
Effects of Plant Communities in Urban Green Spaces on Microclimate and Thermal Comfort

Abstract
1. Introduction
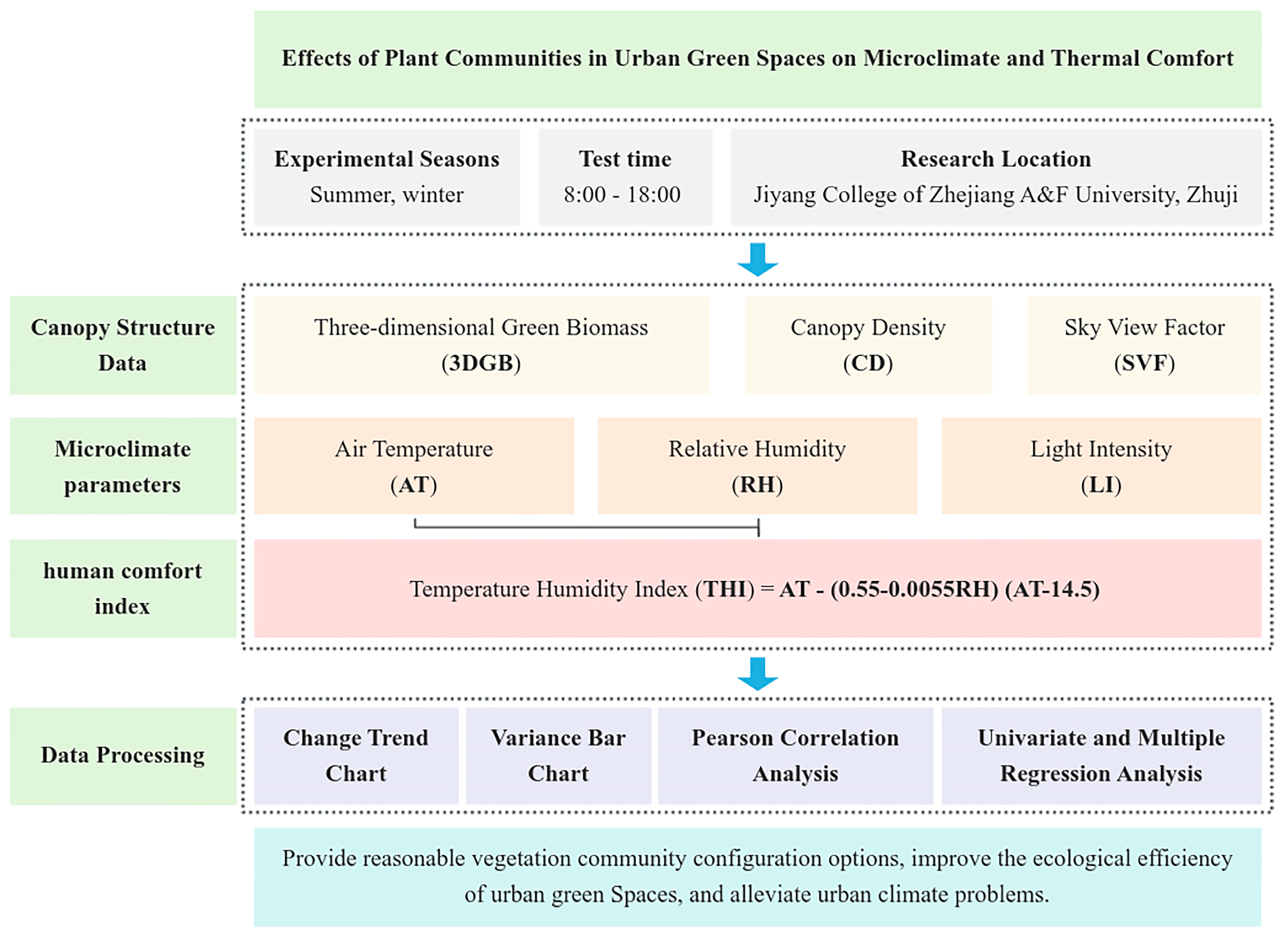
2. Materials and Methods
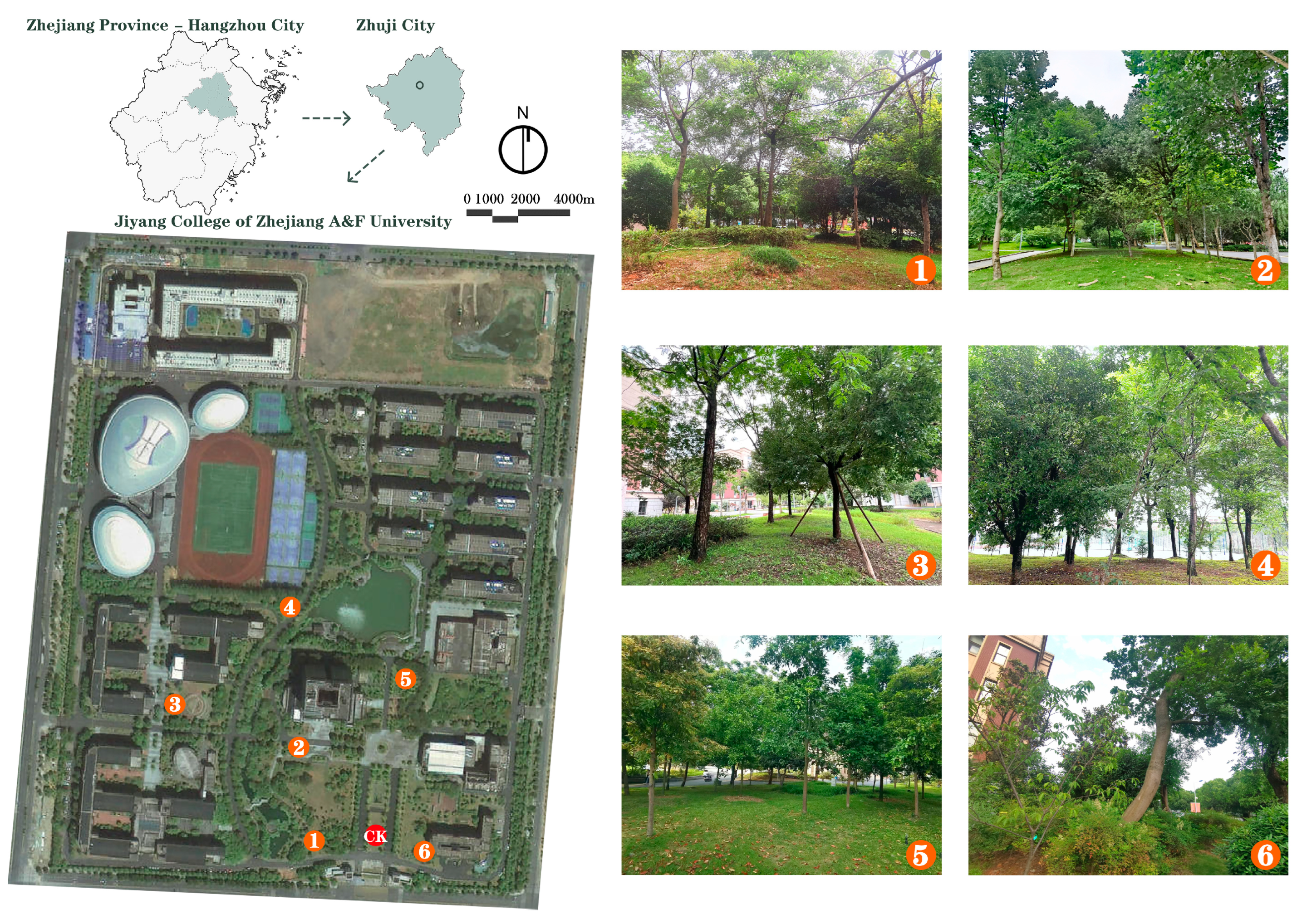
2.1. Study Area and Measurement Sites
2.2. Methods
2.3. Measurement of Microclimatic Parameters and Canopy Structure Indices
2.4. Thermal Comfort Index
2.5. Data Processing and Analysis
3. Results and Analysis
3.1. Quantitative Analysis of Microclimatic Characteristics
3.1.1. Air Temperature
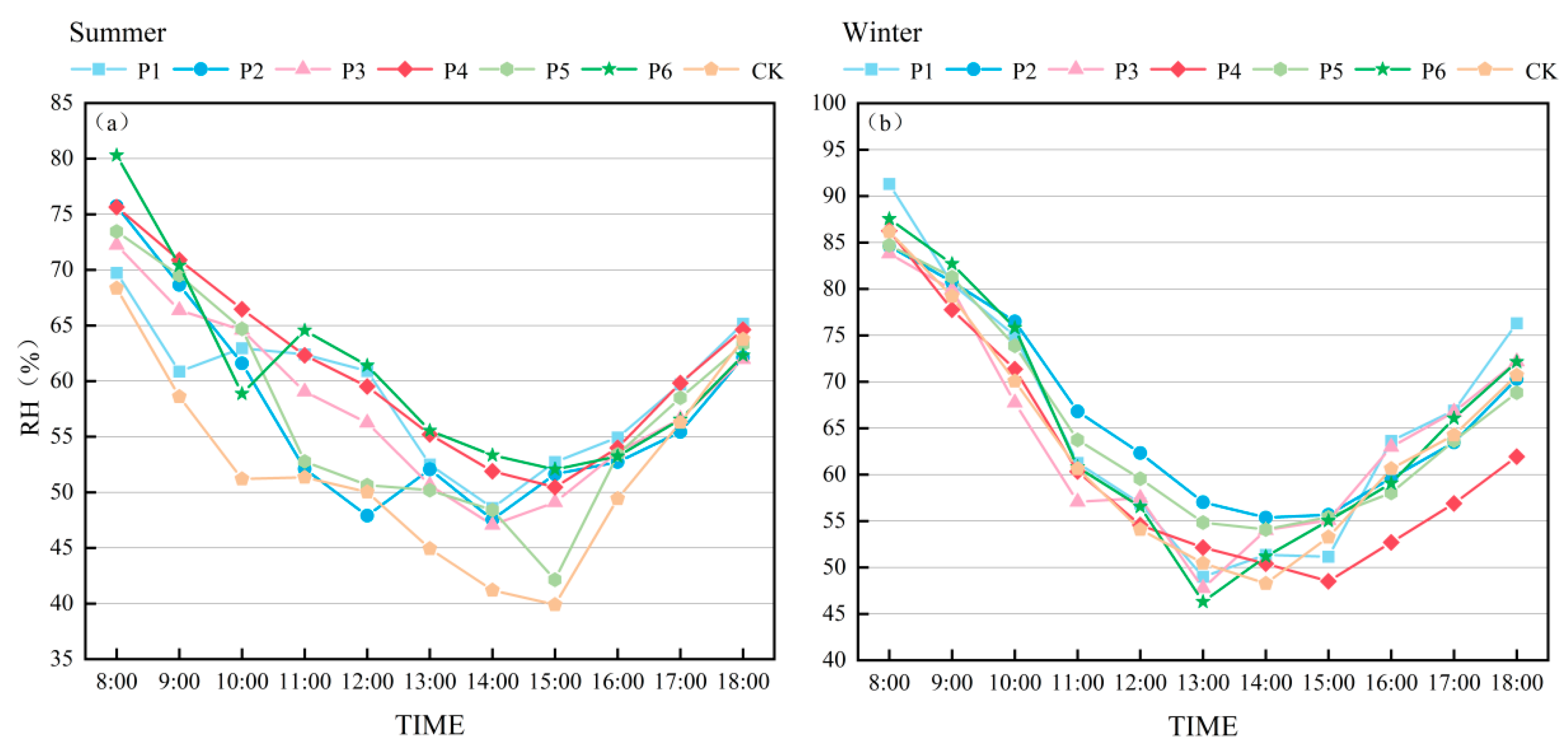
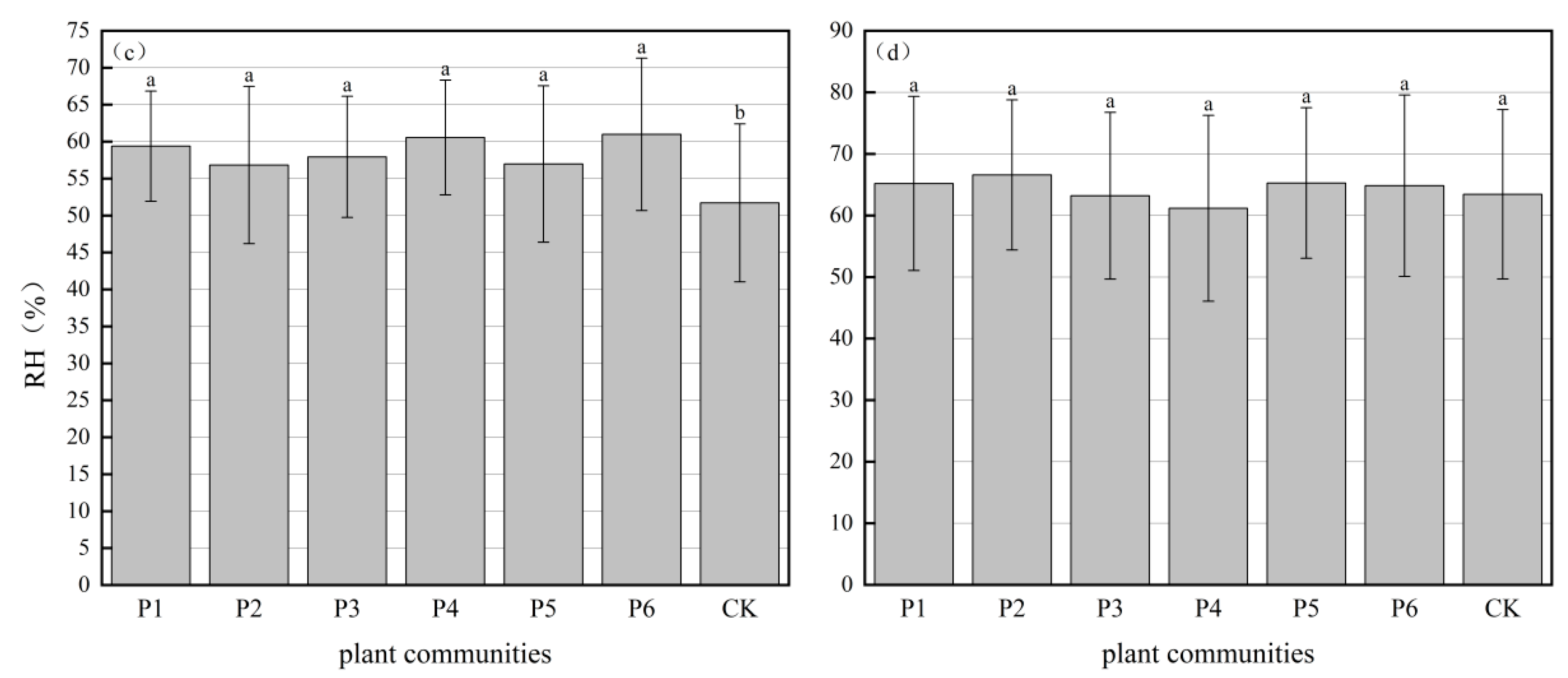
3.1.2. Relative Humidity
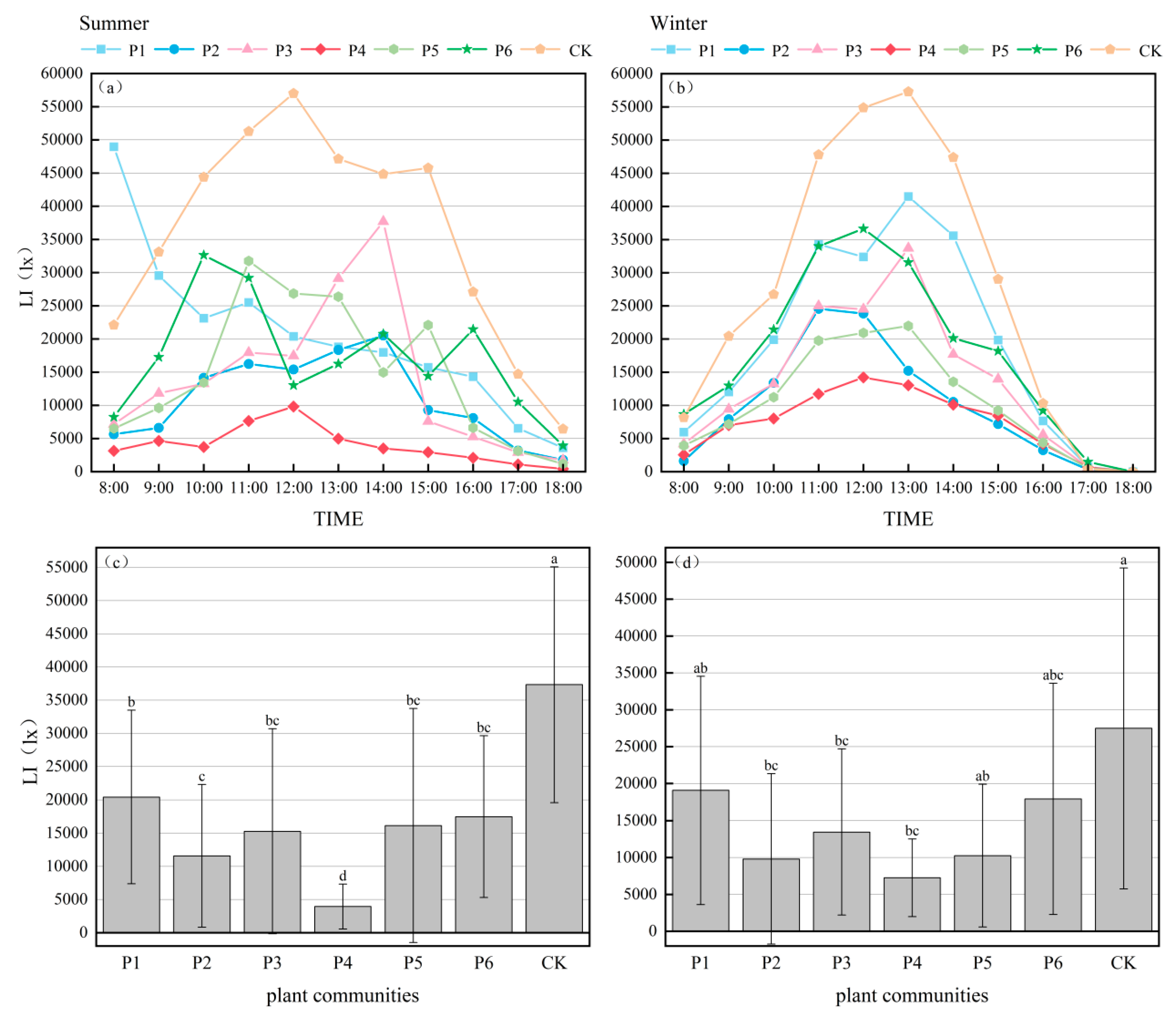
3.1.3. Light Intensity
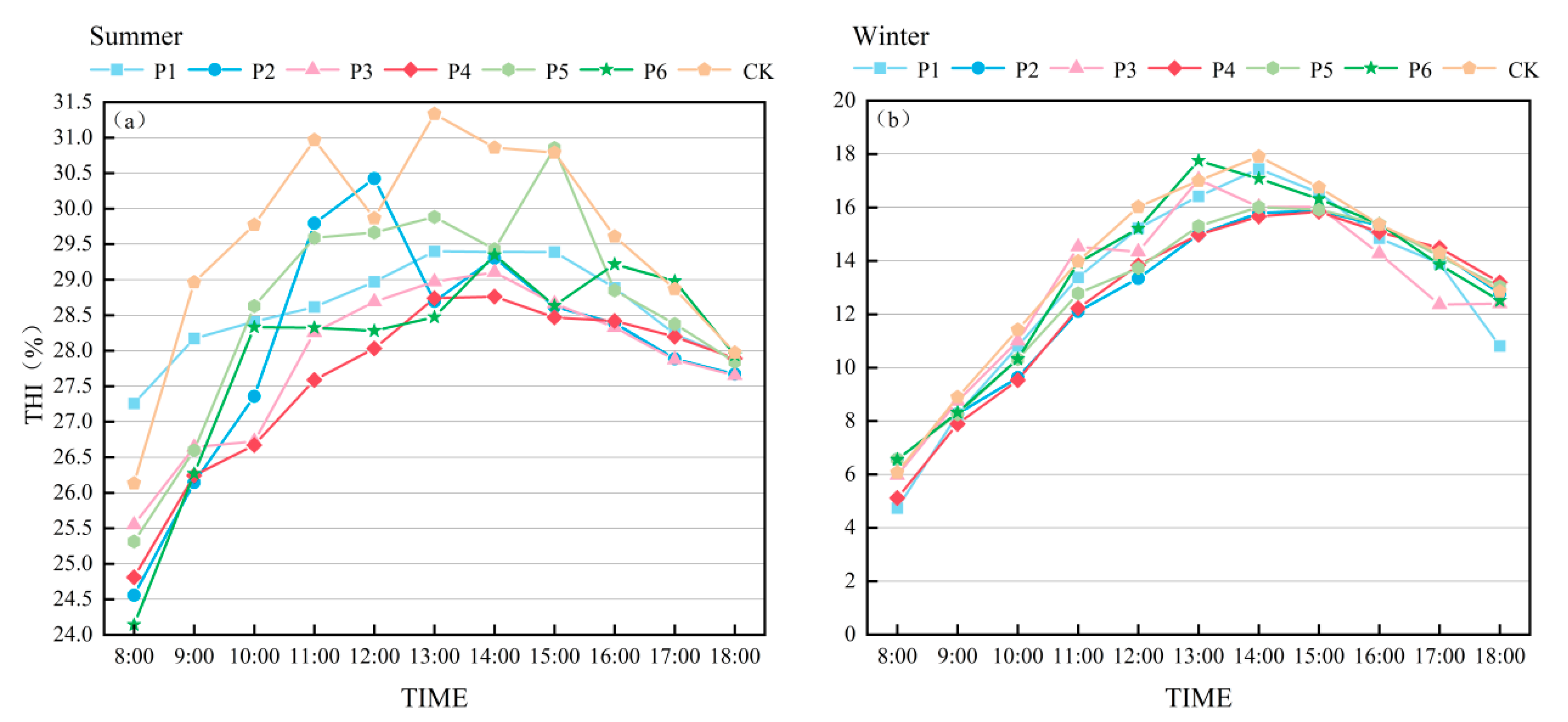
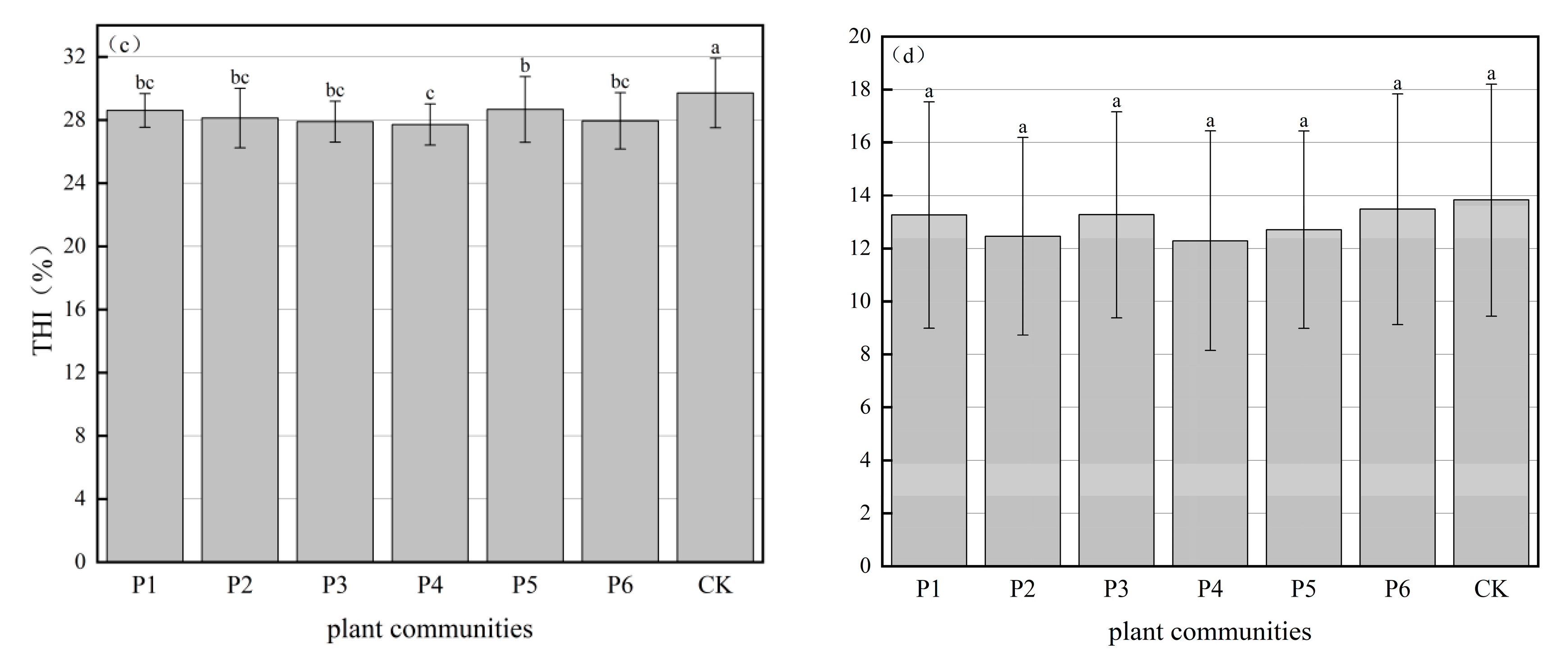
3.2. THI of Experimental Sample Sites
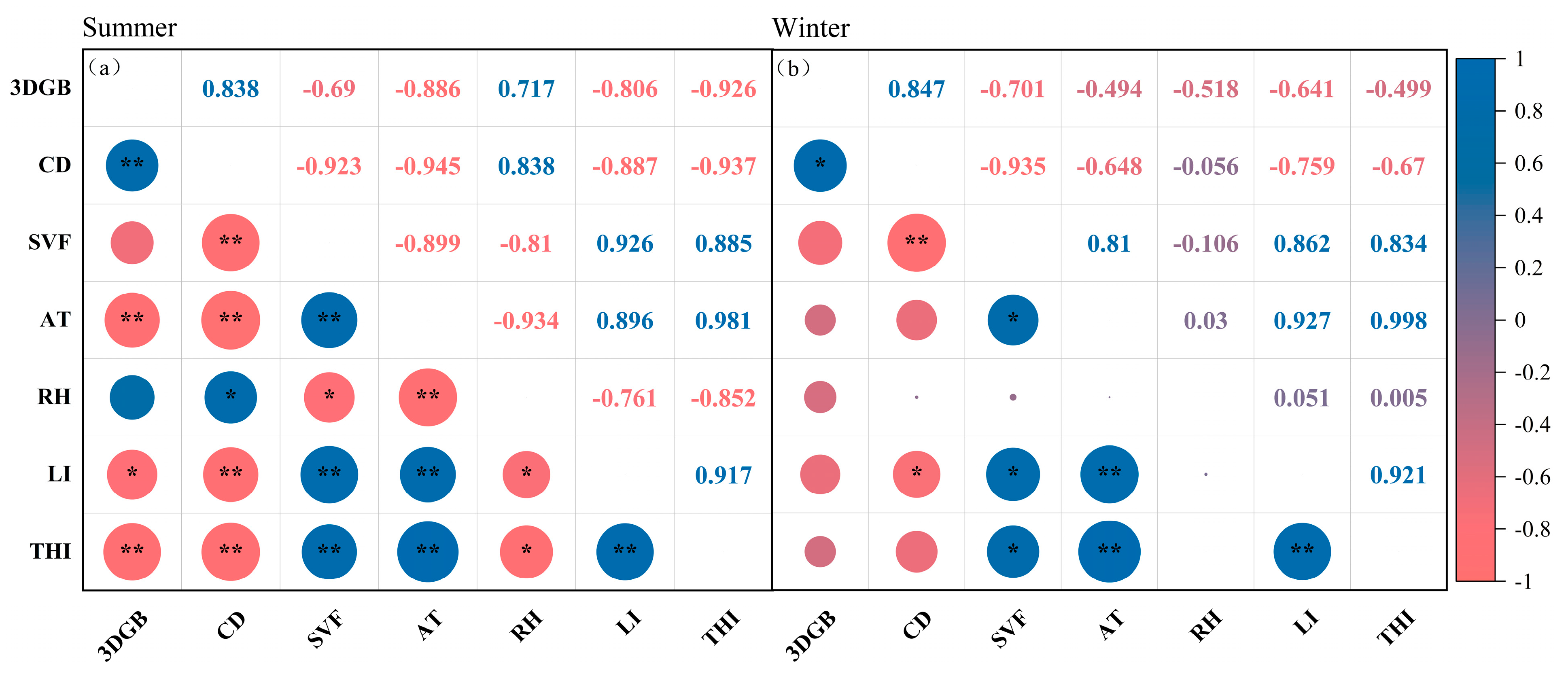
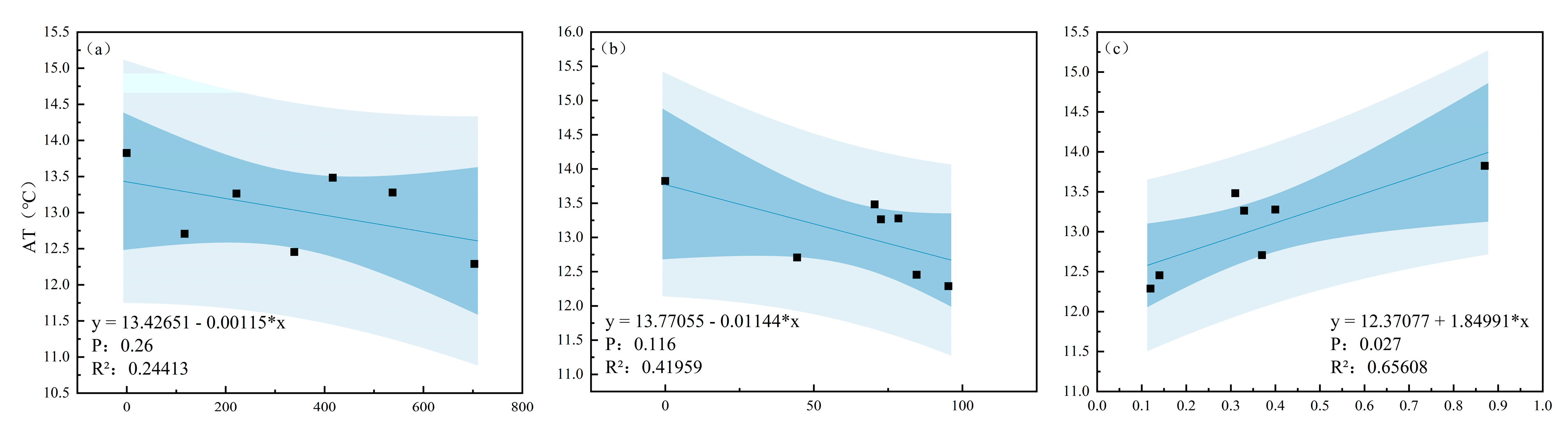
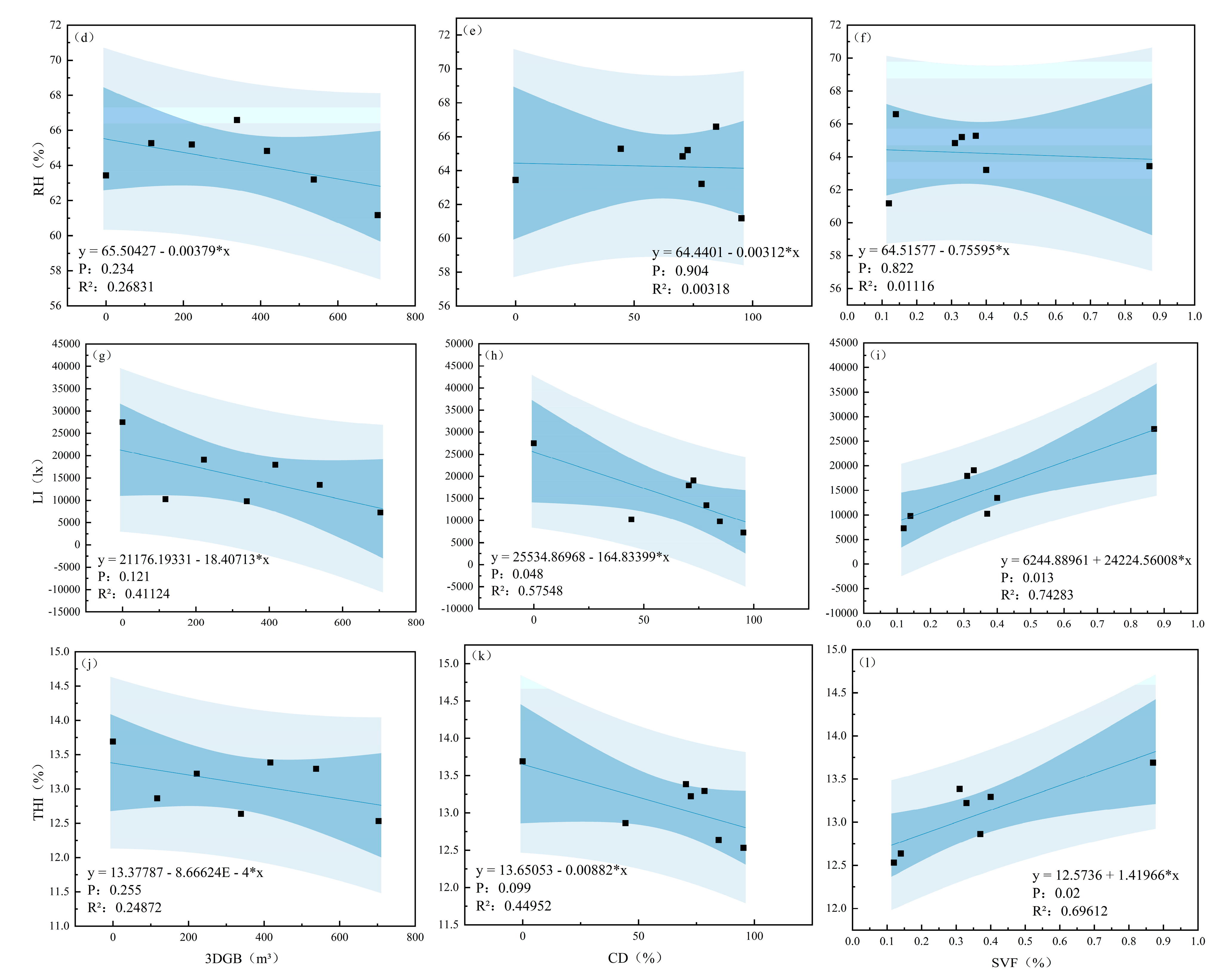
3.3. Relationships Between Microclimate Factors, THI, and Canopy Structural Indices
4. Discussion
4.1. Microclimate Parameter Comparison
4.2. Human Thermal Comfort Comparison
4.3. Effect of Canopy Structure on Microclimate and Thermal Comfort
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3DGB | Three-dimensional Green Biomass |
| CD | Canopy Density |
| SVF | Sky-View Factor |
| AT | Air Temperature |
| RH | Relative Humidity |
| LI | Light Intensity |
| THI | Temperature–Humidity Index |
| WS | Wind Speed |
| LAI | Leaf Area Index |
| ET | Evergreen Tree |
| DT | Deciduous Tree |
References
- Shevchenko, O.; Snizhko, S.; Matzarakis, A. Recent trends on human thermal bioclimate conditions in Kyiv, Ukraine. Geogr. Pol. 2020, 93, 89–106. [Google Scholar] [CrossRef]
- Field, C.; Barros, V.; Mach, K.; Mastrandrea, M.; Yohe, G. Climate Change 2014: Impacts Adaptation & Vulnerability; Technical Summary of the IPCC Working Group II AR5 Report; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Akbari, H.; Pomerantz, M.; Taha, H. Cool surfaces and shade trees to reduce energy use and improve air quality in urban areas. Sol. Energy 2001, 70, 295–310. [Google Scholar] [CrossRef]
- Grimmond, S.U.E. Urbanization and global environmental change: Local effects of urban warming. Geogr. J. 2007, 173, 83–88. [Google Scholar] [CrossRef]
- Park, M.; Hagishima, A.; Tanimoto, J.; Narita, K.I. Effect of urban vegetation on outdoor thermal environment: Field measurement at a scale model site. Build. Environ. 2012, 56, 38–46. [Google Scholar] [CrossRef]
- Renterghem, T.V.; Botteldooren, D.; Verheyen, K. Road traffic noise shielding by vegetation belts of limited depth. J. Sound Vib. 2012, 331, 2404–2425. [Google Scholar] [CrossRef]
