

Prediction of the Potentially Suitable Area for Anoplophora glabripennis (Coleoptera: Cerambycidae) in China Based on MaxEnt
Abstract
1. Introduction
2. Materials and Methods
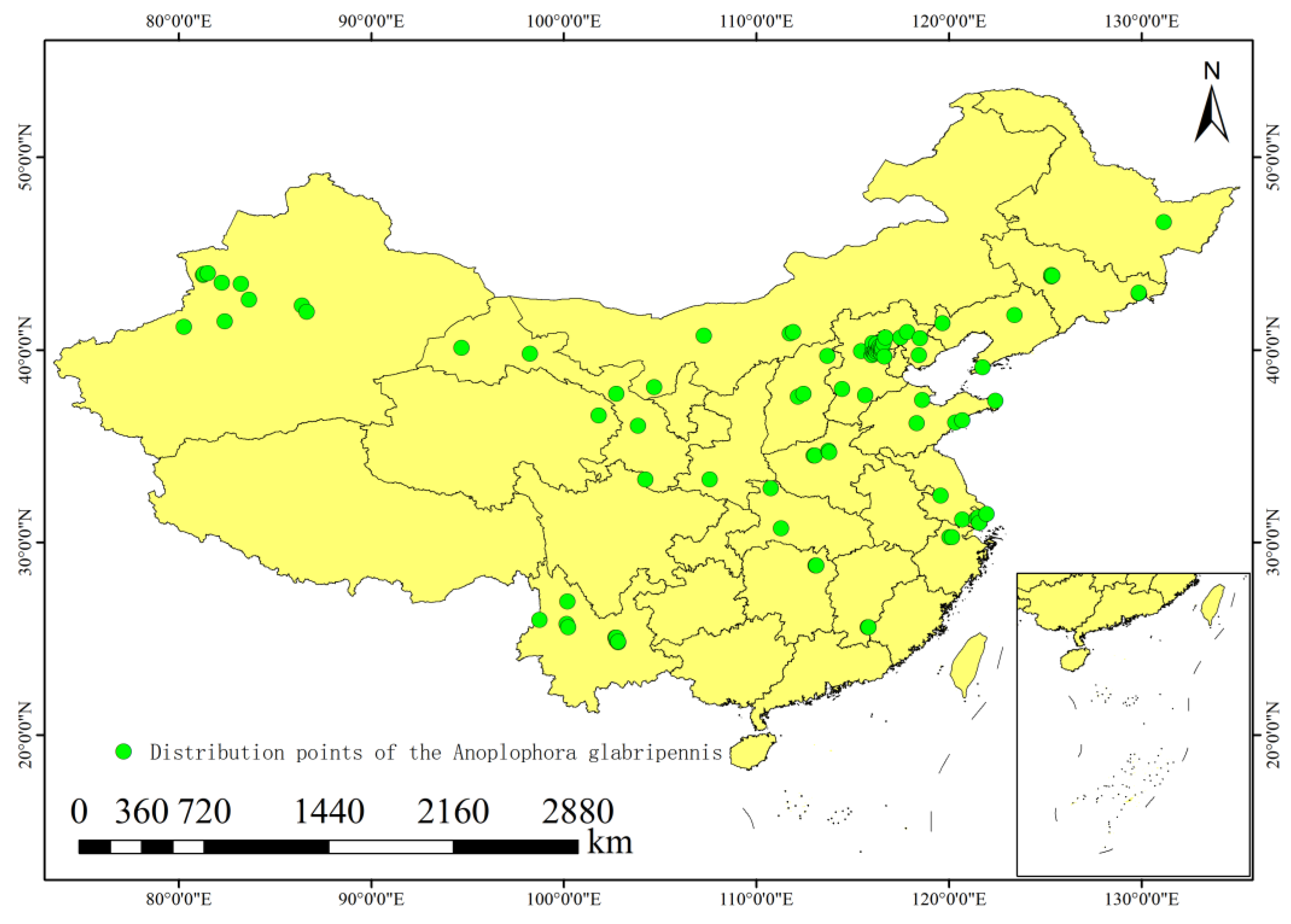
2.1. Occurrence Point Data for A. glabripennis (Asian longhorned beetle, ALB)
2.2. Environmental Variable Data
2.3. Geographic Data
2.4. Variable Selection
2.5. Software and Parameters
3. Results
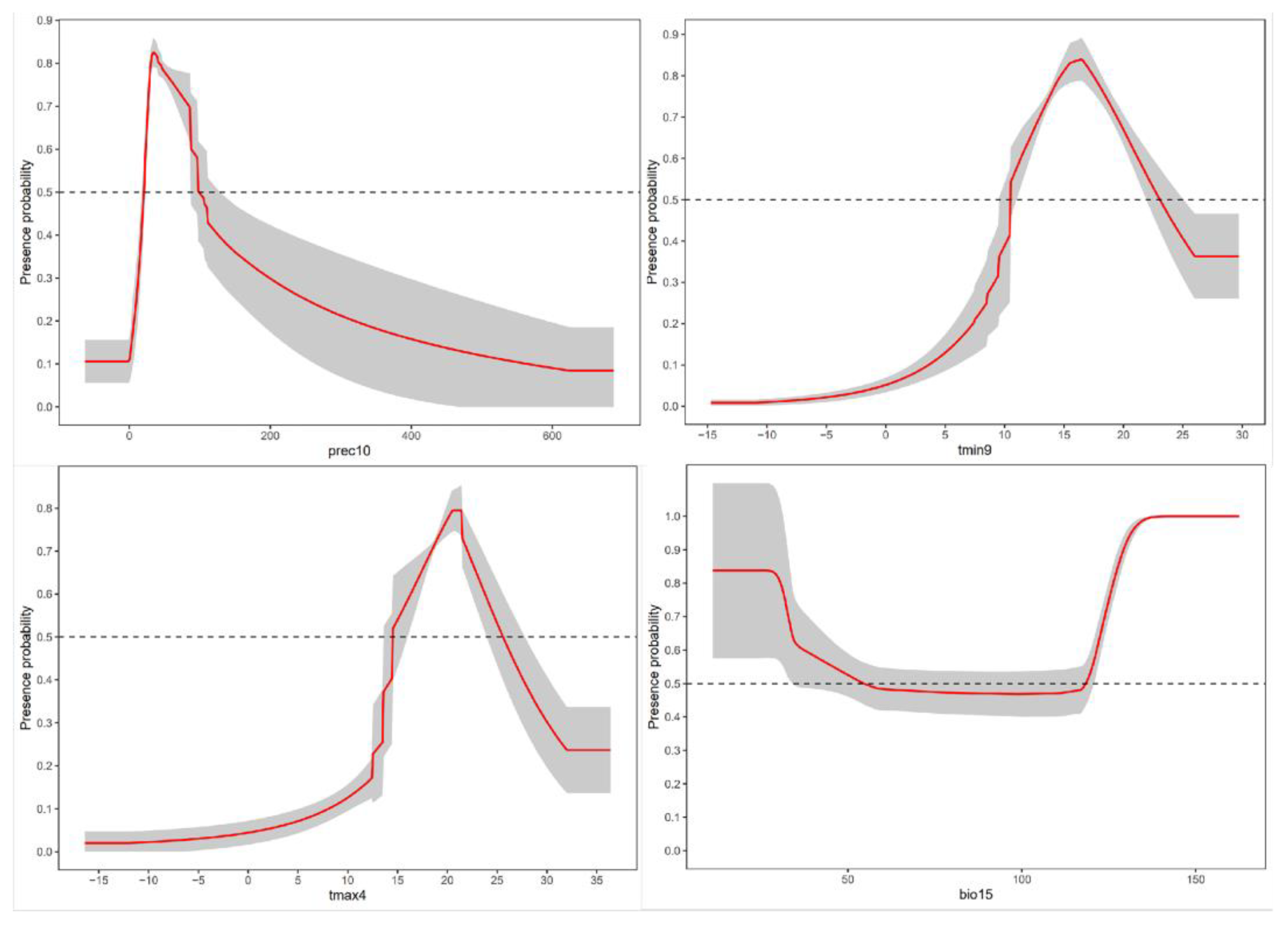
3.1. Key Environmental Variables Affecting the Distribution of Potentially Suitable Areas for A. glabripennis (Asian longhorned beetle, ALB)
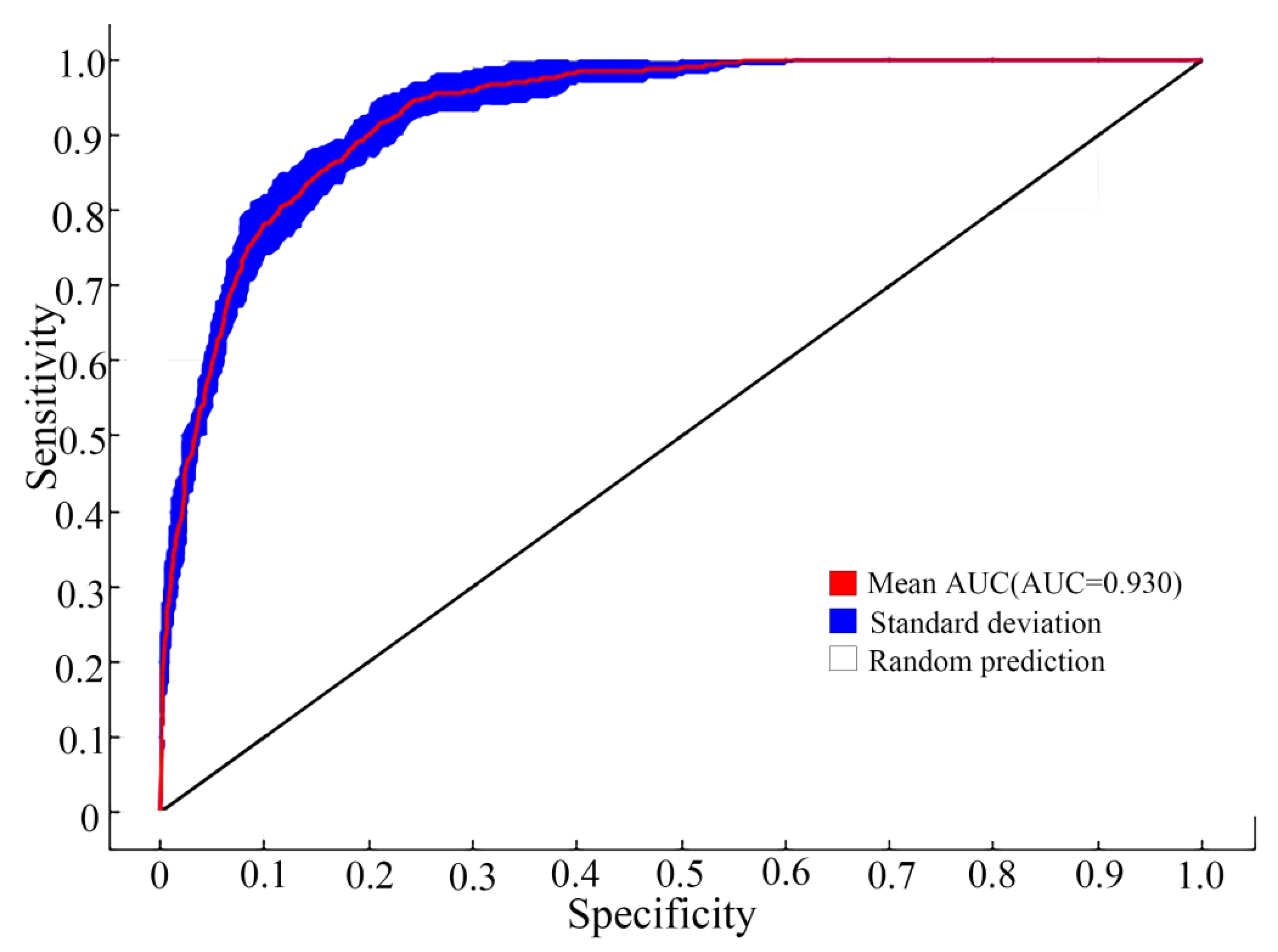
3.2. Evaluation of Model Accuracy
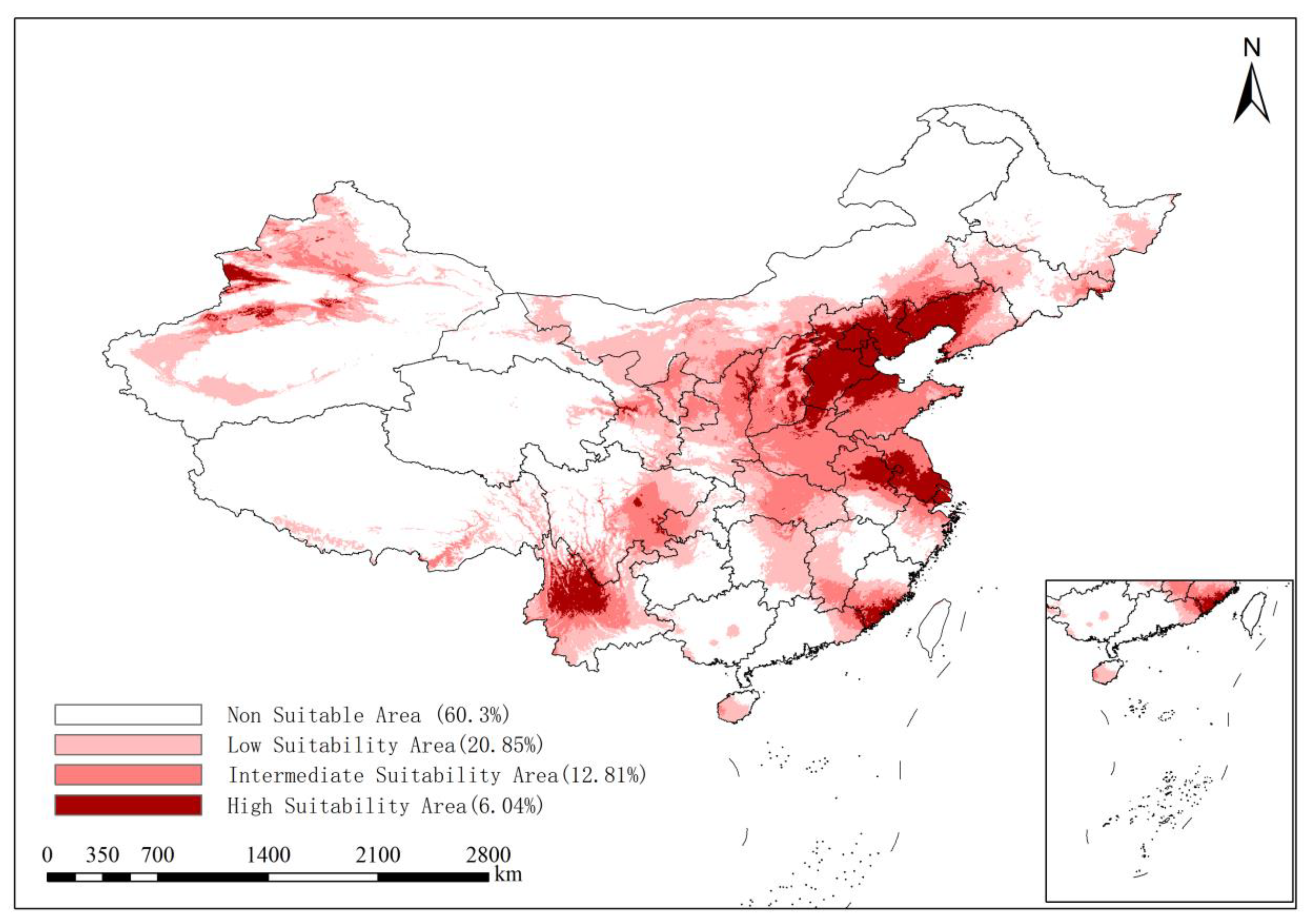
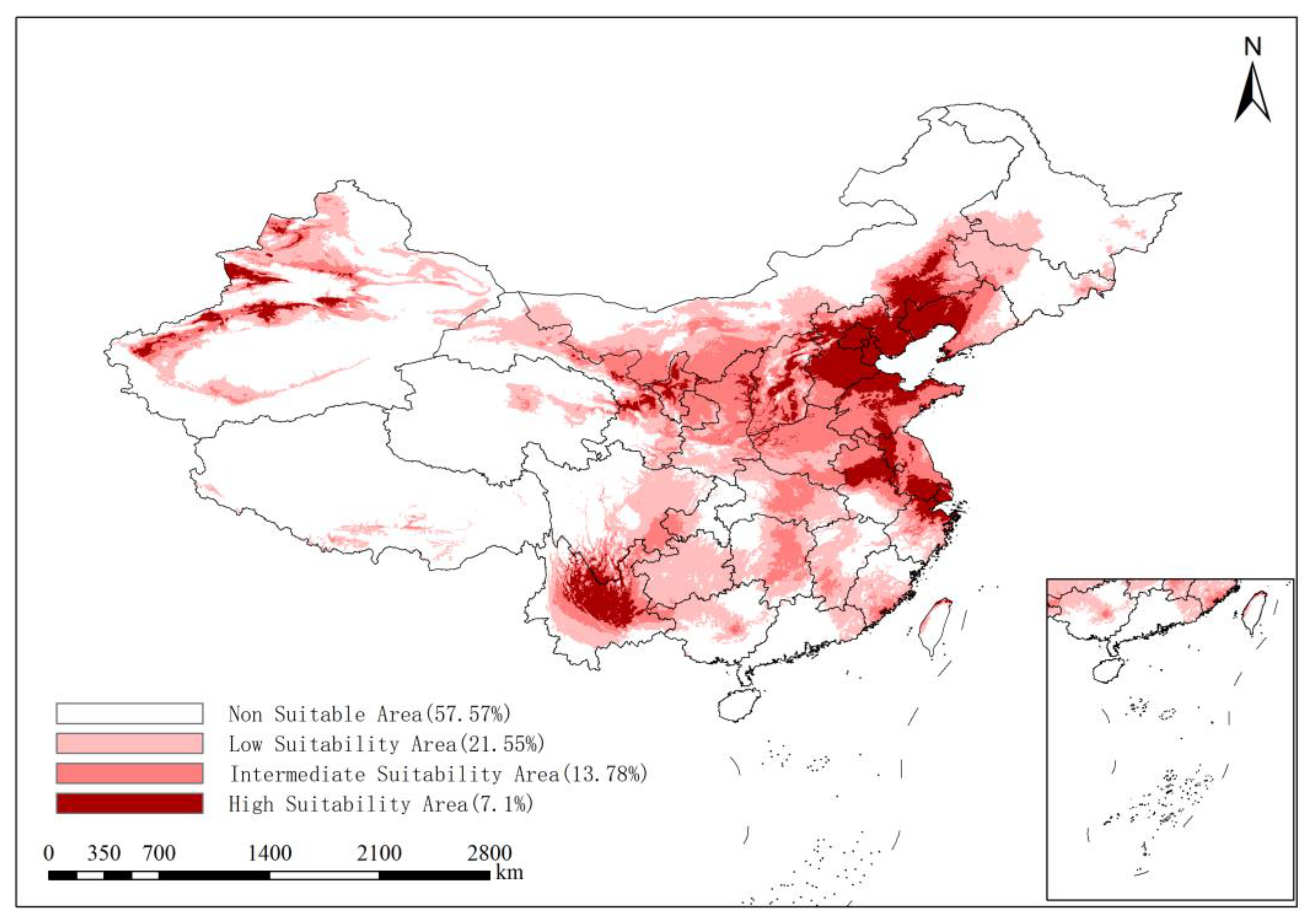
3.3. Prediction of the Distribution of the Suitable Area for ALB in the Present Period
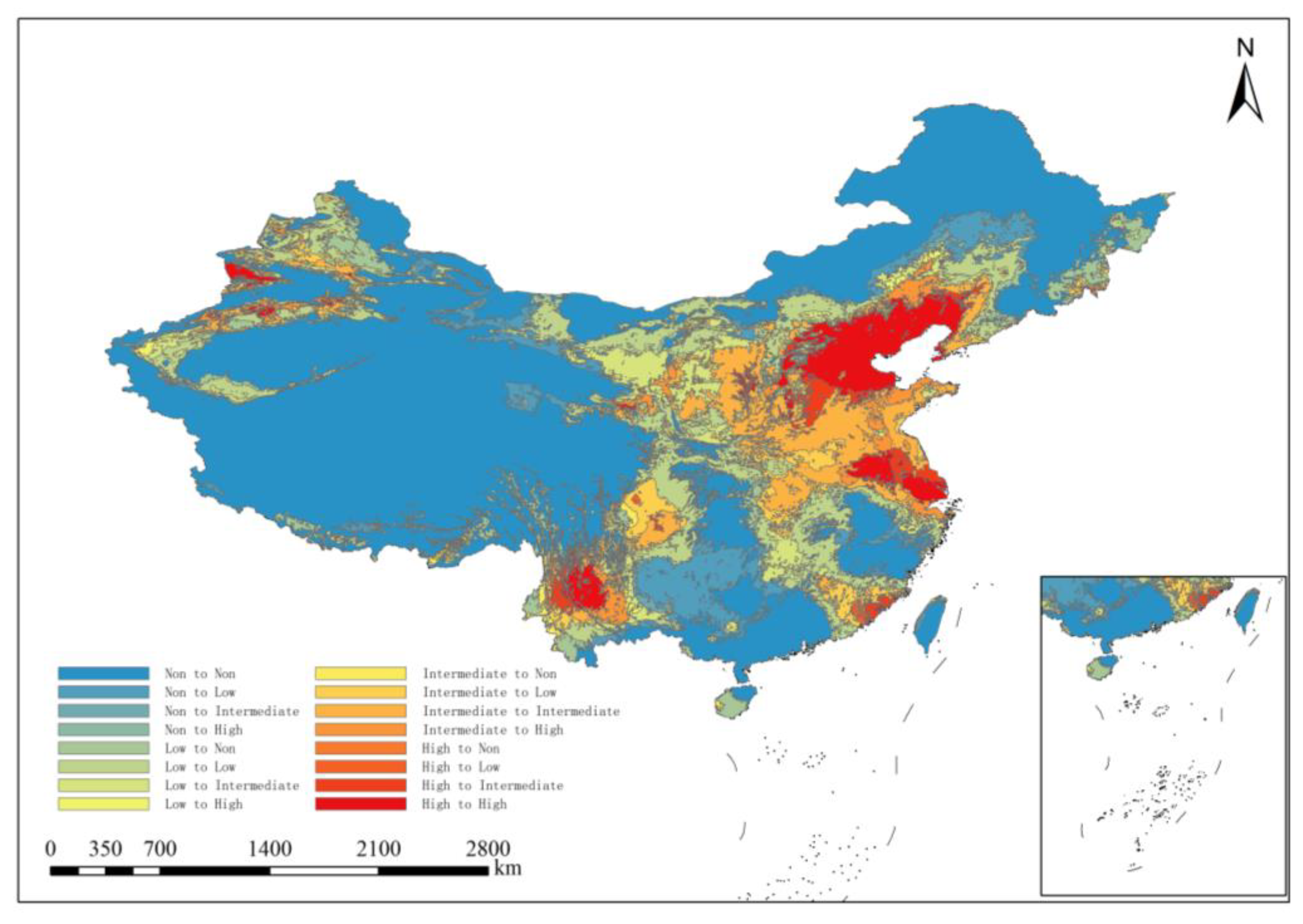
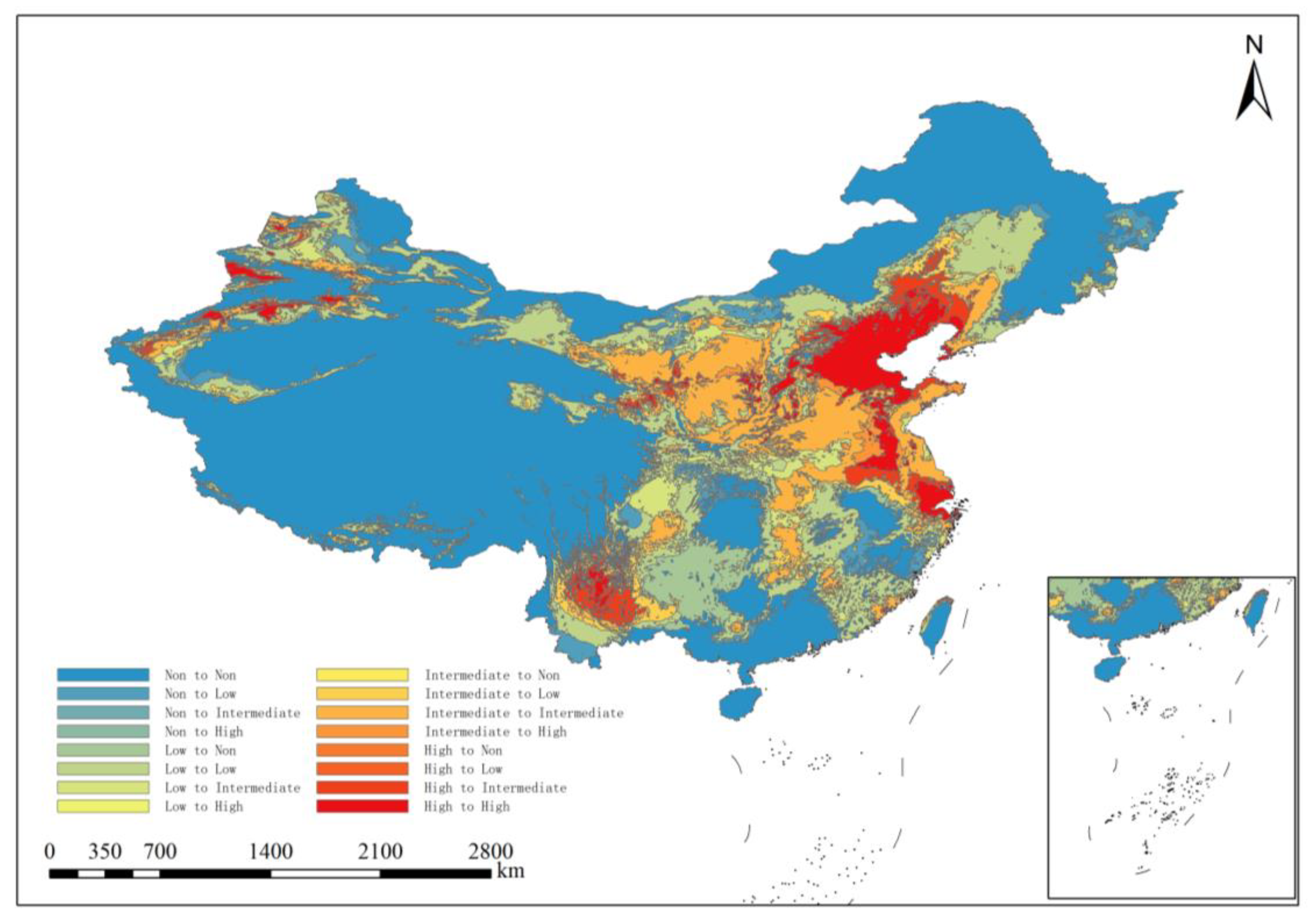
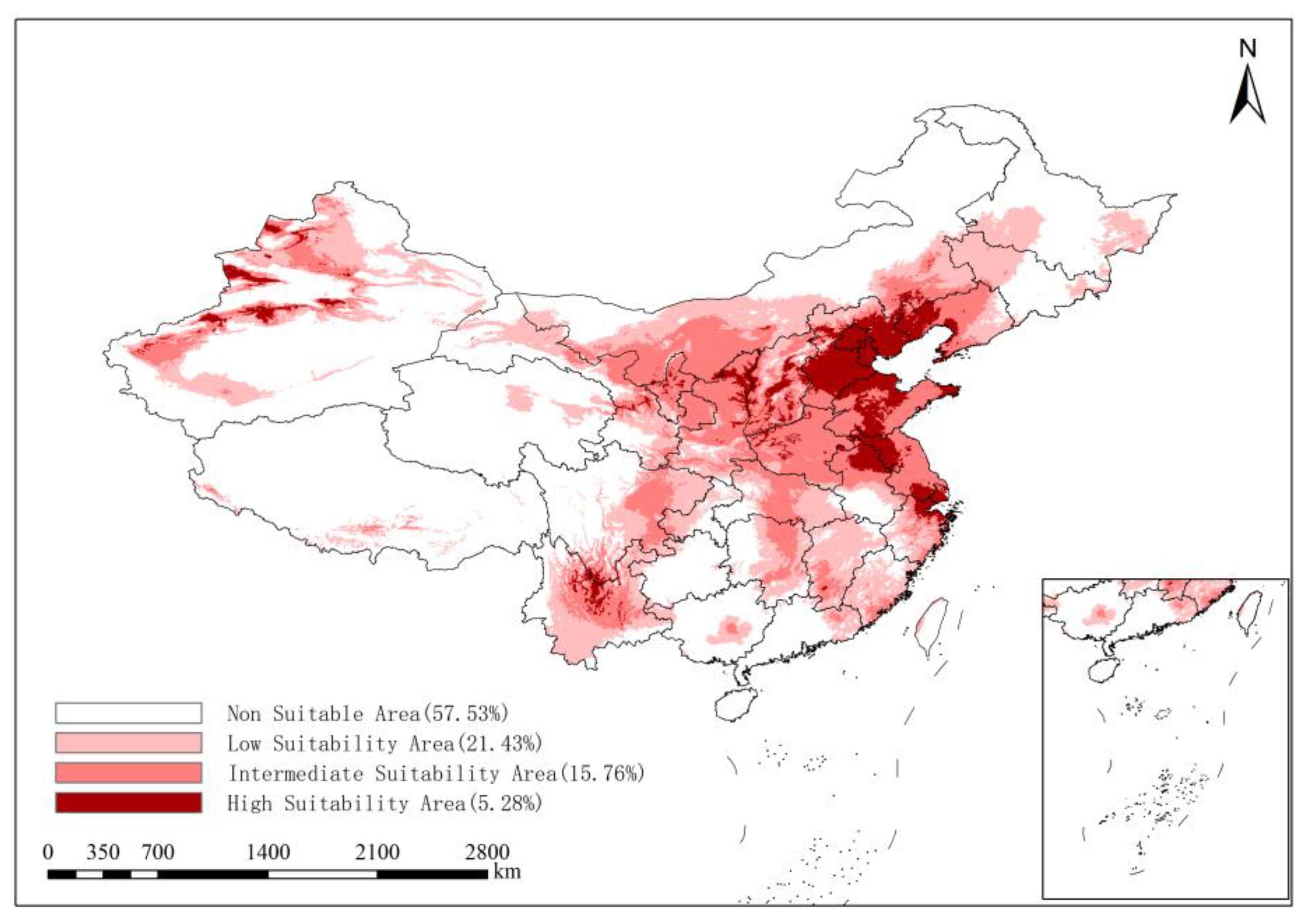
3.4. Future Distribution Changes in ALB in China
4. Discussion
4.1. Dominant Ecological Factors Affecting the Distribution of A. glabripennis (Asian longhorned Beetle, ALB)
4.2. Changes in the Potential Distribution Area of ALB Under Future Climate Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, J.; Angeli, S.; Schuetz, S.; Luo, Y.; Hajek, A.E. Ecology and management of exotic and endemic Asian longhorned beetle Anoplophora glabripennis. Agric. For. Entomol. 2009, 11, 359–375. [Google Scholar] [CrossRef]
- Yasui, H.; Fujiwara-Tsujii, N.; Kugimiya, S.; Shibuya, K.; Mishiro, K.; Uechi, N. Anoplophora glabripennis, an invasive longhorned beetle, has the potential to damage fruit trees in Japan. Sci. Rep. 2024, 14, 12708. [Google Scholar] [CrossRef] [PubMed]
