A 36-Year Assessment of Mangrove Ecosystem Dynamics in China Using Kernel-Based Vegetation Index


Abstract
1. Introduction
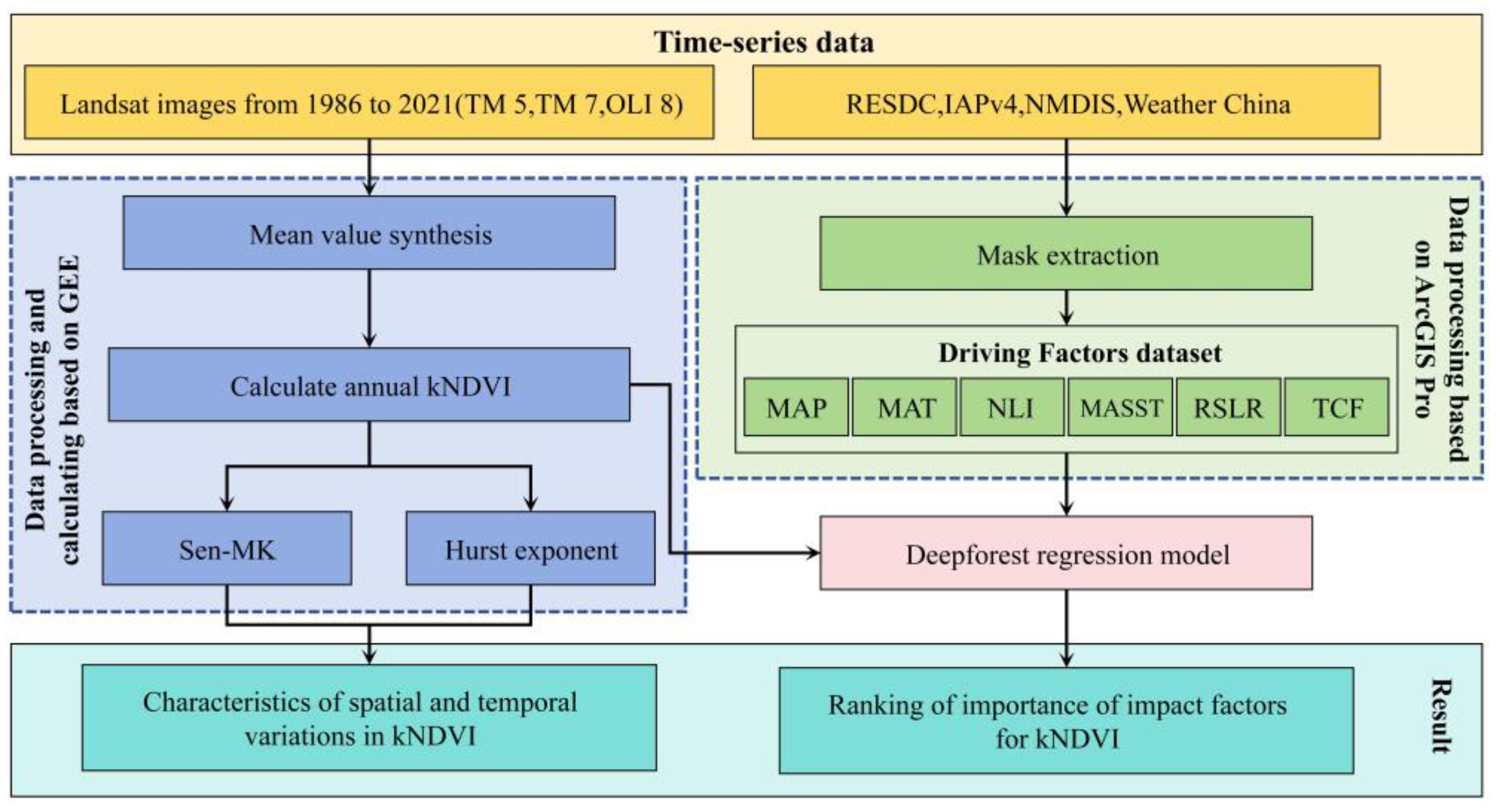
2. Materials and Methods
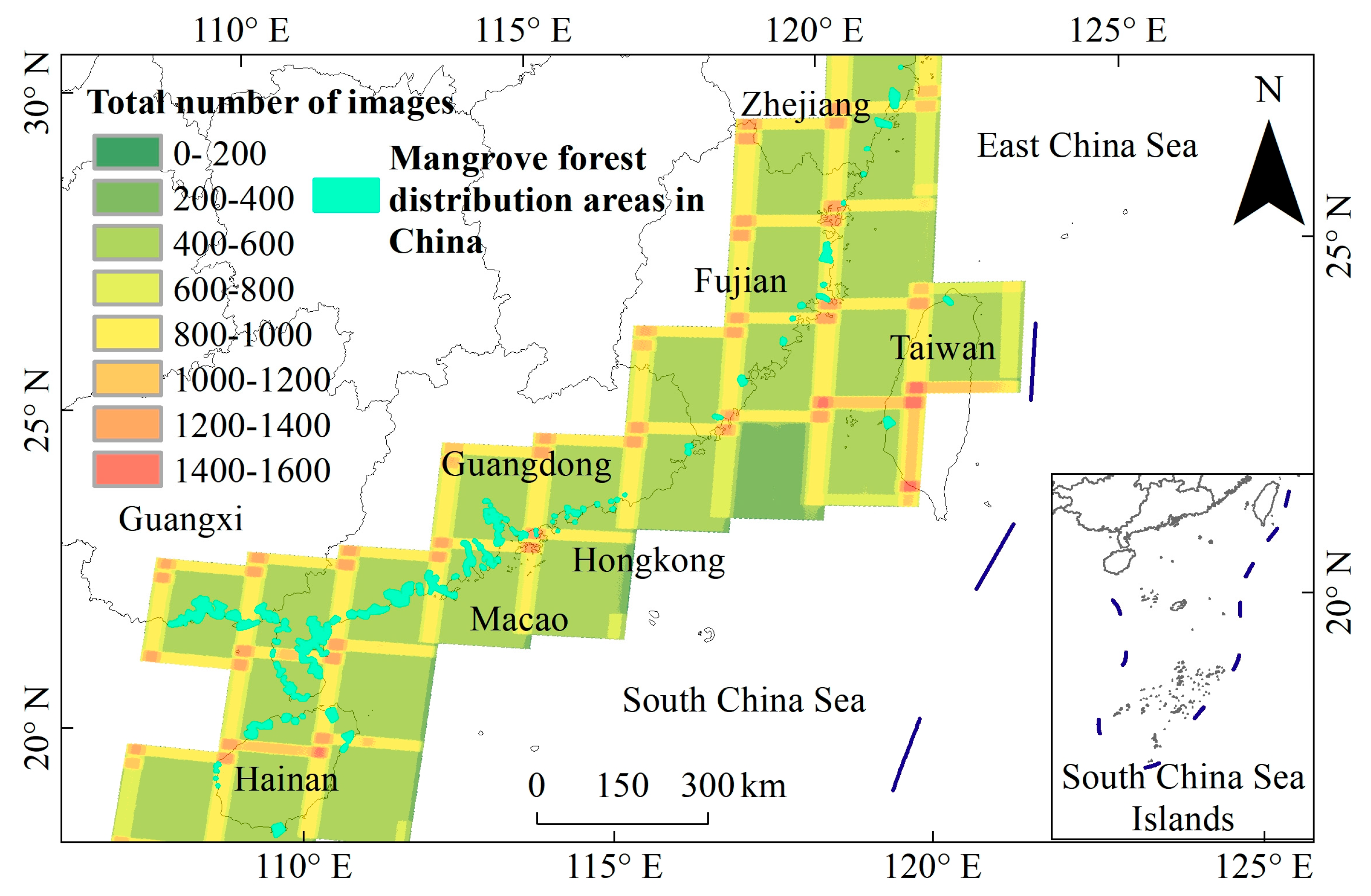
2.1. Study Area
2.2. Data Source and Preprocessing
2.2.1. Landsat Data Collection and Preprocessing
2.2.2. Driving Factor Dataset
- (1)
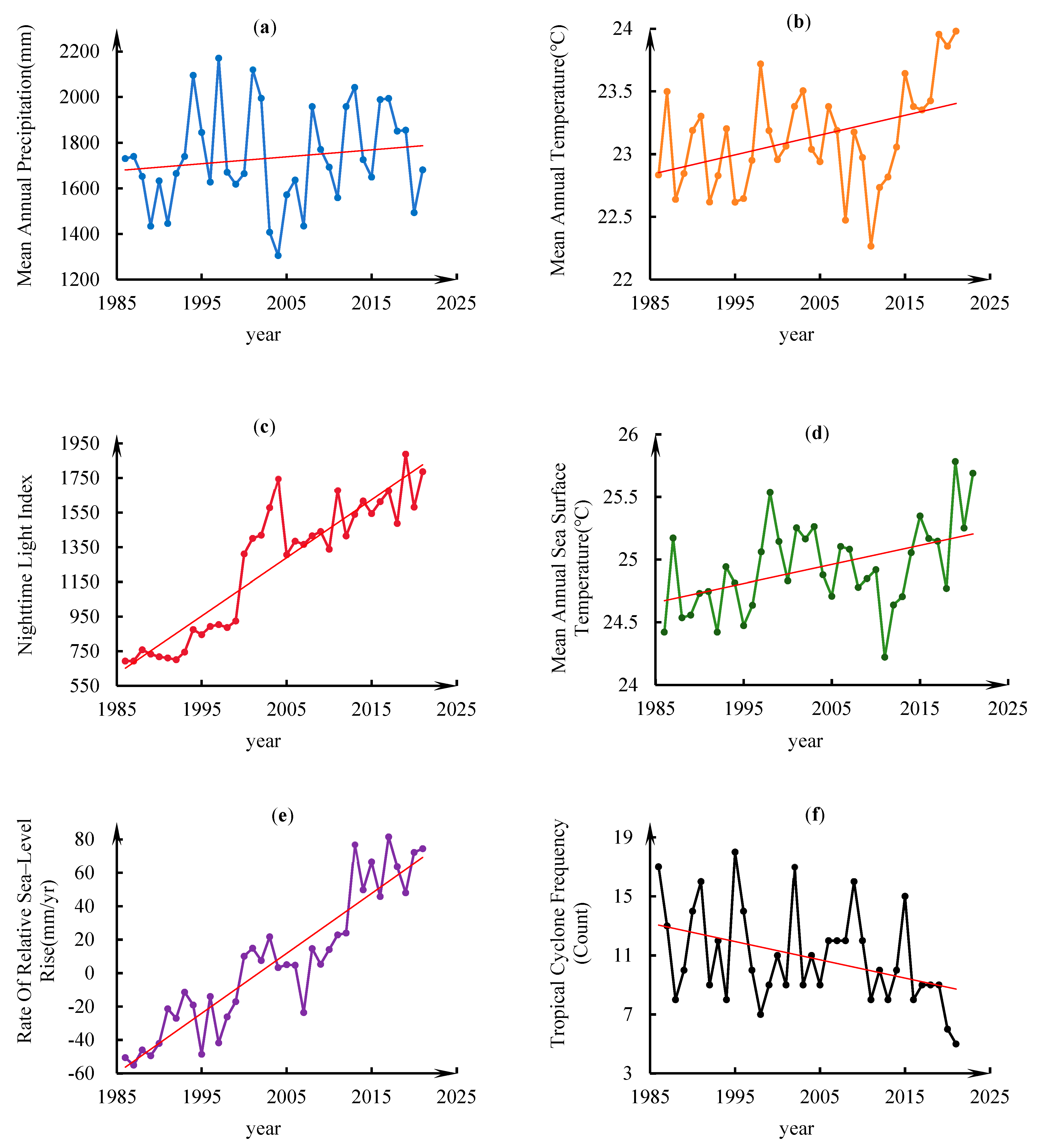
- Climatic parameters: Mean annual precipitation (MAP) and mean annual temperature (MAT), gridded datasets at 1 km spatial resolution, were acquired from the Resource and Environment Science and Data Center (RESDC). MAP was selected for its critical role in regulating freshwater input and soil salinity gradients, which directly influence mangrove water balance and salt stress responses. MAT serves as a key thermodynamic control on mangrove photosynthesis efficiency and species distribution thresholds.
- (2)
- Anthropogenic activity proxy: The nighttime light index (NLI), a 1 km resolution raster covering a continuous inland belt extending 50 km from the coastline, was obtained from RESDC to