
Kennaugh Elements Allow Early Detection of Bark Beetle Infestation in Temperate Forests Using Sentinel-1 Data

 , ,
, ,  and
and
Abstract

1. Introduction
1.1. Remote Detection of Bark Beetle Infestation
1.2. Research Questions
- Is there a distinct correlation between radar backscatter recorded by Sentinel-1 time series and bark beetle infestation in spruce forests?
- Can we predict an ongoing or even future bark beetle infestation with the help of radar before it is visible in optical earth observation data?
2. Materials
2.1. Study Areas
2.2. Data and Preprocessing
2.2.1. From Sentinel-1 Data to Consistent Time Series
2.2.2. From Reference Data to Reliable Labels
2.2.3. Environmental and Hydrological Context Data
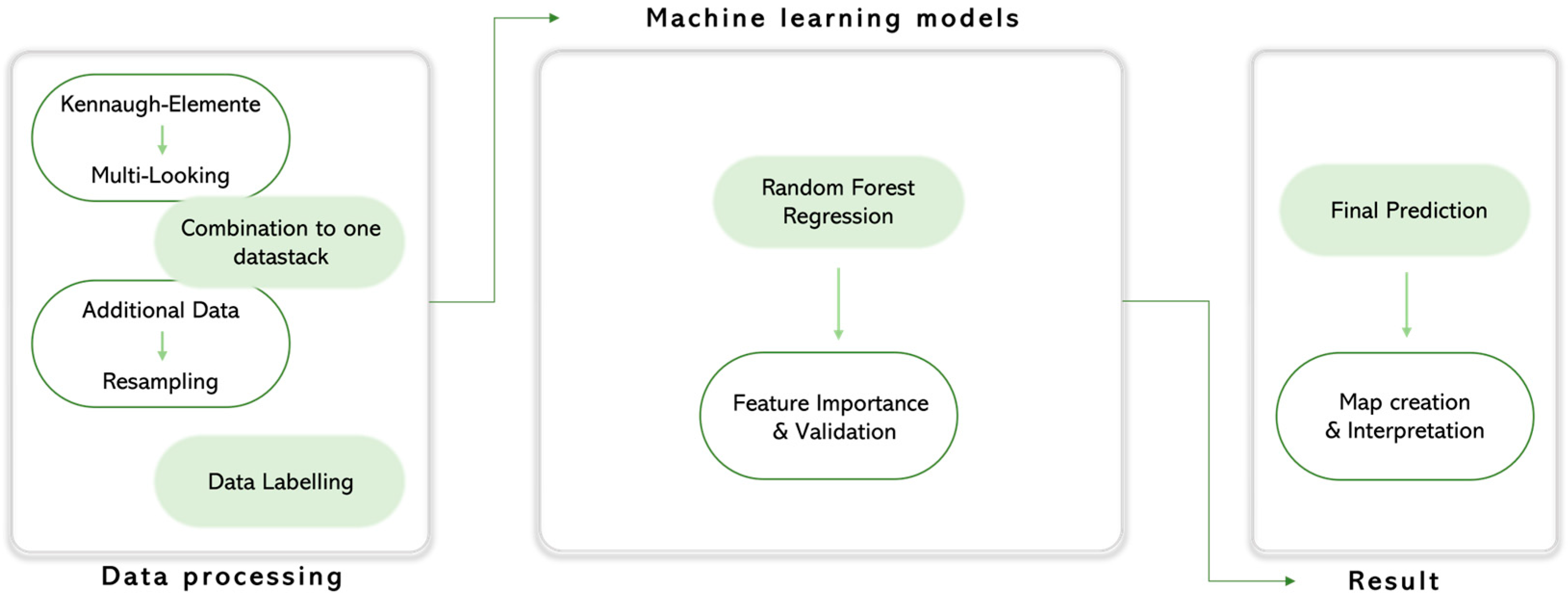
3. Methods
3.1. Trend Analysis
3.2. Vitality Prediction
3.3. Regional Transfer
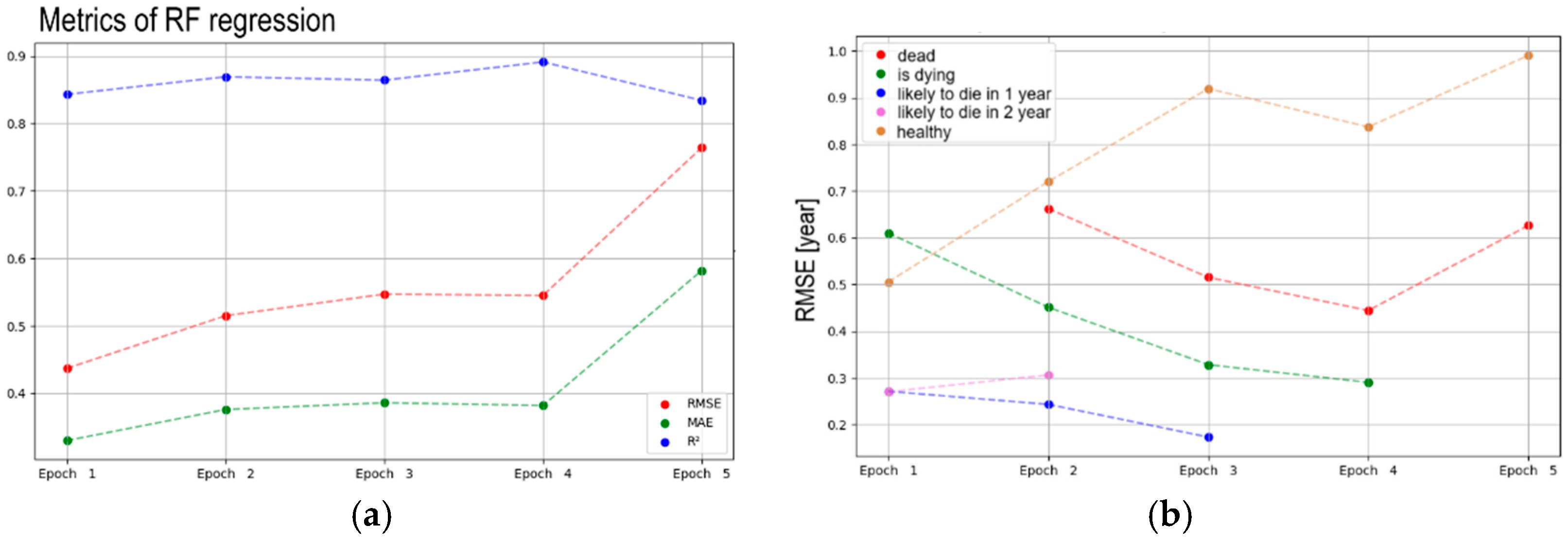
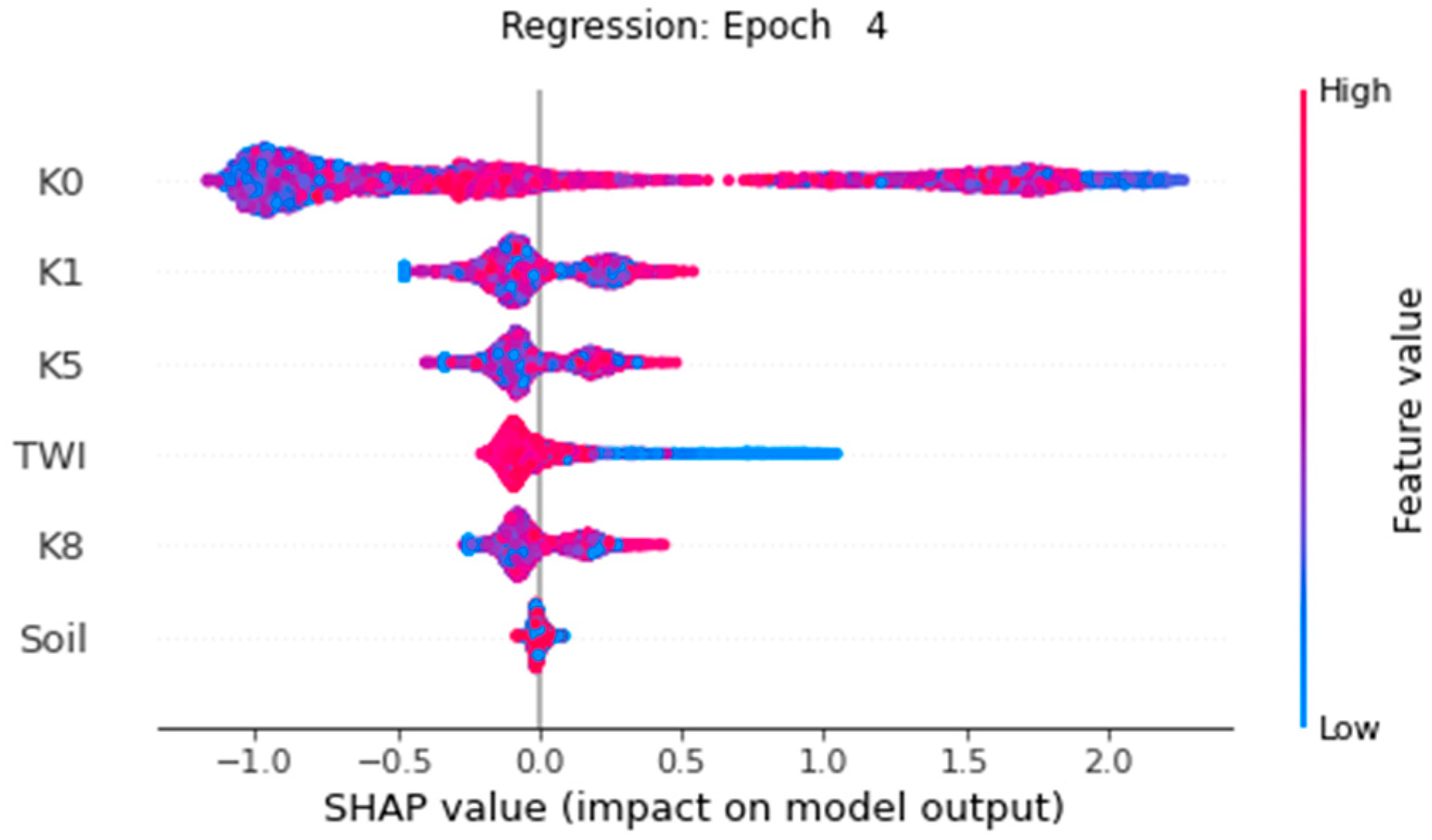
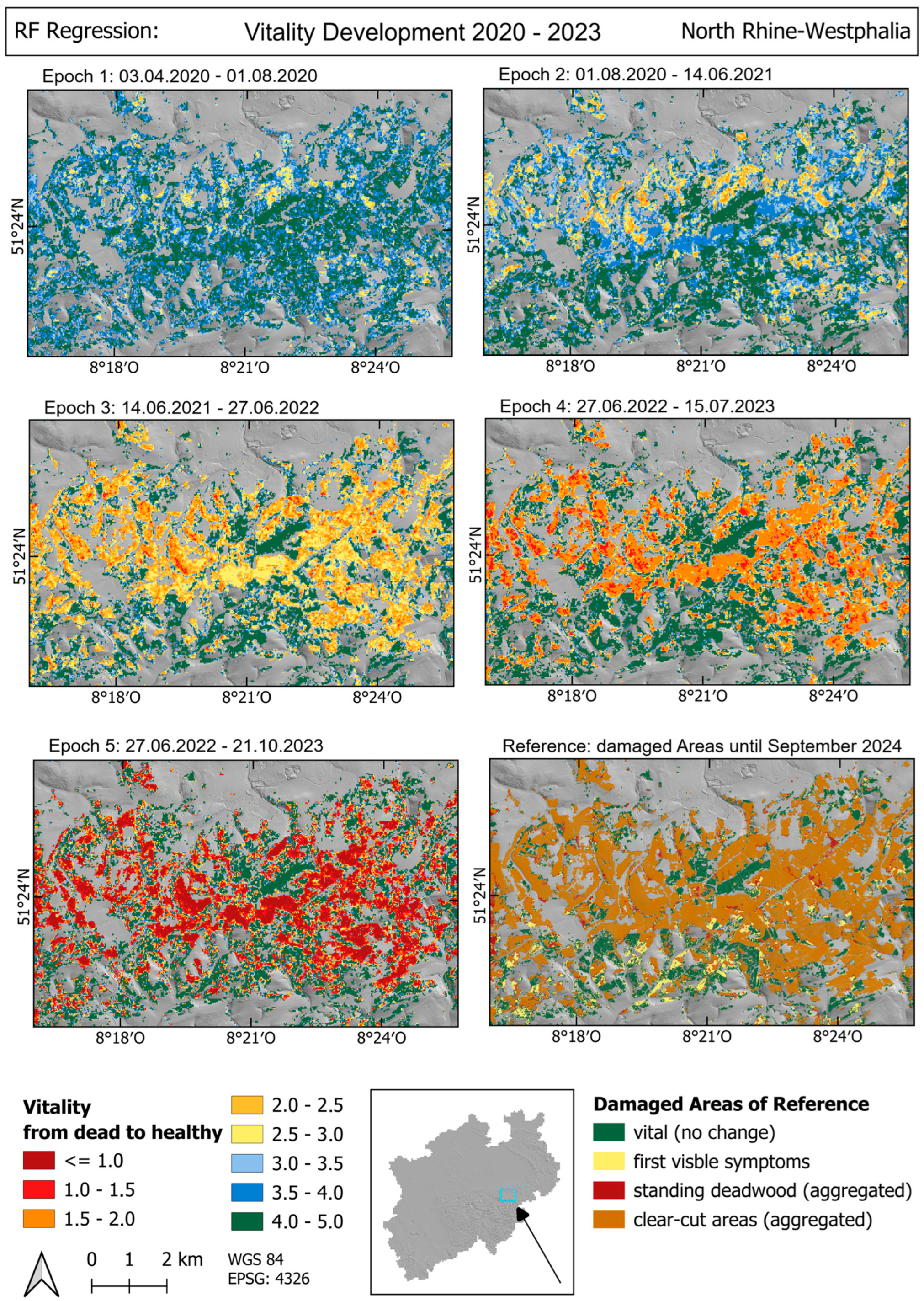
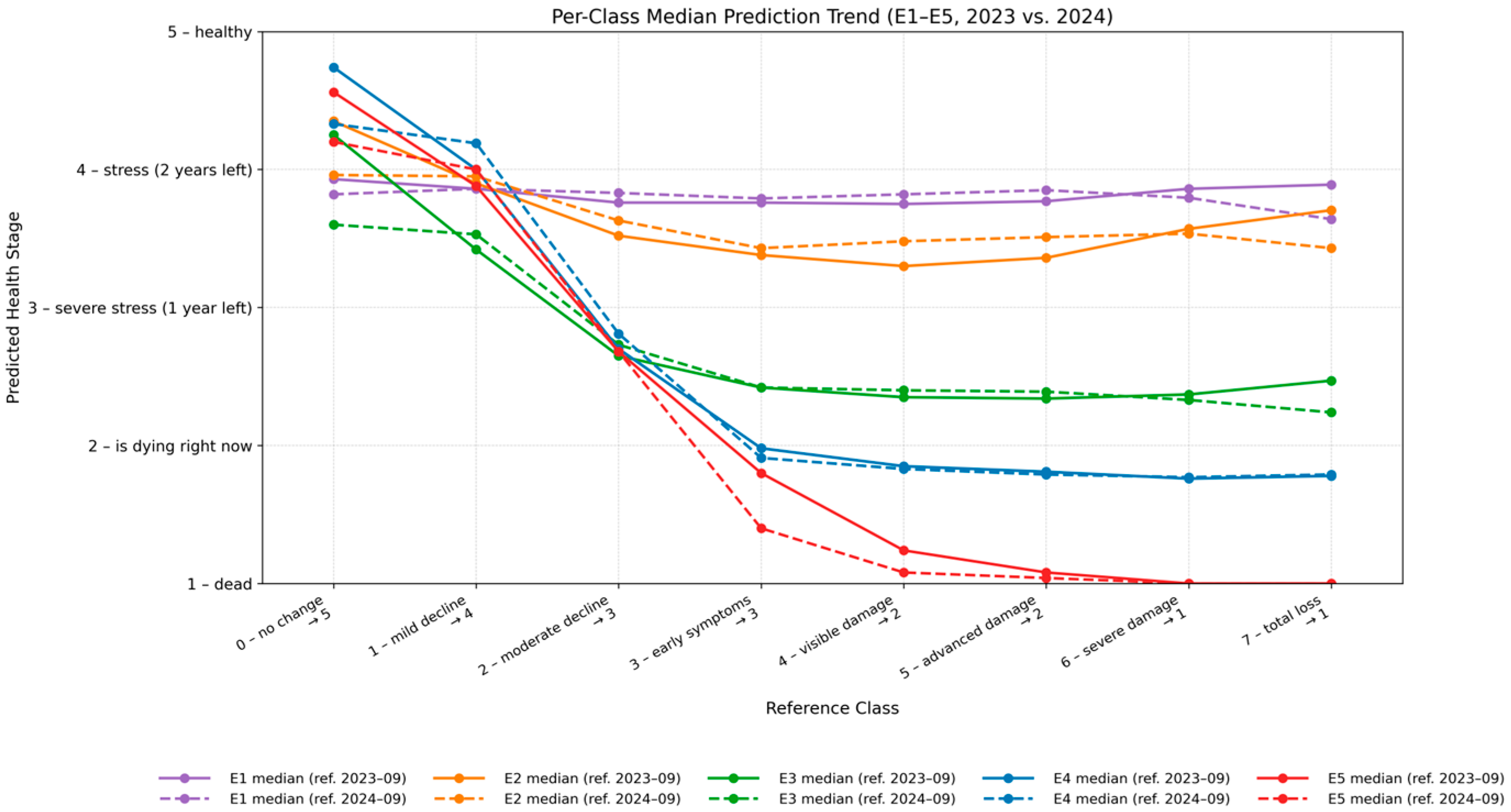
4. Results
4.1. Trend Analysis
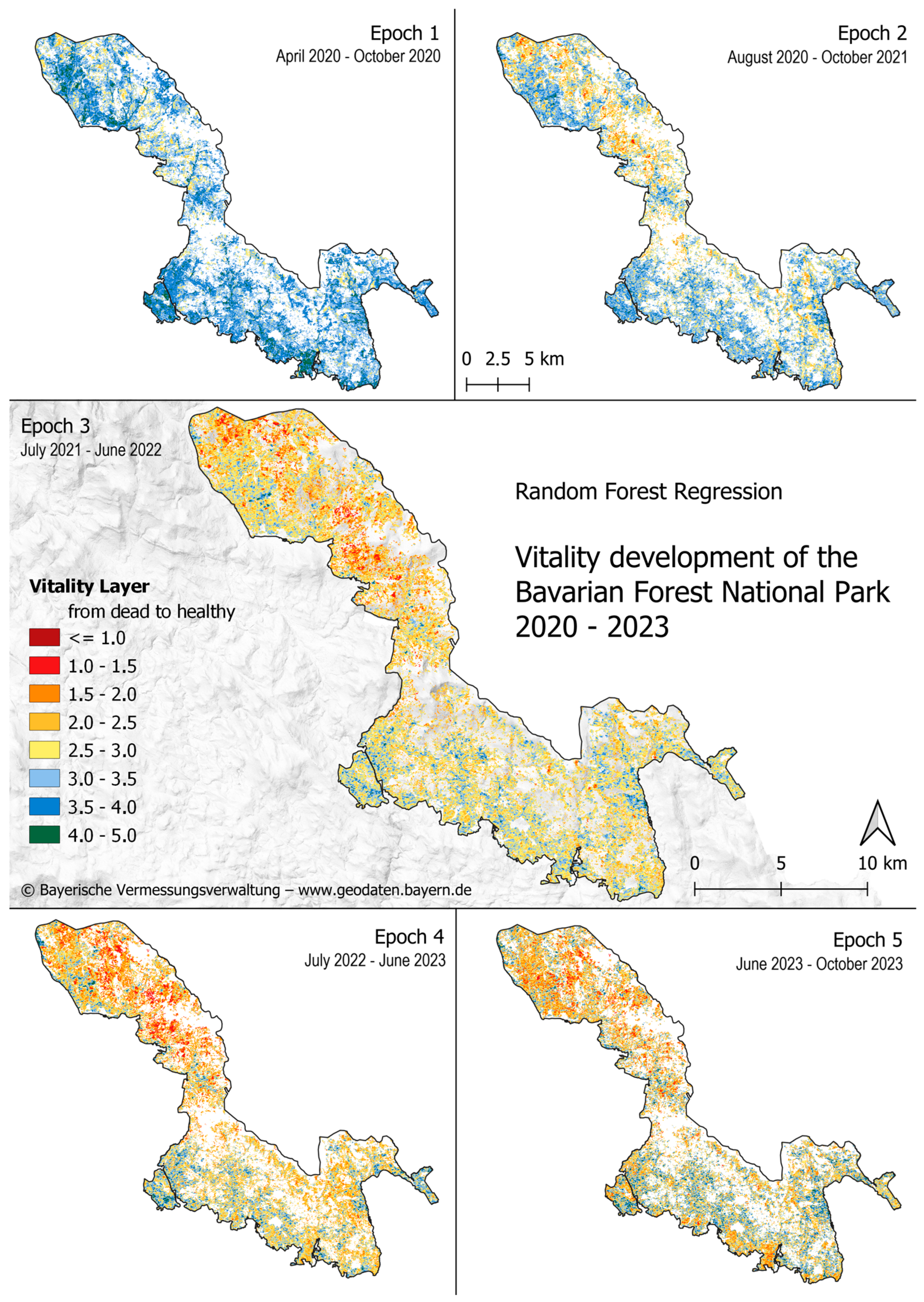
4.2. Vitality Prediction
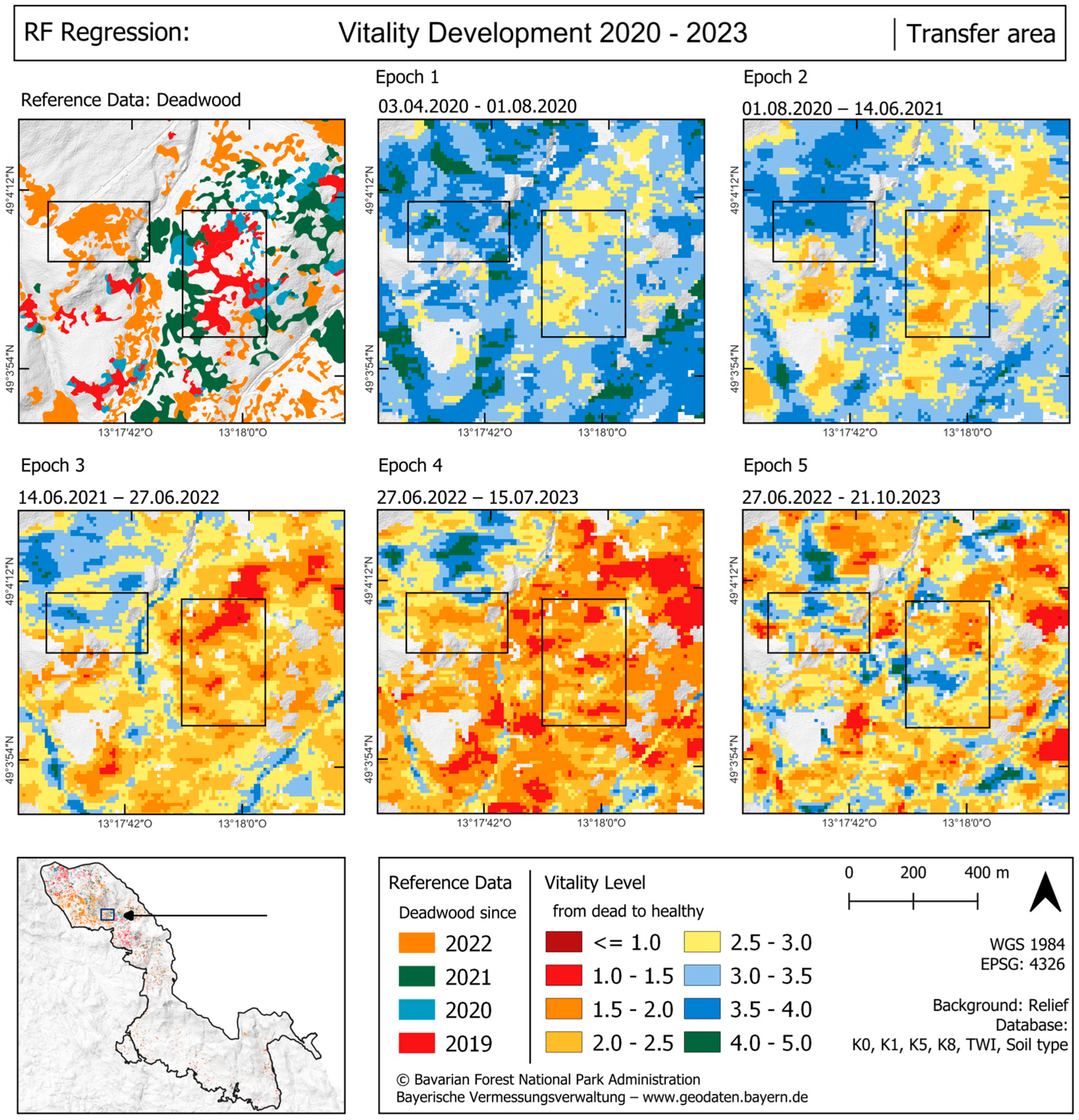
4.3. Regional Transfer
5. Discussion
5.1. Trend Analysis
5.2. Vitality Prediction
5.3. Regional Transfer
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aquino, C.; Balzarolo, M.; Chiriacò, M.V.; Santini, M. Advancing Forest Cover and Forest Cover Change Mapping for SDG 15: A Novel Approach Using Copernicus Data Products. In Proceedings of the Copernicus Meetings, EGU24-15509, Vienna, Austria, 14–19 April 2024. [Google Scholar] [CrossRef]
- Espíndola, R.P.; Ebecken, N.F.F. Advances in remote sensing for sustainable forest management: Monitoring and protecting natural resources. Rev. Caribeña Cienc. Soc. 2023, 12, 1605–1617. [Google Scholar] [CrossRef]
