Using Silviculture to Influence Carbon Sequestration in Southern Appalachian Spruce-Fir Forests

Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
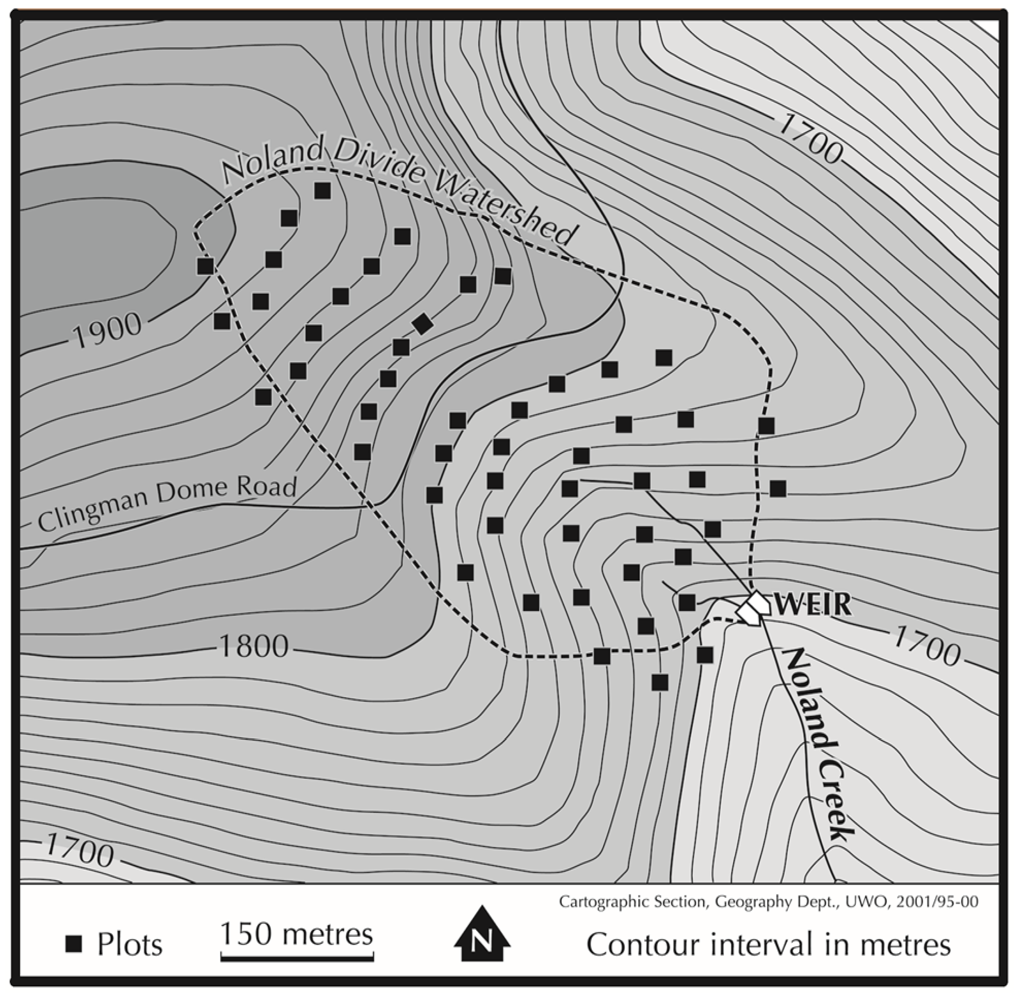
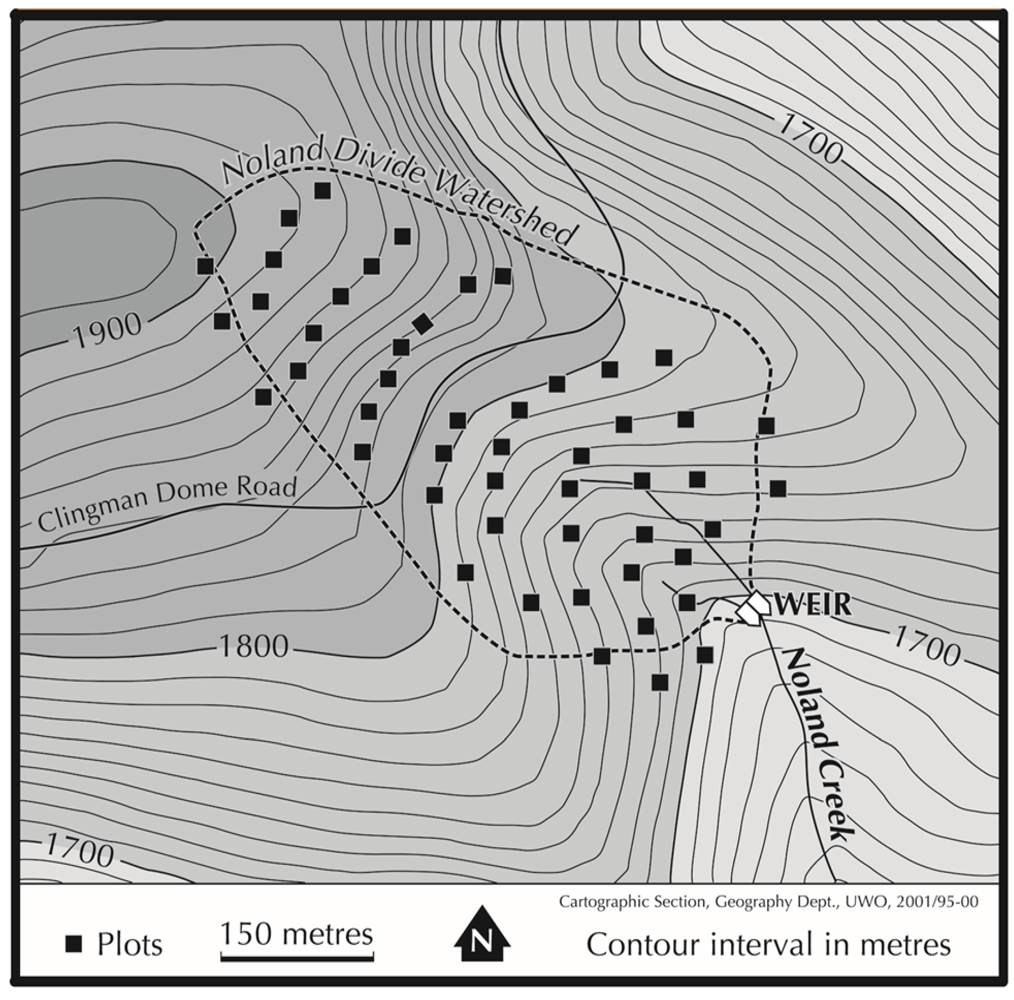
2.1. Study Area
2.2. Data Collection
2.3. Data Preparation
2.4. Silvicultural Scenarios
2.5. Carbon Accounting
3. Results
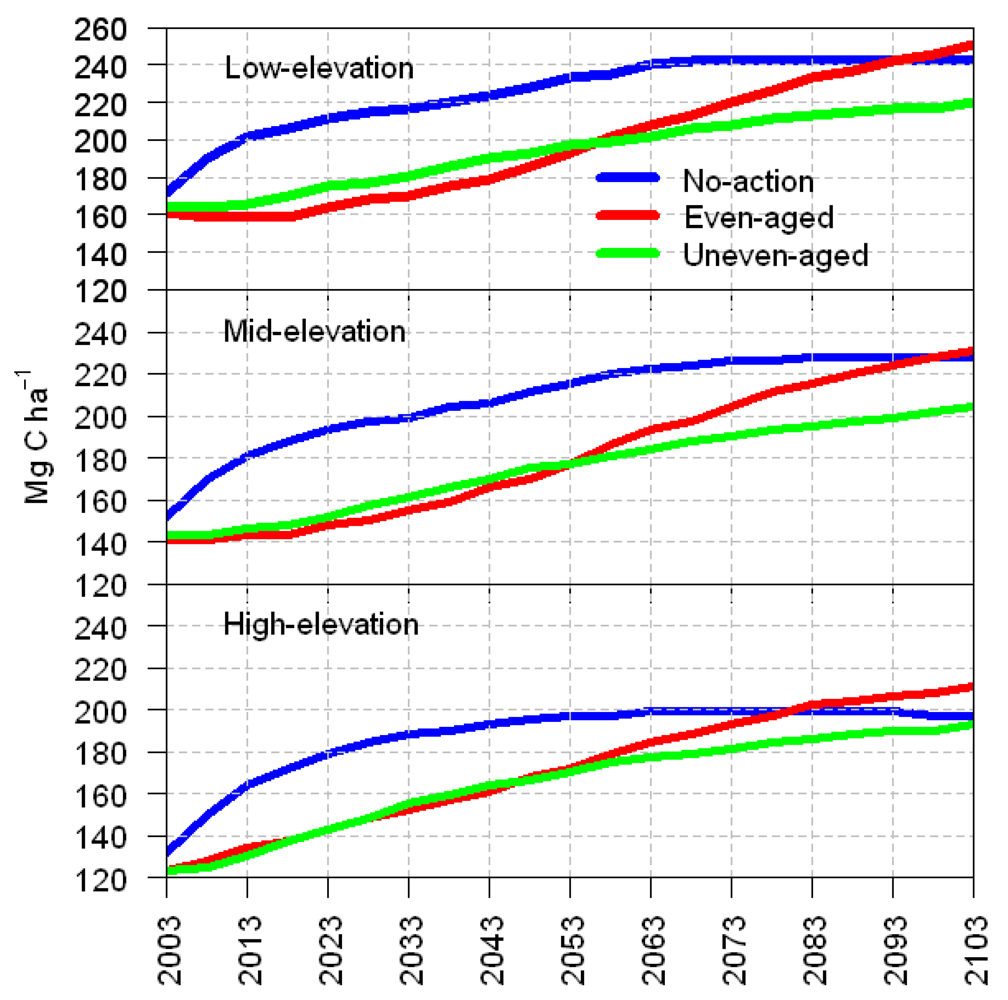
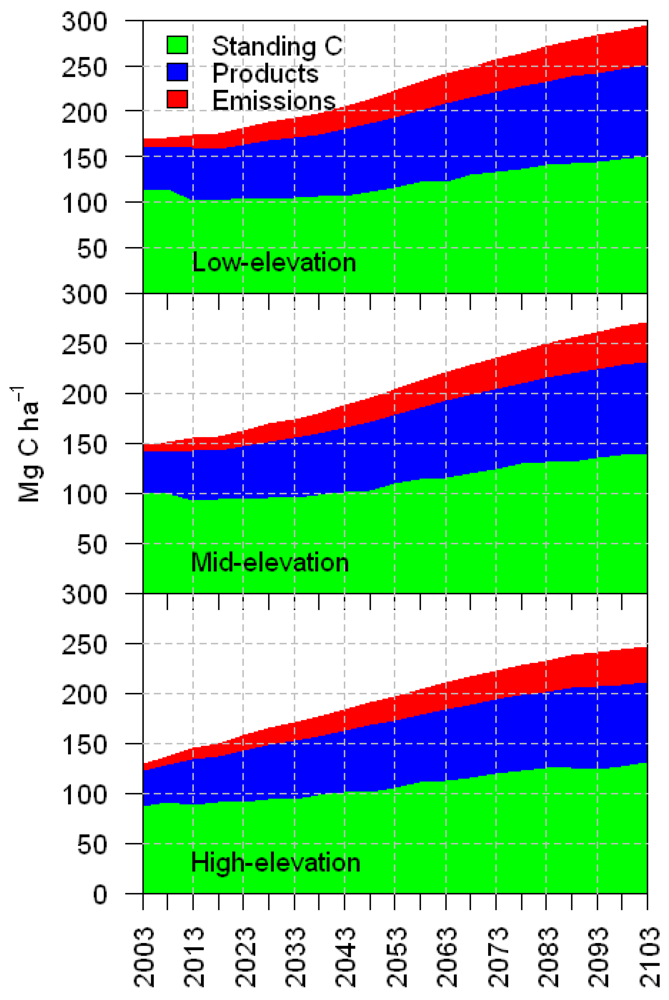
3.1. Total Carbon Sequestration
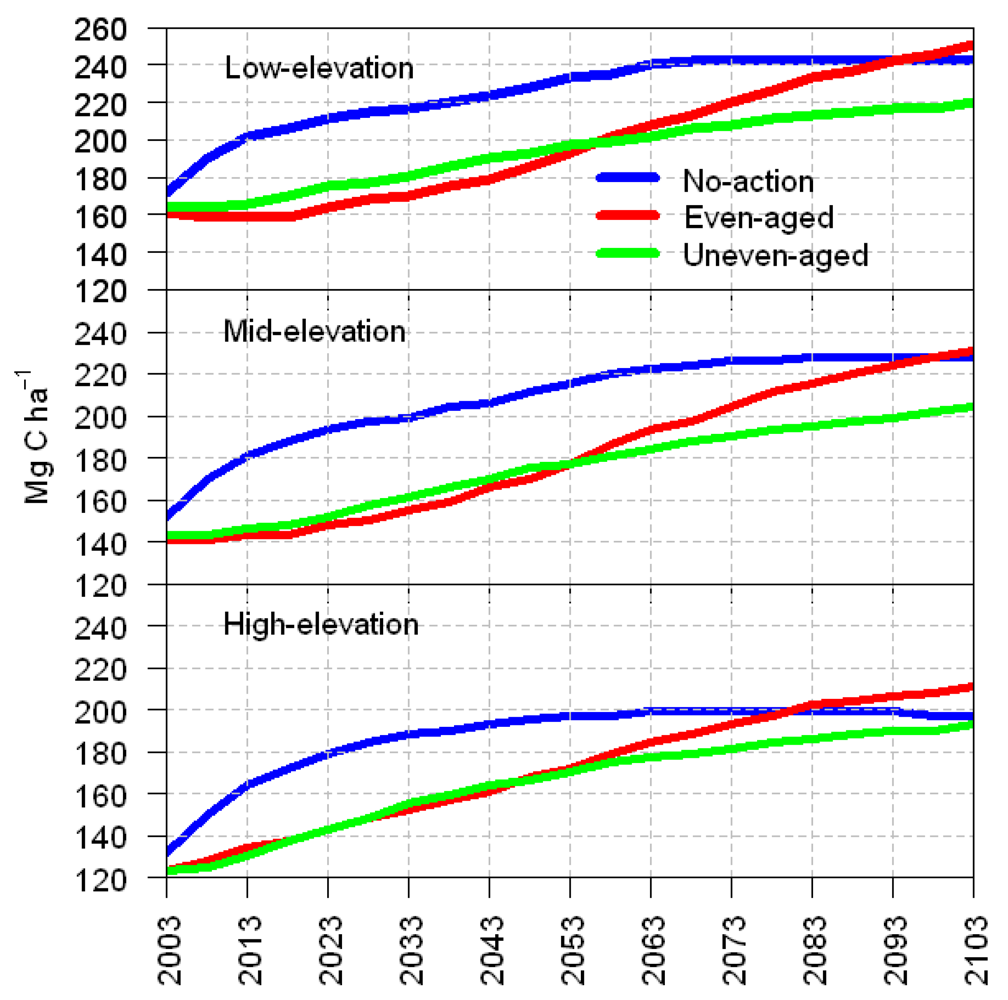
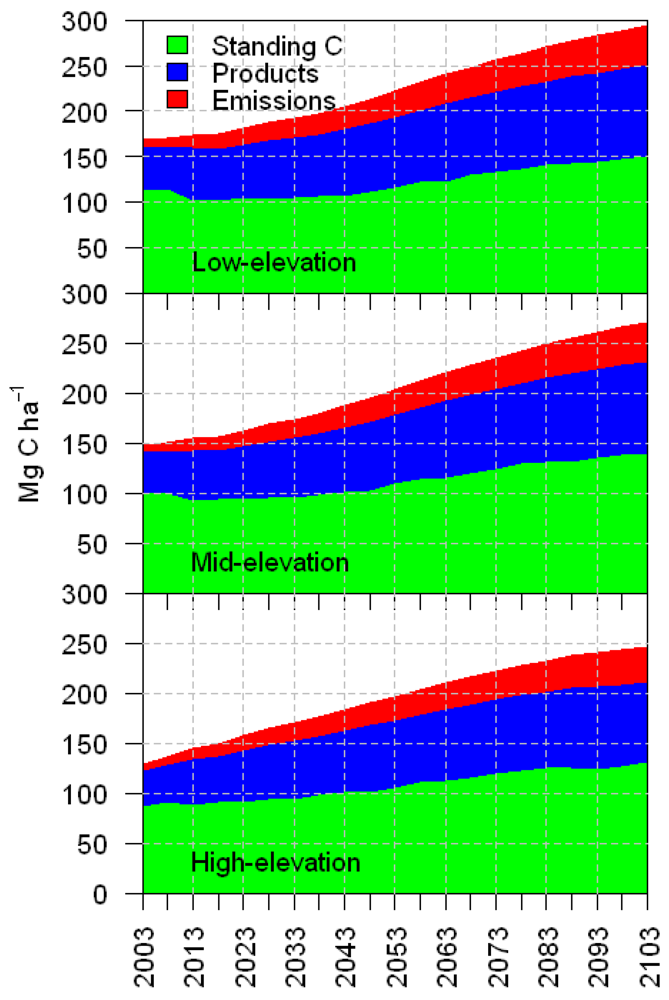
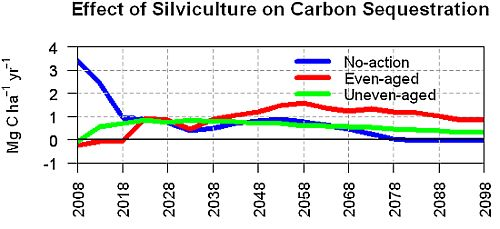
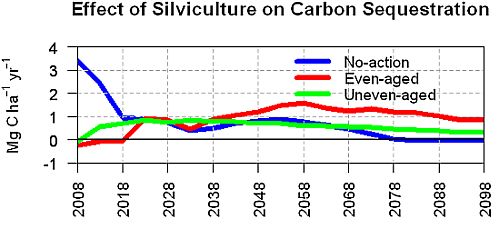
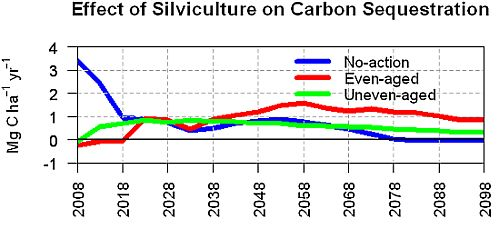
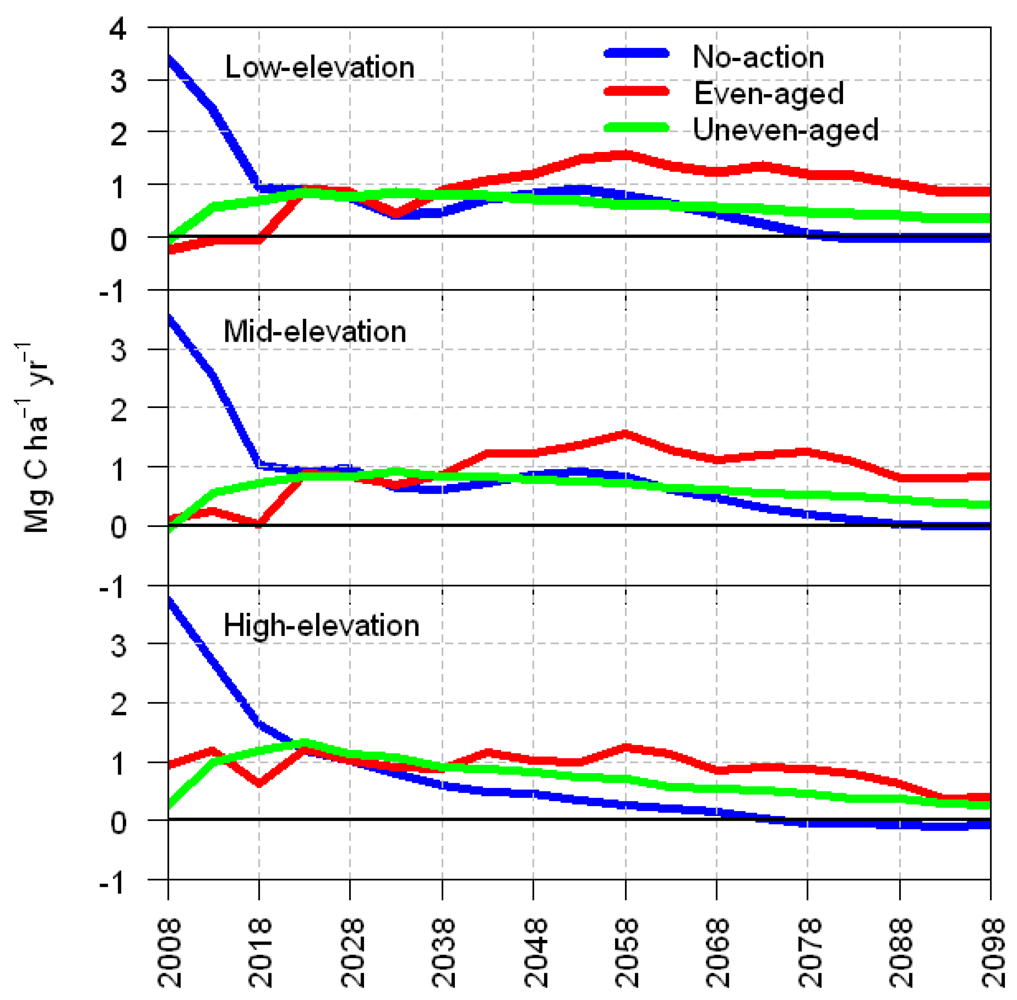
3.2. Average Annual Change in Carbon
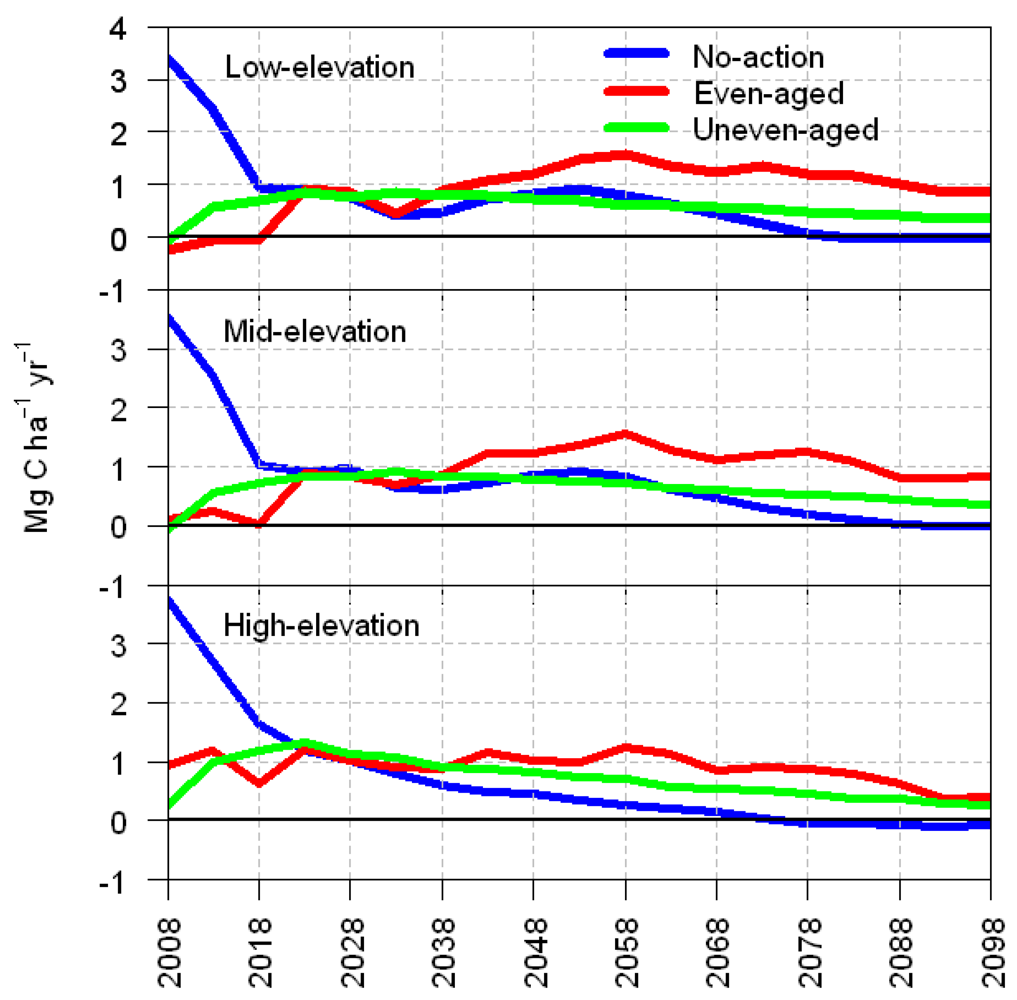
