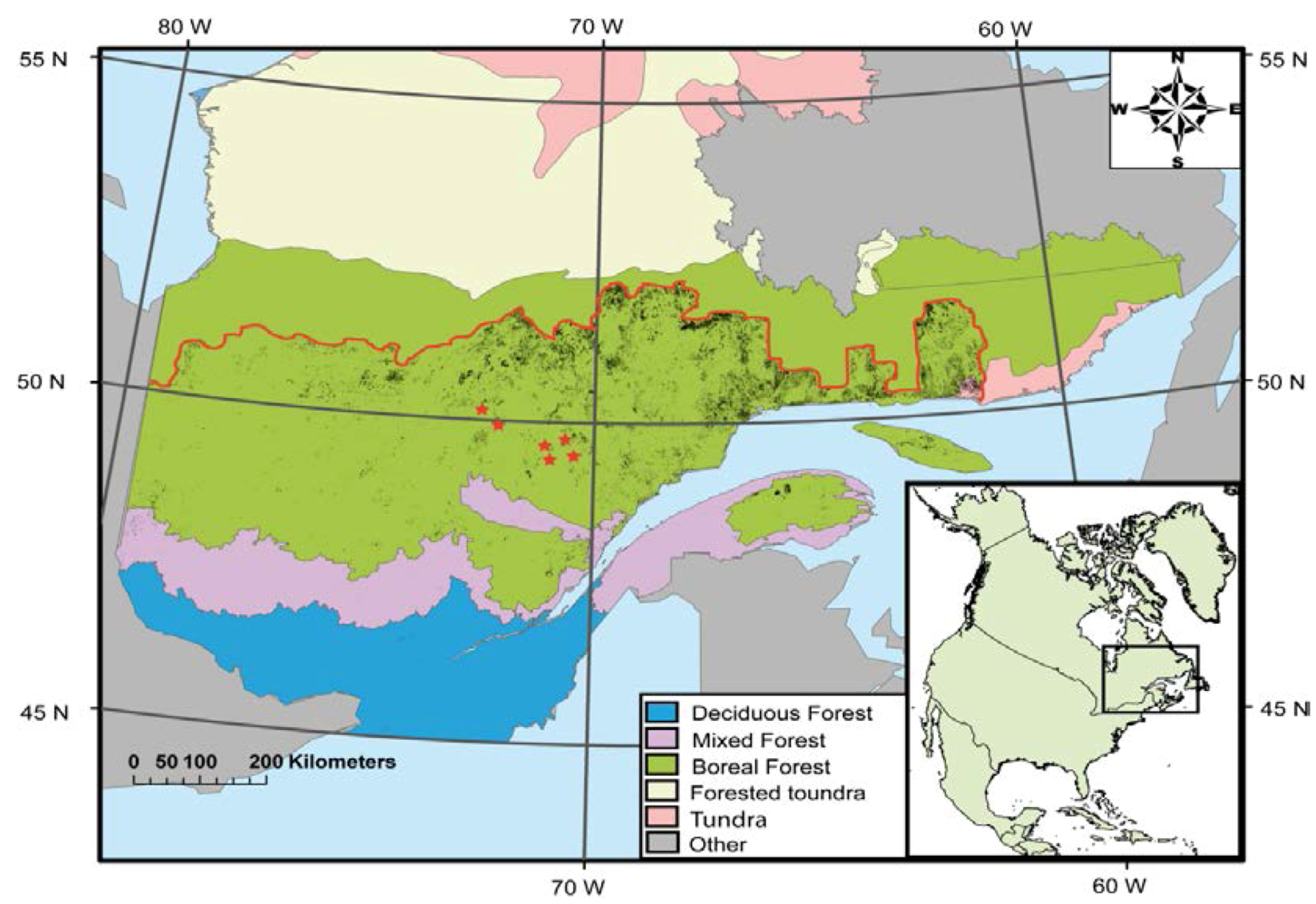
2.1. Site Description
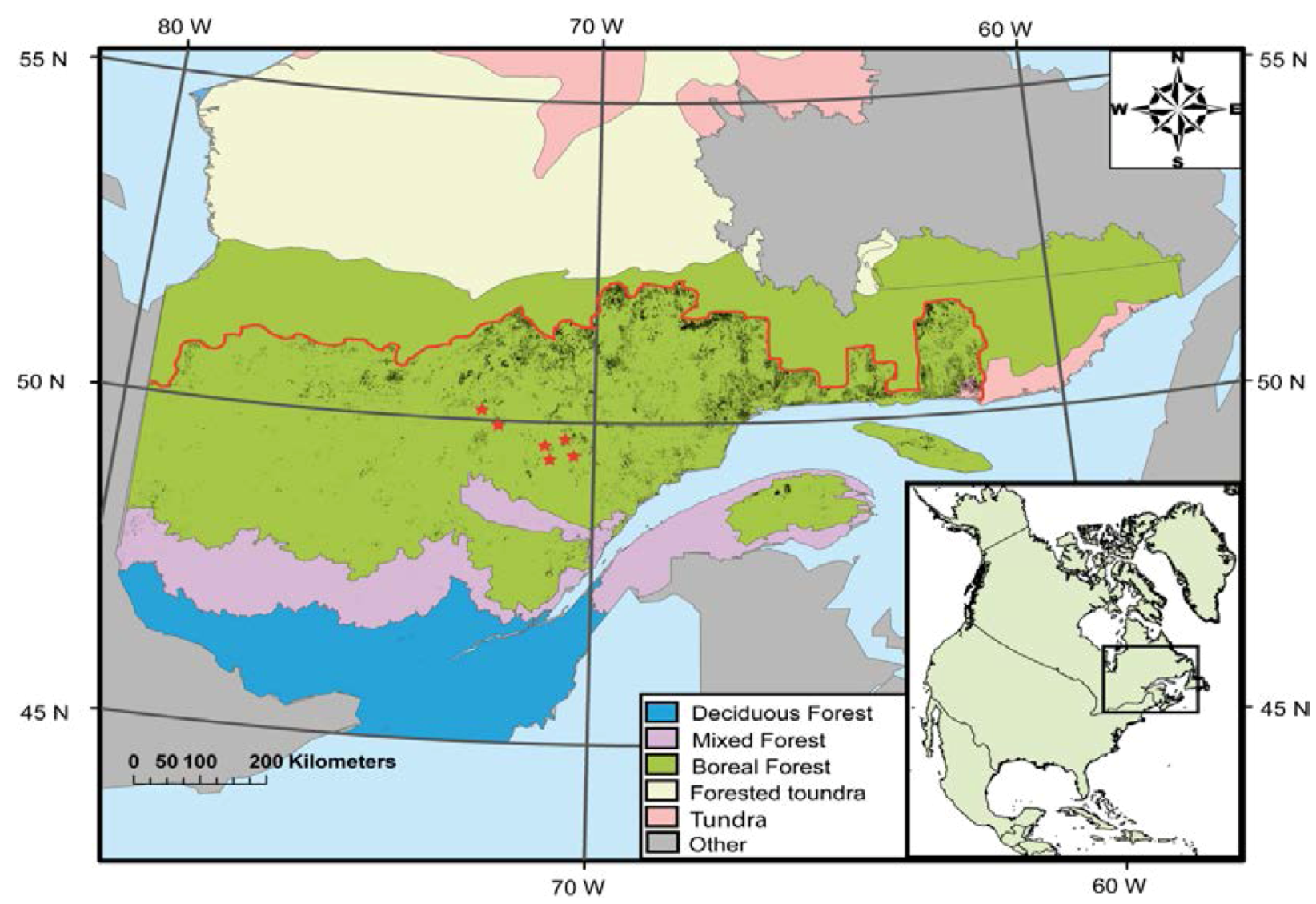
2.2. Experimental Design and Biological Material
2.3. Physiological Measurements
2.4. Survival and Morphological Measurements
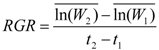 where W2 is total biomass of seedling at t2 (fall 2006) and W1 is total seedling biomass at t1 (fall 2005). This equation takes into account initial seedling size and yields an unbiased estimate of RGR under all conditions [30]. The same calculations were applied on Ht and Ds, but these results are not presented as they were similar to those with biomass.