- Haeussermann, I.; Hasselmann, M. Complex European invasion history of Anoplophora glabripennis (Motschulsky): New insights in its population genomic differentiation using genotype-by-sequencing. Sci. Rep. 2024, 14, 4263. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; Tramontini, S.; Antoniou, A.; Gilioli, G.; Krusteva, R.; Sabbatini, P.G.; Rzepecka, D.; Scala, M.; Sánchez, B.; Nougadère, A.V.S. Anoplophora glabripennis—Pest Report to support the ranking of EU candidate priority pests. EFSA Support. Publ. 2025, 22, 9446E. [Google Scholar]
- Dhandapani, R.K.; Gurusamy, D.; Duan, J.; Palli, S.R. RNAi for management of Asian long-horned beetle, Anoplophora glabripennis: Identification of target genes. J. Pest Sci. 2020, 93, 823–832. [Google Scholar] [CrossRef]
- Byeon, D.-h.; Kim, S.-H.; Jung, J.-M.; Jung, S.; Kim, K.-H.; Lee, W.-H. Climate-based ensemble modelling to evaluate the global distribution of Anoplophora glabripennis (Motschulsky). Agric. For. Entomol. 2021, 23, 569–583. [Google Scholar] [CrossRef]
- Zhang, Q.-C.; Wang, J.-G.; Lei, Y.-H. Predicting distribution of the Asian longhorned beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and its natural enemies in China. Insects 2022, 13, 687. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, C.; Luo, Y.; Wang, G.; Dou, Z.; Haq, I.U.; Shang, S.; Cui, M. Current and future control of the wood-boring pest Anoplophora glabripennis. Insect Sci. 2023, 30, 1534–1551. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.J.W.; Bonebrake, T.C.; da Silva, K.D.; Dongmo, M.A.K.; França, F.M.; Gregory, N.; Kitching, R.L.; Ledger, M.J.; Lewis, O.T.; Sharp, A.C.; et al. Causes and consequences of insect decline in tropical forests. Nat. Rev. Biodivers. 2025, 1, 315–331. [Google Scholar] [CrossRef]
- Huang, J.; Lu, X.; Liu, H.; Zong, S. The driving forces of Anoplophora glabripennis have spatial spillover effects. Forests 2021, 12, 1678. [Google Scholar] [CrossRef]
- Zhou, Y.; Ge, X.; Zou, Y.; Guo, S.; Wang, T.; Zong, S. Prediction of the potential global distribution of the Asian longhorned beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) under climate change. Agric. For. Entomol. 2021, 23, 557–568. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, P.; Xie, G.; Wang, W. Impacts of climate change conditions on the potential distribution of Anoplophora glabripennis and its host plants, Salix babylonica and Salix matsudana, in China. Ecol. Evol. 2024, 14, e70692. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Fan, M.; Liu, Y.; Huang, P.; Gaisberger, H.; Li, C.; Zheng, Y.; Lin, F. Habitat suitability modeling of a nearly extinct rosewood species (Dalbergia odorifera) under current, and future climate conditions. J. For. Res. 2025, 36, 58. [Google Scholar] [CrossRef]
- Yong, D.; Xu, D.; Deng, X.; He, Z.; Zhuo, Z. Potential distribution of Anoplophora horsfieldii Hope in China based on MaxEnt and its response to climate change. Insects 2025, 16, 484. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhuo, Z.; Ali, H.; Mureed, S.; Liu, Q.; Yang, X.; Xu, D. Predicting potential habitat distribution of the invasive species Rhynchophorus ferrugineus Olivier in China based on MaxEnt modelling technique and future climate change. Bull. Entomol. Res. 2024, 114, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, D.; Liao, W.; Xu, Y.; Zhuo, Z. Predicting the current and future distributions of Frankliniella occidentalis (Pergande) based on the MaxEnt species distribution model. Insects 2023, 14, 458. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lim, C.; Kang, J.H.; Bae, Y.J. The effect of climate change on indicator wetland insects: Predicting the current and future distribution of two giant water bugs (Hemiptera: Belostomatidae) in South Korea. Insects 2024, 15, 820. [Google Scholar] [CrossRef] [PubMed]
- Pagad, S.; Bisset, S.; Genovesi, P.; Groom, Q.; Hirsch, T.; Jetz, W.; Ranipeta, A.; Schigel, D.; Sica, Y.V.; McGeoch, M.A. Country compendium of the global register of introduced and invasive species. Sci. Data 2022, 9, 391. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Luo, J.; Zhang, R.; Liu, J.; Cao, D.; Su, Z.; Wei, J. Methyl jasmonate enhances the resistance of Populus alba var. pyramidalis against Anoplophora glabripennis (Coleoptera: Cerambycidae). Insects 2025, 16, 153. [Google Scholar] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Marchi, M.; Sinjur, I.; Bozzano, M.; Westergren, M. Evaluating WorldClim version 1 (1961–1990) as the baseline for sustainable use of forest and environmental resources in a changing climate. Sustainability 2019, 11, 3043. [Google Scholar] [CrossRef]
