
Spatial Patterning and Growth of Naturally Regenerated Eastern White Pine in a Northern Hardwood Silviculture Experiment

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
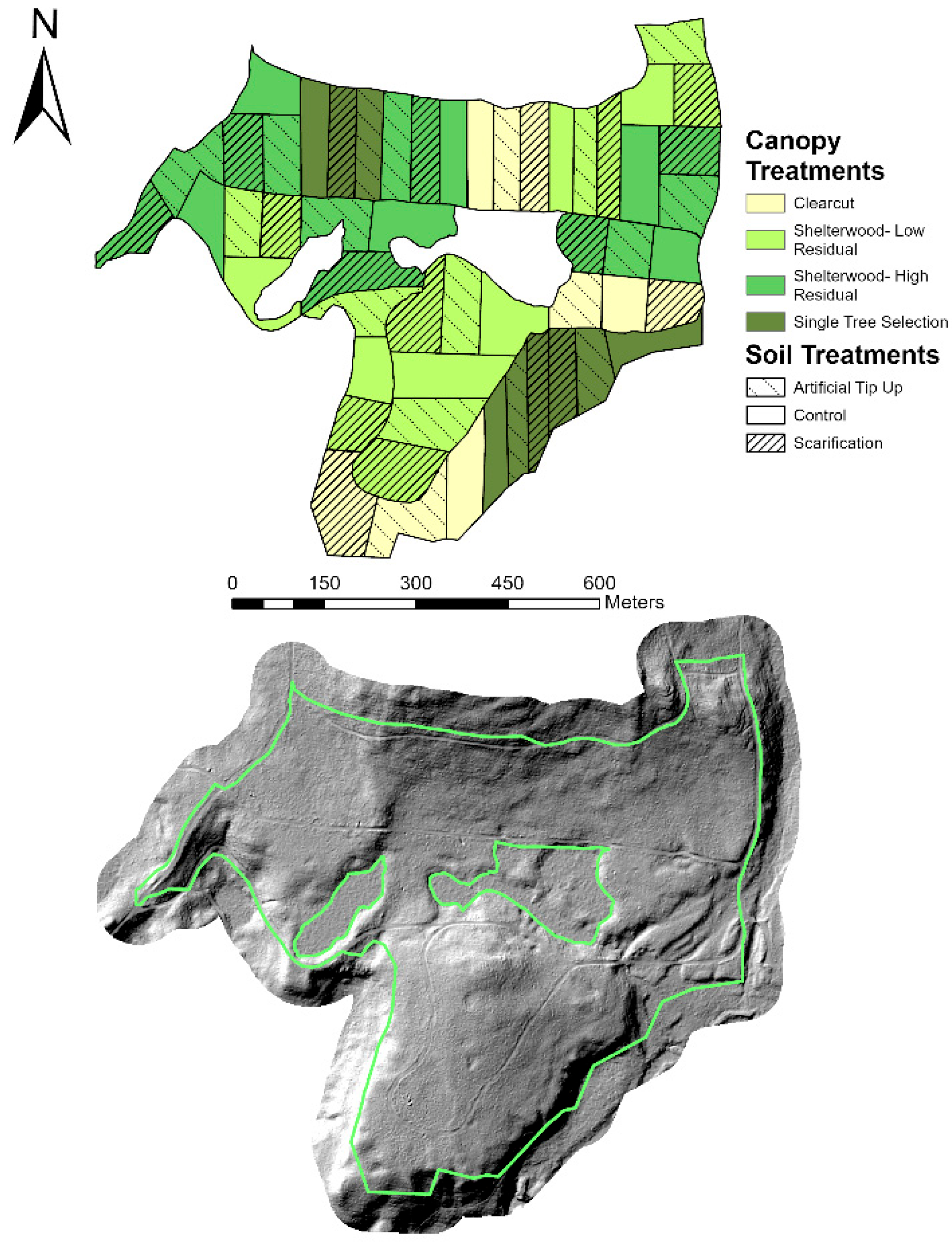
2.2. The NH-SEED Experiment
2.3. Field Data Collection
2.4. Analysis Methods
3. Results
3.1. Density
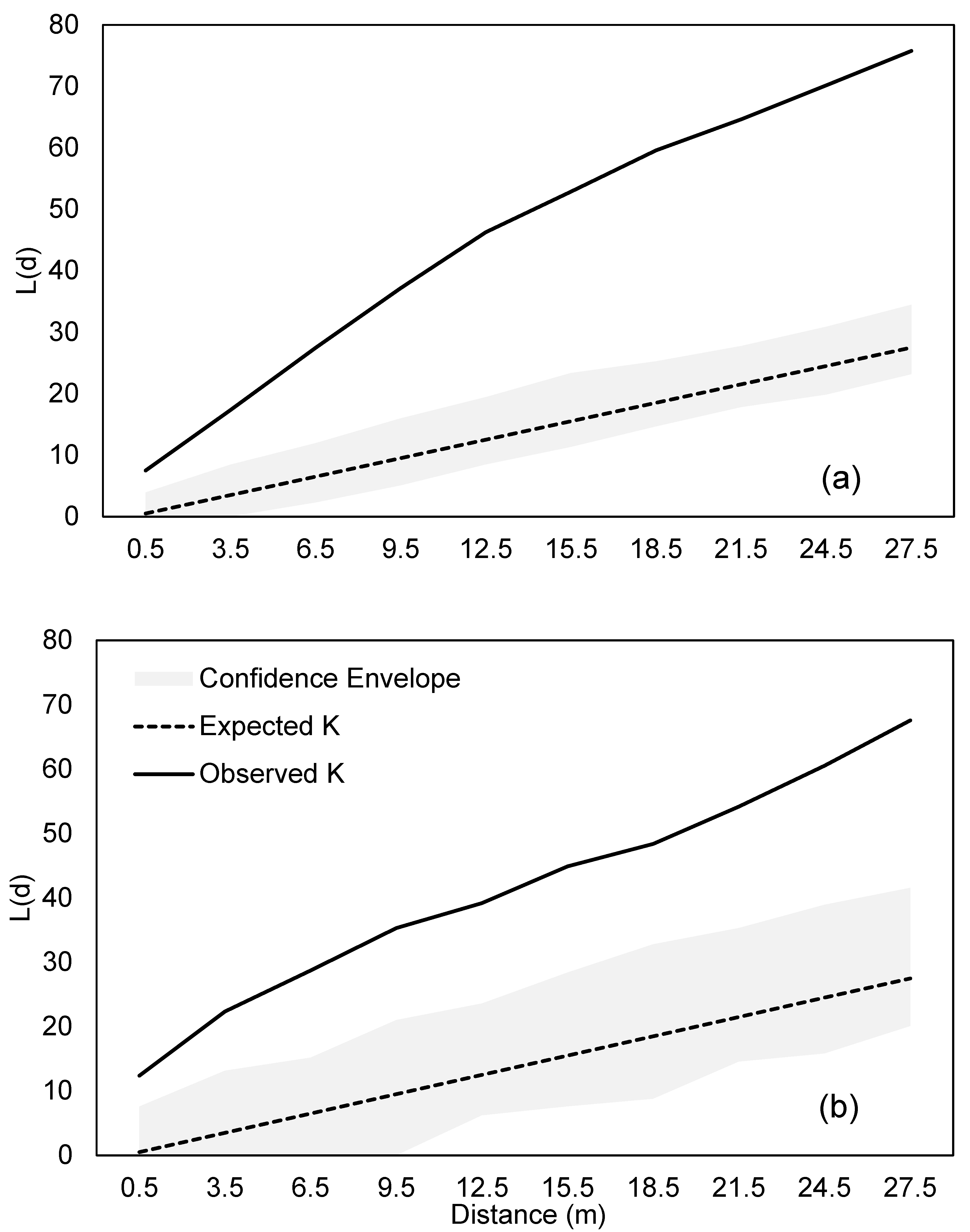
3.2. Spatial Pattern
3.3. Internode Growth
3.4. Damaging Agents
4. Discussion
5. Study Limitations
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palik, B.J.; Pregitzer, K.S. White pine seed-tree legacies in an aspen landscape: Influences on post-disturbance white pine population structure. For. Ecol. Manag. 1994, 67, 191–201. [Google Scholar] [CrossRef]
- Lutz, J.A.; Larson, A.J.; Swanson, M.E.; Freund, J.A. Ecological importance of large-diameter trees in a temperate mixed-conifer forest. PLoS ONE 2012, 7, e36131. [Google Scholar] [CrossRef] [PubMed]
- Fahey, R.T.; Lorimer, C.G. Restoring a midtolerant pine species as a component of late-successional forests: Results of gap-based planting trials. For. Ecol. Manag. 2013, 292, 139–149. [Google Scholar] [CrossRef]
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; Gonzalez-Olabarria, J.R.; Koricheva, J.; Meurisse, N.; Brockerhoff, E.G. Tree diversity drives forest stand resistance to natural disturbances. Curr. For. Rep. 2017, 3, 223–243. [Google Scholar] [CrossRef]
- Fahey, R.T.; Lorimer, C.G.; Mladenoff, D.J. Habitat heterogeneity and life-history traits influence presettlement distributions of early-successional tree species in a late-successional, hemlock-hardwood landscape. Landsc. Ecol. 2012, 27, 999–1013. [Google Scholar] [CrossRef]
- Walker, B.; Kinzig, A.; Langridge, J. Plant attribute diversity, resilience, and ecosystem function: The nature and significance of dominant and minor Species. Ecosystems 1999, 2, 95–113. [Google Scholar] [CrossRef]
- Espelta, J.M.; Cruz-Alonso, V.; Alfaro-Sánchez, R.; Hampe, A.; Messier, C.; Pino, J. Functional diversity enhances tree growth and reduces herbivory damage in secondary broadleaf forests, but does not influence resilience to drought. J. Appl. Ecol. 2020, 57, 2362–2372. [Google Scholar] [CrossRef]
- Hermy, M.; Verheyen, K. Legacies of the past in the present-day forest biodiversity: A review of past land-use effects on forest plant species composition and diversity. Ecol. Res. 2007, 22, 361–371. [Google Scholar] [CrossRef]
- Tripler, C.E.; Canham, C.D.; Inouye, R.S.; Schnurr, J.L. Competitive hierarchies of temperate tree species: Interactions between resource availability and white-tailed deer. Écoscience 2005, 12, 494–505. [Google Scholar] [CrossRef]