quantify human settlement intensity. This variable was prioritized as it effectively captures coastal urbanization patterns and land-use changes that induce habitat fragmentation and pollution pressures on mangrove ecosystems.
- (3)
- Marine environmental drivers: Mean annual sea surface temperature (MASST), derived from the 1°-gridded Global Ocean Temperature and Heat Content Dataset (IAPv4) via the Ocean and Climate Team portal (http://www.ocean.iap.ac.cn, accessed on 18 February 2025), was incorporated as a critical thermal constraint on mangrove growth, aligning with established drivers in coastal biogeographic studies in which sea-level proximity governs thermal exposure. The rate of relative sea-level rise (RSLR), with annual mean values for coastal waters adjacent to the study area, was extracted from the National Marine Data and Information Service (NMDIS; https://mds.nmdis.org.cn, accessed on 18 February 2025), as it governs tidal inundation regimes and sediment dynamics critical for mangrove establishment [30]. The frequency of tropical cyclones (TCF) was determined based on event counts from 1986 to 2021, as reported by the China Meteorological Administration (https://weather.com.cn, accessed on 18 February 2025). This approach was employed to account for extreme climatic disturbances, which have the potential to cause mechanical damage and salinity fluctuations.
2.3. Calculation of kNDVI
2.4. Trend Analysis Method
2.4.1. Mann–Kendall Test and Sen’s Slope Estimator Model
2.4.2. Hurst Exponent
2.5. Detection of Variable Importance Using the Deep Forest Algorithm
3. Results
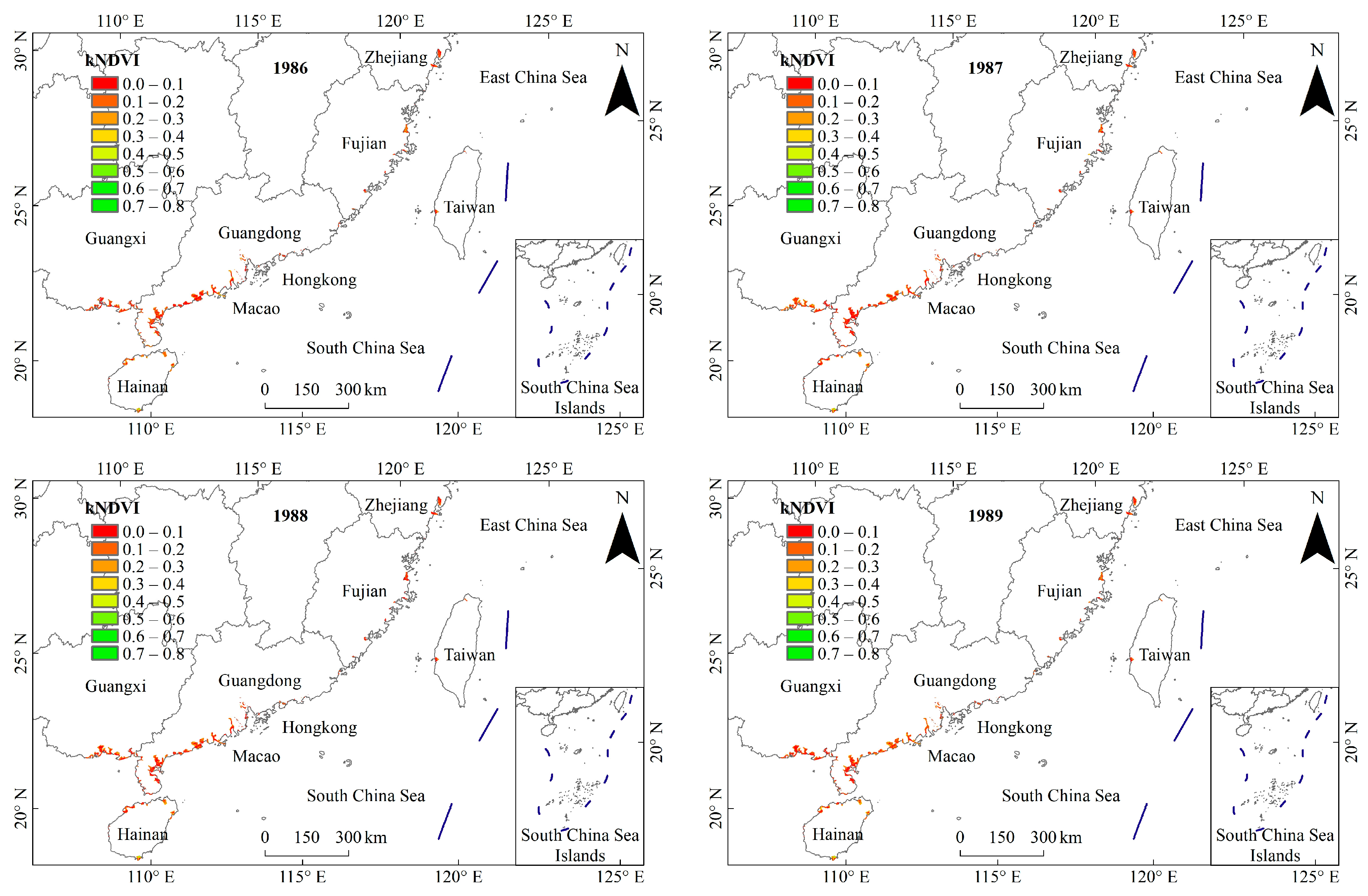
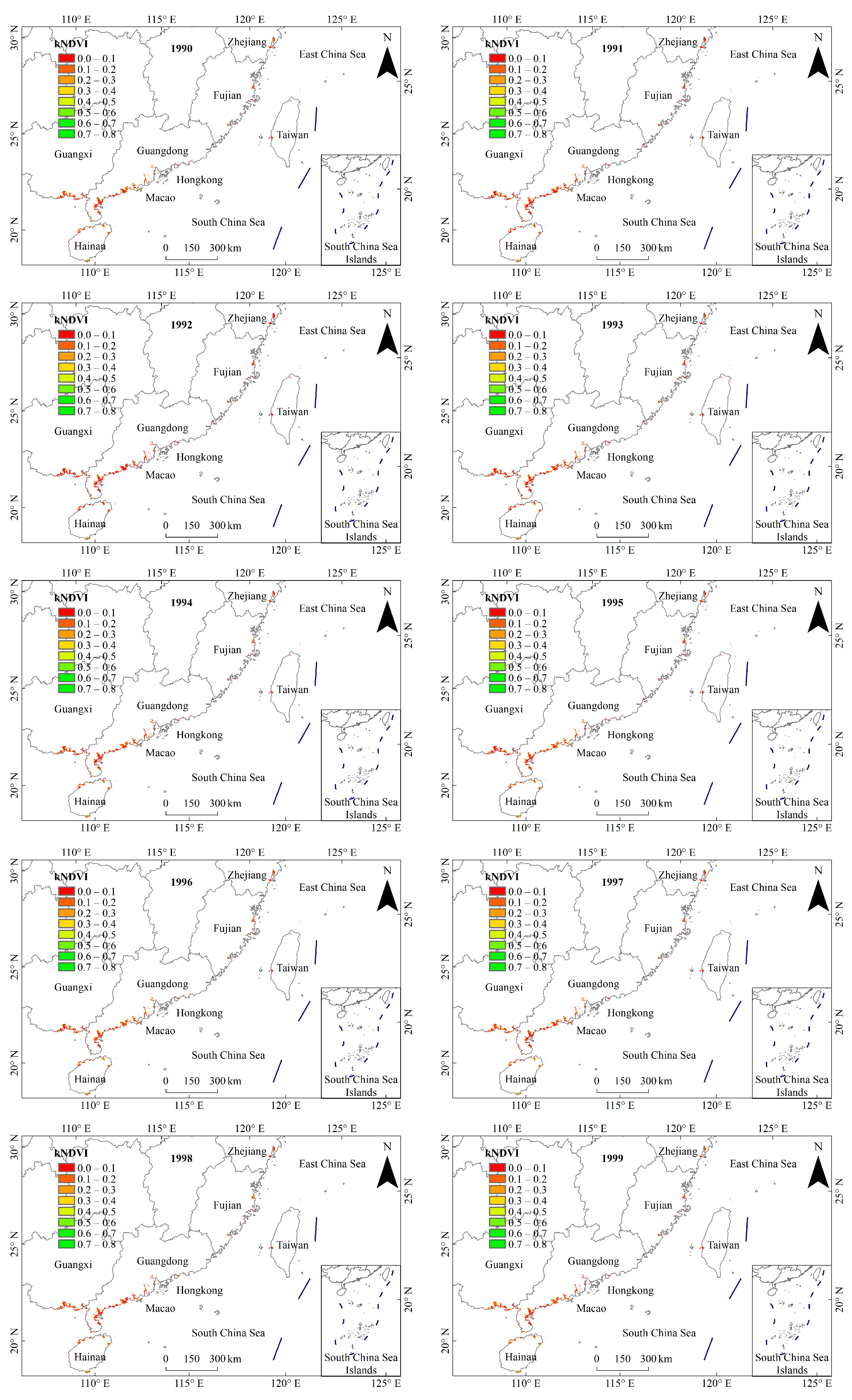
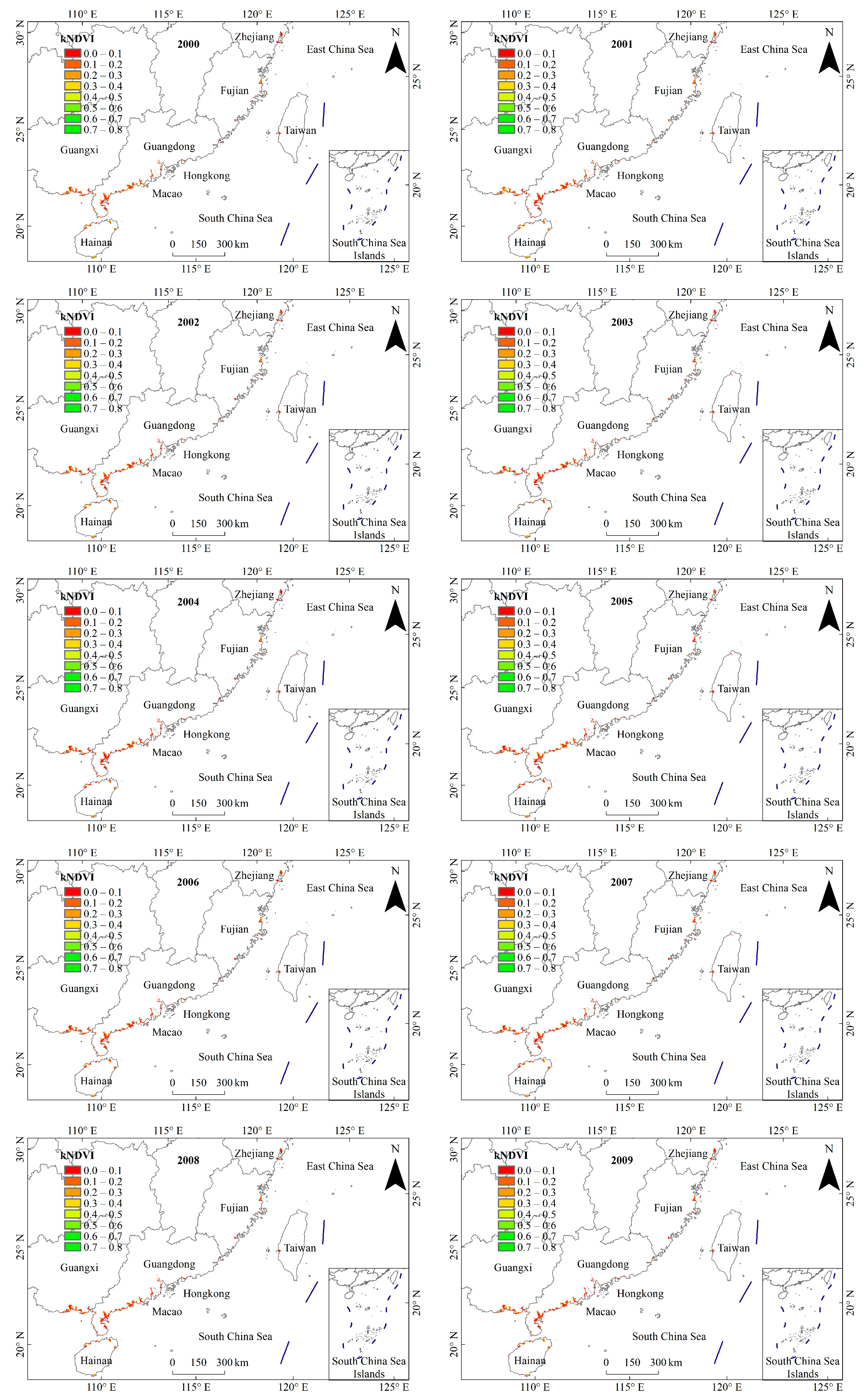
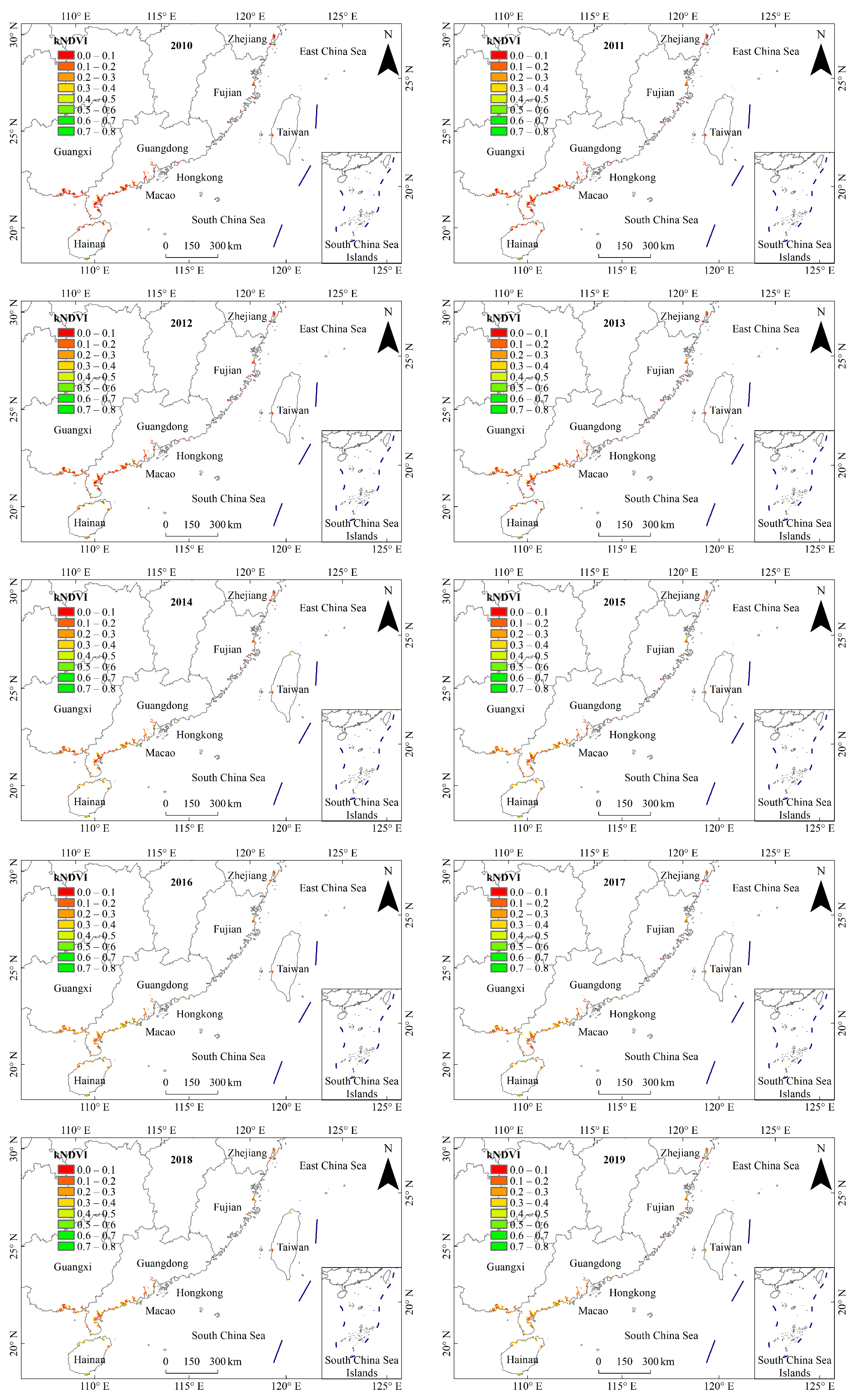
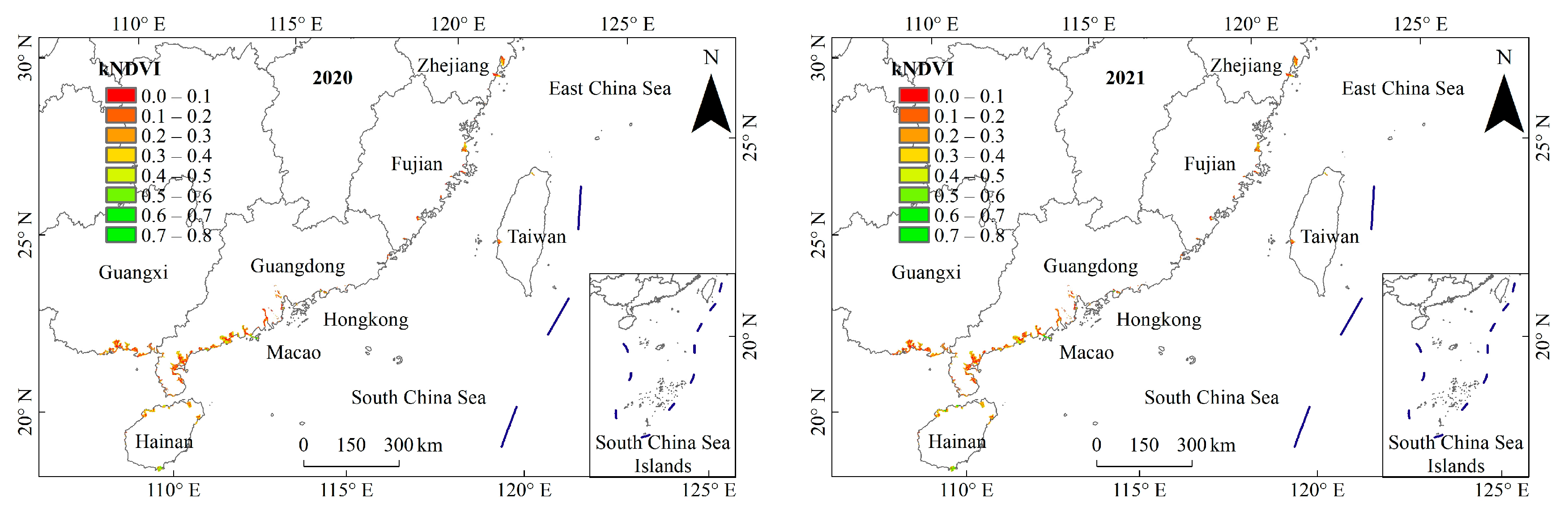
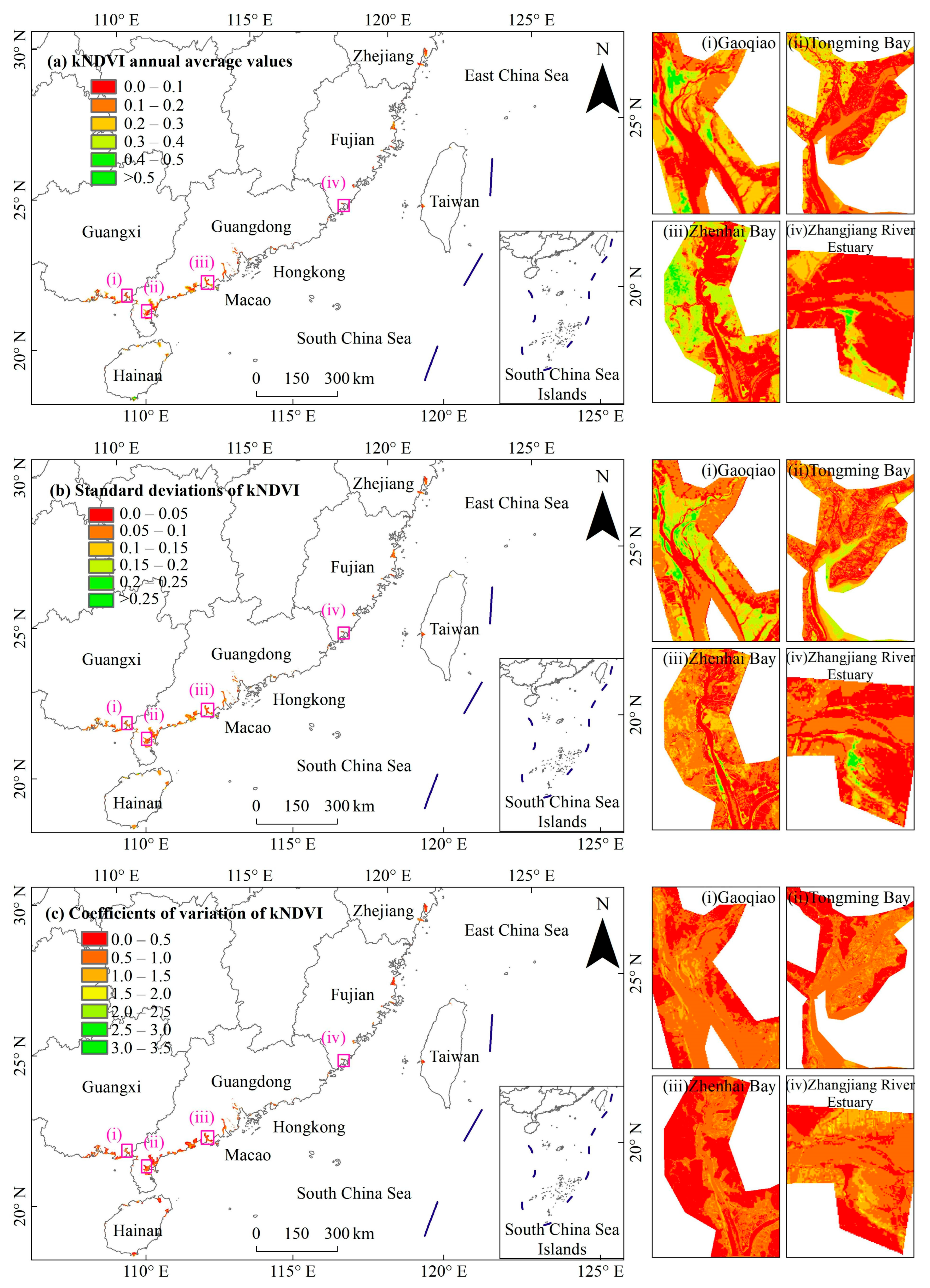
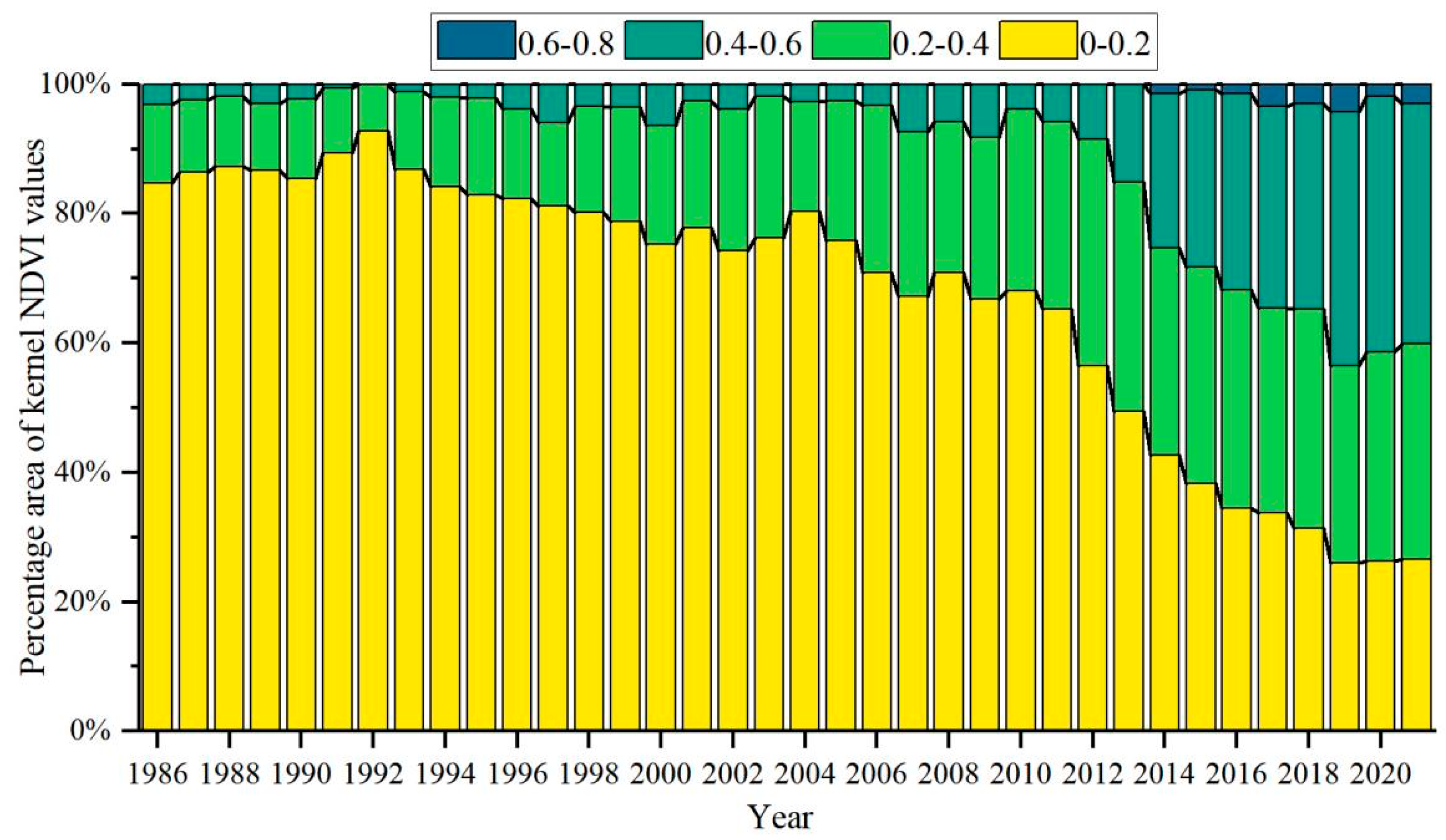
3.1. General Characteristics of Mangrove kNDVI
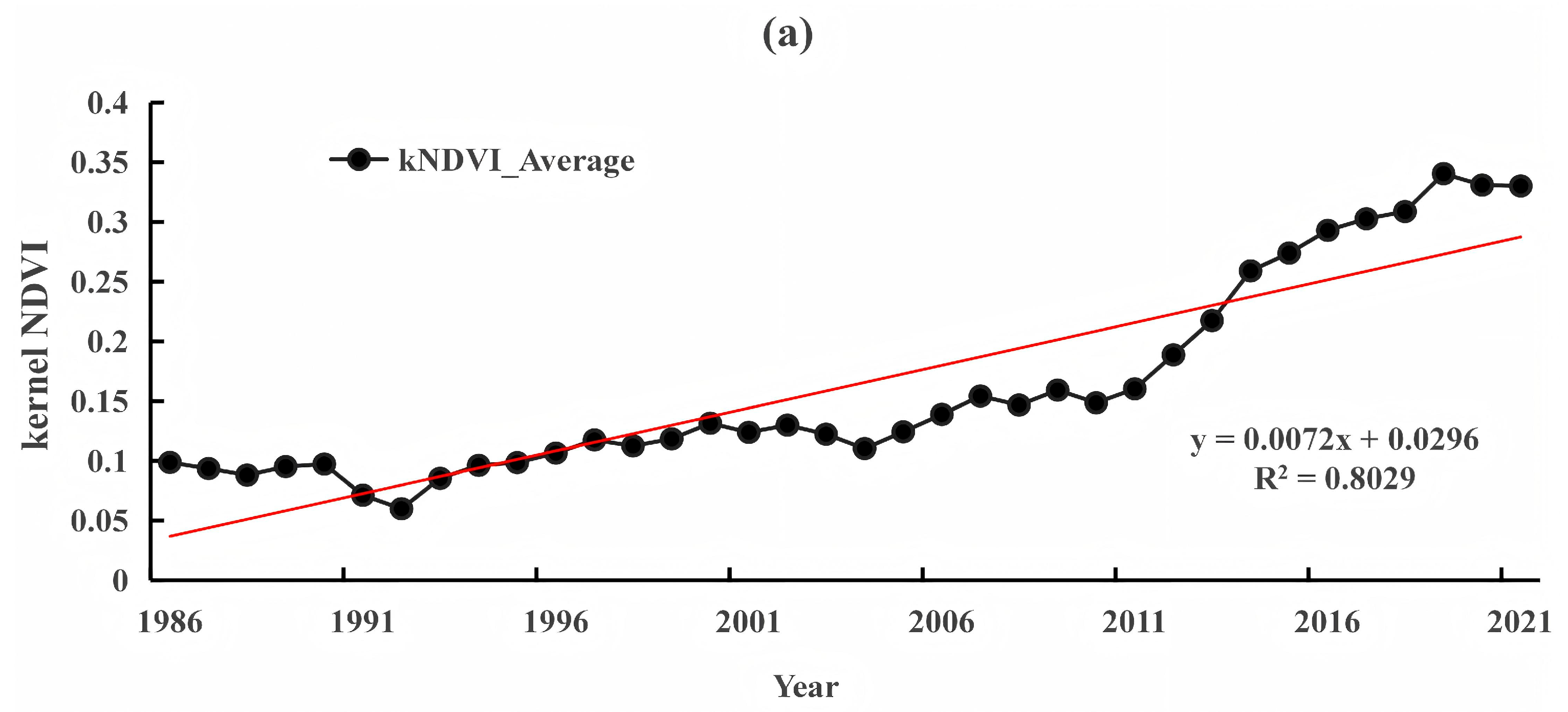
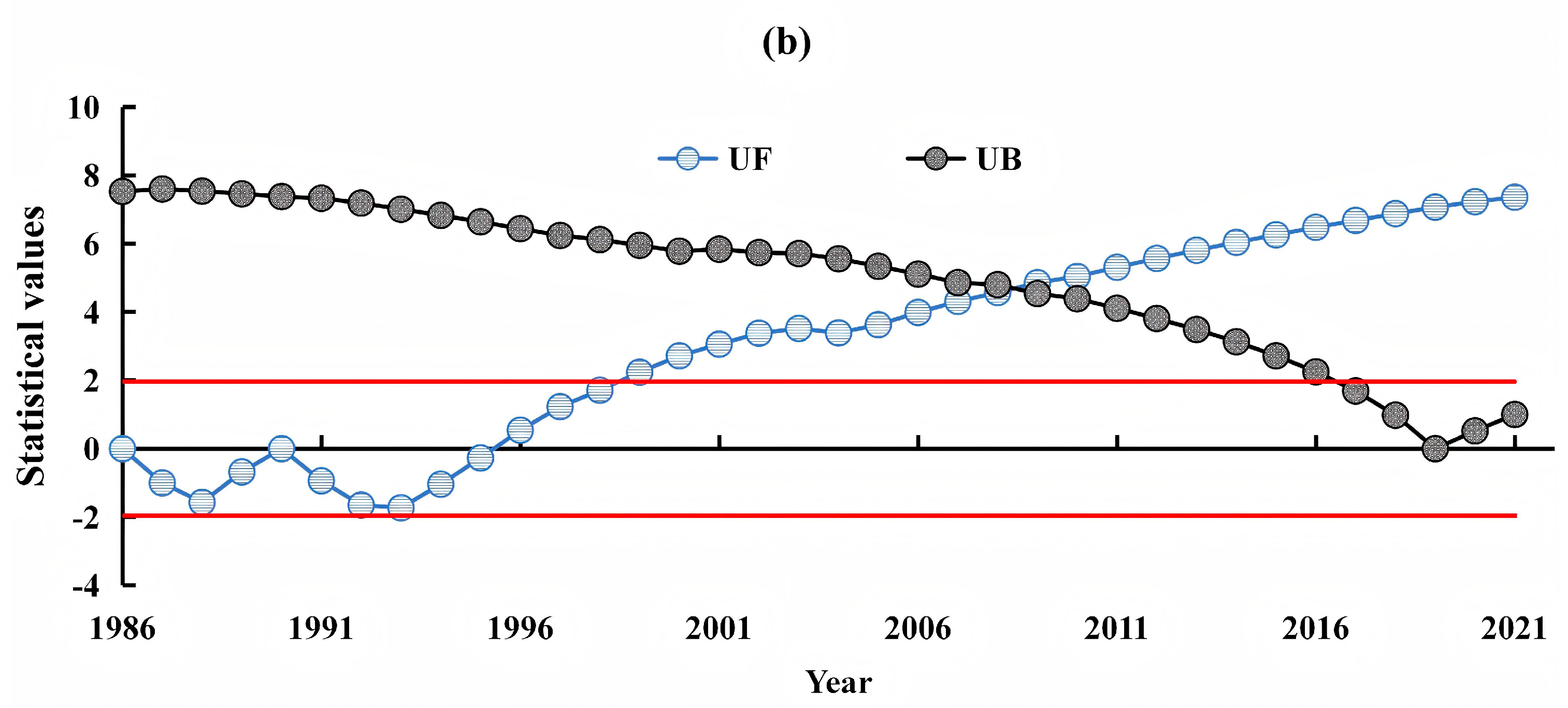
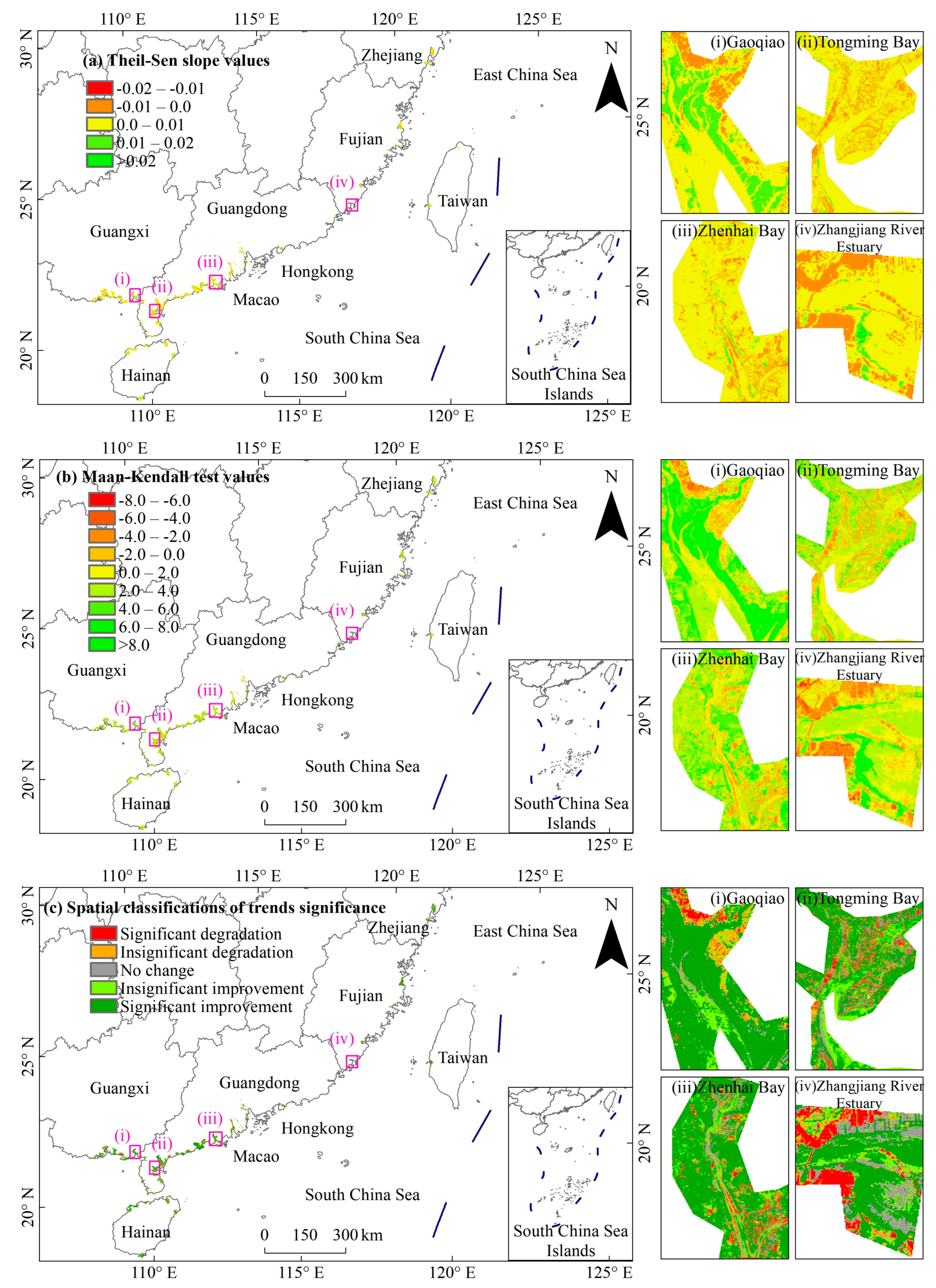
3.2. Dynamic TRENDS in Mangrove kNDVI During 1986–2021
3.3. Spatial Differences in Mangrove kNDVI Dynamics Trends
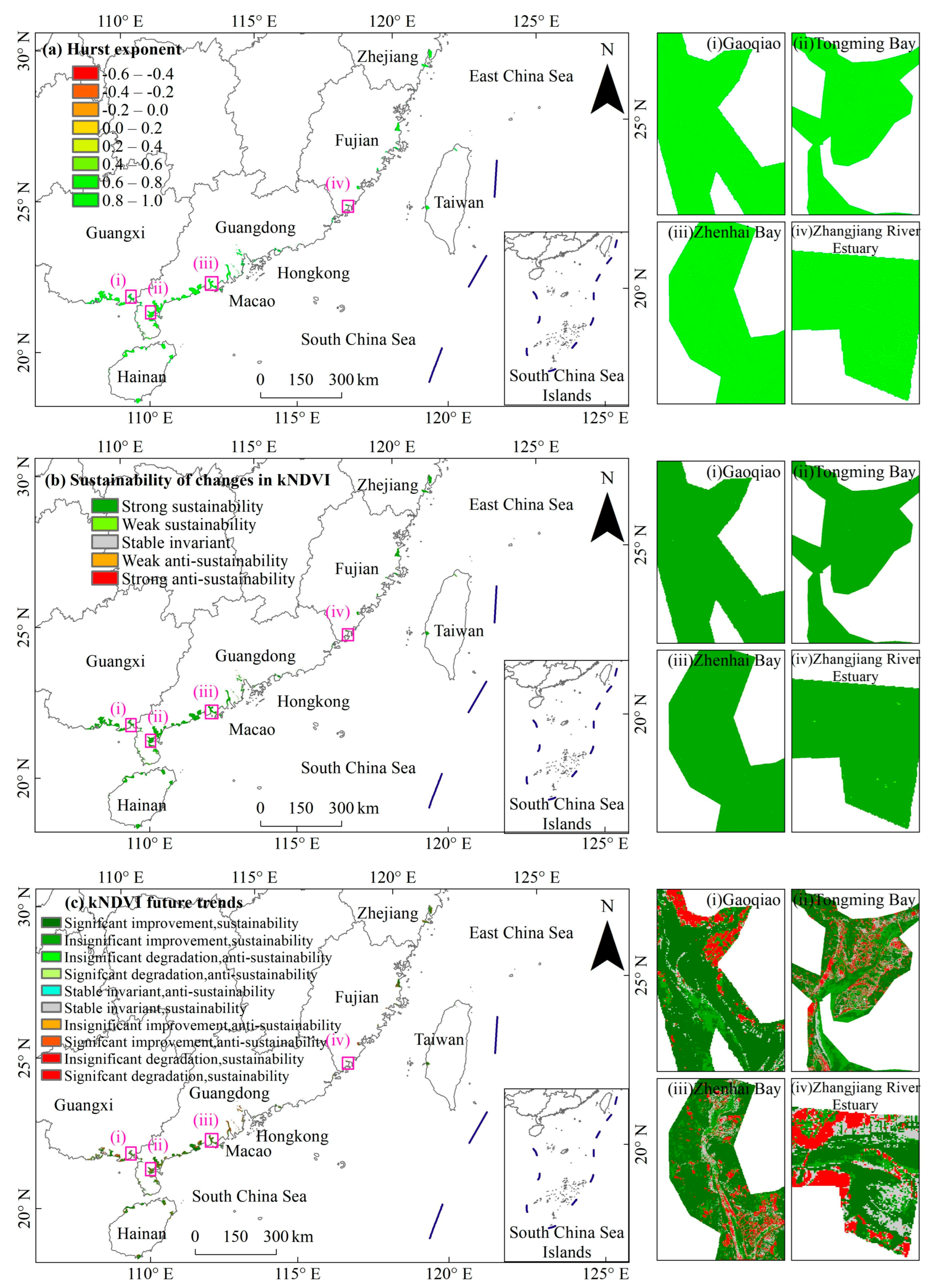