- Akhtar, N.; Alharthi, M.F. Enhancing accuracy in modeling highly multicollinear data using alternative shrinkage parameters for ridge regression methods. Sci. Rep. 2025, 15, 10774. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Zheng, F.; Zhang, W.; Xu, G.; Li, D.; Yang, S.; Jin, G.; Clements, D.R.; Nikkel, E.; Chen, A.; et al. Potential distribution and ecological impacts of Acmella radicans (Jacquin) R.K. Jansen (a new Yunnan invasive species record) in China. BMC Plant Biol. 2024, 24, 494. [Google Scholar] [CrossRef] [PubMed]
- Pulighe, G.; Lupia, F.; Manente, V. Climate-driven invasion risks of Japanese beetle (Popillia japonica Newman) in Europe predicted through species distribution modelling. Agriculture 2025, 15, 684. [Google Scholar] [CrossRef]
- Wu, J.; Wei, X.; Wang, Z.; Peng, Y.; Liu, B.; Zhuo, Z. Mapping the distribution of curculio davidi Fairmaire 1878 under climate change via geographical data and the MaxEnt model (CMIP6). Insects 2024, 15, 583. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yang, J.; Xu, D.; Zhuo, Z. Global distribution prediction of Cyrtotrachelus buqueti guer (Coleoptera: Curculionidae) insights from the optimised MaxEnt model. Insects 2024, 15, 708. [Google Scholar] [CrossRef] [PubMed]
- John, A.; Riat, A.K.; Bhat, K.A.; Ganie, S.A.; endarto, O.; Nugroho, C.; Handoko, H.; Wani, A.K. Adapting to climate extremes: Implications for insect populations and sustainable solutions. J. Nat. Conserv. 2024, 79, 126602. [Google Scholar] [CrossRef]