- Jim, C.Y.; Chen, W.Y. Assessing the ecosystem service of air pollutant removal by urban trees in Guangzhou (China)—Science Direct. J. Environ. Manag. 2008, 88, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Armson, D.; Stringer, P.; Ennos, A.R. The effect of tree shade and grass on surface and globe temperatures in an urban area. Urban For. Urban Green. 2012, 11, 245–255. [Google Scholar] [CrossRef]
- Wu, Y.; Patuano, A.; Mashhoodi, B.; Lenzholzer, S.; Acred, A.; Zertuche, L.N. How small green spaces cool urban neighbourhoods: Optimising distribution, size and shape. Landsc. Urban Plan. 2025, 253, 105224. [Google Scholar] [CrossRef]
- Ariani, R.; Fitri, R.; Wisdianti, D. Designing Pocket Park at Dense Building Area with Micro Climate Approach. Sustain. Civ. Build. Manag. Eng. J. 2022, 1, 1–9. [Google Scholar] [CrossRef]
- Ma, D. Cooling Effect of Pocket Parks in Urban Areas. Environ. Sci. Pollut. Res. 2023, 30, 23135–23154. [Google Scholar] [CrossRef]
- Rui, L.; Buccolieri, R.; Gao, Z.; Gatto, E.; Ding, W. Study of the effect of green quantity and structure on thermal comfort and air quality in an urban-like residential district by ENVI-met modelling. Build. Simul. 2019, 12, 183–194. [Google Scholar] [CrossRef]
- Lu, F.; Gao, Y.; Jiang, L.; Chen, Y.; Hao, Z. The effect of greening layout on microclimate in urban residential areas in hot summer—Cold winter zones. Atmosphere 2023, 14, 1824. [Google Scholar] [CrossRef]
- Rasul, N.; Motesaddi, S.Z.; Mohammad, B.; Abdollah, A. Design a protocol to investigate the effects of climate change in vivo. Environ. Res. 2022, 212, 113482. [Google Scholar]
- Antoniadis, D.; Katsoulas, N.; Papanastasiou, D.; Christidou, V.; Kittas, C. Evaluation of thermal perception in schoolyards under Mediterranean climate conditions. Int. J. Biometeorol. 2016, 60, 319–334. [Google Scholar] [CrossRef]
- Öğüztürk, G.E.; Sünbül, S.; Alparslan, C. The effect of green areas on urban microclimate: A university campus model case. Appl. Sci. 2025, 15, 4358. [Google Scholar] [CrossRef]
- Antoniadis, D.; Katsoulas, N.; Papanastasiou, D.K. Thermal Environment of Urban Schoolyards: Current and Future Design with Respect to Children’s Thermal Comfort. Atmosphere 2020, 11, 1144. [Google Scholar] [CrossRef]
- Zhang, S.; Li, S.; Shu, L.; Xiao, T.; Shui, T. Landscape Configuration Effects on Outdoor Thermal Comfort across Campus—A Case Study. Atmosphere 2023, 14, 270. [Google Scholar] [CrossRef]
- Ghaffarianhoseini, A.; Berardi, U.; Ghaffarianhoseini, A.; Al-Obaidi, K. Analyzing the thermal comfort conditions of outdoor spaces in a university campus in Kuala Lumpur, Malaysia. Sci. Total Environ. 2019, 666, 1327–1345. [Google Scholar] [CrossRef]
- Nastos, P.T.; Polychroni, I.D. Modeling and in situ measurements of biometeorological conditions in microenvironments within the Athens University Campus, Greece. Int. J. Biometeorol. 2016, 8, 1137. [Google Scholar] [CrossRef]