4. Discussion
5. Management Implications
Acknowledgments
Conflict of Interest
References
- Metz, B.; Davidson, O.R.; Bosch, P.R.; Dave, R.; Meyer, L.A. Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Ryan, M.G.; Harmon, M.E.; Bridsey, R.A.; Giargina, C.P.; Heath, L.S.; Houghton, R.A.; Jackson, R.B.; McKinley, D.C.; Morrison, J.F.; Murray, B.C.; et al. A synthesis of the science on forests and carbon for U.S. forests. Ecol. Soc. Am. Issue Ecol. 2010, 13, 1–16. [Google Scholar]
- Skog, K.; Nicholson, G. Carbon cycling through wood products: The role of wood and paper products in carbon sequestration. For. Prod. J. 1998, 48, 75–83. [Google Scholar]
- Harmon, M.E.; Marks, B. Effects of silvicultural practices on carbon stores in Douglas-fir-western hemlock forests in the Pacific Northwest, USA: Results from a simulation model. Can. J. For. Res. 2002, 32, 863–877. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Borner, A.; Knohl, A.; Hessenmoller, D.; Law, B.E.; Philippe, C.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar]
- Van Deusen, P. Carbon sequestration potential of forest land: Management for products and bioenergy versus preservation. Biomass Bioenergy 2010, 34, 1687–1694. [Google Scholar] [CrossRef]
- Miner, R. The 100-year method for forecasting carbon sequestration in forest products in use. Mitig. Adapt. Strateg. Glob. Change 2006. [Google Scholar]
- Hoover, C.M.; Heath, L.S. Potential gains in C storage on productive forestlands in the northeastern United States through stocking management. Ecol. Appl. 2011, 21, 1154–1161. [Google Scholar] [CrossRef] [Green Version]