- Keena, M.A.; Moore, P.M. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) larvae and pupae. Environ. Entomol. 2010, 39, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Fang, G.; Chai, S.; He, C.; Sun, S.; Zhao, G.; Lin, X. Can ecological niche models be used to accurately predict the distribution of invasive insects? A case study of Hyphantria cunea in China. Ecol. Evol. 2024, 14, e11159. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Jiang, Z.; Li, W.; Hou, Q.; Li, L. Changes in extreme temperature over China when global warming stabilized at 1.5 °C and 2.0 °C. Sci. Rep. 2019, 9, 14982. [Google Scholar] [CrossRef] [PubMed]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wu, Y.; Javal, M.; Giguère, I.; Roux, G.; Andres, J.A.; Keena, M.; Shi, J.; Wang, B.; Braswell, E.; et al. Genome-scale phylogeography resolves the native population structure of the Asian longhorned beetle, Anoplophora glabripennis (Motschulsky). Evol. Appl. 2022, 15, 934–953. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment Variables | Description | Environment Variables | Description |
|---|---|---|---|
| Bio1 | mean annual temperature | Bio12 | Average annual precipitation |
| Bio2 | Average temperature difference between day and night | Bio13 | Precipitation in the wettest month |
| Bio3 | Isothermality | Bio14 | Precipitation in the driest month |
| Bio4 | Temperature seasonality | Bio15 | Seasonal variation coefficient of precipitation |
| Bio5 | The highest temperature of the hottest month | Bio16 | Wettest quarterly precipitation |
| Bio6 | The lowest temperature in the coldest month | Bio17 | Precipitation in the driest quarter |
| Bio7 | Annual temperature variation range | Bio18 | Warmest quarterly precipitation |