- Singh, M.; Acharya, N.; Jamshidi, S.; Jiao, J.; Yang, Z.L.; Coudert, M.; Baumer, Z.; Niyogi, D. Urban precipitation downscaling using deep learning: A smart city application over Austin, Texas, USA. arXiv 2022, arXiv:2209.06848. [Google Scholar]
- Song, Y.; Newman, G.D.; Huang, A.; Ye, X. Factors influencing long-term city park visitations for mid-sized US cities: A big data study using smartphone user mobility. Sustain. Cities Soc. 2022, 80, 103815. [Google Scholar] [CrossRef]
- Yang, Y.; Gatto, E.; Gao, Z.; Buccolieri, R.; Morakinyo, T.E.; Lan, H. The ‘plant evaluation model’ for the assessment of the impact of vegetation on outdoor microclimate in the urban environment. Build. Environ. 2019, 159, 106151.1–106151.13. [Google Scholar] [CrossRef]
- Streiling, S.; Matzarakis, A. Influence of single and small clusters of trees on the bioclimate of a city: A case study. J. Arboric. 2003, 29, 309–316. [Google Scholar] [CrossRef]
- Lin, B.S.; Lin, Y.J. Cooling Effect of Shade Trees with Different Characteristics in a Subtropical Urban Park. HortScience 2010, 45, 83–86. [Google Scholar] [CrossRef]
- Yang, A.S.; Juan, Y.H.; Wen, C.Y.; Chang, C.J. Numerical simulation of cooling effect of vegetation enhancement in a subtropical urban park. Appl. Energy 2017, 192, 178–200. [Google Scholar] [CrossRef]
- Li, R.; Zeng, F.; Zhao, Y.; Wu, Y.; Niu, J.; Wang, L.L.; Gao, N.; Shi, X. CFD simulations of the tree effect on the outdoor microclimate by coupling the canopy energy balance model. Build. Environ. 2023, 230, 109995. [Google Scholar] [CrossRef]
- Kurn, D.M.; Bretz, S.E.; Huang, B.; Akbari, H. The Potential for Reducing Urban Air Temperatures and Energy Consumption Through Vegetative Cooling; Lawrence Berkeley National Laboratory (LBNL): Berkeley, CA, USA, 1994. [Google Scholar]
- Frenne, P.D.; Lenoir, J.; Luoto, M.; Scheffers, B.R.; Hylander, K. Forest microclimates and climate change: Importance, drivers and future research agenda. Glob. Change Biol. 2021, 27, 1109–1124. [Google Scholar] [CrossRef]
- Florian, Z.; Pieter, D.F.; Jonathan, L.; Pieter, V.; Kris, V.; Markus, B.R.; Lander, B.; Radim, H.; Imre, B.; Jrg, B. Forest microclimate dynamics drive plant responses to warming. Science 2020, 368, 772–775. [Google Scholar]
- Pieter, F.D.; Francisco, R.; Anthony, D.C.; Lander, B.; Gorik, V.; Mark, V.; Markus, B.; Brown, C.D.; Brunet, J.; Johnny, C. Microclimate moderates plant responses to macroclimate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18561–18565. [Google Scholar]
- Geiger, R. The Climate Near the Ground; Harvard University Press: Cambridge, MA, USA, 1957. [Google Scholar]