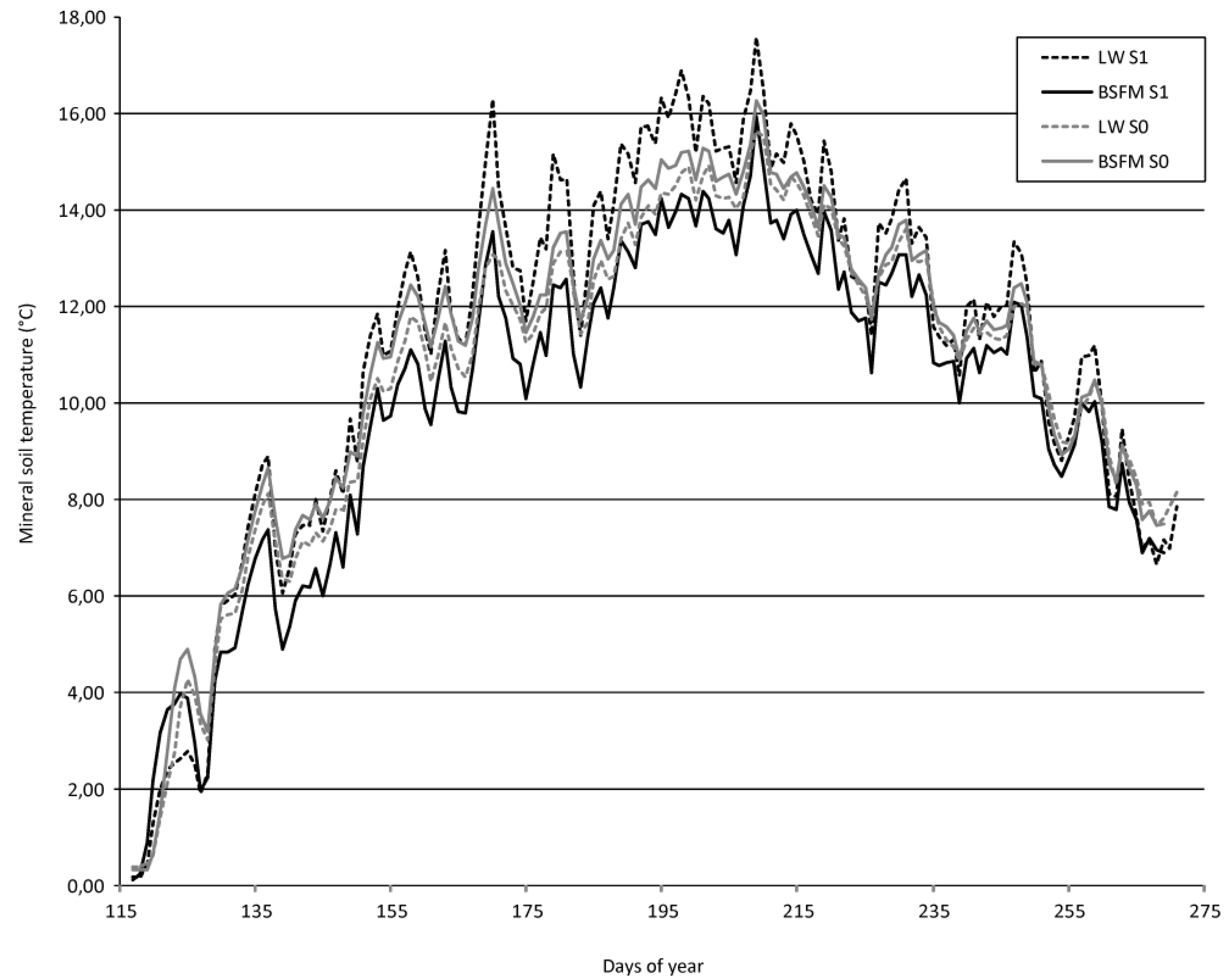
where W2 is total biomass of seedling at t2 (fall 2006) and W1 is total seedling biomass at t1 (fall 2005). This equation takes into account initial seedling size and yields an unbiased estimate of RGR under all conditions [30]. The same calculations were applied on Ht and Ds, but these results are not presented as they were similar to those with biomass.2.5. Abiotic Variable Measurements
2.6. Statistical Analysis
3. Results
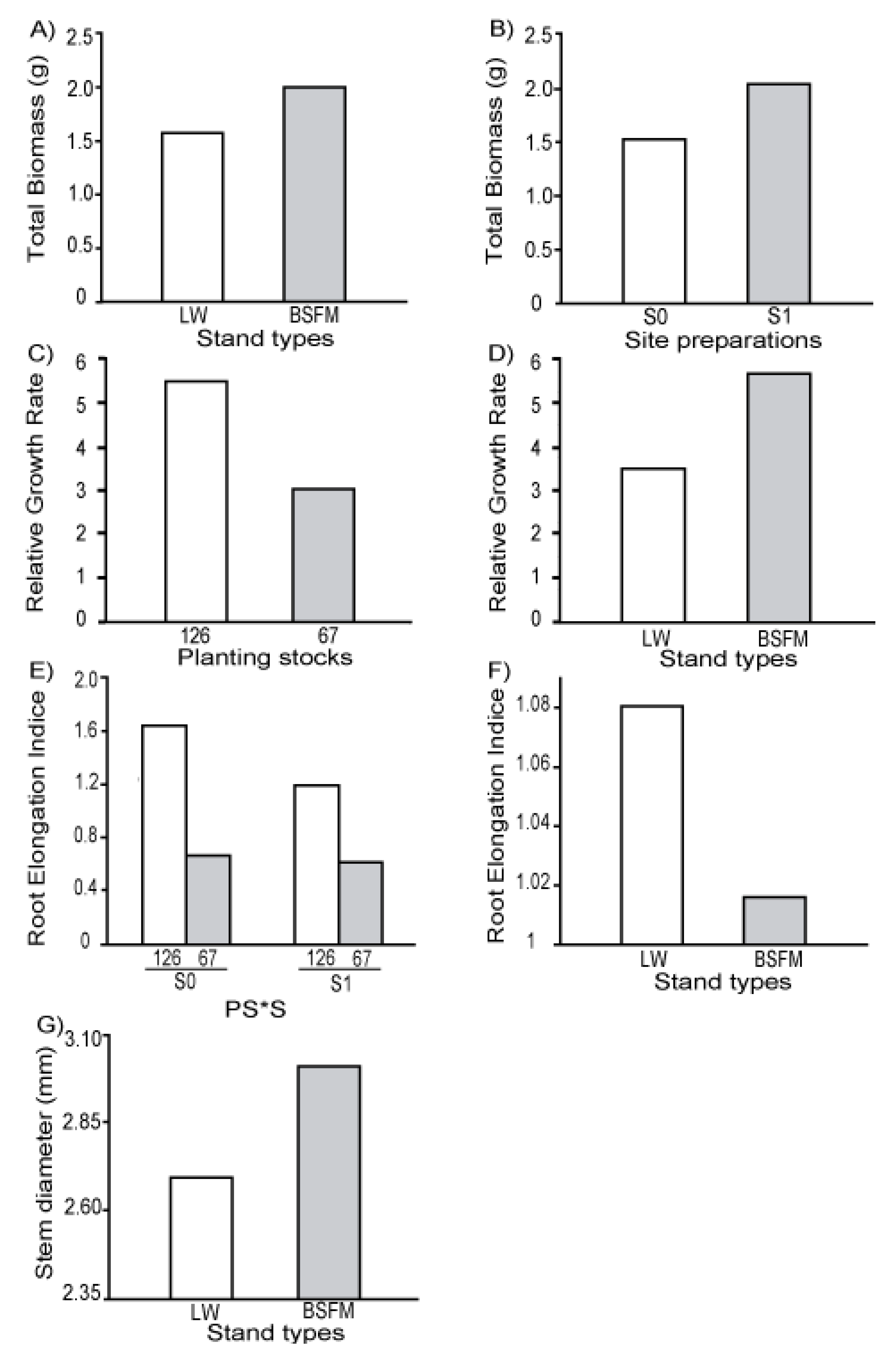
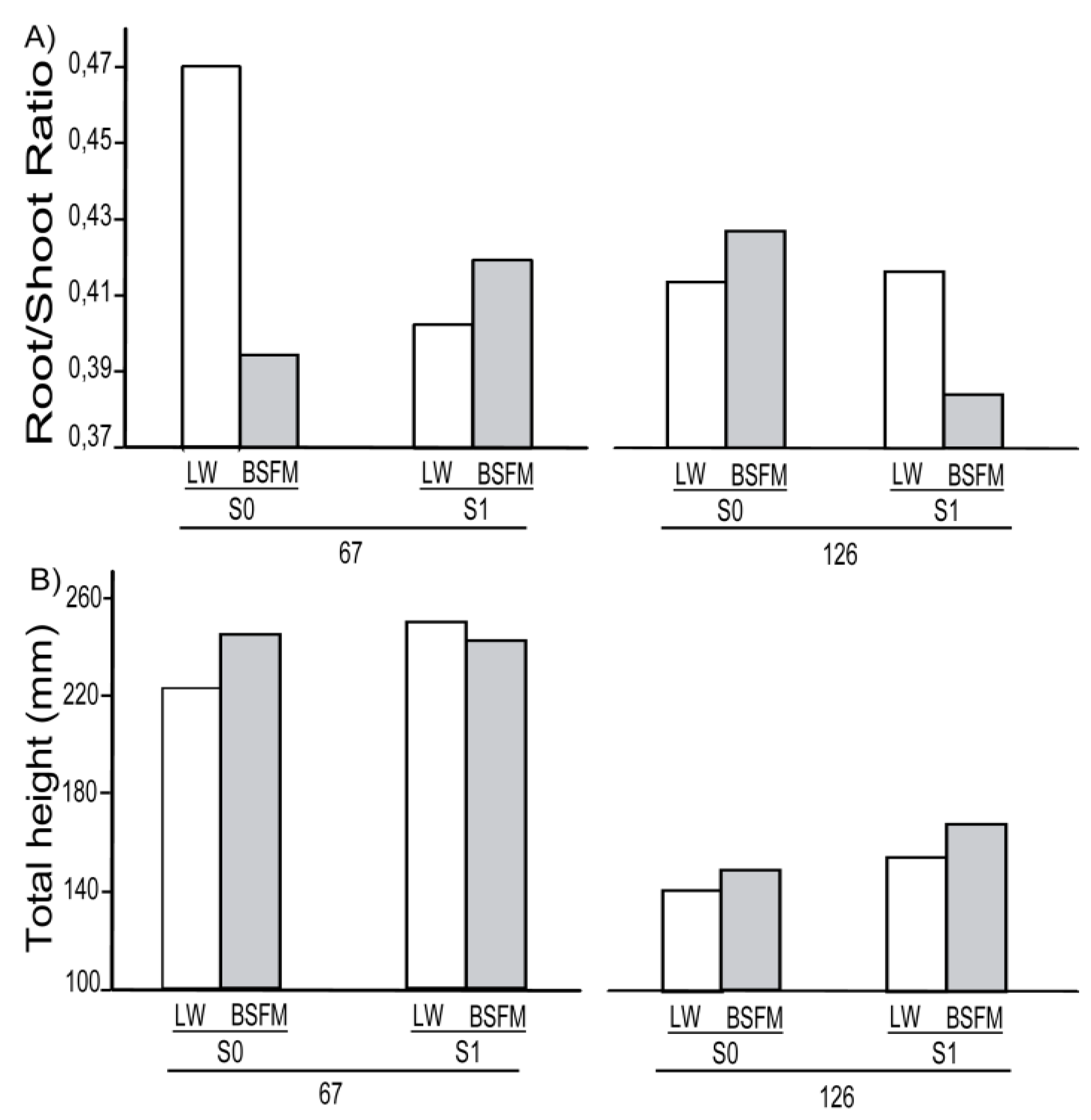
3.1. Seedling Survival and Growth

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources of variation | BT | HT | DS | RGR | Survival | R/SRatio | REI * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ndf | ddf | p | ddf | p | ddf | p | ddf | p | ddf | p | ddf | p | ddf | p | |
| Block | 5 | 5 | 0.2577 | 5.1 | 0.2252 | 5.09 | 0.1816 | 5 | 0.1337 | 3.996 | 0.8460 | 4.999 | 0.7512 | 5.08 | 0.0451 |
| Stand Types (ST) | 1 | 5 | 0.0116 | 5.108 | 0.0683 | 5.096 | 0.0097 | 5 | 0.0018 | 4.056 | 0.0811 | 5.045 | 0.2240 | 5.16 | 0.0447 |
| Site preparation (S) | 1 | 10 | 0.0363 | 10.05 | 0.0538 | 10.05 | 0.1516 | 10 | 0.0543 | 10.19 | 0.0012 | 9.688 | 0.6031 | 10.05 | 0.0029 |
| ST * S | 1 | 10 | 0.8284 | 10.05 | 0.4126 | 10.05 | 0.6826 | 10 | 0.6524 | 10.19 | 0.2238 | 9.696 | 0.5308 | 10.05 | 0.2298 |
| Planting Stock Sizes (PS) | 1 | 20 | <0.0001 | 19.99 | <0.0001 | 20.15 | <0.0001 | 20 | 0.0011 | 19.6 | 0.0006 | 20.9 | 0.0246 | 20.39 | <0.0001 |
| ST * PS | 1 | 20 | 0.5809 | 19.99 | 0.7759 | 20.15 | 0.6494 | 20 | 0.4026 | 19.6 | 0.8607 | 20.92 | 0.4756 | 20.39 | 0.5422 |
| S * PS | 1 | 20 | 0.4358 | 19.99 | 0.6217 | 20.15 | 0.9345 | 20 | 0.2368 | 19.6 | 0.9917 | 20.9 | 0.2778 | 20.39 | 0.0221 |
| ST * S * PS | 1 | 20 | 0.8871 | 19.99 | 0.0462 | 20.15 | 0.7170 | 20 | 0.5570 | 19.6 | 0.7443 | 20.92 | 0.0190 | 20.39 | 0.6858 |
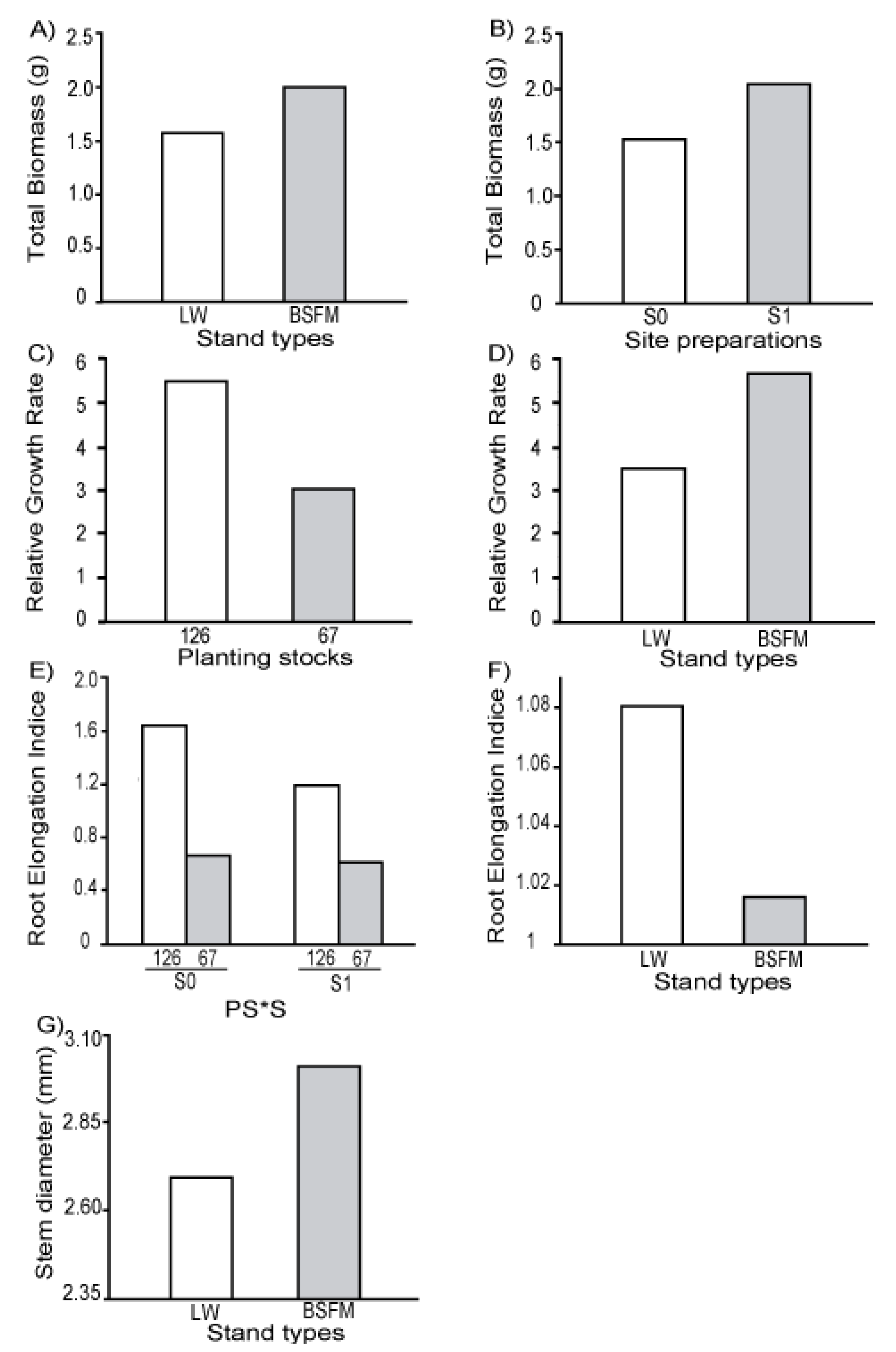
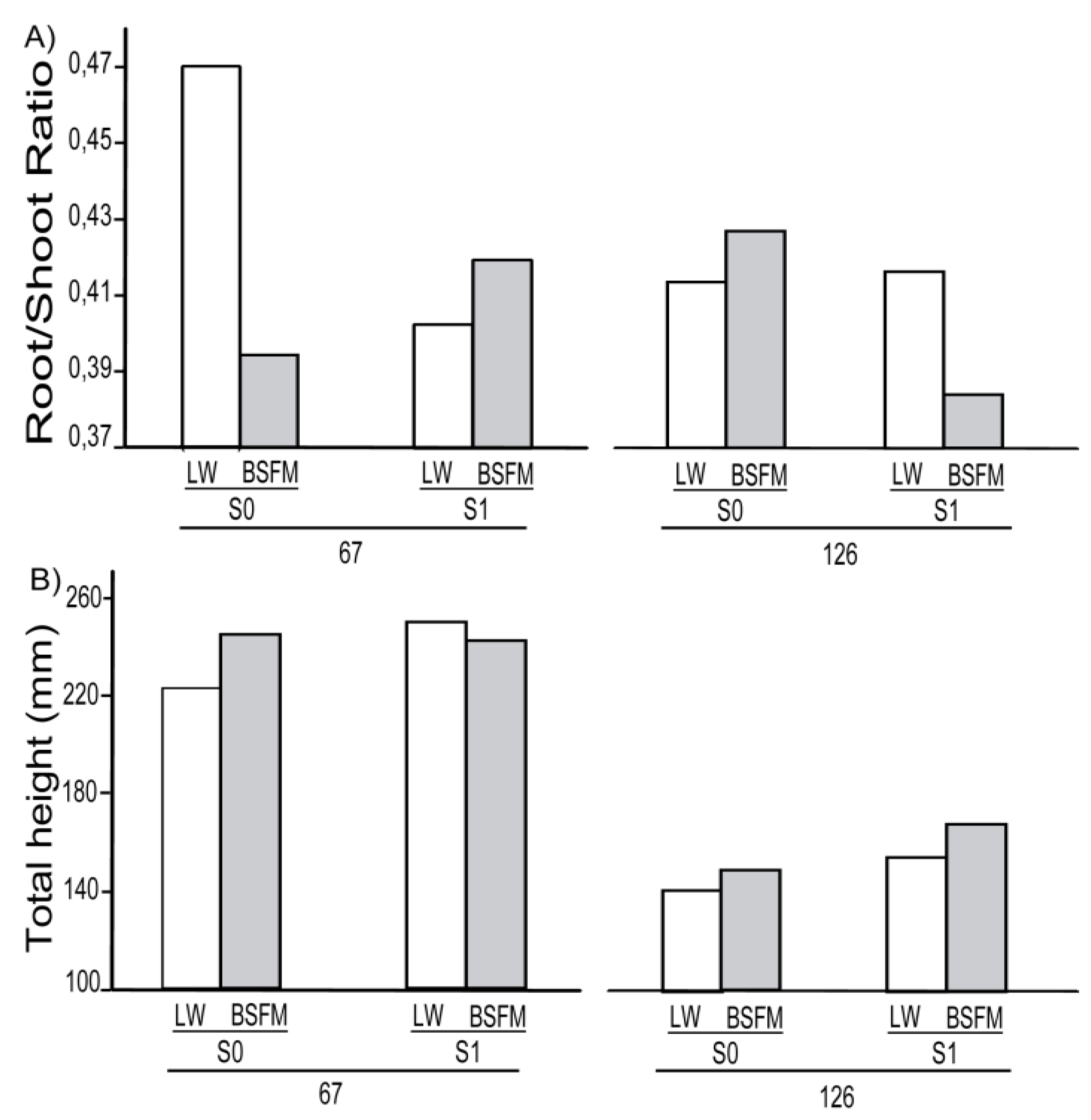
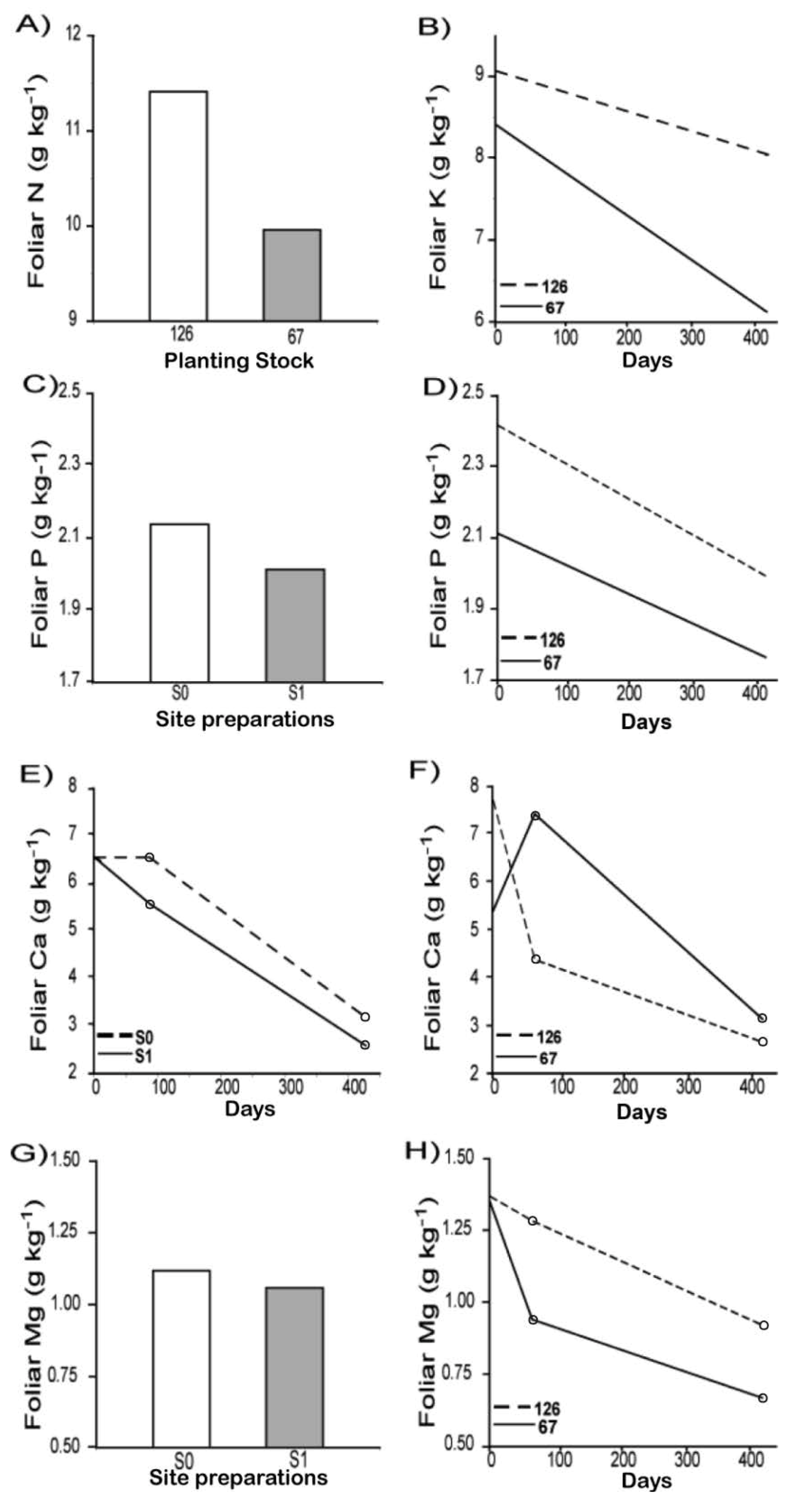
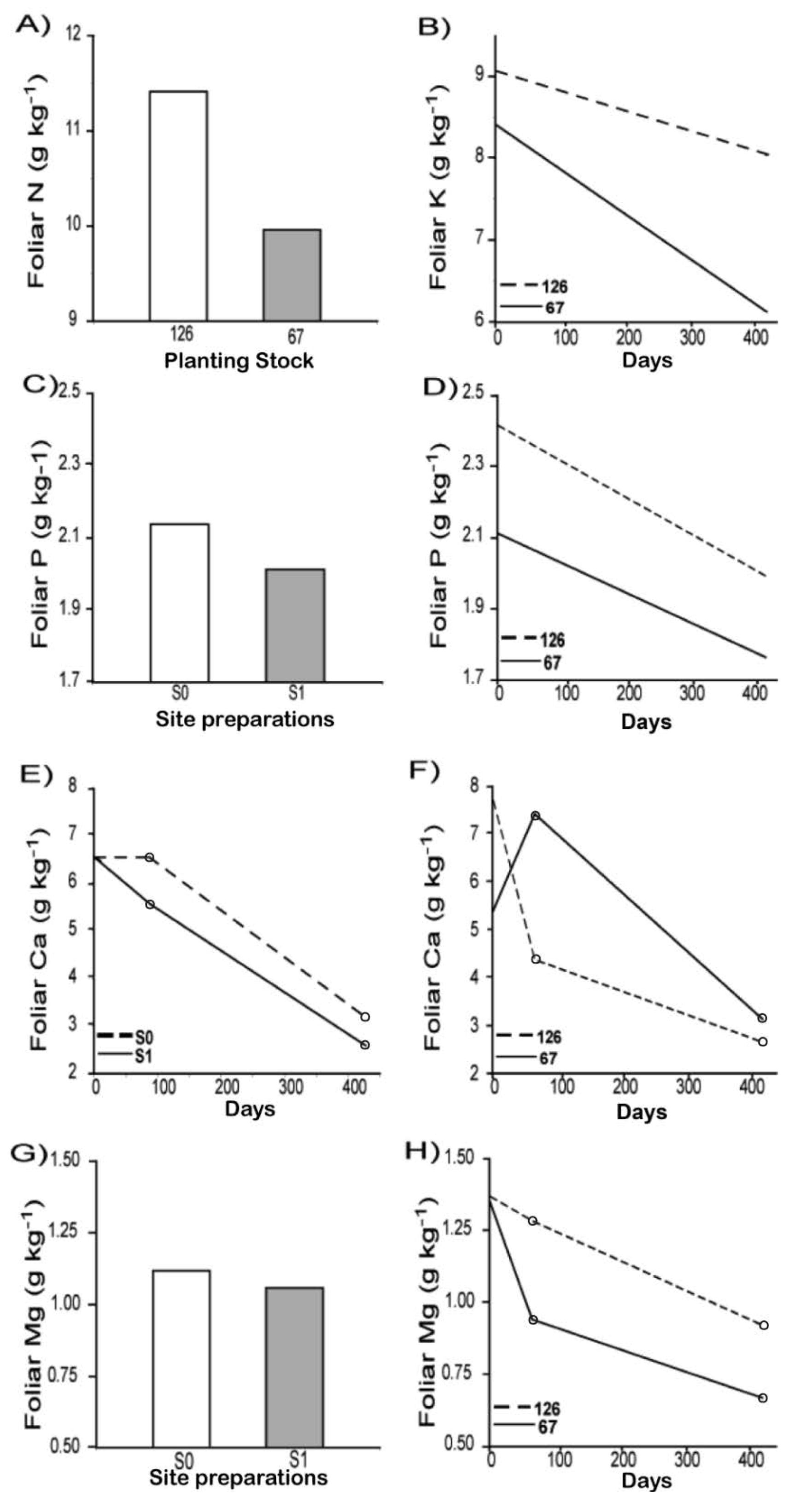
3.2. Physiological Response
