
Changes in Plant Nitrogen Uptake Strategies Following Vegetation Recovery in Karst Regions

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Field Hydroponic Experimental Design
2.3. Plant Root Functional Traits
2.4. Plant Nitrogen Uptake Strategies
2.5. Available Soil Nitrogen
2.6. Data Statistics and Analysis
3. Results
3.1. Soil Nitrogen Availability
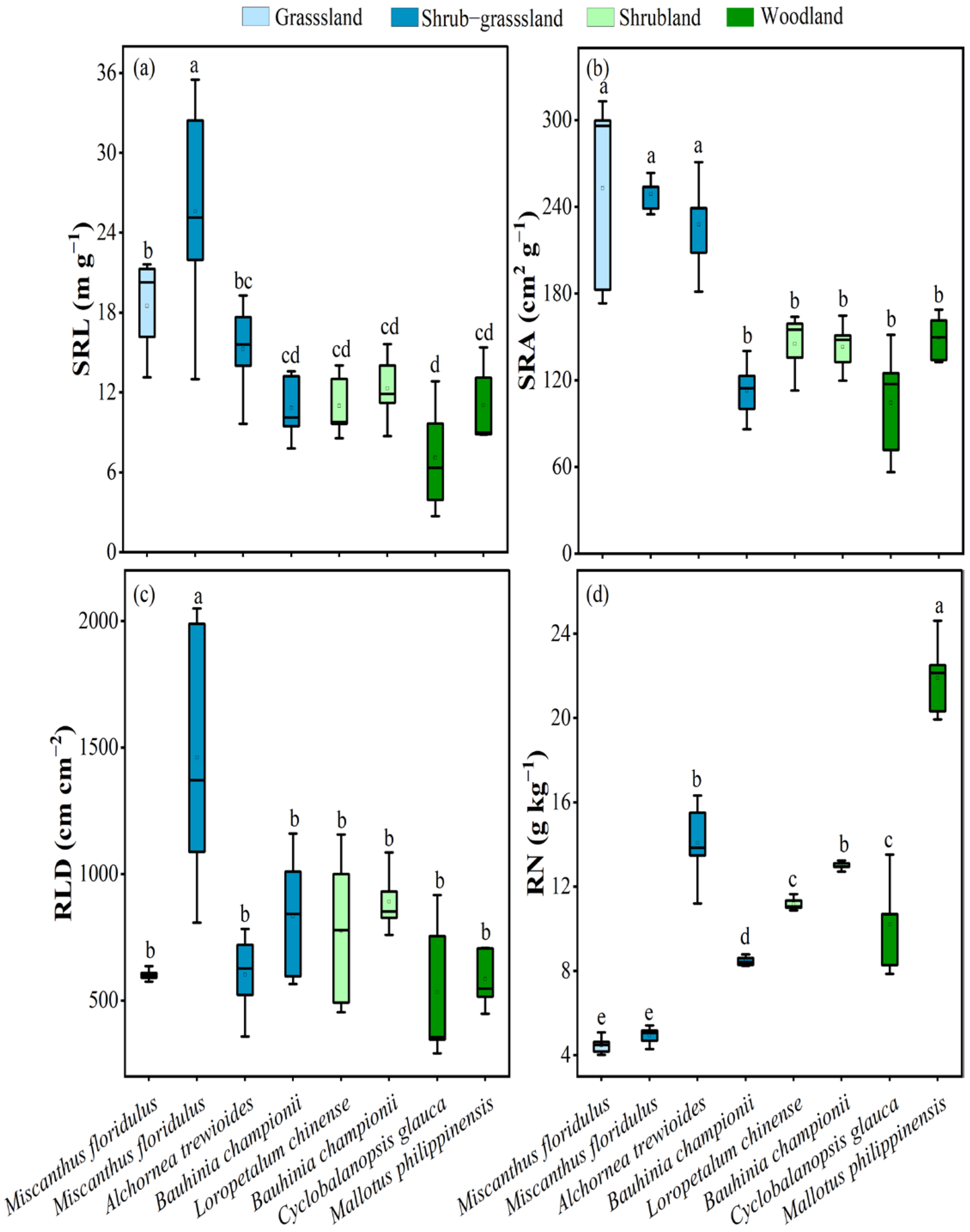
3.2. Root Functional Traits of Dominant Plants
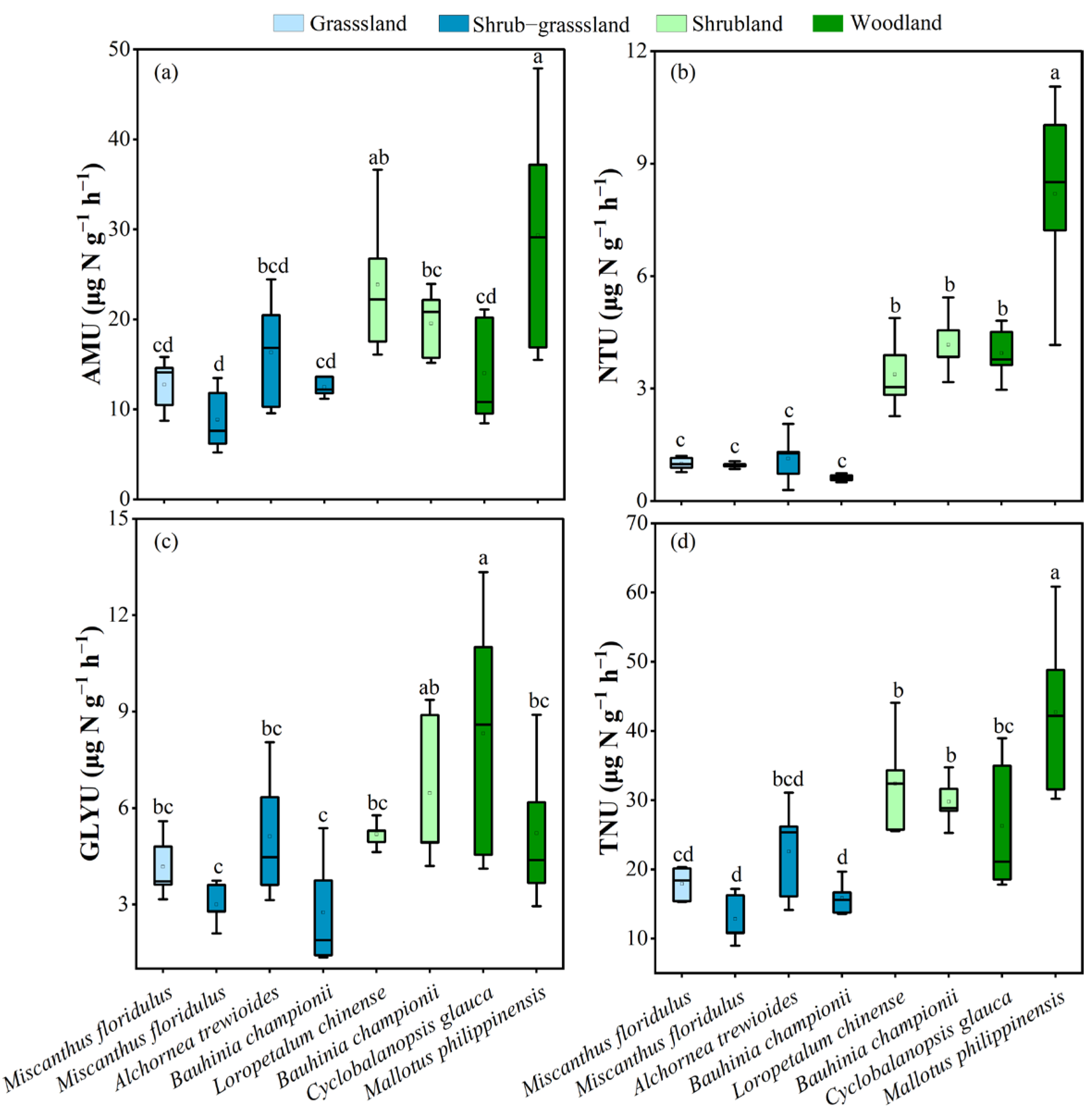
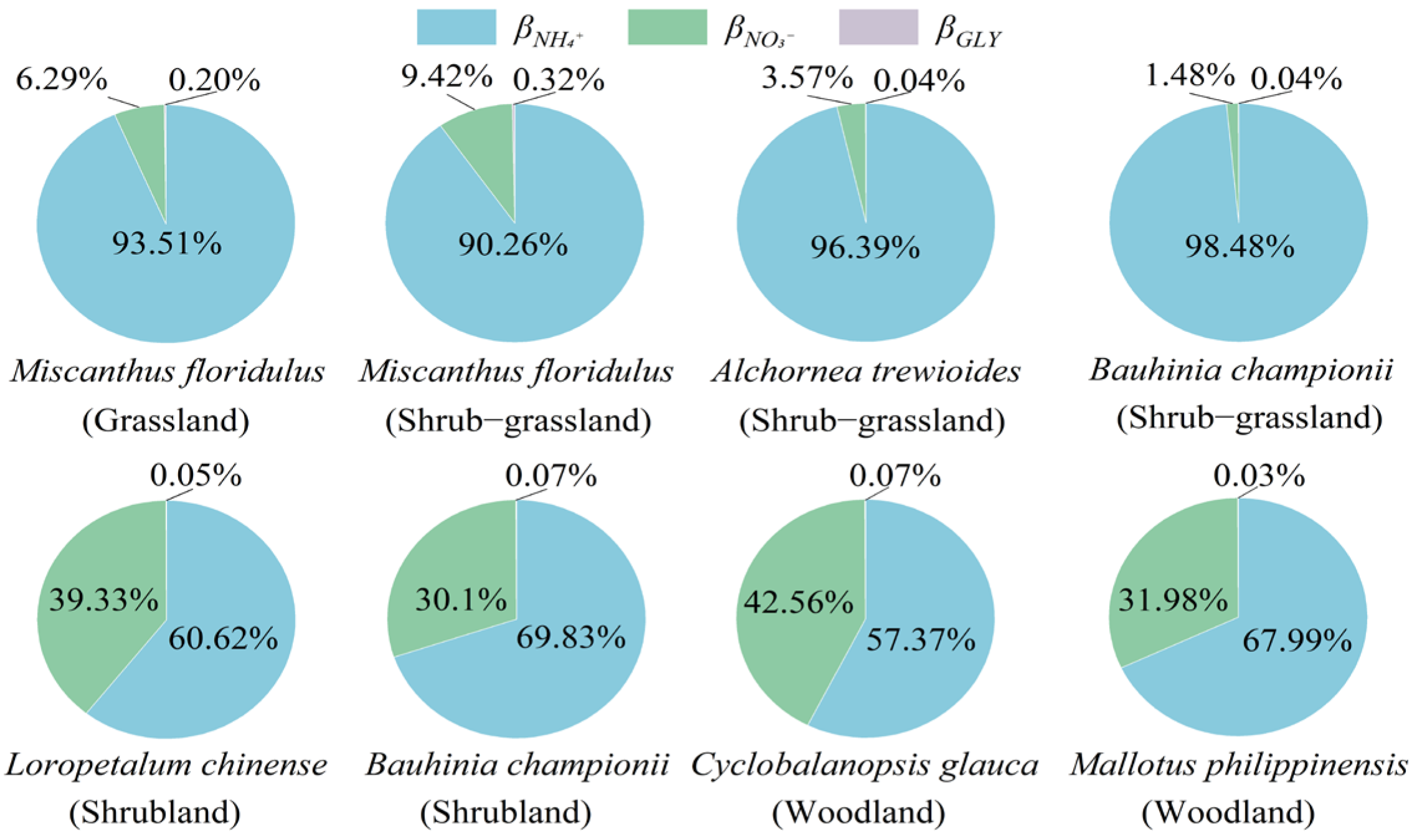
3.3. Nitrogen Uptake Rate and Ratio of Dominant Plants
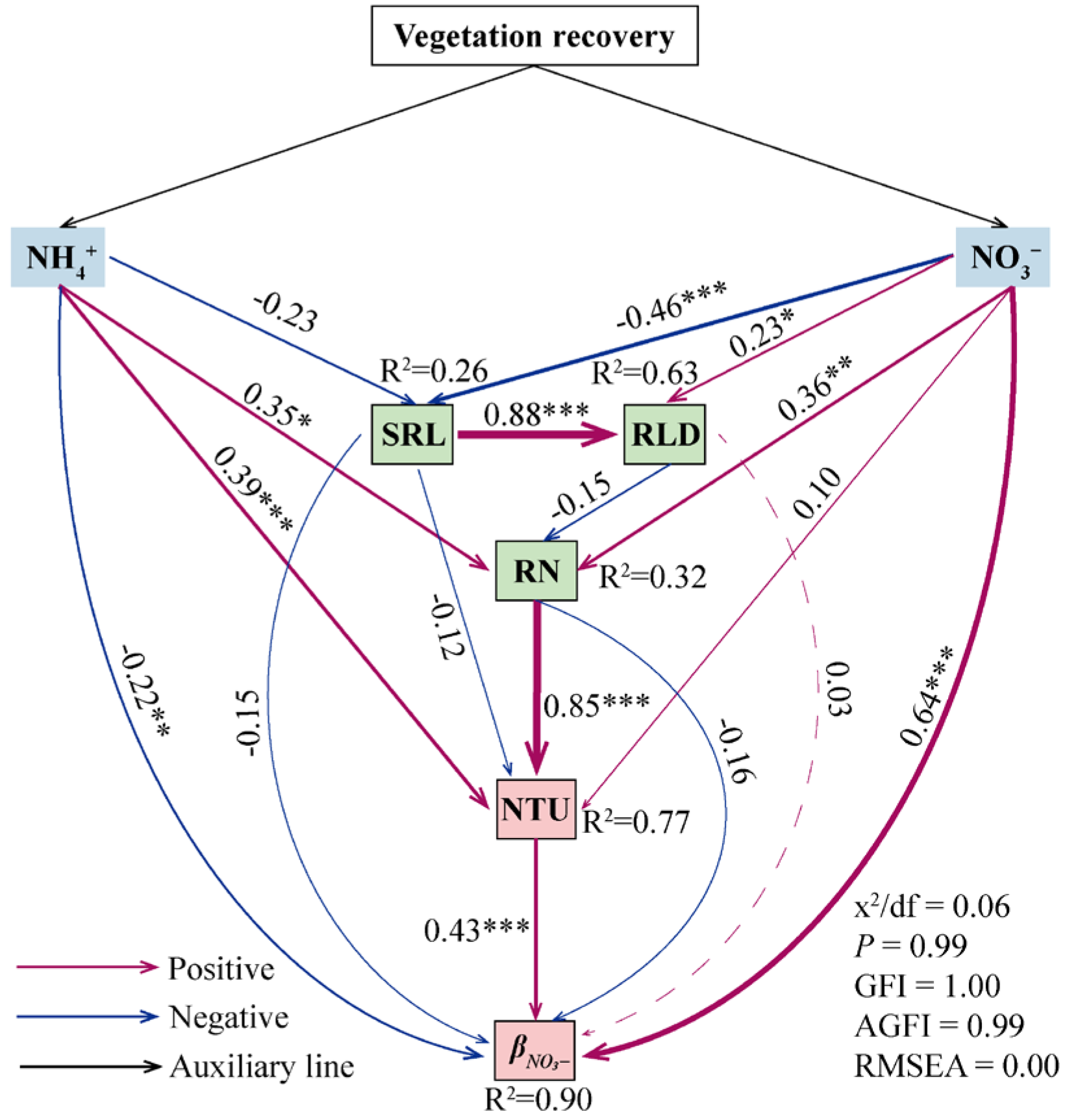
3.4. Drivers of the Nitrogen Uptake Rate and Ratio of Dominant Plants
4. Discussion
4.1. Plant Nitrogen Uptake Strategies During Vegetation Recovery
4.2. Relationship Between Plant Root Functional Traits and Plant Nitrogen Uptake Strategies During Vegetation Recovery
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.L.; Zhao, Y.; Xiao, D.; Xu, Z.H.; Zhang, W.; Xiao, J.; Wang, K.L. Dynamics of soil nitrogen availability following vegetation restoration along a climatic gradient of a subtropical karst region in China. J. Soil Sediments 2021, 21, 2167–2178. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 367–382. [Google Scholar] [CrossRef]
- Elrys, A.S.; Chen, Z.X.; Wang, J.; Uwiragiye, Y.; Helmy, A.M.; Desoky, E.S.M.; Cheng, Y.; Zhang, J.B.; Cai, Z.C.; Müeller, C. Global patterns of soil gross immobilization of ammonium and nitrate in terrestrial ecosystems. Glob. Change Biol. 2022, 28, 4472–4488. [Google Scholar] [CrossRef]
- Liu, L.J.; Zhu, Q.L.; Yang, L.; Elrys, A.S.; Sun, J.F.; Ni, K.; Meng, L.; Zhu, T.B. Afforestation increases soil inorganic N supply capacity and lowers plant N limitation in subtropical karst areas. Geoderma 2024, 443, 116848. [Google Scholar] [CrossRef]
- Wen, D.N.; Huang, Y.Y.; Huang, Y.F.; Ding, N.N.; Ni, K.; Wang, H.; Elrys, A.S.; Meng, L.; Zhu, T.B.; Gessert, A.; et al. Karst rocky desertification restoration increases soil inorganic N supply to reduce plant N limitation. Catena 2024, 241, 108012. [Google Scholar] [CrossRef]