- Colter, K.R.; Middel, A.C.; Martin, C.A. Effects of natural and artificial shade on human thermal comfort in residential neighborhood parks of Phoenix, Arizona, USA. Urban For. Urban Green. 2019, 44, 126446. [Google Scholar] [CrossRef]
- Wang, R.; Chen, Q.; Wang, D.X. Effects of Altitude, Plant Communities, and Canopies on the Thermal Comfort, Negative Air Ions, and Airborne Particles of Mountain Forests in Summer. Sustainability 2022, 14, 3882. [Google Scholar] [CrossRef]
- Wu, H.; Yang, C.; Chen, J.; Yang, S.; Lu, T.; Lin, X. Effects of Green space landscape patterns on particulate matter in Zhejiang Province, China. Atmos. Pollut. Res. 2018, 9, 923–933. [Google Scholar] [CrossRef]
- Zaki, S.A.; Syahidah, S.W.; Shahidan, M.F.; Ahmad, M.I.; Yakub, F.; Hassan, M.Z.; Daud, M.Y.M. Assessment of Outdoor Air Temperature with Different Shaded Area within an Urban University Campus in Hot-Humid Climate. Sustainability 2020, 12, 5741. [Google Scholar] [CrossRef]
- Terschanski, J.; Nunes, H.M.; Aalto, I.; Pellikka, P.; Wekesa, C.; Maeda, E.E. The role of vegetation structural diversity in regulating the microclimate of human-modified tropical ecosystems. J. Environ. Manag. 2024, 360, 121128. [Google Scholar] [CrossRef] [PubMed]
- Teshnehdel, S.; Akbari, H.; Giuseppe, E.D.; Brown, R.D. Effect of tree cover and tree species on microclimate and pedestrian comfort in a residential district in Iran. Build. Environ. 2020, 178, 106899. [Google Scholar] [CrossRef]
- Fung, C.K.W.; Jim, C.Y. Microclimatic resilience of subtropical woodlands and urban-forest benefits—Science Direct. Urban For. Urban Green. 2019, 42, 100–112. [Google Scholar] [CrossRef]
- Speak, A.; Montagnani, L.; Wellstein, C.; Zerbe, S. The influence of tree traits on urban ground surface shade cooling. Landsc. Urban Plan. 2020, 197, 103748. [Google Scholar] [CrossRef]
- Potchter, O.; Cohen, P.; Lin, T.P.; Matzarakis, A. Outdoor human thermal perception in various climates: A comprehensive review of approaches, methods and quantification. Sci. Total Environ. 2018, 631–632, 390–406. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.G. An Analysis of Some Observations of Thermal Comfort in an Equatorial Climate. Br. J. Ind. Med. 1959, 16, 297–310. [Google Scholar] [CrossRef]
- Thom, E.C. The Discomfort Index. Weatherwise 1959, 12, 57–60. [Google Scholar] [CrossRef]
- Sahar, S.; Huiwen, Z.; Xiaoli, C.; Felix, M.; Li, H. The influence of spatial configuration of green areas on microclimate and thermal comfort. Urban For. Urban Green. 2018, 34, 85–96. [Google Scholar]
- Dong, H.L. Study on the microclimatic characteristics and human comfort of park plant communities in summer. Procedia Environ. Sci. 2012, 13, 755–765. [Google Scholar]