- Weyenberg, S.A.; Frelich, L.E.; Reich, P.B. Logging versus fire: How does disturbance type influence the abundance of Pinus strobus regeneration? Silva Fenn. 2004, 38, 179–194. [Google Scholar] [CrossRef]
- Cornett, M.W.; Reich, P.B.; Puettmann, K.J. Canopy feedbacks and microtopography regulate conifer seedling distribution in two Minnesota conifer-deciduous forests. Écoscience 1997, 4, 353–364. [Google Scholar] [CrossRef]
- Webster, C.R.; Dickinson, Y.L.; Burton, J.I.; Frelich, L.E.; Jenkins, M.A.; Kern, C.C.; Raymond, P.; Saunders, M.R.; Walters, M.B.; Willis, J.L. Promoting and maintaining diversity in contemporary hardwood forests: Confronting contemporary drivers of change and the loss of ecological memory. For. Ecol. Manag. 2018, 421, 98–108. [Google Scholar] [CrossRef]
- Gibson, A.L.; Espeland, E.K.; Wagner, V.; Nelson, C.R. Can local adaptation research in plants inform selection of native plant materials? An analysis of experimental methodologies. Evol. Appl. 2016, 9, 1219–1228. [Google Scholar] [CrossRef]
- Granger, J.J.; Buckley, D.S. Performance of white oak (Quercus alba) and three pine species in novel multi-cropped plantations in Tennessee, USA. For. Ecol. Manag. 2021, 489, 119060. [Google Scholar] [CrossRef]
- Clabo, D.C.; Clatterbuck, W.K. Site preparation techniques for the establishment of mixed pine-hardwood stands: 22-year results. For. Sci. 2015, 61, 790–799. [Google Scholar] [CrossRef]
- Whitney, G.G. An ecological history of the Great Lakes forest of Michigan. J. Ecol. 1987, 75, 667–684. [Google Scholar] [CrossRef]
- Pastor, J.; Broschart, M. The spatial pattern of a northern conifer-hardwood landscape. Landsc. Ecol. 1990, 4, 55–68. [Google Scholar] [CrossRef]
- Goebel, P.C.; Palik, B.J.; Pregitzer, K.S. Structure and composition of riparian forests in an old-growth northern hardwood–hemlock watershed. For. Ecol. Manag. 2012, 280, 52–61. [Google Scholar] [CrossRef]
- Neithercut, M.E. The White Pine Industry and the Transformation of Nineteenth-Century Michigan. Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 1984. Available online: https://open.library.ubc.ca/collections/ubctheses/831/items/1.0096560 (accessed on 7 March 2024).
- Uprety, Y.; Asselin, H.; Bergeron, Y. Preserving ecosystem services on indigenous territory through restoration and management of a cultural keystone species. Forests 2017, 8, 194. [Google Scholar] [CrossRef]
- Naylor, B.J. Managing wildlife habitat in red pine and white pine forests of central Ontario. For. Chron. 1994, 70, 411–419. [Google Scholar] [CrossRef]