| Sources of variation | Foliar N | Foliar P | Foliar K | Foliar Ca | Foliar Mg | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ndf | ddf | p | ddf | p | ddf | p | ddf | p | ddf | p | |
| Block (B) | 5 | 8.867 | 0.8042 | 9.056 | 0.3573 | 9.03 | 0.6334 | 8.54 | 0.4567 | 9.12 | 0.4805 |
| Date (D) | 2 | 8.886 | 0.4977 | 9.059 | 0.3156 | 9.03 | 0.1790 | 8.593 | <0.0001 | 9.13 | <0.0001 |
| Stand types (ST) | 1 | 13.87 | 0.1201 | 14.16 | 0.6597 | 13.77 | 0.8819 | 14.03 | 0.0652 | 13.56 | 0.5662 |
| D * ST | 2 | 13.87 | 0.2141 | 14.16 | 0.9292 | 13.77 | 0.5256 | 14.02 | 0.2887 | 13.56 | 0.8379 |
| Site preparation (S) | 1 | 27.84 | 0.2584 | 27.45 | 0.0095 | 27.28 | 0.9884 | 26.84 | <0.0001 | 27.35 | 0.0415 |
| D * S | 2 | 27.85 | 0.7031 | 27.45 | 0.0684 | 27.28 | 0.2891 | 26.84 | 0.0034 | 27.35 | 0.1759 |
| Contrasts | |||||||||||
| D (Linear) * S | - | - | - | - | - | - | 26.80 | 0.0002 | - | - | |
| D (Quad) * S | - | - | - | - | - | - | 26.88 | 0.0001 | - | - | |
| S0 vs. S1 (D 427) | - | - | - | - | - | - | 26.78 | 0.0024 | - | - | |
| ST * S | 1 | 27.84 | 0.5920 | 27.45 | 0.6095 | 27.28 | 0.6999 | 26.84 | 0.3278 | 27.35 | 0.9673 |
| D * ST * S | 2 | 27.85 | 0.9234 | 27.45 | 0.6169 | 27.28 | 0.6759 | 26.84 | 0.7372 | 27.35 | 0.6749 |
| Planting Stock size (PS) | 1 | 54.90 | 0.0002 | 55.03 | 0.0034 | 54.99 | <0.0001 | 54.33 | 0.3094 | 55.08 | <0.0001 |
| D * PS | 2 | 54.90 | 0.2282 | 55.03 | 0.0338 | 55.00 | <0.0001 | 54.33 | <0.0001 | 55.09 | 0.0088 |
| Contrasts | |||||||||||
| D (Linear) * PS | 1 | - | - | 54.73 | 0.0011 | 54.69 | <0.0001 | 54.21 | 0.8724 | 54.81 | 0.00014 |
| D (Quad) * PS | 1 | - | - | 55.35 | 0.0373 | 55.33 | 0.1309 | 54.45 | 0.1916 | 55.38 | 0.00013 |
| 126 vs. 67 (D 427) | 1 | - | - | 54.63 | 0.0242 | 54.58 | <0.0001 | 54.17 | 0.4629 | 54.17 | 0.4629 |
| ST * PS | 1 | 54.90 | 0.9805 | 55.03 | 0.6403 | 54.99 | 0.7788 | 54.33 | 0.9998 | 55.08 | 0.9890 |
| S * PS | 1 | 54.90 | 0.3652 | 55.03 | 0.8205 | 54.99 | 0.5825 | 54.33 | 0.6941 | 55.08 | 0.6729 |
| D * ST * PS | 2 | 54.90 | 0.8622 | 55.03 | 0.7129 | 55.00 | 0.7515 | 54.33 | 0.9413 | 55.09 | 0.6763 |