3.4. Consistency of Future Trends in Mangrove kNDVI Dynamics
3.5. Analysis of Influencing Factors
4. Discussion
4.1. Spatiotemporal Variation Trends of kNDVI in China’s Mangrove Forests over the Past 36 Years
4.2. Driving Factors of kNDVI Changes in China’s Mangrove Forests
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Atwood, T.B.; Connolly, R.M.; Almahasheer, H.; Carnell, P.E.; Duarte, C.M.; Ewers Lewis, C.J.; Irigoien, X.; Kelleway, J.J.; Lavery, P.S.; Macreadie, P.I. Global patterns in mangrove soil carbon stocks and losses. Nat. Clim. Change 2017, 7, 523–528. [Google Scholar] [CrossRef]
- McKee, K.L.; Cahoon, D.R.; Feller, I.C. Caribbean mangroves adjust to rising sea level through biotic controls on change in soil elevation. Glob. Ecol. Biogeogr. 2007, 16, 545–556. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Z.; Mao, D.; Ren, C.; Song, K.; Zhao, C.; Wang, C.; Xiao, X.; Wang, Y. Mapping global distribution of mangrove forests at 10-m resolution. Sci. Bull. 2023, 68, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Sandilyan, S.; Kathiresan, K. Mangrove conservation: A global perspective. Biodivers. Conserv. 2012, 21, 3523–3542. [Google Scholar] [CrossRef]