| Bio8 | The average temperature of the wettest quarter | Bio19 | Precipitation in the coldest quarter |
| Bio9 | The average temperature of the driest quarter | Preci (i = 1, 2, …, 12) | The average precipitation in month i |
| Bio10 | The average temperature of the hottest quarter | tmaxi (i = 1, 2, …, 12) | The average maximum temperature in month i |
| Bio11 | The average temperature of the coldest quarter | tmini (i = 1, 2, …, 12) | The average minimum temperature in the ith month |
| Environment Variables | Description | Percent Contribution % |
|---|---|---|
| Bio15 | Seasonal variation coefficient of precipitation | 17.7 |
| tmin9 | The average minimum temperature in September | 15.4 |
| prec10 | The average precipitation in October | 8.9 |
| prec4 | The average precipitation in April | 5.2 |
| prec1 | The average precipitation in January | 4.4 |
| prec11 | The average precipitation in November | 4.1 |
| tmax4 | The average maximum temperature in April | 3.5 |
| prec6 | The average precipitation in June | 2.7 |
| Bio3 | Isothermality | 2.7 |
| Bio7 | Annual temperature variation range | 2.3 |
| prec9 | The average precipitation in September | 2.1 |
| Tmax6 | The average maximum temperature in June | 1.3 |
| Porportion China | 2021–2040 | |||||
|---|---|---|---|---|---|---|
| Non | Low | Intermediate | High | Total | ||
| Present | Non | 0.92 | 0.08 | 0.01 | 0.00 | 1 |
| Low | 0.33 | 0.52 | 0.14 | 0.01 | 1 | |
| Intermediate | 0.03 | 0.34 | 0.52 | 0.11 | 1 | |
| High | 0.00 | 0.07 | 0.32 | 0.61 | 1 | |
| Total | 0.6 | 0.2 | 0.1 | 0.1 | 1 | |
| Porportion China | 2041–2060 | |||||
|---|---|---|---|---|---|---|