- Sorenson, C.D.; Finkral, A.J.; Kolb, T.E.; Huang, C.H. Short- and long-term effects of thinning and prescribed fire on carbon stocks in ponderosa pine stands in northern Arizona. For. Ecol. Manag. 2011, 261, 460–472. [Google Scholar]
- Keyser, T.L. Thinning and site quality influence aboveground tree carbon stocks in yellow-poplar forests of the southern Appalachians. Can. J. For. Res. 2010, 40, 659–667. [Google Scholar] [CrossRef]
- Long, J.N.; Smith, F.W. Relation between size and density in developing stands: A description and possible mechanisms. For. Ecol. Manag. 1984, 7, 191–206. [Google Scholar] [CrossRef]
- Long, J.N.; Shaw, J.D. The influence of compositional and structural diversity on forest productivity. Forestry 2010, 83, 121–128. [Google Scholar] [CrossRef]
- Long, J.N. A practical approach to density management. For. Chron. 1985, 61, 23–27. [Google Scholar]
- Korstian, C.F. Perpetuation of spruce on cut-over and burned lands in the higher southern Appalachian. Ecol. Monogr. 1937, 7, 125–167. [Google Scholar] [CrossRef]
- Moore, P.T.; van Miegrot, H.; Nicholas, N.S. Relative role of understory and overstory in carbon and nitrogen cycling in a southern Appalachian spruce-fir forest. Can. J. For. Res. 2007, 37, 2689–2700. [Google Scholar] [CrossRef]
- Moore, P.T.; van Miegroet, H.; Nicholas, N.S. Examination of forest recovery scenarios in a southern Appalachian Picea-Abies forest. Forestry 2008, 81, 183–194. [Google Scholar] [CrossRef]
- Fahey, T.J.; Woodbury, P.B.; Battles, J.J.; Goodale, C.L.; Hamburg, S.P.; Ollinger, S.V.; Woodall, C.W. Forest carbon storage: Ecology, management, and policy. Front. Ecol. Environ. 2010, 8, 245–252. [Google Scholar]
- Nicholas, N.S. Stand Structure, Growth and Mortality in Southern Appalachian Spruce-Fir.
- Dull, C.W.; Ward, J.E.; Brown, H.D.; Ryan, G.W.; Clerke, W.H.; Uhler, R.J. Evaluation of Spruce and Fir Mortality in the Southern Appalachian Mountains; USDA Forest Service Southern Region(R8): Atlanta, GA, USA, 1988. [Google Scholar]
- Pearson, T.R.H.; Brown, S.L.; Birdsey, R.A. Measurement Guidelines for the Sequestration of Forest Carbon; Gen. Tech. Rep. NRS-18; USDA Forest Service: Newtown Square, PA, USA, 2007. Available online: http://www.treesearch.fs.fed.us/pubs/13292 (accessed on 21 February 2012).