- Costanza, K.K.L.; Whitney, T.D.; McIntire, C.D.; Livingston, W.H.; Gandhi, K.J.K. A synthesis of emerging health issues of white pine (Pinus strobus) in North America. For. Ecol. Manag. 2018, 423, 3–17. [Google Scholar] [CrossRef]
- Zenner, E.K.; Peck, J.E. Maintaining a pine legacy in Itasca State Park. Nat. Areas J. 2009, 29, 157–166. [Google Scholar] [CrossRef]
- Willis, J.L.; Walters, M.B.; Farinosi, E. Local seed source availability limits young seedling populations for some species more than other factors in northern hardwood forests. For. Sci. 2016, 62, 440–448. [Google Scholar] [CrossRef]
- Wendel, G.W.; Smith, H.C. Eastern White Pine. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; Volume 1. Conifers. Agric. Handb. 654; United States Department of Agriculture, Forest Service: Washington, DC, USA, 1990; pp. 476–488. [Google Scholar]
- Hupperts, S.F.; Webster, C.R.; Froese, R.E.; Dickinson, Y.L. Seedling and sapling recruitment following novel silvicultural treatments in Great Lakes northern hardwoods. For. Ecol. Manag. 2020, 462, 117983. [Google Scholar] [CrossRef]
- Hupperts, S.F.; Webster, C.R.; Froese, R.E.; Lilleskov, E.A.; Marcarelli, A.M.; Dickinson, Y.L. Increasing ground-layer plant taxonomic diversity masks declining phylogenetic diversity along a silvicultural disturbance gradient. Can. J. For. Res. 2020, 50, 1259–1267. [Google Scholar] [CrossRef]
- Bartlick, C.I.; Burton, J.I.; Webster, C.R.; Froese, R.E.; Dickinson, Y.L. An experimental approach to identify drivers of tree regeneration diversity, composition, and heterogeneity in northern hardwood forests. For. Ecol. Manag. 2023, 546, 121320. [Google Scholar] [CrossRef]
- Webster, C.R.; Dickinson, Y.L.; Froese, R.E. Introducing the NH-SEED: A holistic experiment to revisit classical and test novel silvicultural systems in a changing world. In Proceedings of the First Biennial Northern Hardwood Conference 2021: Bridging Science and Management for the Future, Online, 15–16 June 2021; Gen. Tech. Rep. NRS-P-211. USDA Forest Service: Madison, WI, USA, 2023; pp. 131–135. [Google Scholar] [CrossRef]
- Soil Survey Staff, Natural Resources Conservation Service, United States Department of Agriculture. Official Soil Series Descriptions. Available online: https://websoilsurvey.nrcs.usda.gov/app/ (accessed on 7 March 2024).
- Burger, T.L.; Kotar, J. A Guide to Forest Communities and Habitat Types of Michigan; University of Wisconsin: Madison, WI, USA, 2003. [Google Scholar]