- Ruan, L.; Yan, M.; Zhang, L.; Fan, X.; Yang, H. Spatial-temporal NDVI pattern of global mangroves: A growing trend during 2000–2018. Sci. Total Environ. 2022, 844, 157075. [Google Scholar] [CrossRef] [PubMed]
- van Bijsterveldt, C.E.; Herman, P.M.; van Wesenbeeck, B.K.; Ramadhani, S.; Heuts, T.S.; van Starrenburg, C.; Tas, S.A.; Triyanti, A.; Helmi, M.; Tonneijck, F.H. Subsidence reveals potential impacts of future sea level rise on inhabited mangrove coasts. Nat. Sustain. 2023, 6, 1565–1577. [Google Scholar] [CrossRef]
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove Forests: One of the World’s Threatened Major Tropical Environments: At least 35% of the area of mangrove forests has been lost in the past two decades, losses that exceed those for tropical rain forests and coral reefs, two other well-known threatened environments. Bioscience 2001, 51, 807–815. [Google Scholar]
- Duke, N.C.; Meynecke, J.-O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D. A world without mangroves? Science 2007, 317, 41–42. [Google Scholar] [CrossRef]
- Hu, L.; Li, W.; Xu, B. Monitoring mangrove forest change in China from 1990 to 2015 using Landsat-derived spectral-temporal variability metrics. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 88–98. [Google Scholar] [CrossRef]
- Alatorre, L.C.; Sánchez-Carrillo, S.; Miramontes-Beltrán, S.; Medina, R.J.; Torres-Olave, M.E.; Bravo, L.C.; Wiebe, L.C.; Granados, A.; Adams, D.K.; Sánchez, E. Temporal changes of NDVI for qualitative environmental assessment of mangroves: Shrimp farming impact on the health decline of the arid mangroves in the Gulf of California (1990–2010). J. Arid Environ. 2016, 125, 98–109. [Google Scholar] [CrossRef]
- Nepita-Villanueva, M.R.; Berlanga-Robles, C.A.; Ruiz-Luna, A.; Morales Barcenas, J.H. Spatio-temporal mangrove canopy variation (2001–2016) assessed using the MODIS enhanced vegetation index (EVI). J. Coast. Conserv. 2019, 23, 589–597. [Google Scholar] [CrossRef]
- Pham, L.T.; Brabyn, L. Monitoring mangrove biomass change in Vietnam using SPOT images and an object-based approach combined with machine learning algorithms. ISPRS J. Photogramm. Remote Sens. 2017, 128, 86–97. [Google Scholar] [CrossRef]