- Huete, A.R. Vegetation Indices, Remote Sensing and Forest Monitoring. Geogr. Compass 2012, 6, 513–532. [Google Scholar] [CrossRef]
- Massey, R.; Berner, L.T.; Foster, A.C.; Goetz, S.J.; Vepakomma, U. Remote Sensing Tools for Monitoring Forests and Tracking Their Dynamics. In Boreal Forests in the Face of Climate Change: Sustainable Management; Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 637–655. [Google Scholar] [CrossRef]
- Økland, B.; Flø, D.; Schroeder, M.; Zach, P.; Cocos, D.; Martikainen, P.; Siitonen, J.; Mandelshtam, M.Y.; Musolin, D.L.; Neuvonen, S.; et al. Range expansion of the small spruce bark beetle Ips amitinus: A newcomer in northern Europe. Agric. For. Entomol. 2019, 21, 286–298. [Google Scholar] [CrossRef]
- Biedermann, P.H.; Müller, J.; Grégoire, J.-C.; Gruppe, A.; Hagge, J.; Hammerbacher, A.; Hofstetter, R.W.; Kandasamy, D.; Kolarik, M.; Kostovcik, M.; et al. Bark Beetle Population Dynamics in the Anthropocene: Challenges and Solutions. Trends Ecol. Evol. 2019, 34, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Nationalparkverwaltung Bayerischer Wald. Habitats in the Bavarian Forest National Park. Available online: https://www.nationalpark-bayerischer-wald.bayern.de/english/nature/habitats/index.htm (accessed on 27 September 2024).
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess forest mortality is consistently linked to drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef]
- Singh, V.V.; Naseer, A.; Mogilicherla, K.; Trubin, A.; Zabihi, K.; Roy, A.; Jakuš, R.; Erbilgin, N. Understanding bark beetle outbreaks: Exploring the impact of changing temperature regimes, droughts, forest structure, and prospects for future forest pest management. Rev. Environ. Sci. Biotechnol. 2024, 23, 257–290. [Google Scholar] [CrossRef]
- Marvasti-Zadeh, S.M.; Goodsman, D.; Ray, N.; Erbilgin, N. Early Detection of Bark Beetle Attack Using Remote Sensing and Machine Learning: A Review. ACM Comput. Surv. 2023, 56, 1–40. [Google Scholar] [CrossRef]