| D * S * PS | 2 | 54.90 | 0.6127 | 55.03 | 0.8093 | 55.00 | 0.6828 | 54.33 | 0.9556 | 55.09 | 0.9260 |
| ST * S * PS | 1 | 54.90 | 0.1929 | 55.03 | 0.6372 | 54.99 | 0.9855 | 54.33 | 0.8922 | 55.08 | 0.7861 |
| D * ST * S * PS | 2 | 54.90 | 0.5862 | 55.03 | 0.9430 | 55.00 | 0.7234 | 54.33 | 0.9672 | 55.09 | 0.9696 |
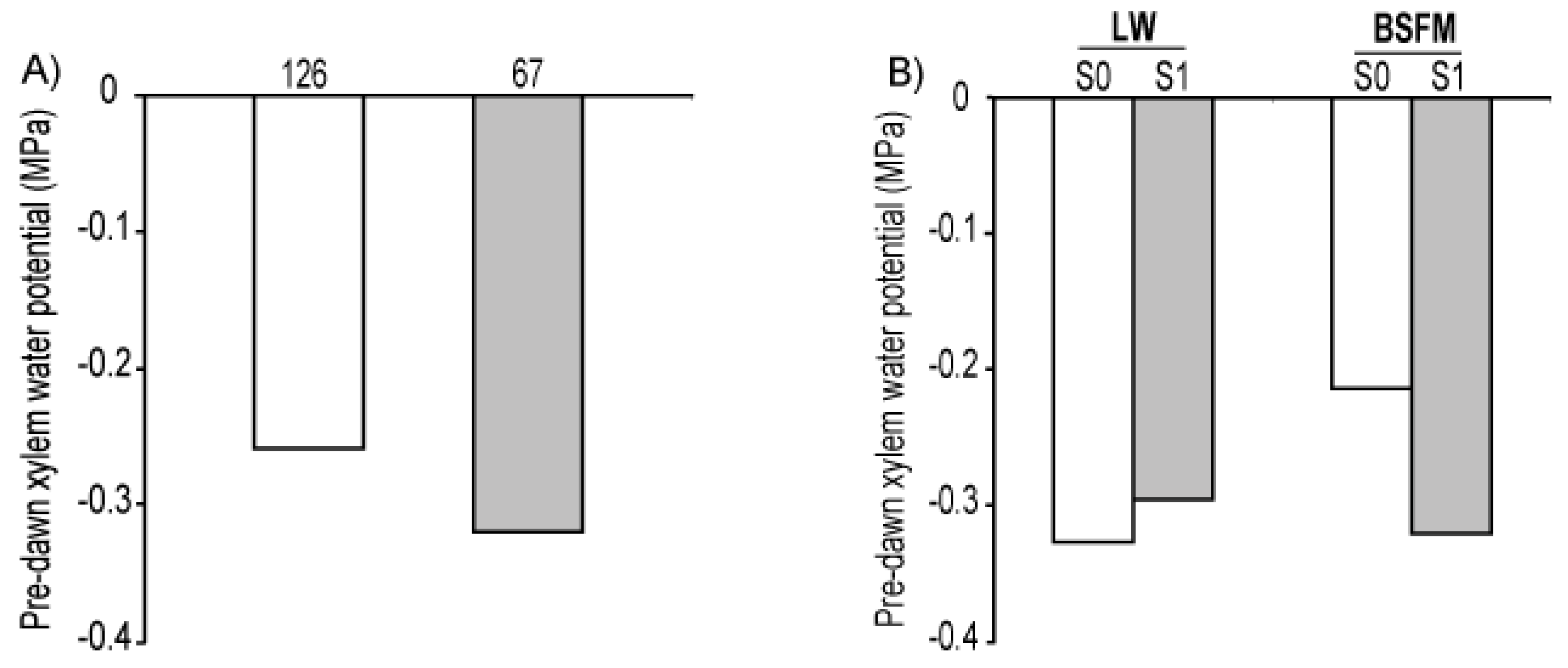
| Sources of variation | A | gs * | WUE | ||||
|---|---|---|---|---|---|---|---|
| ndf | ddf | p | ddf | p | ddf | p | |
| Block | 3 | 3.079 | 0.2727 | 2.991 | 0.7195 | 2.893 | 0.1593 |
| Date (D) | 1 | 3.08 | 0.0516 | 2.991 | 0.4087 | 2.893 | 0.0455 |
| Stand type (ST) | 1 | 6.172 | 0.6319 | 5.342 | 0.4799 | 3.986 | 0.8498 |
| D * ST | 1 | 6.172 | 0.1506 | 5.342 | 0.5902 | 3.986 | 0.2679 |
| Site preparation (S) | 1 | 11.73 | 0.3687 | 8.797 | 0.8045 | 9.998 | 0.8657 |
| D * S | 1 | 11.73 | 0.9808 | 8.797 | 0.7468 | 9.998 | 0.6900 |
| ST * S | 1 | 11.73 | 0.7314 | 8.797 | 0.5180 | 9.998 | 0.9575 |
| D * ST * S | 1 | 11.73 | 0.4305 | 8.797 | 0.2651 | 9.998 | 0.5686 |
| Planting stock sizes (PS) | 1 | 24.4 | 0.0669 | 18.58 | 0.1875 | 23.48 | 0.1136 |
| D * PS | 1 | 24.4 | 0.1872 | 18.58 | 0.2814 | 23.48 | 0.1211 |
| ST * PS | 1 | 24.37 | 0.4089 | 18.58 | 0.9452 | 23.46 | 0.4749 |
| S * PS | 1 | 24.39 | 0.7785 | 18.59 | 0.0788 | 23.48 | 0.6637 |
| ST * S * PS | 1 | 24.37 | 0.4348 | 18.58 | 0.2722 | 23.46 | 0.3353 |
| D * ST * PS | 1 | 24.39 | 0.5721 | 18.59 | 0.7157 | 23.48 | 0.5757 |
| D * S * PS | 1 | 24.39 | 0.4501 | 18.59 | 0.5247 | 23.48 | 0.0792 |
| D * ST * S * PS | 1 | 24.39 | 0.4122 | 18.59 | 0.6049 | 23.48 | 0.1543 |
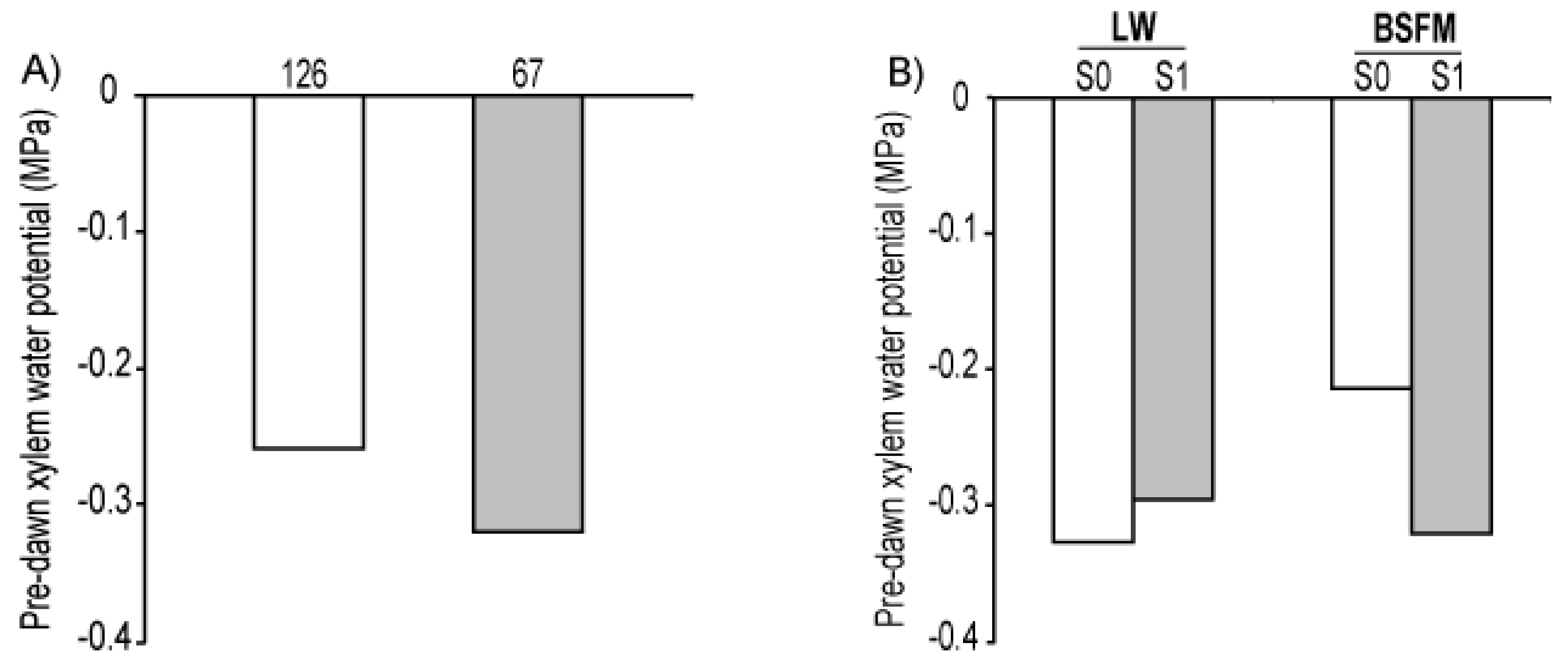
| Sources of variation | Ψxpd | ||
|---|---|---|---|
| ndf | ddf | P | |
| Block | 3 | 3.03 | 0.1302 |
| Stand type (ST) | 1 | 3.05 | 0.3873 |
| Site preparation (S) | 1 | 6.21 | 0.1511 |
| ST * S | 1 | 6.21 | 0.0232 |
| Planting stock sizes (PS) | 1 | 12.30 | 0.0474 |
| ST * PS | 1 | 12.30 | 0.1860 |
| S * PS | 1 | 12.30 | 0.7778 |
| ST * S * PS | 1 | 12.30 | 0.9613 |
| Stand type | N | K | Ca | Mg | Mn | Al | Fe | Na | S |
|---|---|---|---|---|---|---|---|---|---|
| BSFM | 1.09 | 13.01 | 79.65 | 9.08 | 1.95 | 206.24 | 72.61 | 6.92 | 45.32 |