- Du, E.; Terrer, C.; Pellegrini, A.F.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; We, X.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Ågren, G.I.; Wetterstedt, J.Å.M.; Billberger, M.F.K. Nutrient limitation on terrestrial plant growth–modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef]
- Lan, J.C.; Hu, N.; Fu, W.L. Soil carbon-nitrogen coupled accumulation following the natural vegetation restoration of abandoned farmlands in a karst rocky desertification region. Ecol. Eng. 2020, 158, 106033. [Google Scholar] [CrossRef]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef]
- Zhang, J.B.; Wang, J.; Müller, C.; Cai, Z.C. Ecological and practical significances of crop species preferential N uptake matching with soil N dynamics. Soil Boil. Biochem. 2016, 103, 63–70. [Google Scholar] [CrossRef]
- Zhou, X.L.; Wang, A.; Hobbie, E.A.; Zhu, F.F.; Qu, Y.Y.; Dai, L.M.; Li, D.J.; Liu, X.Y.; Zhu, W.X.; Koba, K.; et al. Mature conifers assimilate nitrate as efficiently as ammonium from soils in four forest plantations. New Phytol. 2021, 229, 3184–3194. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, 45. [Google Scholar] [CrossRef]
- Khokon, A.M.; Janz, D.; Polle, A. Ectomycorrhizal diversity, taxon-specific traits and root N uptake in temperate beech forests. New Phytol. 2023, 239, 739–751. [Google Scholar] [CrossRef]
- Yi, R.J.; Liu, Q.Y.; Yang, F.T.; Dai, X.Q.; Meng, S.W.; Fu, X.L.; Li, S.G.; Kou, L.; Wang, H.M. Complementary belowground strategies underlie species coexistence in an early successional forest. New Phytol. 2023, 238, 612–623. [Google Scholar] [CrossRef]
- Bergmann, J.; Weigelt, A.; Plas, F.V.W.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.R.; Valverde-Barrantes, O.J.; Bruelheide, H.; Freschet, G.T.; Iversen, C.M.; et al. The fungal collaboration gradient dominates the root economics space in plants. Sci. Adv. 2020, 6, eaba3756. [Google Scholar] [CrossRef]
- Guo, W.J.; Zhang, Z.L.; Liu, Q.; Xiao, J.; Yin, H.J. Seasonal variations in plant nitrogen acquisition in an ectomycorrhizal alpine forest on the eastern Tibetan Plateau, China. Plant Soil 2021, 459, 79–91. [Google Scholar] [CrossRef]