- Wu, Z.; Zohner, C.M.; Zhou, Y.; Crowther, T.W.; Wang, H.; Wang, Y.; Peuelas, J.; Gong, Y.; Zhang, J.; Zou, Y. Tree species composition governs urban phenological responses to warming. Nat. Commun. 2025, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gkatsopoulos, P. A methodology for calculating cooling from vegetation evapotranspiration for use in urban space microclimate simulations. Procedia Environ. Sci. 2017, 38, 477–484. [Google Scholar] [CrossRef]
- Kotzen, B. An investigation of shade under six different tree species of the Negev desert towards their potential use for enhancing micro-climatic conditions in landscape architectural development. J. Arid Environ. 2003, 55, 231–274. [Google Scholar] [CrossRef]
- Wei, R.; Xu, C.; Song, D.; Tong, H.; Chen, Z. Study on the Correlation Analysis between Urban Morphological Factors and Microclimate Based on Empirical Methods on a University Campus in a Hot-Summer–Cold-Winter Region. Buildings 2023, 13, 25. [Google Scholar] [CrossRef]
- Smith, W.K.; Vogelmann, T.C.; Schaberg, P.G. Clouds homogenize shoot temperatures, transpiration, and photosynthesis within crowns of Abies fraseri (Pursh.) Poiret. Oecologia 2017, 183, 673–683. [Google Scholar]
- Sun, S.; Xu, X.; Lao, Z.; Liu, W.; Li, Z.; Higueras Garcia, E.; He, L.; Zhu, J. Evaluating the impact of urban green space and landscape design parameters on thermal comfort in hot summer by numerical simulation. Build. Environ. 2017, 123, 277–288. [Google Scholar] [CrossRef]
- Zheng, S.L.; Guldmann, J.M.; Wang, Z.Y.; Qiu, Z.N.; He, C.W.; Wang, K. Experimental and theoretical study of urban tree instantaneous and hourly transpiration rates and their cooling effect in hot and humid area. Sustain. Cities Soc. 2021, 68, 102808. [Google Scholar] [CrossRef]
- Suwanmanee, N.; Kronprasert, N.; Sutheerakul, C.; Arunotayanun, K.; Rinchumphu, D. Investigation of outdoor thermal comfort for campus pedestrian walkways in Thailand. Sustainability 2023, 15, 12345. [Google Scholar] [CrossRef]
- Li, J.; Zhai, Z.; Ding, Y.; Ding, Y.; Deng, Y.; Ye, L.; Zhai, Z.; Li, H.; Chen, S. Effect of optimal allocation of urban trees on the outdoor thermal environment in hot and humid areas: A case study of a university campus in Guangzhou, China. Energy Build. 2023, 300, 113640. [Google Scholar] [CrossRef]
- Lin, W.; Zeng, C.C.; Nie, W.B.; Nan, X.G.; Shen, S.S.; Shi, Y.; Yan, H.; Yang, F.; Wu, R.W.; Bao, Z.Y. Study of the Vertical Structures, Thermal Comfort, Negative Air Ions, and Human Physiological Stress of Forest Walking Spaces in Summer. Forests 2022, 13, 335. [Google Scholar] [CrossRef]
- Fan, L.; Zhao, M.; Huo, J.; Sha, Y.; Zhou, Y. The Impact of Vegetation Layouts on Thermal Comfort in Urban Main Streets: A Case Study of Youth Street in Shenyang. Sustainability 2025, 17, 1755. [Google Scholar] [CrossRef]