- National Centers for Environmental Information, National Oceanic and Atmospheric Administration. U.S. Climate Normals Quick Access, 1981–2010. Available online: https://www.ncei.noaa.gov/access/us-climate-normals/#dataset=normals-monthly&timeframe=81&location=MI&station=USC00200089 (accessed on 20 April 2023).
- Midwestern Regional Climate Center. (1981–2010) Snow and Precipitation Summaries for Station USC00203744—HERMAN, MI. Available online: https://mrcc.purdue.edu (accessed on 20 April 2023).
- Graham, S.A. Climax forests of the Upper Peninsula of Michigan. Ecology 1941, 22, 355–362. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Butaye, J.; Hermy, M. Influence of environmental and spatial variables on regional distribution of forest plant species in a fragmented and changing landscape. Ecography 2003, 26, 768–776. [Google Scholar] [CrossRef]
- Reed, D.D.; Holmes, M.J.; Johnson, J.A. A 22-year study of stand development and financial return in northern hardwoods. North. J. Appl. For. 1986, 3, 35–38. [Google Scholar] [CrossRef]
- Erickson, M.D.; Reed, D.D.; Mroz, G.D. Stand development and economic analysis of alternative cutting methods in northern hardwoods: 32-year results. North. J. Appl. For. 1990, 7, 153–158. [Google Scholar] [CrossRef]
- Schwartz, J.W.; Nagel, L.M.; Webster, C.R. Effects of uneven-aged management on diameter distribution and species composition of northern hardwoods in Upper Michigan. For. Ecol. Manag. 2005, 211, 356–370. [Google Scholar] [CrossRef]
- Bartlick, C.I.; Burton, J.I.; Webster, C.R.; Froese, R.E.; Hupperts, S.F.; Dickinson, Y.L. Artificial tip-up mounds influence tree seedling composition in a managed northern hardwood forest. Can. J. For. Res. 2023, 53, 893–904. [Google Scholar] [CrossRef]
- Harlow, W.M.; Harrar, E.S.; Hardin, J.W.; White, F.M. Textbook of Dendrology, Seventh Edition; McGraw-Hill: New York, NY, USA, 1991. [Google Scholar]
- Ripley, B.D. The second-order analysis of stationary processes. J. Appl. Prob. 1976, 13, 255–266. [Google Scholar] [CrossRef]
- Besag, J. Contribution to the discussion on Dr. Ripley’s paper. J. R. Stat. Soc. B 1977, 39, 192–212. [Google Scholar] [CrossRef]
- Ripley, B.D. Modeling Spatial Patterns. J. R. Stat. Soc. B 1977, 39, 172–192. [Google Scholar] [CrossRef]
- Esri Inc. Multi-Distance Spatial Cluster Analysis. Available online: https://pro.arcgis.com/en/pro-app/latest/tool-reference/spatial-statistics/multi-distance-spatial-cluster-analysis.htm (accessed on 18 June 2025).
- Esri Inc. How Multi-Distance Spatial Cluster Analysis (Ripley’s K-Function) Works. Available online: https://pro.arcgis.com/en/pro-app/latest/tool-reference/spatial-statistics/h-how-multi-distance-spatial-cluster-analysis-ripl.htm (accessed on 18 June 2025).