- Yan, H.; Freschet, G.T.; Wang, H.; Hogan, J.A.; Li, S.; Valverde-Barrantes, O.J.; Fu, X.L.; Wang, R.L.; Dai, X.Q.; Jiang, L.; et al. Mycorrhizal symbiosis pathway and edaphic fertility frame root economics space among tree species. New Phytol. 2022, 234, 1639–1653. [Google Scholar] [CrossRef]
- Xie, L.L.; Hu, X.F.; Li, W.T.; Liu, Q.H.; Yin, C.Y. Plant-plant interactions affect seasonal nitrogen uptake of subalpine conifer seedlings by altering root traits and soil nitrogen availabilities. Physiol. Plantarum. 2024, 176, e14204. [Google Scholar] [CrossRef]
- Ito, T.; Tanaka-Oda, A.; Masumoto, T.; Akatsuki, M.; Makita, N. Different relationships of fine root traits with root ammonium and nitrate uptake rates in conifer forests. J. Forestry Res. 2023, 28, 25–32. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Li, N.; Xiao, J.; Zhao, C.Z.; Zou, T.T.; Li, D.D.; Liu, Q.; Yin, H.J. Changes in plant nitrogen acquisition strategies during the restoration of spruce plantations on the eastern Tibetan Plateau, China. Soil Boil. Biochem. 2018, 119, 50–58. [Google Scholar] [CrossRef]
- Ding, J.X.; Kong, D.L.; Zhang, Z.L.; Cai, Q.; Xiao, J.; Liu, Q.; Yin, H.J. Climate and soil nutrients differentially drive multidimensional fine root traits in ectomycorrhizal-dominated alpine coniferous forests. J. Ecol. 2020, 108, 2544–2556. [Google Scholar] [CrossRef]
- White, P.J.; George, T.S.; Gregory, P.J.; Bengough, A.G.; Hallett, P.D.; McKenzie, B.M. Matching roots to their environment. Ann. Bot. 2013, 112, 207–222. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenotypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef]
- Liu, M.; Xu, X.L.; Yang, B.; Zhang, N.L.; Ma, Z.Q.; van Dam, N.M.; Bruelheide, H. Niche partitioning in nitrogen uptake among subtropical tree species enhances biomass production. Sci. Total Environ. 2022, 823, 153716. [Google Scholar] [CrossRef]
- Wang, K.L.; Zhang, C.H.; Chen, H.S.; Yue, Y.M.; Zhang, W.; Zhang, M.Y.; Qi, X.K.; Fu, Z.Y. Karst landscapes of China: Patterns, ecosystem processes and services. Landsc. Ecol. Eng. 2019, 34, 2743–2763. [Google Scholar] [CrossRef]