| LW | 1.31 | 14.64 | 43.48 | 5.67 | 1.43 | 200.4 | 82.38 | 7.63 | 49.3 |
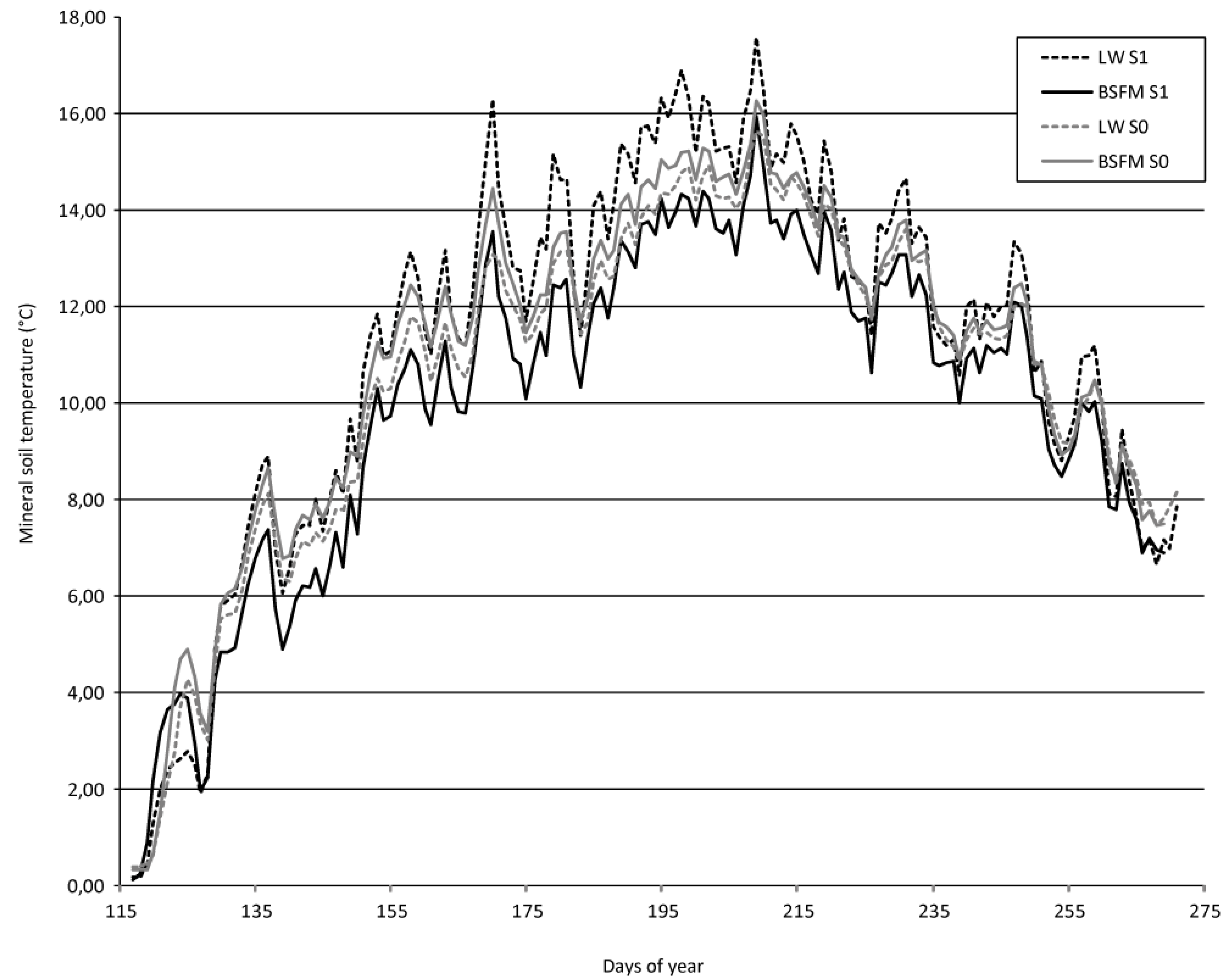
4. Discussion
4.1. Seedling Response to Stand Type
4.2. Planting Stock Size
4.3. Seedling Response to Site Preparation
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Coulombe, G.; Huot, J.; Arsenault, J.; Bauce, É.; Bernard, J.T.; Bouchard, A.; Liboiron, M.A.; Szaraz, G. Commission D’étude sur la Gestion de la Forêt Publique Québécoise; Bibliothèque Nationale du Québec: Québec, Canada, 2004; p. 307. [Google Scholar]
- Arseneault, D.; Payette, S. A postfire shift from lichen-spruce to lichen-tundra vegetation at tree line. Ecology 1992, 73, 1067–1081. [Google Scholar] [CrossRef]
- Payette, S.; Bhiry, N.; Delwaide, A.; Simard, M. Origin of the lichen woodland at its southern range limit in eastern Canada: The catastrophic impact of insect defoliators and fire on the spruce-moss forest. Can. J. For. Res. 2000, 30, 288–305. [Google Scholar] [CrossRef]
- Gagnon, R.; Morin, H. Les forêts d’épinette noire du Québec: Dynamique, perturbations et biodiversité. Nat. Can. 2001, 125, 26–35. [Google Scholar]
- Girard, F.; Payette, S.; Gagnon, R. Rapid expansion of lichen woodlands within the closed-crown boreal forest zone over the last 50 years caused by stand disturbances in eastern Canada. Can. J. Biogeogr. 2008, 35, 529–537. [Google Scholar] [CrossRef]
- Jasinski, J.P.P.; Payette, S. The creation of alternative stable states in the southern boreal forest, Québec, Canada. Ecol. Monogr. 2005, 75, 561–583. [Google Scholar] [CrossRef]
- Hébert, F.; Boucher, J.F.; Bernier, P.Y.; Lord, D. Growth response and water relations of 3-year-old planted black spruce and jack pine seedlings in site prepared lichen woodlands. For. Ecol. Manag. 2006, 223, 226–236. [Google Scholar] [CrossRef]
- Côté, D.; Girard, F.; Hébert, F.; Bouchard, S.; Gagnon, R.; Lord, D. Is the closed-crown boreal forest resilient after successive stand disturbances? A quantitative demonstration from a case study. J. Veg. Sci. 2013, 24, 664–674. [Google Scholar] [CrossRef]