- Zhang, R.; Jia, M.; Wang, Z.; Zhou, Y.; Mao, D.; Ren, C.; Zhao, C.; Liu, X. Tracking annual dynamics of mangrove forests in mangrove National Nature Reserves of China based on time series Sentinel-2 imagery during 2016–2020. Int. J. Appl. Earth Obs. Geoinf. 2022, 112, 102918. [Google Scholar] [CrossRef]
- Wulder, M.A.; Loveland, T.R.; Roy, D.P.; Crawford, C.J.; Masek, J.G.; Woodcock, C.E.; Allen, R.G.; Anderson, M.C.; Belward, A.S.; Cohen, W.B. Current status of Landsat program, science, and applications. Remote Sens. Environ. 2019, 225, 127–147. [Google Scholar] [CrossRef]
- Yang, J.; Yan, D.; Yu, Z.; Wu, Z.; Wang, H.; Liu, W.; Liu, S.; Yuan, Z. NDVI variations of different terrestrial ecosystems and their response to major driving factors on two side regions of the Hu-Line. Ecol. Indic. 2024, 159, 111667. [Google Scholar] [CrossRef]
- Rouse, J.W., Jr.; Haas, R.H.; Deering, D.; Schell, J.; Harlan, J.C. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; NASA: Washington, DC, USA, 1974.