- Philip, G.M.; Watson, D.F. A precise method for determining contoured surfaces. Aust. Pet. Explor. Assoc. J. 1982, 22, 205–212. [Google Scholar] [CrossRef]
- Watson, D.F.; Philip, G.M. A refinement of inverse distance weighted interpolation. Geoprocessing 1985, 2, 315–327. [Google Scholar]
- Silverman, B.W. Density Estimation for Statistics and Data Analysis. Chapman and Hall: New York, NY, USA, 1986. [Google Scholar] [CrossRef]
- Andersen, M. Mechanistic models for the seed shadows of wind-dispersed plants. Am. Nat. 1991, 137, 476–497. [Google Scholar] [CrossRef]
- Horn, H.S.; Nathan, R.; Kaplan, S.R. Long-distance dispersal of tree seeds by wind. Ecol. Res. 2001, 16, 877–885. [Google Scholar] [CrossRef]
- Domke, G.M.; Caspersen, J.P.; Jones, T.A. Light attenuation following selection harvesting in northern hardwood forests. For. Ecol. Manag. 2007, 239, 182–190. [Google Scholar] [CrossRef]
- Roberts, D.; Ciuti, S.; Barber, Q.E.; Willier, C.; Nielsen, S.E. Accelerated seed dispersal along linear disturbances in the Canadian oil sands region. Sci. Rep. 2018, 8, 4828. [Google Scholar] [CrossRef]
- Frothingham, E.H. United States Department of Agriculture Agricultural Bulletin; 13; United States Department of Agriculture: Washington, DC, USA, 1914.
- Lancaster, K.F.; Leak, W.B. A Silviculture Guide for White Pine in the Northeast; Gen. Tech. Rep. NE-41; United States Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Broomall, PA, USA, 1978; p. 13.
- Wilson, R.W.; McQuilkin, W.E. Silvical Characteristics of Eastern White Pine (Pinus strobus); Res. Paper NE-13; United States Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Broomall, PA, USA, 1963; p. 29.
- Wetzel, S.; Burgess, D. Understorey environment and vegetation response after partial cutting and site preparation in Pinus strobus L. stands. For. Ecol. Manag. 2001, 151, 43–59. [Google Scholar] [CrossRef]
- Burgess, D.; Wetzel, S. Recruitment and early growth of eastern white pine (Pinus strobus) regeneration after partial cutting and site preparation. Forestry 2002, 75, 419–423. [Google Scholar] [CrossRef]
- Raymond, P.; Munson, A.D.; Ruel, J.C.; Nolet, P. Group and single-tree selection cutting in mixed tolerant hardwood–white pine stands: Early establishment dynamics of white pine and associated species. For. Chron. 2003, 79, 1093–1106. [Google Scholar] [CrossRef]
- Bleher, B.; Oberrath, R.; Böhning-Gaese, K. Seed dispersal, braeding system, tree density and the spatial pattern of trees—A simulation approach. Basic Appl. Ecol. 2002, 3, 115–123. [Google Scholar] [CrossRef]
- Fahey, R.T.; Lorimer, C.G. Habitat associations and 150 years of compositional change in white pine-hemlock-hardwood forests based on resurvey of public land survey corners. J. Torrey Bot. Soc. 2014, 141, 277–293. [Google Scholar] [CrossRef]
- Saunders, M.R.; Puettmann, K.J. Effects of overstory and understory competition and simulated herbivory on growth and survival of white pine seedlings. Can. J. For. Res. 1999, 29, 536–546. [Google Scholar] [CrossRef]