- Abreu-Harbich, D.V.L.; Labaki, C.L.; Matzarakis, A. Effect of tree planting design and tree species on human thermal comfort in the tropics. Landsc. Urban Plan. 2015, 138, 99–109. [Google Scholar] [CrossRef]
- Yan, H.; Wu, F.; Dong, L. Influence of a large urban park on the local urban thermal environment. Sci. Total Environ. 2018, 622–623, 882–891. [Google Scholar] [CrossRef]
- Liu, Z.; Brown, R.D.; Zheng, S.; Jiang, Y.; Zhao, L. An In-Depth Analysis of the Effect of Trees on Human Energy Fluxes. Urban For. Urban Green. 2020, 50, 126646. [Google Scholar] [CrossRef]
- Karimi, A.; Sanaieian, H.; Farhadi, H.; Saeid, N.M. Evaluation of the thermal indices and thermal comfort improvement by different vegetation species and materials in a medium-sized urban park. Energy Rep. 2020, 6, 1670–1684. [Google Scholar] [CrossRef]
- Lu, T.R.; Jiang, Y.J.; Wu, Z.; Yan, M.; Qiu, J.; Wu, C. Transpiration characteristics of typical evergreen and deciduous tree species in the subtropical karst region and their responses to environmental factors. Acta Ecol. Sin. 2022, 42, 1047–1058. [Google Scholar]
- Shao, Y.C.; Li, J.J.; Fu, D.F.; Li, E.H.; Zhuang, J.R. Study on the Photosynthetic and Transpiration Characteristics of Eight Deciduous Broad-leaved Tree Species in Summer in Shanghai Area. J. Northwest For. Univ. 2015, 30, 30–38. [Google Scholar]
- Zhang, Y.P.; Zhou, G.M.; Zhou, Y.F.; Shi, Y.J.; Li, C.; Li, M.; Lv, Y.L.; He, Y. Photosynthetic Response Characteristics of Five Common Greening Tree Species to Extreme High Temperature. J. Zhejiang AF Univ. 2017, 34, 301–309. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Community Composition | Community Structure | 3DGB/m3 | CD /% | SVF/% | Fisheye Photograph |
|---|---|---|---|---|---|---|
| P1 | Hovenia acerba × 2, Machilus thunbergia × 1, Ligustrum vulgare × 1, Ligustrum japonicum ‘Howardii’, Loropetalum chinense, Pittosporum tobira, Rhododendron simsii, Ophiopogon japonicus (ET:DT = 6:3) | Tree- Shrub- Herb | 247.19 | 81.9 | 0.14 |  |
| P2 | Malus halliana × 4, Camphora officinarum × 2, Yulania denudata × 2, Platanus × acerifolia × 1 (ET:DT = 2:7) | Tree | 401.89 | 93.5 | 0.05 |  |
| P3 | Albizia julibrissin × 4, Camphora officinarum × 1, Chaenomeles japonica, Ophiopogon japonicus (ET:DT = 2:5) | Tree- Shrub- Herb | 685.02 | 89.6 | 0.10 |  |
| P4 | Camphora officinarum × 1, Osmanthus × 2, Ligustrum × 1, Sapindus saponaria × 1, PopulusL × 2 (ET:DT = 4:3) | Tree | 734.36 | 100.0 | 0.05 |  |
| P5 | Ormosia hosiei × 1, Elaeocarpus sylvestris × 2, Phoebe chekiangensis × 2, Yulania denudata × 1 (ET:DT = 5:1) | Tree | 150.20 | 46.9 | 0.15 |  |
| P6 | magnolia grandiflora × 2, Camellia japonica × 2, Celtis L. × 1, Ulmus parvifolia × 1, Ostrya rehderiana × 1, Prunus campanulata × 1, Nandina domestica (ET:DT = 5:4) | Large tree- Small tree | 503.39 | 79.6 | 0.18 |  |