| Non | Low | Intermediate | High | Total | ||
| 2021–2040 | Non | 0.93 | 0.07 | 0.00 | 0.00 | 1 |
| Low | 0.19 | 0.67 | 0.14 | 0.00 | 1 | |
| Intermediate | 0.01 | 0.20 | 0.62 | 0.18 | 1 | |
| High | 0.00 | 0.00 | 0.19 | 0.81 | 1 | |
| Total | 0.6 | 0.2 | 0.1 | 0.1 | 1 | |
| Area Ratio (%) | SSP | Nonsuitability Area | Low Suitability Area | Intermediate Suitability Area | High Suitability Area |
|---|---|---|---|---|---|
| Time (Year) | |||||
| 1970–2000 | 60.30 | 20.85 | 12.81 | 6.04 | |
| 2021–2040 | SSP245 | 57.57 | 21.55 | 13.78 | 7.10 |
| 2041–2060 | SSP245 | 57.53 | 21.43 | 15.76 | 5.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, K.; Zhou, M.; Hu, H.; Dong, N.; Tang, C. Prediction of the Potentially Suitable Area for Anoplophora glabripennis (Coleoptera: Cerambycidae) in China Based on MaxEnt. Forests 2025, 16, 1239. https://doi.org/10.3390/f16081239
Tan K, Zhou M, Hu H, Dong N, Tang C. Prediction of the Potentially Suitable Area for Anoplophora glabripennis (Coleoptera: Cerambycidae) in China Based on MaxEnt. Forests. 2025; 16(8):1239. https://doi.org/10.3390/f16081239
Chicago/Turabian StyleTan, Kaiwen, Mingwang Zhou, Hongjiang Hu, Ning Dong, and Cheng Tang. 2025. "Prediction of the Potentially Suitable Area for Anoplophora glabripennis (Coleoptera: Cerambycidae) in China Based on MaxEnt" Forests 16, no. 8: 1239. https://doi.org/10.3390/f16081239
APA StyleTan, K., Zhou, M., Hu, H., Dong, N., & Tang, C. (2025). Prediction of the Potentially Suitable Area for Anoplophora glabripennis (Coleoptera: Cerambycidae) in China Based on MaxEnt. Forests, 16(8), 1239. https://doi.org/10.3390/f16081239