- Krueger, J.A.; Puettmann, K.J. Growth and injury patterns of eastern white pine (Pinus strobus L.) seedlings as affected by hardwood overstory density and weeding treatments. North. J. Appl. For. 2004, 21, 61–68. [Google Scholar] [CrossRef]
- Bevilacqua, E.; Puttock, D.; Blake, T.J.; Burgess, D. Long-term differential stem growth responses in mature eastern white pine following release from competition. Can. J. For. Res. 2005, 35, 511–520. [Google Scholar] [CrossRef]
- Garret, P. Resistance of eastern white pine (Pinus strobus L.) provenances to the white-pine weevil (Pissodes strobi Peck). Silvae Genet. 1972, 21, 119–121. [Google Scholar]
- Wilkinson, R.C. Leader and growth characteristics of eastern white pine associated with white pine weevil attack susceptibility. Can. J. For. Res. 1983, 13, 78–84. [Google Scholar] [CrossRef]
- Stiell, W.M.; Berry, A.B. Limiting white pine weevil attacks by side shade. For. Chron. 1985, 61, 5–9. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Density ᵻ (stems ha−1) | Height (m) | DBH ᵻ (cm) | Number of Whorls † | Internode Growth ¶ (m yr−1) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |||||||||||||
| Advance Regeneration (n = 156) | 3.84 | ± | 0.91 | 2.31 | ± | 0.12 | 3.56 | ± | 0.27 | 13.30 | ± | 0.39 | 0.21 | ± | 0.01 | ||
| Clearcut (n = 48) | 7.41 | ± | 2.84 | 2.75 | ± | 0.23 | 4.24 | ± | 0.47 | 13.40 | ± | 0.63 | 0.25 | ± | 0.02 | ||
| Control * (n = 39) | 9.26 | ± | 3.87 | 2.76 | ± | 0.27 | 4.26 | ± | 0.57 | 13.62 | ± | 0.76 | 0.24 | ± | 0.02 | ||
| Scarification (n = 9) | 3.72 | ± | 2.45 | 2.72 | ± | 0.31 | 4.14 | ± | 0.67 | 12.44 | ± | 0.63 | 0.28 | ± | 0.03 | ||
| Shelterwood―High Residual (n = 23) | 1.54 | ± | 0.40 | 2.33 | ± | 0.38 | 3.51 | ± | 0.73 | 13.43 | ± | 1.11 | 0.19 | ± | 0.03 | ||
| Control (n = 14) | 1.42 | ± | 0.42 | 2.15 | ± | 0.31 | 2.98 | ± | 0.36 | 12.36 | ± | 1.18 | 0.21 | ± | 0.04 | ||
| Scarification (n = 9) | 1.76 | ± | 0.84 | 2.59 | ± | 0.83 | 4.48 | ± | 1.86 | 15.11 | ± | 2.04 | 0.17 | ± | 0.04 | ||
| Shelterwood―Low Residual (n = 43) | 3.03 | ± | 0.90 | 2.38 | ± | 0.21 | 3.35 | ± | 0.51 | 12.72 | ± | 0.76 | 0.25 | ± | 0.02 | ||
| Control (n = 36) | 3.81 | ± | 1.25 | 2.34 | ± | 0.22 | 3.10 | ± | 0.53 | 12.69 | ± | 0.74 | 0.25 | ± | 0.02 | ||
| Scarification (n = 7) | 1.47 | ± | 0.66 | 2.56 | ± | 0.65 | 5.60 | ± | 1.26 | 12.86 | ± | 2.74 | 0.26 | ± | 0.05 | ||
| Single-Tree Selection (n = 42) | 6.52 | ± | 3.80 | 1.71 | ± | 0.19 | 2.53 | ± | 0.43 | 13.71 | ± | 0.75 | 0.15 | ± | 0.02 | ||
| Control (n = 34) | 7.89 | ± | 5.46 | 1.75 | ± | 0.23 | 2.86 | ± | 0.51 | 14.12 | ± | 0.87 | 0.15 | ± | 0.02 | ||
| Scarification (n = 8) | 3.78 | ± | 2.54 | 1.57 | ± | 0.24 | 1.35 | ± | 0.10 | 12.00 | ± | 1.09 | 0.18 | ± | 0.02 | ||
| New Recruits (n = 81) | 1.89 | ± | 0.46 | 0.63 | ± | 0.04 | 0.97 | ± | 0.21 | 4.46 | ± | 0.14 | 0.15 | ± | 0.01 | ||
| Clearcut (n = 19) | 2.76 | ± | 0.91 | 0.78 | ± | 0.09 | 0.75 | ± | 0.18 | 4.58 | ± | 0.23 | 0.16 | ± | 0.01 | ||
| Control (n = 17) | 3.74 | ± | 1.14 | 0.72 | ± | 0.07 | 0.50 | ± | 0.00 | 4.47 | ± | 0.24 | 0.16 | ± | 0.01 | ||
| Scarification (n = 2) | 0.80 | ± | 0.66 | 1.23 | ± | 0.45 | 1.00 | ± | 0.00 | 5.50 | ± | 0.35 | 0.21 | ± | 0.07 | ||