| CK | lawn | / | 0 | 0 | 0.76 |  |
| No. | Community Composition | Community Structure | 3DGB/m3 | CD /% | SVF/% | Fisheye Photograph |
|---|---|---|---|---|---|---|
| P1 | Hovenia acerba × 2, Machilus thunbergii × 1, Ligustrum vulgare × 1, Ligustrum japonicum ‘Howardii’, Loropetalum chinense, Pittosporum tobira, Rhododendron simsii, Ophiopogon japonicus (ET:DT = 6:3) | Tree- Shrub- Herb | 221.87 | 72.6 | 0.33 |  |
| P2 | Malus halliana × 4, Camphora officinarum × 2, Yulania denudata × 2, Platanus × acerifolia × 1 (ET:DT = 2:7) | Tree | 338.82 | 84.6 | 0.14 |  |
| P3 | Albizia julibrissin × 4, Camphora officinarum × 1, Chaenomeles japonica, Ophiopogon japonicus (ET:DT = 2:5) | Tree- Shrub- Herb | 537.85 | 78.5 | 0.40 |  |
| P4 | Camphora officinarum × 1, Osmanthus × 2, Ligustrum × 1, Sapindus saponaria × 1, PopulusL × 2 (ET:DT = 4:3) | Tree | 703.10 | 95.3 | 0.12 |  |
| P5 | Ormosia hosiei × 1, Elaeocarpus sylvestris × 2, Phoebe chekiangensis × 2, Yulania denudata × 1 (ET:DT = 5:1) | Tree | 117.04 | 44.4 | 0.37 |  |
| P6 | magnolia grandiflora × 2, Camellia japonica × 2, Celtis L. × 1, Ulmus parvifolia × 1, Ostrya rehderiana × 1, Prunus campanulata × 1, Nandina domestica (ET:DT = 5:4) | Large tree- Small tree | 416.76 | 70.5 | 0.31 |  |
| CK | lawn | / | 0 | 0 | 0.87 |  |
| Level | Perception | THI |
|---|---|---|
| 1 | Excessive cold | <−10 |
| 2 | Very cold | −10–−1.8 |
| 3 | Cold | −1.8–13 |
| 4 | Cool | 13–15 |
| 5 | Comfortable | 15–20 |
| 6 | Hot | 20–26.5 |
| 7 | Very hot | 26.5–30 |
| 8 | Stuffy | >30 |
| Season | Community | AT/°C | RH/% | LI/lx | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| α | β | γ | α | β | γ | α | β | γ | ||
| Summer | P1 | 32.8 | 36.5 | 7.9 | 59.4 | 76.4 | −14.8 | 20,437.4 | 56,656.0 | 45.3 |
| P2 | 32.6 | 41.9 | 8.2 | 56.8 | 81.8 | −9.9 | 11,569.4 | 39,175.0 | 69.0 | |
| P3 | 32.0 | 35.3 | 9.9 | 57.9 | 80.1 | −12.0 | 14,036.0 | 54,083.0 | 62.4 | |
| P4 | 31.5 | 34.7 | 11.5 | 60.6 | 76.4 | −17.1 | 3954.7 | 18,762.0 | 89.4 | |
| P5 | 33.3 | 41.2 | 6.3 | 57.0 | 74.9 | −10.2 | 16,134.3 | 61,373.0 | 56.8 | |
| P6 | 31.8 | 38.0 | 10.5 | 61.0 | 85.8 | −17.9 | 17,479.6 | 55,517.0 | 53.2 | |
| CK | 35.6 | 48.4 | / | 51.7 | 69.6 | / | 37,346.1 | 63,127.0 | / | |
| Winter | P1 | 13.3 | 20.0 | 4.1 | 65.2 | 91.9 | −2.8 | 19,077.3 | 50,832.0 | 30.6 |
| P2 | 12.5 | 18.6 | 9.9 | 66.6 | 90.4 | −5.0 | 9793.5 | 45,769.0 | 64.4 | |
| P3 | 13.3 | 19.4 | 4.0 | 63.2 | 91.7 | 0.4 | 13,445.3 | 38,943.0 | 51.1 | |
| P4 | 12.3 | 18.3 | 11.1 | 61.2 | 93.3 | 3.6 | 7261.0 | 18,395.0 | 73.6 | |
| P5 | 12.7 | 18.5 | 8.1 | 65.3 | 89.6 | −2.9 | 10,242.5 | 37,538.0 | 72.7 | |
| P6 | 13.5 | 20.1 | 2.5 | 64.8 | 91.0 | −2.2 | 17,939.1 | 58,668.0 | 34.7 | |
| CK | 13.8 | 20.9 | / | 63.4 | 88.8 | / | 27,485.8 | 61,359.0 | / | |