- Hu, C.P.; Liu, Z.Q.; Xiong, K.N.; Lyu, X.X.; Li, Y.; Zhang, R.K. Temporal and Spatial Variations in Carbon/Nitrogen Output in the Karst Critical Zone and Its Response to the Forest Ecosystem of Karst Desertification Control. Forests 2023, 14, 1121. [Google Scholar] [CrossRef]
- Hu, G.; Pang, Q.L.; Hu, C.; Xu, C.H.; Zhang, Z.H.; Zhong, C.F. Beta diversity patterns and determinants among vertical layers of tropical seasonal rainforest in karst peak-cluster depressions. Forests 2024, 15, 365. [Google Scholar] [CrossRef]
- Tong, X.W.; Brandt, M.; Yue, Y.M.; Ciais, P.; Rudbeck, J.M.; Penuelas, J.; Wigneron, J.; Xiao, X.M.; Song, X.P.; Horion, S.; et al. Forest management in southern China generates short term extensive carbon sequestration. Nat. Commun. 2020, 11, 1191–1197. [Google Scholar] [CrossRef]
- Dobson, A.; Rowe, Z.; Berger, J.; Wholey, P.; Caro, T. Biodiversity loss due to more than climate change. Science 2021, 374, 699–700. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.H.; Cao, Y.Q.; Zhang, Z.F.; Zhou, L.W.; Li, X.Q. The Grain for Green Program Promotes Soil Organic Matter Accumulation and Improves Soil Fungal Diversity in the Southwestern Karst Region. Forests 2025, 16, 121. [Google Scholar] [CrossRef]
- Ma, T.S.; Deng, X.W.; Chen, L.; Xiang, W.H. The soil properties and their effects on plant diversity in different degrees of rocky desertification. Sci. Total Environ. 2020, 736, 139667. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.D.; Dai, Q.H.; Ding, G.J.; Shi, D.M.; Li, C.L. Impact of vegetation restoration on soil properties in near-surface fissures located in karst rocky desertification regions. Soil Till. Res. 2020, 200, 104620. [Google Scholar] [CrossRef]
- Xiao, J.; Xiong, K.N. A review of agroforestry ecosystem services and its enlightenment on the ecosystem improvement of rocky desertification control. Sci. Total Environ. 2022, 852, 158538. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.J.; Li, D.J.; Chen, H.S.; Wang, K.L. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Song, M.; He, T.G.; Chen, H.; Wang, K.L.; Li, D.J. Dynamics of soil gross nitrogen transformations during post-agricultural succession in a subtropical karst region. Geoderma 2019, 341, 1–9. [Google Scholar] [CrossRef]