| Shelterwood―High Residual (n = 28) | 1.75 | ± | 1.06 | 0.55 | ± | 0.04 | - | 4.43 | ± | 0.23 | 0.13 | ± | 0.01 | ||||
| Control (n = 8) | 0.88 | ± | 0.39 | 0.54 | ± | 0.04 | - | 5.00 | ± | 0.31 | 0.11 | ± | 0.01 | ||||
| Scarification (n = 20) | 3.48 | ± | 2.95 | 0.56 | ± | 0.05 | - | 4.20 | ± | 0.28 | 0.15 | ± | 0.02 | ||||
| Shelterwood―Low Residual (n = 21) | 1.53 | ± | 0.56 | 0.66 | ± | 0.09 | 1.40 | ± | 0.00 | 4.19 | ± | 0.30 | 0.16 | ± | 0.01 | ||
| Control (n = 16) | 1.75 | ± | 0.81 | 0.64 | ± | 0.11 | 1.40 | ± | 0.00 | 4.25 | ± | 0.31 | 0.15 | ± | 0.02 | ||
| Scarification (n = 5) | 1.08 | ± | 0.32 | 0.73 | ± | 0.18 | - | 4.00 | ± | 0.75 | 0.17 | ± | 0.02 | ||||
| Single-tree Selection (n = 13) | 2.04 | ± | 0.93 | 0.54 | ± | 0.06 | - | 4.77 | ± | 0.33 | 0.12 | ± | 0.01 | ||||
| Control (n = 7) | 1.65 | ± | 1.03 | 0.57 | ± | 0.09 | - | 5.57 | ± | 0.19 | 0.10 | ± | 0.02 | ||||
| Scarification (n = 6) | 2.83 | ± | 1.78 | 0.50 | ± | 0.04 | - | 3.83 | ± | 0.44 | 0.15 | ± | 0.02 | ||||
| Treatment | Nearest Neighbor (m) | Nearest Seed Tree (m) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Mean ± SE | Max | Min | Mean ± SE | Max | |||||||
| Advance Regeneration (n = 156) | 0.1 | 9.8 | ± | 0.9 | 60.8 | 2.4 | 61.6 | ± | 3.5 | 199.4 | ||
| Clearcut (n = 48) | 0.3 | 8.8 | ± | 1.6 | 60.8 | 2.4 | 48.2 | ± | 5.6 | 133.6 | ||
| Control * (n = 39) | 0.3 | 8.6 | ± | 1.8 | 60.8 | 2.4 | 41.0 | ± | 6.4 | 133.6 | ||
| Scarification (n = 9) | 1.6 | 9.8 | ± | 3.4 | 31.5 | 68.4 | 79.6 | ± | 3.3 | 99.2 | ||
| Shelterwood―High Residual (n = 23) | 1.7 | 14.5 | ± | 2.5 | 44.9 | 9.1 | 73.6 | ± | 9.7 | 153.2 | ||
| Control (n = 14) | 5.5 | 15.1 | ± | 3.0 | 43.1 | 9.1 | 72.1 | ± | 11.3 | 143.3 | ||
| Scarification (n = 9) | 1.7 | 13.7 | ± | 4.4 | 44.9 | 16.1 | 75.8 | ± | 17.5 | 153.2 | ||
| Shelterwood―Low Residual (n = 43) | 1.7 | 12.4 | ± | 1.7 | 48.3 | 2.5 | 63.8 | ± | 8.6 | 199.4 | ||
| Control (n = 36) | 1.7 | 11.1 | ± | 1.8 | 48.3 | 2.5 | 60.1 | ± | 8.9 | 199.4 | ||
| Scarification (n = 7) | 6.5 | 18.9 | ± | 4.7 | 48.3 | 31.8 | 82.7 | ± | 25.0 | 192.2 | ||
| Single-Tree Selection (n = 42) | 0.1 | 5.8 | ± | 1.2 | 45.2 | 11.9 | 68.2 | ± | 3.7 | 139.5 | ||
| Control (n = 34) | 0.1 | 5.2 | ± | 1.4 | 45.2 | 11.9 | 69.0 | ± | 4.3 | 139.5 | ||
| Scarification (n = 8) | 2.0 | 8.6 | ± | 2.0 | 20.2 | 51.4 | 64.8 | ± | 6.2 | 108.2 | ||
| New Recruits (n = 81) | 0.1 | 12.3 | ± | 2.1 | 107.8 | 3.9 | 46.9 | ± | 4.3 | 212.6 | ||
| Clearcut (n = 19) | 1.6 | 10.6 | ± | 2.4 | 45.6 | 5.3 | 48.4 | ± | 7.9 | 135.0 | ||
| Control (n = 17) | 1.7 | 11.0 | ± | 2.7 | 45.6 | 5.3 | 44.2 | ± | 8.2 | 135.0 | ||
| Scarification (n = 2) | 1.6 | 6.5 | ± | 3.5 | 11.5 | 67.2 | 84.7 | ± | 12.4 | 102.3 | ||
| Shelterwood―High Residual (n = 28) | 0.1 | 11.4 | ± | 4.0 | 107.2 | 9.6 | 31.2 | ± | 4.4 | 103.3 | ||
| Control (n = 8) | 2.0 | 27.0 | ± | 11.8 | 107.2 | 9.6 | 53.2 | ± | 11.9 | 103.3 | ||
| Scarification (n = 20) | 0.1 | 5.2 | ± | 1.5 | 27.3 | 11.6 | 22.5 | ± | 1.4 | 31.9 | ||
| Shelterwood―Low Residual (n = 21) | 3.4 | 11.4 | ± | 2.3 | 54.7 | 3.9 | 52.8 | ± | 11.3 | 212.6 | ||