| Season | Community | 8:00 | 9:00 | 10:00 | 11:00 | 12:00 | 13:00 | 14:00 | 15:00 | 16:00 | 17:00 | 18:00 | Average |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Summer | P1 | 27.3 | 28.2 | 28.4 | 28.6 | 29.0 | 29.4 | 29.4 | 29.4 | 28.9 | 28.2 | 27.9 | 28.6 bc |
| P2 | 24.6 | 26.1 | 27.4 | 29.8 | 30.4 | 28.7 | 29.3 | 28.6 | 28.4 | 27.9 | 27.7 | 28.1 bc | |
| P3 | 25.6 | 26.6 | 26.7 | 28.3 | 28.7 | 29.0 | 29.1 | 28.7 | 28.3 | 27.9 | 27.7 | 27.9 bc | |
| P4 | 24.8 | 26.2 | 26.7 | 27.6 | 28.0 | 28.7 | 28.8 | 28.5 | 28.4 | 28.2 | 27.9 | 27.7 c | |
| P5 | 25.3 | 26.6 | 28.6 | 29.6 | 29.7 | 29.9 | 29.4 | 30.9 | 28.8 | 28.4 | 27.8 | 28.7 b | |
| P6 | 24.1 | 26.3 | 28.3 | 28.3 | 28.3 | 28.5 | 29.3 | 28.6 | 29.2 | 29.0 | 27.9 | 27.9 bc | |
| CK | 26.1 | 29.0 | 29.8 | 31.0 | 29.9 | 31.3 | 30.9 | 30.8 | 29.6 | 28.9 | 28.0 | 29.7 a | |
| Winter | P1 | 4.8 | 8.3 | 10.8 | 13.4 | 15.2 | 16.4 | 17.4 | 16.6 | 14.9 | 13.9 | 10.8 | 13.2 a |
| P2 | 6.6 | 8.3 | 9.6 | 12.1 | 13.3 | 15.0 | 15.8 | 15.9 | 15.3 | 14.3 | 12.8 | 12.6 a | |
| P3 | 6.0 | 8.7 | 11.0 | 14.5 | 14.3 | 17.0 | 16.0 | 16.0 | 14.3 | 12.4 | 12.4 | 13.3 a | |
| P4 | 5.1 | 7.9 | 9.5 | 12.3 | 13.8 | 15.0 | 15.7 | 15.8 | 15.1 | 14.5 | 13.2 | 12.5 a | |
| P5 | 6.6 | 8.3 | 10.3 | 12.8 | 13.7 | 15.3 | 16.0 | 16.0 | 15.4 | 14.2 | 13.0 | 12.9 a | |
| P6 | 6.6 | 8.3 | 10.3 | 13.9 | 15.2 | 17.8 | 17.1 | 16.3 | 15.4 | 13.9 | 12.5 | 13.4 a | |
| CK | 6.1 | 8.9 | 11.4 | 14.0 | 16.0 | 17.0 | 17.9 | 16.8 | 15.4 | 14.3 | 12.9 | 13.7 a |
| Season | Dependent Variable | Pr > F | R2 | Independent Variable | Parameter Estimation |
|---|---|---|---|---|---|
| Summer | AT | 0.001 | 0.893 | CD | −0.035 |
| RH | 0.027 | 0.656 | SVF | −10.155 | |
| LI | 0.003 | 0.857 | SVF | 38,084.694 | |
| THI | <0.001 | 0.910 | CD | −0.018 | |
| Winter | AT | 0.027 | 0.656 | SVF | 1.270 |
| RH | No variables enter the equation | ||||
| LI | 0.013 | 0.743 | SVF | 24,224.560 | |
| THI | 0.020 | 0.696 | SVF | 1.420 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Pan, P.; Fang, D.; Guo, C. Effects of Plant Communities in Urban Green Spaces on Microclimate and Thermal Comfort. Forests 2025, 16, 799. https://doi.org/10.3390/f16050799
Li W, Pan P, Fang D, Guo C. Effects of Plant Communities in Urban Green Spaces on Microclimate and Thermal Comfort. Forests. 2025; 16(5):799. https://doi.org/10.3390/f16050799
Chicago/Turabian StyleLi, Wenjie, Pinwei Pan, Dongming Fang, and Chao Guo. 2025. "Effects of Plant Communities in Urban Green Spaces on Microclimate and Thermal Comfort" Forests 16, no. 5: 799. https://doi.org/10.3390/f16050799
APA StyleLi, W., Pan, P., Fang, D., & Guo, C. (2025). Effects of Plant Communities in Urban Green Spaces on Microclimate and Thermal Comfort. Forests, 16(5), 799. https://doi.org/10.3390/f16050799