- Liang, J.H.; Cui, X.D.; Wen, L.Y.; Liu, D.; Yi, C.X.; Huang, K.Z.; Wang, J. Comparison of soil calcium and magnesium fractions transport in classic karst and non-karst region, Guilin. Carsologica Sin. 2022, 41, 220–227. (In Chinese) [Google Scholar]
- Rowley, M.C.; Grand, S.; Verrecchia, E.P. Calcium-mediated stabilization of soil organic carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef]
- Zhu, X.A.; Shen, Y.X.; Yuan, X.; Yuan, C.; Jin, L.Y.; Zhao, Z.M.; Chen, F.J.; Yang, B.; Jiang, X.J.; Liu, W.J. High levels of soil calcium and clay facilitate the recovery and stability of organic carbon: Insights from different land uses in the karst of China. Environ. Sci. Pollut. R. 2024, 31, 34234–34248. [Google Scholar] [CrossRef]
- Yang, H.; Zhu, T.B.; Wu, X.; Wu, H.Y.; Tang, W.; Lan, G.Y.; Christoph, M. Effect of sugar orange short-term planting on soil nitrogen conversion process in karst area. Carsologica Sin. 2023, 42, 52–60. (In Chinese) [Google Scholar]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Li, D.J.; Wen, L.; Xiao, K.C.; Song, T.Q.; Wang, K.L. Responses of soil gross nitrogen transformations to three vegetation restoration strategies in a subtropical karst region. Land Degrad. Dev. 2021, 32, 2520–2527. [Google Scholar] [CrossRef]
- Lipson, D.A.; Raab, T.K.; Schmidt, S.K.; Monson, R.K. An empirical model of amino acid transformations in an alpine Soil. Soil Boil. Biochem. 2001, 33, 189–198. [Google Scholar] [CrossRef]
- Liu, M.; Li, C.C.; Xu, X.L.; Wanek, W.; Jiang, N.; Wang, H.M.; Yang, X.D. Organic and inorganic nitrogen uptake by 21 dominant tree species in temperate and tropical forests. Tree Physiol. 2017, 37, 1515–1526. [Google Scholar] [CrossRef]
- Liu, Q.Y.; Wang, H.M.; Xu, X.L. Root nitrogen acquisition strategy of trees and understory species in a subtropical pine plantation in southern China. Eur. J. Forest Res. 2020, 139, 791–804. [Google Scholar] [CrossRef]
- Li, C.C.; Li, Q.R.; Qiao, N.; Xu, X.L.; Li, Q.K.; Wang, H.M. Inorganic and organic nitrogen uptake by nine dominant subtropical tree species. iForest 2015, 9, 253. [Google Scholar] [CrossRef]