- Hoover, C.M. Field Measurements for Forest Carbon Monitoring: A Landscape-Scale Approach; Springer: New York, NY, USA, 2008. [Google Scholar]
- Smith, J.E.; Heath, L.S.; Skog, K.E.; Birdsey, R.A. Methods for Calculating Forest Ecosystem and Harvested Carbon with Standard Estimates for Forest Types of the United States; Gen. Tech. Rep. NE-343; USDA Forest Service Northeastern Research Station: Newtown Square, PA, USA, 2006; p. 216. Available online: http://www.treesearch.fs.fed.us/pubs/22954 (accessed on 21 February 2012).
- Van Deusen, P.; Heath, L.S. COLE Web Applications Suite; NCASI and USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012. Available online: http://www.ncasi2.org/COLE/ (accessed on 21 February 2012).
- Crookston, N.L.; Dixon, G.E. The forest vegetation simulator: A review of its structure, content, and applications. Comput. Electron. Agric. 2005, 49, 60–80. [Google Scholar] [CrossRef]
- Reinhardt, E.D.; Crookston, N.L.; Rebain, S.A. The Fire and Fuels Extension to the Forest Vegetation Simulator; RMRS-GTR-116; USDA Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2007; p. 220. Available online: http://www.fs.fed.us/rm/pubs/rmrs_gtr116.html (accessed on 21 February 2012).
- Hoover, C.M.; Rebain, S.A. Forest Carbon Estimation Using the Forest Vegetation Simulator: Seven Things You Need to Know; Gen. Tech. Rep. NRS-77; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2011; p. 20. Available online: http://www.treesearch.fs.fed.us/pubs/37449 (accessed on 21 February 2012).
- Hurteau, M.D.; Koch, G.W.; Hungate, B.A. Carbon protection and fire risk reduction: Toward a full accounting of forest carbon offsets. Front. Ecol. Environ. 2008, 6, 493–498. [Google Scholar] [CrossRef]
- Hurteau, M.; North, M. Fuel treatment effects on tree-based forest carbon storage and emission under modeled wildfire scenarios. Front. Ecol. Environ. 2009, 7, 409–414. [Google Scholar] [CrossRef]
- Malmshimer, R.W.; Bowyer, J.L.; Fried, J.S.; Gee, E.; Izlar, R.L.; Reid, R.A.; Munn, I.A.; Oneil, E.; Stewart, W.C. Managing forests because carbon matters: Integrating energy, products, and land management policy. J. For. 2011, 109, S7–S50. [Google Scholar]
- Broekhoff, D.; Nickerson, J.; Raven, H. Forest Project Protocol, version 3.1; Climate Action Reserve: Los Angeles, CA, USA, 2009. [Google Scholar]
- Regional Greenhouse Gas Initiative Home Page. Available online: http://www.rggi.org (accessed on 21 February 2012).
- Pyle, C. The type and extent of anthropogenic vegetation disturbance in the Great Smoky Mountains before National Park Service acquisition. Castanea 1988, 53, 183–196. [Google Scholar]
- Barker, M.; van Miegroet, H.; Nicholas, N.S.; Creed, I.F. Variation in overstory nitrogen uptake in a small, high-elevation southern Appalachian spruce-fir watershed. Can. J. For. Res. 2002, 32, 1741–1752. [Google Scholar] [CrossRef]
- Pauley, E.F.; Clebsch, E.E.C. Patterns of Abies fraseri in a Great Smoky Mountains spruce fir forest. Bull. Torr. Bot. Club 1990, 117, 375–381. [Google Scholar] [CrossRef]
- McCracken, R.J.; Shanks, R.E.; Clebsch, E.E.C. Soil morphology and genesis at higher elevations of the Great Smoky Mountains. Soil Sci. Soc. Am. Proc. 1962, 26, 384–388. [Google Scholar]
- Van Miegroet, H.; Johnson, D.W.; Todd, D.E. Foliar response of red spruce saplings to fertilization with Ca and Mg in the Great Smoky Mountains National Park. Can. J. For. Res. 1993, 23, 89–95. [Google Scholar] [CrossRef]
- Van Miegroet, H.; Moore, P.T.; Tewksbury, C.E.; Nicholas, N.S. Carbon sources and sinks in high-elevation spruce-fir forests of the southeastern US. For. Ecol. Manage. 2007, 238, 249–260. [Google Scholar] [CrossRef]
- Johnson, D.W.; van Miegroet, H.; Lindberg, S.E.; Todd, D.E. Nutrient cycling in red spruce forests of the Great Smokey Mountains. Can. J. For. Res. 1991, 21, 769–787. [Google Scholar] [CrossRef]
- Shanks, R.E. Climates of the Great Smoky Mountains. Ecology 1954, 35, 353–361. [Google Scholar]
- Zedaker, S.M.; Nicholas, N.S. Quality Assurance and Methods Manual for Forest Site Classification and Field Measurements; EPA/600/3-90/082; U.S. Environmental Protection Agency: Corvallis, OR, USA, 1990; pp. 44–45. [Google Scholar]
- Keyser, C.E. Southern (SN) Variant Overview Forest Vegetation Simulator; US Forest Service, Forest Management Service Center: Fort Collins, CO, USA, 2008; p. 68. Available online: http://www.fs.fed.us/fmsc/ftp/fvs/docs/overviews/FVSsn_Overview.pdf (accessed on 21 February 2012).