- Fahse, L.; Heurich, M. Simulation and analysis of outbreaks of bark beetle infestations and their management at the stand level. Ecol. Model. 2011, 222, 1833–1846. [Google Scholar] [CrossRef]
- Kautz, M.; Feurer, J.; Adler, P. Early detection of bark beetle (Ips typographus) infestations by remote sensing—A critical review of recent research. For. Ecol. Manag. 2024, 556, 121595. [Google Scholar] [CrossRef]
- Ghorbanian, M.; Karimi-Malati, A.; Jalaeian, M.; Sangani, M.F. Maximum entropy modelling to predict the impact of abiotic variables on the potential distribution of Orthotomicus erosus (Wollaston) (Coleoptera, Curculionidae, Scolytinae). J. Insect Biodivers. Syst. 2023, 9, 711–725. [Google Scholar] [CrossRef]
- Lissovsky, A.A.; Dudov, S.V. Species-Distribution Modeling: Advantages and Limitations of Its Application. 2. MaxEnt. Biol. Bull. Rev. 2021, 11, 265–275. [Google Scholar] [CrossRef]
- Öhrn, P.; Långström, B.; Lindelöw, Å.; Björklund, N. Seasonal flight patterns of Ips typographus in southern S weden and thermal sums required for emergence. Agric. For. Entomol. 2014, 16, 147–157. [Google Scholar] [CrossRef]
- Heurich, M.; Beudert, B.; Rall, H.; Křenová, Z. National Parks as Model Regions for Interdisciplinary Long-Term Ecological Research: The Bavarian Forest and Šumavá National Parks Underway to Transboundary Ecosystem Research. In Long-Term Ecological Research; Müller, F., Baessler, C., Schubert, H., Klotz, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 327–344. [Google Scholar] [CrossRef]
- Marini, L.; Lindelöw, Å.; Jönsson, A.M.; Wulff, S.; Schroeder, L.M. Population dynamics of the spruce bark beetle: A long-term study. Oikos 2013, 122, 1768–1776. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Harding, S.; Bärring, L.; Ravn, H.P. Impact of climate change on the population dynamics of Ips typographus in southern Sweden. Agric. For. Meteorol. 2007, 146, 70–81. [Google Scholar] [CrossRef]