- Yuan, X.Y.; She, W.W.; Guo, Y.P.; Qiao, Y.G.; Liu, L.; Song, C.Y.; Qin, S.G.; Zhang, Y.Q. Linkage between plant nitrogen preference and rhizosphere effects on soil nitrogen transformation reveals a plant resource adaptive strategies in nitrogen-limited soils. Plant Soil 2025. [Google Scholar] [CrossRef]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Bukovská, P.; Sudová, R.; Jansa, J. Benefits in plant N uptake via the mycorrhizal pathway in ample soil moisture persist under severe drought. Soil Boil. Biochem. 2023, 187, 109220. [Google Scholar] [CrossRef]
- Guan, M.; Pan, X.C.; Sun, J.K.; Chen, J.X.; Kong, D.L.; Feng, Y.L. Nitrogen acquisition strategy and its effects on invasiveness of a subtropical invasive plant. Front. Plant Sci. 2023, 14, 1243849. [Google Scholar] [CrossRef]
- Reuter, R.; Ferlian, O.; Tarkka, M.; Eisenhauer, N.; Pritsch, K.; Simon, J. Tree species rather than type of mycorrhizal association drive inorganic and organic nitrogen acquisition in tree–tree interactions. Tree Physiol. 2021, 41, 2096–2108. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.; Gong, X.H.; Shu, Y.G. Effects of vegetation restoration in karst areas on soil nitrogen mineralisation. PeerJ 2024, 12, e18582. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, X.l.; Wanek, W.; Sun, J.; Bardgett, R.D.; Tian, Y.Q.; Cui, X.Y.; Jiang, L.L.; Ma, Z.Q.; Kuzyakov, Y.; et al. Nitrogen availability in soil controls uptake of different nitrogen forms by plants. New Phytol. 2025, 245, 1450–1467. [Google Scholar] [CrossRef]
- Elrys, A.S.; Wang, J.; Metwally, M.A.; Cheng, Y.; Zhang, J.B.; Cai, Z.C.; Chang, S.X.; Müller, C. Global gross nitrification rates are dominantly driven by soil carbon-to-nitrogen stoichiometry and total nitrogen. Glob. Change Biol. 2021, 27, 6512–6524. [Google Scholar] [CrossRef]
- Li, D.J.; Yang, Y.; Chen, H.; Xiao, K.C.; Song, T.Q.; Wang, K.L. Soil gross nitrogen transformations in typical karst and nonkarst forests, southwest China. J. Geophys. Res. Biogeo. 2017, 122, 2831–2840. [Google Scholar] [CrossRef]
- Ren, H.; Gao, G.; Ma, Y.; Li, Z.; Wang, S.; Gu, J. Shift of root nitrogen-acquisition strategy with tree age is mediated by root functional traits along the collaboration gradient of the root economics space. Tree Physiol. 2023, 43, 1341–1353. [Google Scholar] [CrossRef]