- Gaboury, S.; Boucher, J.F.; Villeneuve, C.; Lord, D.; Gagnon, R. Estimating the net carbon balance of boreal open woodland afforestation: A case study in Québec’s closed-crown boreal forest. For. Ecol. Manag. 2009, 257, 483–494. [Google Scholar] [CrossRef]
- Mallik, A.U. Ecology of a forest weed of Newfoundland: Vegetative regeneration strategy of Kalmia angustifolia. Can. J. Bot. 1993, 71, 161–166. [Google Scholar] [CrossRef]
- Mallik, A.U. Growth and physiological responses of black spruce (Picea mariana) to sites dominated by Ledum groenlandicum. J. Chem. Ecol. 1996, 22, 575–585. [Google Scholar] [CrossRef]
- Yamasaki, S.H.; Fyles, J.W.; Egger, K.N.; Titus, B.D. The effect of Kalmia angustifolia on the growth, nutrition, and ectomycorrhizal symbiont community of black spruce. For. Ecol. Manag. 1998, 105, 197–207. [Google Scholar] [CrossRef]
- Thiffault, N.; Cyr, G.; Prégent, G.; Jobidon, R.; Charrette, L. Régénération artificielle des pessières noires à éricacées: Effets du scarifiage, de la fertilisation et du type de plants après 10 ans. For. Chron. 2004, 80, 141–149. [Google Scholar]
- Thiffault, N.; Jobidon, R. How to shift unproductive Kalmia angustifolia–Rhododendron groenlandicum heath to productive conifer plantation. Can. J. For. Res. 2006, 36, 2364–2376. [Google Scholar] [CrossRef]
- LeBel, P.; Thiffault, N.; Bradley, R. Kalmia removal increases nutrient supply and growth of black spruce seedlings: An effect fertilizer cannot emulate. For. Ecol. Manag. 2008, 256, 1780–1784. [Google Scholar] [CrossRef]
- Moroni, M.T.; Thiffault, N.; Titus, B.D.; Mante, C.; Makeschin, F. Controlling Kalmia and reestablishing conifer dominance enhances soil fertility indicators in central Newfoundland, Canada. Can. J. For. Res. 2009, 39, 1270–1279. [Google Scholar] [CrossRef]
- Steijlen, I.; Nilsson, M.C.; Zackrisson, O. Seed regeneration of scots pine in boreal forest stands dominated by lichen and feather moss. Can. J. For. Res. 1995, 25, 713–723. [Google Scholar] [CrossRef]
- Fisher, R.F. Possible allelopathic effects of reindeer-moss (Cladonia) on jack pine and white spruce. For. Sci. 1979, 25, 256–260. [Google Scholar]