| Control (n = 16) | 3.4 | 9.0 | ± | 1.0 | 18.8 | 3.9 | 39.3 | ± | 9.9 | 164.6 | ||
| Scarification (n = 5) | 6.5 | 19.1 | ± | 8.1 | 54.7 | 38.8 | 95.8 | ± | 27.9 | 212.6 | ||
| Single-tree Selection (n = 13) | 1.0 | 18.2 | ± | 7.8 | 107.8 | 34.8 | 69.0 | ± | 8.5 | 141.2 | ||
| Control (n = 7) | 1.0 | 18.5 | ± | 13.8 | 107.8 | 48.7 | 80.0 | ± | 12.6 | 141.2 | ||
| Scarification (n = 6) | 10.1 | 17.9 | ± | 5.1 | 45.2 | 34.8 | 56.1 | ± | 8.5 | 98.5 | ||
| Treatment | Mechanical Damage (%) | Browse (%) | White Pine Weevil (%) | # of Terminal Leaders Affected by Weevil Damage | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | ± | SE | ||||||
| Advance Regeneration | 23.1 | 41.7 | 30.1 | 1.3 | ± | 0.1 | ||
| Clearcut | 7.1 | 10.9 | 11.5 | 1.4 | ± | 0.2 | ||
| Control * | 4.5 | 10.3 | 7.7 | 1.3 | ± | 0.2 | ||
| Scarification | 2.6 | 0.6 | 3.8 | 1.8 | ± | 0.5 | ||
| Shelterwood—High Residual | 5.1 | 2.6 | 7.1 | 1.2 | ± | 0.2 | ||
| Control | 3.8 | 1.9 | 2.6 | 1.3 | ± | 0.3 | ||
| Scarification | 1.3 | 0.6 | 4.5 | 1.1 | ± | 0.2 | ||
| Shelterwood—Low Residual | 8.3 | 7.7 | 8.3 | 1.2 | ± | 0.2 | ||
| Control | 7.1 | 6.4 | 8.3 | 1.2 | ± | 0.2 | ||
| Scarification | 1.3 | 1.3 | 0.0 | 0.0 | ± | 0.0 | ||
| Single-Tree Selection | 2.6 | 20.5 | 3.2 | 1.0 | ± | 0.1 | ||
| Control | 2.6 | 16.0 | 3.2 | 1.0 | ± | 0.2 | ||
| Scarification | 0.0 | 4.5 | 0.0 | 0.0 | ± | 0.0 | ||
| New Recruits | 5.8 | 14.1 | 3.8 | 1.0 | ± | 0.1 | ||
| Clearcut | 1.9 | 5.8 | 0.6 | 1.0 | ± | 0.2 | ||
| Control | 0.6 | 5.8 | 0.6 | 1.0 | ± | 0.2 | ||
| Scarification | 1.3 | 0.0 | 0.0 | 0.0 | ± | 0.0 | ||
| Shelterwood—High Residual | 0.6 | 1.9 | 3.2 | 1.0 | ± | 0.2 | ||
| Control | 0.0 | 1.3 | 1.3 | 1.0 | ± | 0.3 | ||
| Scarification | 0.6 | 0.6 | 1.9 | 1.0 | ± | 0.2 | ||
| Shelterwood—Low Residual | 1.9 | 3.2 | 0.0 | 0.0 | ± | 0.0 | ||
| Control | 1.9 | 3.2 | 0.0 | 0.0 | ± | 0.0 | ||
| Scarification | 0.0 | 0.0 | 0.0 | 0.0 | ± | 0.0 | ||
| Single-Tree Selection | 1.3 | 3.2 | 0.0 | 0.0 | ± | 0.0 | ||
| Control | 1.3 | 3.2 | 0.0 | 0.0 | ± | 0.0 | ||
| Scarification | 0.0 | 0.0 | 0.0 | 0.0 | ± | 0.0 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kromholz, D.A.; Webster, C.R.; Hyslop, M.D. Spatial Patterning and Growth of Naturally Regenerated Eastern White Pine in a Northern Hardwood Silviculture Experiment. Forests 2025, 16, 1235. https://doi.org/10.3390/f16081235
Kromholz DA, Webster CR, Hyslop MD. Spatial Patterning and Growth of Naturally Regenerated Eastern White Pine in a Northern Hardwood Silviculture Experiment. Forests. 2025; 16(8):1235. https://doi.org/10.3390/f16081235
Chicago/Turabian StyleKromholz, David A., Christopher R. Webster, and Michael D. Hyslop. 2025. "Spatial Patterning and Growth of Naturally Regenerated Eastern White Pine in a Northern Hardwood Silviculture Experiment" Forests 16, no. 8: 1235. https://doi.org/10.3390/f16081235
APA StyleKromholz, D. A., Webster, C. R., & Hyslop, M. D. (2025). Spatial Patterning and Growth of Naturally Regenerated Eastern White Pine in a Northern Hardwood Silviculture Experiment. Forests, 16(8), 1235. https://doi.org/10.3390/f16081235