- Dixon, G.E. Essential FVS: A User’s Guide to the Forest Vegetation Simulator; US Forest Service, Forest Management Service Center: Fort Collins, CO, USA, 2002; p. 208. Available online: www.fs.fed.us/fmsc/fvs/documents/userguides.shtml (accessed on 21 February 2012).
- DeRose, R.J.; Shaw, J.D.; Vacchiano, G.; Long, J.N. Improving longleaf pine mortality predictions in the Southern Variant of the Forest Vegetation Simulator. In Third Forest Vegetation Simulator Conference; RMRS-P-54; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008. [Google Scholar]
- Vacchiano, G.; Shaw, J.D.; DeRose, R.J.; Long, J.N. Inventory-based sensitivity analysis of the large tree diameter growth submodel of the Southern Variant of FVS. In Third Forest Vegetation Simulator Conference; RMRS-P-54; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008. [Google Scholar]
- Herring, N.D. Sensitivity Analysis of the Forest Vegetation Simulator Southern Variant (FVS-SN) for Southern Appalachian Hardwoods.
- Nicholas, N.S.; Zedaker, S.M. Expected stand behavior: Site quality estimation for southern Appalachian red spruce. For. Ecol. Manag. 1992, 47, 39–50. [Google Scholar] [CrossRef]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology, 9th ed; John Wiley & Sons: New York, NY, USA, 1997. [Google Scholar]
- Penman, J.; Kruger, D.; Galbally, I.; Hiraishi, T.; Nyenzi, B.; Emmanul, S.; Buendia, L.; Hoppaus, R.; Martinsen, T.; Meijer, J.; Miwa, K.; Tanabe, K. (Eds.) Intergovernmental Panel on Climate Change. Guidance Good Practice Guidance and Uncertainty Management in National Greenhouse Gas Inventories; IPCC National Greenhouse Gas Inventories Programme, Institute for Global Environmental Strategies: Hayama, Kanagawa, Japan, 2000. Available online: http://www.ipcc-nggip.iges.or.jp/public/gp/english/index.html (accessed on 21 February 2012).
- Perez-Cruzado, C.; Mohren, G.M.J.; Merino, A.; Rodriguez-Soalleiro, R. Carbon balance for different management practices for fast growing tree species planted on former pastureland in southern Europe: A case study using the CO2Fix model. Eur. J. For. Res. 2012, in press.. [Google Scholar]
- Huang, C.-H.; Sorenson, C. The economic value of sellng carbon credits from resotred forests: A case study from the navajo Nation’s tribal forests. West. J. Appl. For. 2011, 26, 37–45. [Google Scholar]
- Kolari, P.; Pumpanen, J.; Rannik, U.; Hanni, I.; Hari, P.; Berninger, F. Carbon balance of different aged Scots pine forests in southern Finland. Glob. Change Biol. 2004, 10, 1106–1119. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar]
- Drew, T.J.; Flewelling, J.W. Some recent Japanese theories of yield-density relationships and their application to Monterey pine plantations. For. Sci. 1977, 23, 517–534. [Google Scholar]
- Birdsey, R.A. Carbon Storage and Accumulation in United States Forest Ecosystems; USDA Forest Service, Washington Office (GTR-WO-59): Washington, DC, USA, 1992; p. 51. [Google Scholar]
- Liski, J.; Palosuo, T.; Peltoniemi, M.; Sievanen, R. Carbon and decomposition model Yasso for forest soils. Ecol. Mod. 2005, 189, 168–182. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Van Miegroet, H.; Olsson, M. Ecosystem disturbance and soil organic carbon—A review. In Soil Carbon in Sensitive European Ecosystems: From Science to Land Management; Jandl, R., Rodeghiero, M., Olsson, M., Eds.; John Wiley & Sons: West Sussex, UK, 2011. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moore, P.T.; DeRose, R.J.; Long, J.N.; Van Miegroet, H. Using Silviculture to Influence Carbon Sequestration in Southern Appalachian Spruce-Fir Forests. Forests 2012, 3, 300-316. https://doi.org/10.3390/f3020300
Moore PT, DeRose RJ, Long JN, Van Miegroet H. Using Silviculture to Influence Carbon Sequestration in Southern Appalachian Spruce-Fir Forests. Forests. 2012; 3(2):300-316. https://doi.org/10.3390/f3020300
Chicago/Turabian StyleMoore, Patrick T., R. Justin DeRose, James N. Long, and Helga Van Miegroet. 2012. "Using Silviculture to Influence Carbon Sequestration in Southern Appalachian Spruce-Fir Forests" Forests 3, no. 2: 300-316. https://doi.org/10.3390/f3020300
APA StyleMoore, P. T., DeRose, R. J., Long, J. N., & Van Miegroet, H. (2012). Using Silviculture to Influence Carbon Sequestration in Southern Appalachian Spruce-Fir Forests. Forests, 3(2), 300-316. https://doi.org/10.3390/f3020300