- Hollaus, M.; Vreugdenhil, M. Radar Satellite Imagery for Detecting Bark Beetle Outbreaks in Forests. Curr. For. Rep. 2019, 5, 240–250. [Google Scholar] [CrossRef]
- Huo, L.; Persson, H.J.; Lindberg, E. Early detection of forest stress from European spruce bark beetle attack, and a new vegetation index: Normalized distance red & SWIR (NDRS). Remote Sens. Environ. 2021, 255, 112240. [Google Scholar] [CrossRef]
- Abdullah, H.; Skidmore, A.K.; Darvishzadeh, R.; Heurich, M. Timing of red-edge and shortwave infrared reflectance critical for early stress detection induced by bark beetle (Ips typographus, L.) attack. Int. J. Appl. Earth Obs. Geoinf. 2019, 82, 101900. [Google Scholar] [CrossRef]
- Ali, A.M.; Abdullah, H.; Darvishzadeh, R.; Skidmore, A.K.; Heurich, M.; Roeoesli, C.; Paganini, M.; Heiden, U.; Marshall, D. Canopy chlorophyll content retrieved from time series remote sensing data as a proxy for detecting bark beetle infestation. Remote Sens. Appl. Soc. Environ. 2021, 22, 100524. [Google Scholar] [CrossRef]
- Pulliainen, J.; Hari, P.; Hallikainen, M.; Patrikainen, N.; Peramaki, M.; Kolari, P. Monitoring of soil moisture and vegetation water content variations in boreal forest from C-band SAR data. In Proceedings of the IGARSS 2004, 2004 IEEE International Geoscience and Remote Sensing Symposium, Anchorage, AK, USA, 20–24 September 2004; pp. 1013–1016. [Google Scholar] [CrossRef]
- Bernardino, P.N.; Oliveira, R.S.; Van Meerbeek, K.; Hirota, M.; Furtado, M.N.; A Sanches, I.; Somers, B. Estimating vegetation water content from Sentinel-1 C-band SAR data over savanna and grassland ecosystems. Environ. Res. Lett. 2024, 19, 034019. [Google Scholar] [CrossRef]