- Houle, G.; Filion, L. The effects of lichens on white spruce seedling establishment and juvenile growth in a spruce-lichen woodland of subarctic Quebec. Ecoscience 2003, 10, 80–84. [Google Scholar]
- Burdett, A.N. Physiological processes in plantation establishment and the development of specifications for forest planting stock. Can. J. For. Res. 1990, 20, 415–427. [Google Scholar] [CrossRef]
- Bernier, P.Y. Comparing natural and planted black spruce seedlings 1. Water relations and growth. Can. J. For. Res. 1993, 23, 2427–2434. [Google Scholar] [CrossRef]
- Girard, F. Remise en Production des Pessières à Lichens de la Forêt Boréale Commerciale: Nutrition et Croissance de Plants D’épinette Noire Trois Ans Après Traitements de Préparation de Terrain; Université du Québec à Chicoutimi: Chicoutimi, Canada, 2004. [Google Scholar]
- Jobidon, R. Light threshold for optimal black spruce (Picea mariana) seedling growth and development under brush competition. Can. J. For. Res. 1994, 24, 1629–1635. [Google Scholar] [CrossRef]
- Lamhamedi, M.; Bernier, P.Y.; Hébert, C. Effect of shoot size on the gas exchange and growth of containerized Picea mariana seedlings under different watering regimes. New For. 1997, 13, 209–223. [Google Scholar] [CrossRef]
- Nabuurs, G.J.; Masera, O.; Andrasko, K.; Benitez-Ponce, P.; Boer, R.; Dutschke, M.; Elsiddig, E.; Ford-Robertson, J.; Frumhoff, P.; Karjalainen, T.; et al. Forestry. In Climate Change 2007: Mitigation. Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; pp. 541–584. [Google Scholar]
- Peters-Stanley, M.; Hamilton, K.; Marcello, T.; Sjardin, M. Back to the Future: State of the Voluntary Carbon Markets 2011; Ecosystem Marketplace: Washington, DC, USA, 2011; p. 93. [Google Scholar]
- Saucier, J.P.; Bergeron, J.F.; Grondin, P.; Robitaille, A. Les Régions Écologiques du Québec Méridional; Ministère des Ressources Naturelles du Québec: Ste-Foy, Canada, 2000. [Google Scholar]
- Environment Canada National Climate Data and Information Archive. Available online: http://climate.weatheroffice.gc.ca/climateData/canada_e.html (accessed on 1 April 2013).