- Camps-Valls, G.; Campos-Taberner, M.; Moreno-Martínez, Á.; Walther, S.; Duveiller, G.; Cescatti, A.; Mahecha, M.D.; Muñoz-Marí, J.; García-Haro, F.J.; Guanter, L. A unified vegetation index for quantifying the terrestrial biosphere. Sci. Adv. 2021, 7, eabc7447. [Google Scholar] [CrossRef]
- Huete, A.; Liu, H.; Batchily, K.; Van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Wannasiri, W.; Nagai, M.; Honda, K.; Santitamnont, P.; Miphokasap, P. Extraction of Mangrove Biophysical Parameters Using Airborne LiDAR. Remote Sens. 2013, 5, 1787–1808. [Google Scholar] [CrossRef]
- Wang, Q.; Moreno-Martínez, Á.; Muñoz-Marí, J.; Campos-Taberner, M.; Camps-Valls, G. Estimation of vegetation traits with kernel NDVI. ISPRS J. Photogramm. Remote Sens. 2023, 195, 408–417. [Google Scholar] [CrossRef]
- Friess, D.A.; Rogers, K.; Lovelock, C.E.; Krauss, K.W.; Hamilton, S.E.; Lee, S.Y.; Lucas, R.; Primavera, J.; Rajkaran, A.; Shi, S. The State of the World’s Mangrove Forests: Past, Present, and Future. Annu. Rev. Environ. Resour. 2019, 44, 89–115. [Google Scholar] [CrossRef]
- Zhou, Z.-H.; Feng, J. Deep forest. Natl. Sci. Rev. 2018, 6, 74–86. [Google Scholar] [CrossRef]
- Narváez Salcedo, S. Spatial and Temporal Trend of EVI in Mangrove Forests of Coastal Lagoons in the Colombian Caribbean. Bachelor’s Thesis, Universidad del Norte, Barranquilla, Colombia, 2022. [Google Scholar]
- Wang, Z.; Liu, K.; Cao, J.; Peng, L.; Wen, X. Annual change analysis of mangrove forests in China during 1986–2021 based on Google Earth engine. Forests 2022, 13, 1489. [Google Scholar] [CrossRef]
- Samanta, S.; Hazra, S.; Mondal, P.P.; Chanda, A.; Giri, S.; French, J.R.; Nicholls, R.J. Assessment and attribution of mangrove Forest changes in the Indian Sundarbans from 2000 to 2020. Remote Sens. 2021, 13, 4957. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2007, 76, 1–13. [Google Scholar] [CrossRef]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Huang, F.; Tang, L.; Li, R. Analysis of Ecological Benefits of Mangrove Protection and RestorationMeasures in the Mainland of China in the Past 40 Years. Acta Sci. Nat. Univ. Pekin. 2023, 59, 813–822. [Google Scholar] [CrossRef]
- Krauss, K.W.; McKee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How mangrove forests adjust to rising sea level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Griffin: Santa Barbara, CA, USA, 1948. [Google Scholar]
- Mann, H.B. Nonparametric tests against trend. Econom. J. Econom. Soc. 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Hurst, H.E. Long-term storage capacity of reservoirs. Trans. Am. Soc. Civ. Eng. 1951, 116, 770–799. [Google Scholar] [CrossRef]
- Zhong, X.; Li, J.; Wang, J.; Zhang, J.; Liu, L.; Ma, J. Linear and nonlinear characteristics of long-term NDVI using trend analysis: A case study of Lancang-Mekong River Basin. Remote Sens. 2022, 14, 6271. [Google Scholar] [CrossRef]
- Jiang, W.; Yuan, L.; Wang, W.; Cao, R.; Zhang, Y.; Shen, W. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecol. Indic. 2015, 51, 117–126. [Google Scholar] [CrossRef]
- Zhao, H.; Xu, X.; Tang, J.; Wang, Z.; Miao, C. Understanding the key factors and future trends of ecosystem service value to support the decision management in the cluster cities around the Yellow River floodplain area. Ecol. Indic. 2023, 154, 110544. [Google Scholar] [CrossRef]
- Al-huqail, A.A.; Islam, Z.; Al-Harbi, H.F. Mangroves trend and their impact on surface temperature in Al-Wajh Lagoon: A study aligned with Saudi Arabia’s vision 2030. Front. Environ. Sci. 2024, 12, 1439425. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Z.; Mao, D.; Huang, C.; Lu, C. Spatial-temporal changes of China’s mangrove forests over the past 50years: An analysis towards the Sustainable Development Goals(SDGs). Chin. Sci. Bull. 2021, 66, 3886–3901. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Murray, A.B. A coupled geomorphic and ecological model of tidal marsh evolution. Proc. Natl. Acad. Sci. USA 2007, 104, 6118–6122. [Google Scholar] [CrossRef]
- Saintilan, N.; Khan, N.; Ashe, E.; Kelleway, J.; Rogers, K.; Woodroffe, C.; Horton, B.P. Thresholds of mangrove survival under rapid sea level rise. Science 2020, 368, 1118–1121. [Google Scholar] [CrossRef]
- Wei, S.; Zhang, H.; Xu, Z.; Lin, G.; Lin, Y.; Liang, X.; Ling, J.; Wee, A.K.S.; Lin, H.; Zhou, Y. Coastal urbanization may indirectly positively impact growth of mangrove forests. Commun. Earth Environ. 2024, 5, 608. [Google Scholar] [CrossRef]
- Sharifian, S.; Kamrani, E.; Saeedi, H. Global biodiversity and biogeography of mangrove crabs: Temperature, the key driver of latitudinal gradients of species richness. J. Therm. Biol. 2020, 92, 102692. [Google Scholar] [CrossRef]
- Ovalle, A.; Rezende, C.; Lacerda, L.; Silva, C.A.R. Factors affecting the hydrochemistry of a mangrove tidal creek, Sepetiba Bay, Brazil. Estuar. Coast. Shelf Sci. 1990, 31, 639–650. [Google Scholar] [CrossRef]
- Gao, X.; Leung, F.; Wang, W. Assessing typhoon impacts on mangroves and post-typhoon recovery patterns to inform projects using mangroves as nature-based solutions. Glob. Ecol. Conserv. 2025, 61, e03667. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Satellite | Operational Period | Sensor | Spatial Resolution (m) | GEE Collection Identifier |
|---|---|---|---|---|
| Landsat-5 | 1984–2011 | TM | 30 m | LANDSAT/LT05/C01/T1_SR/TOA |
| Landsat-7 | 1999–2021 | ETM+ | 30 m | LANDSAT_7/02/T1/TOA |
| Landsat-8 | 2013–2021 | OLI | 30, 15 m | LANDSAT/LC08/C01/T1_TOA |
| Category | Variables | Spatial Resolution | Unit | Data Source |
|---|---|---|---|---|
| Climatic | Mean Annual Precipitation (MAP) | 1 km | mm | Resources and Environmental Science and Data Center, Chinese Academy of SciencesAnnual |
| Mean Annual Temperature (MAT) | 1 km | °C | Resources and Environmental Science and Data Center, Chinese Academy of SciencesAnnual | |
| Mean Annual Sea Surface Temperature (MASST) | 1° | °C | 1°–gridded Global Ocean Temperature and Heat Content Dataset | |
| Marine Environmental Drivers | Rate Of Relative Sea–Level Rise (RSLR) | – | mm/yr | National Marine Science Data Center Typhoon Frequency |
| Tropical Cyclone Frequency (TCF) | – | Count | China Meteorological Administration (CMA) | |
| Anthropogenic | Nighttime Light Index (NLI) | 1 km | – | Resources and Environmental Science and Data Center, Chinese Academy of Sciences |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Y.; Huang, M.; Chen, Y.; Chen, B.; Ma, L.; Zhao, W.; Fu, D. A 36-Year Assessment of Mangrove Ecosystem Dynamics in China Using Kernel-Based Vegetation Index. Forests 2025, 16, 1143. https://doi.org/10.3390/f16071143
Pan Y, Huang M, Chen Y, Chen B, Ma L, Zhao W, Fu D. A 36-Year Assessment of Mangrove Ecosystem Dynamics in China Using Kernel-Based Vegetation Index. Forests. 2025; 16(7):1143. https://doi.org/10.3390/f16071143
Chicago/Turabian StylePan, Yiqing, Mingju Huang, Yang Chen, Baoqi Chen, Lixia Ma, Wenhui Zhao, and Dongyang Fu. 2025. "A 36-Year Assessment of Mangrove Ecosystem Dynamics in China Using Kernel-Based Vegetation Index" Forests 16, no. 7: 1143. https://doi.org/10.3390/f16071143
APA StylePan, Y., Huang, M., Chen, Y., Chen, B., Ma, L., Zhao, W., & Fu, D. (2025). A 36-Year Assessment of Mangrove Ecosystem Dynamics in China Using Kernel-Based Vegetation Index. Forests, 16(7), 1143. https://doi.org/10.3390/f16071143