- Vreugdenhil, M.; Wagner, W.; Bauer-Marschallinger, B.; Pfeil, I.; Teubner, I.; Rüdiger, C.; Strauss, P. Sensitivity of Sentinel-1 Backscatter to Vegetation Dynamics: An Austrian Case Study. Remote Sens. 2018, 10, 1396. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef]
- Abdullah, H.; Skidmore, A.K.; Darvishzadeh, R.; Heurich, M.; Pettorelli, N.; Disney, M. Sentinel-2 accurately maps green-attack stage of European spruce bark beetle (Ips typographus, L.) compared with Landsat-8. Remote Sens. Ecol. Conserv. 2018, 5, 87–106. [Google Scholar] [CrossRef]
- König, S.; Thonfeld, F.; Förster, M.; Dubovyk, O.; Heurich, M. Assessing Combinations of Landsat, Sentinel-2 and Sentinel-1 Time series for Detecting Bark Beetle Infestations. GISci. Remote Sens. 2023, 60, 2226515. [Google Scholar] [CrossRef]
- Bárta, V.; Lukeš, P.; Homolová, L. Early detection of bark beetle infestation in Norway spruce forests of Central Europe using Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102335. [Google Scholar] [CrossRef]
- Hesslerová, P.; Huryna, H.; Pokorný, J.; Procházka, J. The effect of forest disturbance on landscape temperature. Ecol. Eng. 2018, 120, 345–354. [Google Scholar] [CrossRef]
- Tanase, M.A.; Aponte, C.; Mermoz, S.; Bouvet, A.; Le Toan, T.; Heurich, M. Detection of windthrows and insect outbreaks by L-band SAR: A case study in the Bavarian Forest National Park. Remote Sens. Environ. 2018, 209, 700–711. [Google Scholar] [CrossRef]
- Schmitt, A.; Wendleder, A.; Hinz, S. The Kennaugh element framework for multi-scale, multi-polarized, multi-temporal and multi-frequency SAR image preparation. ISPRS J. Photogramm. Remote Sens. 2015, 102, 122–139. [Google Scholar] [CrossRef]
- Hoekman, D.H. Measurements of the backscatter and attenuation properties of forest stands at X-, C- and L-band. Remote Sens. Environ. 1987, 23, 397–416. [Google Scholar] [CrossRef]