- Scholander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap pressure in vascular plants, negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar]
- Hoffman, W.A.; Poorter, H. Avoiding bias in calculation of Relative Growth Rate. Ann. Bot. Lond. 2002, 80, 37–42. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill Publishing Company: New York, NY, USA, 1980; p. 633. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999; p. 266. [Google Scholar]
- Devore, J.; Peck, R. Introductory Statistics, 2nd ed.; West Publishing Company: St. Paul, MN, USA, 1994; p. 640. [Google Scholar]
- Sedia, E.G.; Ehrenfeld, J.G. Lichens and mosses promote alternate stable plant communities in the New Jersey Pinelands. Oikos 2003, 100, 447–458. [Google Scholar] [CrossRef]
- Trofymow, J.A.; van den Driessche, R. Mycorizha. In Mineral Nutrition of Conifer Seedlings; CRC Press: Boston, MA, USA, 1991; pp. 183–229. [Google Scholar]
- Johansson, K.; Nilson, U.; Lee, A.H. Interaction between soil scarification and Norway spruce seedling types. New For. 2007, 33, 13–27. [Google Scholar] [CrossRef]
- Rudinger, M.; Hallgren, S.W.; Steudle, E.; Schulze, E.D. Hydraulic and osmotic properties of spruce roots. J. Exp. Bot. 1994, 45, 1413–1425. [Google Scholar] [CrossRef]
- Kramer, P.J. Water Deficits and Plant Growth. In Water Relations of Plants; Academic Press: New York, NY, USA, 1983; pp. 342–389. [Google Scholar]
- Grossnickle, S.C. Planting stress in newly planted jack pine and white spruce 1. Factors influencing water uptake. Tree Physiol. 1988, 4, 71–83. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Davies, W.J. Control of water balance in transplanted trees. J. Arboric. 1975, 1, 1–10. [Google Scholar]
- Kramer, P.J.; Kozlowski, T.T. Physiology of Woody Plants; Academic Press Inc.: New York, NY, USA, 1979; pp. 58–112. [Google Scholar]
- Folk, R.S.; Grossnickle, S.C. Stock-type patterns of phosphorus uptake, retranslocation, net photosynthesis and morphological development in interior spruce seedlings. New For. 2000, 19, 27–49. [Google Scholar] [CrossRef]
- Colombo, S.J. Second year shoot development in black spruce Picea mariana (Mill) BSP container seedlings. Can. J. For. Res. 1986, 16, 68–73. [Google Scholar] [CrossRef]
- Bigras, F.J.; D’Aoust, A.L. Hardening and dehardening of shoot and roots of containerized black spruce and white spruce seedlings under short and long days. Can. J. For. Res. 1992, 22, 388–396. [Google Scholar] [CrossRef]
- Malik, V.; Timmer, V.R. Growth, nutrient dynamics, and interspecific competition of nutrient-loaded black spruce seedlings on a boreal mixedwood site. Can. J. For. Res. 1996, 26, 1651–1659. [Google Scholar] [CrossRef]
- Timmer, V.R.; Morrow, L.D. Predicting Fertilizer Growth Response and Nutrient Status of Jack Pine by Foliar Diagnosis. In Proceedings of the Forest Soils and Treatment Impacts, Sixth North American Forest Soils Conference, University of Tennessee, Knoxville, TN, USA, 1983; pp. 335–351.
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Funct. Plant Biol. 1988, 15, 63–92. [Google Scholar]
- Canham, C.D. Different responses to gaps among shade-tolerant tree species. Ecology 1989, 70, 548–550. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Wayne, P.M. Coping with Environmental Heterogeneity: The Physiological Ecology of Tree Seedling Regeneration across the Gap-Understory Continuum. In Exploitation of Environmental Heterogeneity by Plants: Ecophysiological Processes Above- and Belowground; Caldwell, M.M., Pearcy, R.W., Eds.; Academic Press Inc.: San Diego, CA, USA, 1994; pp. 349–390. [Google Scholar]
- Walters, M.B.; Reich, P.B. Trade-offs in low-light CO2 exchange: A component of variation in shade tolerance among cold temperate tree seedlings. Funct. Ecol. 2000, 14, 155–165. [Google Scholar] [CrossRef]
- Tan, W.X.; Blake, T.J.; Boyle, T.J.B. Drought tolerance in faster-growing and slower-growing black spruce (Picea mariana) Progenies 1. Stomatal and gas exchange responses to osmotic stress. Physiol. Plant. 1992, 85, 639–644. [Google Scholar] [CrossRef]
- Bassman, J.H. Influence of two site preparation treatments on ecophysiology of planted Picea engelmannii × glauca seedlings. Can. J. For. Res. 1989, 19, 1359–1370. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; Heikurinen, J. Site preparation: Water relations and growth of newly planted jack pine and white spruce. New For. 1989, 3, 99–123. [Google Scholar] [CrossRef]
- Örlander, G.; Gemmel, P.; Hunt, J. Site Preparation: A Swedish Overview; British Columbia Ministry of Forests, Canada/BC Economic & Regional Development Agreement: Victoria, BC, Canada, 1990; p. 61. [Google Scholar]
- Prévost, M. Effet du scarifiage sur les propriétés du sol et de l’ensemencement naturel dans une pessière noire à mousses de la forêt boréale québécoise. Can. J. For. Res. 1996, 26, 72–86. [Google Scholar] [CrossRef]
- Boucher, J.F.; Wetzel, S.; Munson, A.D. Leaf level response of planted eastern white pine (Pinus strobus L.) seven years after intensive silvicultural treatments. For. Ecol. Manag. 1998, 107, 291–307. [Google Scholar] [CrossRef]
- Bedford, L.; Sutton, R.F. Site preparation for establishing lodgepole pine in the sub-boreal spruce zone of interior British Columbia: The Bednesti trial, 10-year results. For. Ecol. Manag. 2000, 126, 227–238. [Google Scholar] [CrossRef]
- Thiffault, N.; Jobidon, R.; Munson, A.D. Performance and physiology of large containerized and bare-root spruce seedlings in relation to scarification and competition in Quebec (Canada). Ann. For. Sci. 2003, 60, 645–655. [Google Scholar] [CrossRef]
- Thiffault, N.; Hébert, F.; Jobidon, R. Planted Picea mariana growth and nutrition as influenced by silviculture × nursery interaction on an ericaceous-dominated site. Silva Fenn. 2012, 46, 667–682. [Google Scholar]
- Bowen, G.D. Soil Temperature, Root Growth, and Plant Function. In Plant Roots: The Hidden Half; Waisel, Y., Eshel, A., KapFkafi, U., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1991; pp. 309–330. [Google Scholar]
- Dodd, I.C.; He, J.; Turnbull, C.G.N.; Lee, S.K.; Critchley, C. The influence of supra-optimal root-zone temperature on growth and stomatal conductance in Capsicum annuum L. J. Exp. Bot. 2000, 51, 239–248. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical & Environmental Plant Physiology; Academic Press: San Diego, CA, USA, 1999; p. 474. [Google Scholar]
- Brais, S. Persistence of soil compaction and effects on seedling growth in Northwestern Quebec. Soil Sci. Soc. Am. J. 2001, 65, 1263–1271. [Google Scholar] [CrossRef]
- Boivin, R.; Miller, B.D.; Timmer, V.R. Late-season fertilisation of Picea mariana seedlings under greenhouse culture: Biomass and nutrient dynamics. Ann. For. Sci. 2002, 59, 255–264. [Google Scholar] [CrossRef]
- Mansuy, N.; Gauthier, S.; Bergeron, Y. Afforestation opportunities when stand productivity is driven by a high risk of natural disturbance: A review of the open lichen woodland in the eastern boreal forest of Canada. Mitig. Adapt. Strateg. Glob. Change 2012, 18, 245–264. [Google Scholar] [CrossRef]
- Boucher, J.-F.; Tremblay, P.; Gaboury, S.; Villeneuve, C. Can boreal afforestation help offset incompressible GHG emissions from Canadian industries? Process Saf. Environ. 2012, 90, 459–466. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tremblay, P.; Boucher, J.-F.; Tremblay, M.; Lord, D. Afforestation of Boreal Open Woodlands: Early Performance and Ecophysiology of Planted Black Spruce Seedlings. Forests 2013, 4, 433-454. https://doi.org/10.3390/f4020433
Tremblay P, Boucher J-F, Tremblay M, Lord D. Afforestation of Boreal Open Woodlands: Early Performance and Ecophysiology of Planted Black Spruce Seedlings. Forests. 2013; 4(2):433-454. https://doi.org/10.3390/f4020433
Chicago/Turabian StyleTremblay, Pascal, Jean-Francois Boucher, Marc Tremblay, and Daniel Lord. 2013. "Afforestation of Boreal Open Woodlands: Early Performance and Ecophysiology of Planted Black Spruce Seedlings" Forests 4, no. 2: 433-454. https://doi.org/10.3390/f4020433
APA StyleTremblay, P., Boucher, J.-F., Tremblay, M., & Lord, D. (2013). Afforestation of Boreal Open Woodlands: Early Performance and Ecophysiology of Planted Black Spruce Seedlings. Forests, 4(2), 433-454. https://doi.org/10.3390/f4020433