- Liu, Q.Y.; Chen, Y.X.; Chen, Y.M. Nitrogen acquisition strategy shifts with tree age depending on root functional traits and soil properties in Larix principis-rupprechtii plantations. Front. Plant Sci. 2024, 15, 1358367. [Google Scholar] [CrossRef]
- Weemstra, M.; Mommer, L.; Visser, E.J.W.; van Ruijven, J.; Kuyper, T.W.; Mohren, G.M.J.; Sterck, F.J. Towards a multidimensional root trait framework: A tree root review. New Phytol. 2016, 211, 1159–1169. [Google Scholar] [CrossRef]
- Ma, Z.Q.; Guo, D.L.; Xu, X.L.; Lu, M.Z.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef]
- Lu, B.H.; Qian, J.; Hu, J.; Wang, P.F.; Jin, W.; Tang, S.J.; He, Y.X.; Zhang, C. The role of fine root morphology in nitrogen uptake by riparian plants. Plant Soil 2022, 472, 527–542. [Google Scholar] [CrossRef]
- Stefan, W.; Peter, R.; Edwards, P.J. Phenotypic plasticity of grass root anatomy in response to light intensity and nutrient supply. Ann. Bot. 2001, 88, 1071–1078. [Google Scholar]
- Ficken, C.D.; Wright, J.P. Nitrogen uptake and biomass resprouting show contrasting relationships with resource acquisitive and conservative plant traits. J. Veg. Sci. 2019, 30, 65–74. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, J.; Gong, L. The morphological and chemical properties of fine roots respond to nitrogen addition in a temperate Schrenk’s spruce (Picea schrenkiana) forest. Sci. Rep. 2021, 11, 3839. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recovery Stages | Longitude and Latitude | Altitude (m) | Slope (°) | Soil Depth (cm) | Interference Conditions |
|---|---|---|---|---|---|
| Grassland | 110°32′ E 25°12′ N | 324–341 | 2–5 | 0–30 | Abandoned, no interference for 3–5 years |
| Shrub–grassland | 110°32′ E 25°12′ N | 322–348 | 4–8 | 0–30 | Abandoned, no interference for 7–10 years |
| Shrubland | 110°53′ E 25°21′ N | 321–343 | 12–15 | 0–25 | Abandoned, no interference for 30–35 years |
| Woodland | 110°52′ E 25°20′ N | 246–300 | 20–25 | 0-20 | Abandoned, no interference for 70–80 years |