- Konings, A.G.; Rao, K.; Steele-Dunne, S.C. Macro to micro: Microwave remote sensing of plant water content for physiology and ecology. New Phytol. 2019, 223, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Buddenbaum, H.; Nink, S.; Hill, J. Potential of Sentinel-1 Data for Spatially and Temporally High-Resolution Detection of Drought Affected Forest Stands. Forests 2022, 13, 2148. [Google Scholar] [CrossRef]
- Bae, S.; Müller, J.; Förster, B.; Hilmers, T.; Hochrein, S.; Jacobs, M.; Leroy, B.M.L.; Pretzsch, H.; Weisser, W.W.; Mitesser, O. Tracking the temporal dynamics of insect defoliation by high-resolution radar satellite data. Methods Ecol. Evol. 2021, 13, 121–132. [Google Scholar] [CrossRef]
- Nationalparkverwaltung Bayerischer Wald. Profile of Bavarian Forest National Park. Available online: https://www.nationalpark-bayerischer-wald.bayern.de/english/about_us/profile/index.htm (accessed on 27 September 2024).
- Seidl, R.; Müeller, J.; Hothorn, T.; Bässler, C.; Heurich, M.; Kautz, M. Small beetle, large-scale drivers: How regional and landscape factors affect outbreaks of the European spruce bark beetle. J. Appl. Ecol. 2016, 53, 530–540. [Google Scholar] [CrossRef]
- Latifi, H.; Holzwarth, S.; Skidmore, A.; Brůna, J.; Červenka, J.; Darvishzadeh, R.; Hais, M.; Heiden, U.; Homolová, L.; Krzystek, P.; et al. A laboratory for conceiving Essential Biodiversity Variables (EBVs)—The “Data pool initiative for the Bohemian Forest Ecosystem”. Methods Ecol. Evol. 2021, 12, 2073–2083. [Google Scholar] [CrossRef]
- Landesbetrieb Wald und Holz Nordrhein-Westfalen. Datenbestand des Landesbetriebes Wald und Holz NRW [Data from the State Forestry and Timber Agency of North Rhine-Westphalia]. 2024. Available online: https://www.wms.nrw.de/umwelt/waldNRW?SERVICE=WMS&REQUEST=GetCapabilities (accessed on 16 June 2025).
- Bertram, A.; Wendleder, A.; Schmitt, A.; Huber, M. Long-term Monitoring of water dynamics in the Sahel region using the Multi-SAR-System. In International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences—ISPRS Archives; International Society for Photogrammetry and Remote Sensing (ISPRS), Publisher: Prague, Czech Republic, 2016; pp. 9–16. [Google Scholar] [CrossRef]
- Small, D. Flattening Gamma: Radiometric Terrain Correction for SAR Imagery. IEEE Trans. Geosci. Remote Sens. 2011, 49, 3081–3093. [Google Scholar] [CrossRef]
- Akhavan, Z.; Hasanlou, M.; Hosseini, M. A Comparison of Tree-based Regression Models for Soil Moisture Estimation using SAR Data. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2023, X-4/W1-202, 37–42. [Google Scholar] [CrossRef]
- Bai, X.; He, B.; Li, X.; Zeng, J.; Wang, X.; Wang, Z.; Zeng, Y.; Su, Z. First Assessment of Sentinel-1A Data for Surface Soil Moisture Estimations Using a Coupled Water Cloud Model and Advanced Integral Equation Model over the Tibetan Plateau. Remote Sens. 2017, 9, 714. [Google Scholar] [CrossRef]
- Le Hegarat-Mascle, S.; Zribi, M.; Alem, F.; Weisse, A.; Loumagne, C. Soil moisture estimation from ERS/SAR data: Toward an operational methodology. IEEE Trans. Geosci. Remote Sens. 2002, 40, 2647–2658. [Google Scholar] [CrossRef]
- Moreira, A.; Prats-Iraola, P.; Younis, M.; Krieger, G.; Hajnsek, I.; Papathanassiou, K.P. A tutorial on synthetic aperture radar. IEEE Geosci. Remote Sens. Mag. 2013, 1, 6–43. [Google Scholar] [CrossRef]
- Touzi, R. Target Scattering Decomposition in Terms of Roll-Invariant Target Parameters. IEEE Trans. Geosci. Remote Sens. 2007, 45, 73–84. [Google Scholar] [CrossRef]
- Landesbetrieb Wald und Holz Nordrhein-Westfalen. Vitalitätsabnahme und Kalamitätskarte Nadelwald NRW [Vitality Decline and Calamity Map of Coniferous Forests in North Rhine-Westphalia]. 2024. Available online: https://open.nrw/dataset/a0fab723-b852-4f8f-adf5-2723699149ff (accessed on 16 June 2025).
- Landesbetrieb Wald und Holz Nordrhein-Westfalen. Vitalitätsabnahme Nadelwald NRW [Decrease in vitality of coniferous forest in North Rhine-Westphalia]. 2024. Available online: https://www.opengeodata.nrw.de/produkte/umwelt_klima/wald_forst/fernerkundung/ (accessed on 16 June 2025).
- Julius Kühn-Institut. Topographischer Feuchteindex. Available online: https://wms.flf.julius-kuehn.de/cgi-bin/twi/qgis_mapserv.fcgi (accessed on 10 February 2024).
- Bayerisches Landesamt für Umwelt. Übersichtsbodenkarte 1:25.000. Available online: https://www.lfu.bayern.de/boden/karten_daten/uebk25/index.htm (accessed on 19 October 2024).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Kranjčić, N.; Cetl, V.; Matijević, H.; Markovinović, D. Comparing Different Machine Learning Options to Map Bark Beetle Infestations in Croatia. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2023, XLVIII-4/W, 83–88. [Google Scholar] [CrossRef]
- Andresini, G.; Appice, A.; Ienco, D.; Malerba, D.; Recchia, V. Potential of Spectral-Spatial Analysis to Map Forest Tree Dieback Due to Bark Beetle Hotspots in Sentinel-2 Images. In Proceedings of the IGARSS 2024—2024 IEEE International Geoscience and Remote Sensing Symposium, Athens, Greece, 7–12 July 2024; pp. 5227–5230. [Google Scholar] [CrossRef]
- Rüetschi, M.; Schaepman, M.E.; Small, D. Using Multitemporal Sentinel-1 C-band Backscatter to Monitor Phenology and Classify Deciduous and Coniferous Forests in Northern Switzerland. Remote Sens. 2018, 10, 55. [Google Scholar] [CrossRef]
- Pirotti, F.; Adedipe, O.; Leblon, B. Sentinel-1 Response to Canopy Moisture in Mediterranean Forests before and after Fire Events. Remote Sens. 2023, 15, 823. [Google Scholar] [CrossRef]
- Drever, C.R.; Peterson, G.; Messier, C.; Bergeron, Y.; Flannigan, M. Can forest management based on natural disturbances maintain ecological resilience? Can. J. For. Res. 2006, 36, 2285–2299. [Google Scholar] [CrossRef]