| Dominant Plants | Recovery Stages | NH4+ (mg kg−1) | NO3− (mg kg−1) | Glycine (mg kg−1) | TFAA (mg kg−1) A | Inorganic N (mg kg−1) | NH4+/NO3− ratio |
|---|---|---|---|---|---|---|---|
| Miscanthus floridulus | Grassland | 8.51 ± 5.72 de | 5.92 ± 2.88 d | 0.03 ± 0.01 b | 0.59 ± 0.12 ab | 14.4 ± 7.62 de | 1.52 ± 1.04 bc |
| Miscanthus floridulus | Shrub–grassland | 5.83 ± 1.00 e | 5.71 ± 3.05 d | 0.06 ± 0.02 a | 0.81 ± 0.10 a | 11.5 ± 3.47 e | 1.31 ± 0.72 cd |
| Alchornea trewioides | 29.3 ± 4.43 a | 13.3 ± 2.92 d | 0.04 ± 0.02 ab | 0.56 ± 0.39 abc | 42.6 ± 3.94 ab | 2.32 ± 0.85 ba | |
| Bauhiniachampionii | 19.8 ± 3.91 b | 6.20 ± 2.70 d | 0.04 ± 0.01 ab | 0.44 ± 0.09 bc | 26.0 ± 6.05 cd | 3.62 ± 1.24 a | |
| Loropetalum chinense | Shrubland | 8.84 ± 4.12 de | 34.6 ± 5.27 ab | 0.03 ± 0.01 b | 0.27 ± 0.11 c | 43.4 ± 9.35 ab | 0.24 ± 0.09 e |
| Bauhinia championii | 12.7 ± 5.09 cd | 26.0 ± 12.1 bc | 0.03 ± 0.01 b | 0.27 ± 0.26 c | 38.8 ± 16.7 bc | 0.52 ± 0.16 de | |
| Cyclobalanopsis glauca | Woodland | 15.3 ± 3.44 bc | 39.2 ± 13.1 a | 0.03 ± 0.00 b | 0.31 ± 0.12 bc | 54.6 ± 15.4 a | 0.43 ± 0.17 de |
| Mallotus philippinensis | 15.0 ± 4.40 bc | 24.0 ± 8.54 c | 0.04 ± 0.02 ab | 0.40 ± 0.20 bc | 39.0 ± 8.79 bc | 0.70 ± 0.38 ced |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Liu, L.; Yang, H.; Yang, S.; Zhu, T.; Meng, L.; Wen, D. Changes in Plant Nitrogen Uptake Strategies Following Vegetation Recovery in Karst Regions. Forests 2025, 16, 839. https://doi.org/10.3390/f16050839
Yang L, Liu L, Yang H, Yang S, Zhu T, Meng L, Wen D. Changes in Plant Nitrogen Uptake Strategies Following Vegetation Recovery in Karst Regions. Forests. 2025; 16(5):839. https://doi.org/10.3390/f16050839
Chicago/Turabian StyleYang, Lin, Lijun Liu, Hui Yang, Shuting Yang, Tongbin Zhu, Lei Meng, and Dongni Wen. 2025. "Changes in Plant Nitrogen Uptake Strategies Following Vegetation Recovery in Karst Regions" Forests 16, no. 5: 839. https://doi.org/10.3390/f16050839
APA StyleYang, L., Liu, L., Yang, H., Yang, S., Zhu, T., Meng, L., & Wen, D. (2025). Changes in Plant Nitrogen Uptake Strategies Following Vegetation Recovery in Karst Regions. Forests, 16(5), 839. https://doi.org/10.3390/f16050839