- Matuszkiewicz, J.M.; Affek, A.N.; Zaniewski, P.; Kołaczkowska, E. Early response of understory vegetation to the mass dieback of Norway spruce in the European lowland temperate forest. For. Ecosyst. 2024, 11, 100177. [Google Scholar] [CrossRef]
- Loaiciga, H.A.; Michaelsen, J.; Hudak, P.F. Truncated Distributions in Hydrologic Analysis. JAWRA J. Am. Water Resour. Assoc. 1992, 28, 853–863. [Google Scholar] [CrossRef]
- Yu, K.; Guo, X.; Liu, L.; Li, J.; Wang, H.; Ling, Z.; Wu, X. Causality-based Feature Selection: Methods and Evaluations. ACM Comput. Surv. 2020, 53, 1–36. [Google Scholar] [CrossRef]
- Štofko, P.; Kodrík, M. Are there any differences in the root branch architecture of Norway spruce trees growing on two sites with different water regime? Folia For. Polonica. Ser. A Forestry 2009, 51, 181–184. [Google Scholar] [CrossRef]
- Jin, S.; Su, Y.; Gao, S.; Hu, T.; Liu, J.; Guo, Q. The Transferability of Random Forest in Canopy Height Estimation from Multi-Source Remote Sensing Data. Remote Sens. 2018, 10, 1183. [Google Scholar] [CrossRef]
- Hoppe, H.; Dietrich, P.; Marzahn, P.; Weiß, T.; Nitzsche, C.; von Lukas, U.F.; Wengerek, T.; Borg, E. Transferability of Machine Learning Models for Crop Classification in Remote Sensing Imagery Using a New Test Methodology: A Study on Phenological, Temporal, and Spatial Influences. Remote Sens. 2024, 16, 1493. [Google Scholar] [CrossRef]
- Qiu, C.; Li, H.; Guo, W.; Chen, X.; Yu, A.; Tong, X.; Schmitt, M. Transferring Transformer-Based Models for Cross-Area Building Extraction From Remote Sensing Images. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2022, 15, 4104–4116. [Google Scholar] [CrossRef]
- Schmitt, A.; Wendleder, A.; Kleynmans, R.; Hell, M.; Roth, A.; Hinz, S. Multi-Source and Multi-Temporal Image Fusion on Hypercomplex Bases. Remote Sens. 2020, 12, 943. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
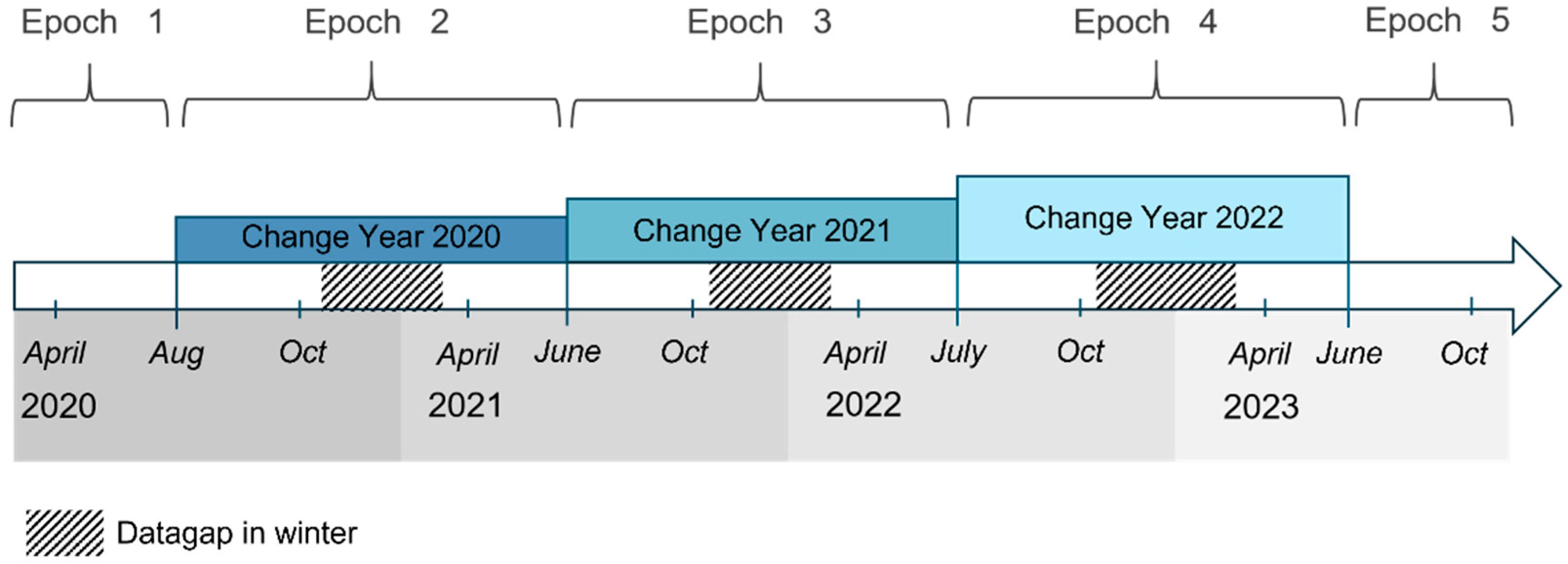
| Change Year | Start Date | Stop Date |
|---|---|---|
| 2019 | 27 June 2019 | 1 August 2020 |
| 2020 | 1 August 2020 | 14 June 2021 |
| 2021 | 14 June 2021 | 27 June 2022 |
| 2022 | 27 June 2022 | 15 July 2023 |
| Epoch | Epoch Time Frame (EO Data) | Predicted “Dead in Year” | Years Before Death (Δt) | Assigned Vitality Class | Predicted Vitality Stage |
|---|---|---|---|---|---|
| E1 | April 2020– October 2020 | 2020 | 0 | 1 | Dead |
| 2021 | +1 | 2 | Is dying right now | ||
| 2022 | +2 | 3 | Severe stress (early indicator) | ||
| E2 | August 2020– October 2021 | 2021 | 0 | 1 | Dead |
| 2022 | +1 | 2 | Is dying right now | ||
| 2023 | +2 | 3 | Severe stress (early indicator) | ||
| E3 | July 2021– June 2022 | 2022 | 0 | 1 | Dead |
| 2023 | +1 | 2 | Is dying right now | ||
| 2024 | +2 | 3 | Severe stress (early indicator) | ||
| E4 | July 2022– June 2023 | 2023 | 0 | 1 | Dead |
| 2024 | +1 | 2 | Is dying right now | ||
| 2025 | +2 | 3 | Severe stress (early indicator) | ||
| E5 | June 2023– October 2023 | 2024 | 0 | 1 | Dead |
| 2025 | +1 | 2 | Is dying right now | ||
| 2026 | +2 | 3 | Severe stress (early indicator) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hechtl, C.; Hauser, S.; Schmitt, A.; Heurich, M.; Wendleder, A. Kennaugh Elements Allow Early Detection of Bark Beetle Infestation in Temperate Forests Using Sentinel-1 Data. Forests 2025, 16, 1272. https://doi.org/10.3390/f16081272
Hechtl C, Hauser S, Schmitt A, Heurich M, Wendleder A. Kennaugh Elements Allow Early Detection of Bark Beetle Infestation in Temperate Forests Using Sentinel-1 Data. Forests. 2025; 16(8):1272. https://doi.org/10.3390/f16081272
Chicago/Turabian StyleHechtl, Christine, Sarah Hauser, Andreas Schmitt, Marco Heurich, and Anna Wendleder. 2025. "Kennaugh Elements Allow Early Detection of Bark Beetle Infestation in Temperate Forests Using Sentinel-1 Data" Forests 16, no. 8: 1272. https://doi.org/10.3390/f16081272
APA StyleHechtl, C., Hauser, S., Schmitt, A., Heurich, M., & Wendleder, A. (2025). Kennaugh Elements Allow Early Detection of Bark Beetle Infestation in Temperate Forests Using Sentinel-1 Data. Forests, 16(8), 1272. https://doi.org/10.3390/f16081272