
Enhancement of Bioactivity of Common Ash and Manna Ash Leaf Extracts Against Spongy Moth Larvae Using a Chitosan–Gelatin Biopolymer Matrix
 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemical Analysis of the Prepared Extracts
2.3. Formulation Preparation
2.4. Insect Material
2.4.1. Feeding Deterrent Activity
2.4.2. Larval Growth and Consumption
2.4.3. Digestive and Contact Toxicity
2.5. Statistical Analysis
3. Results
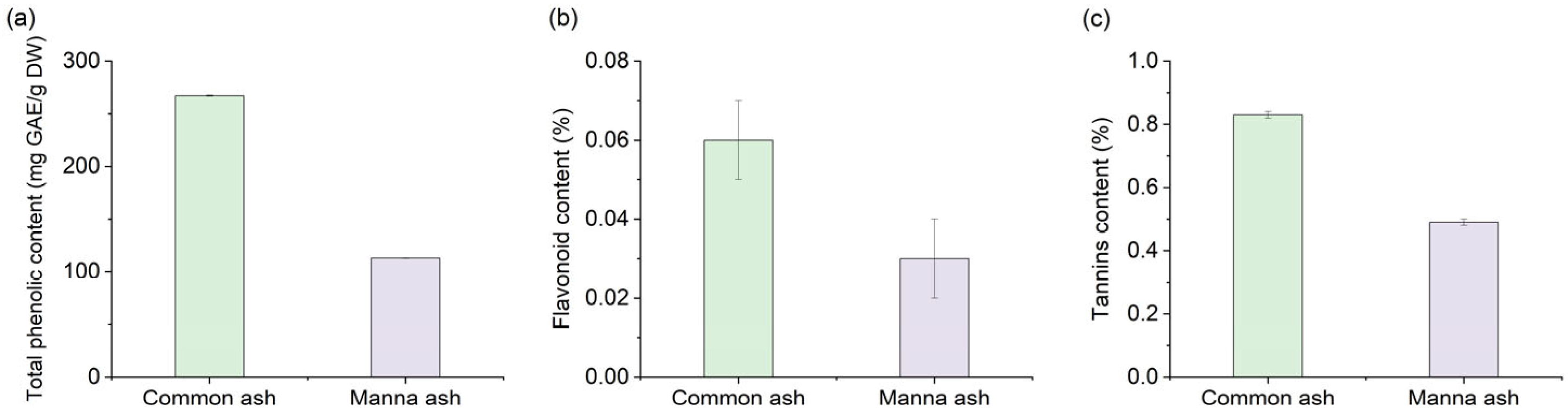
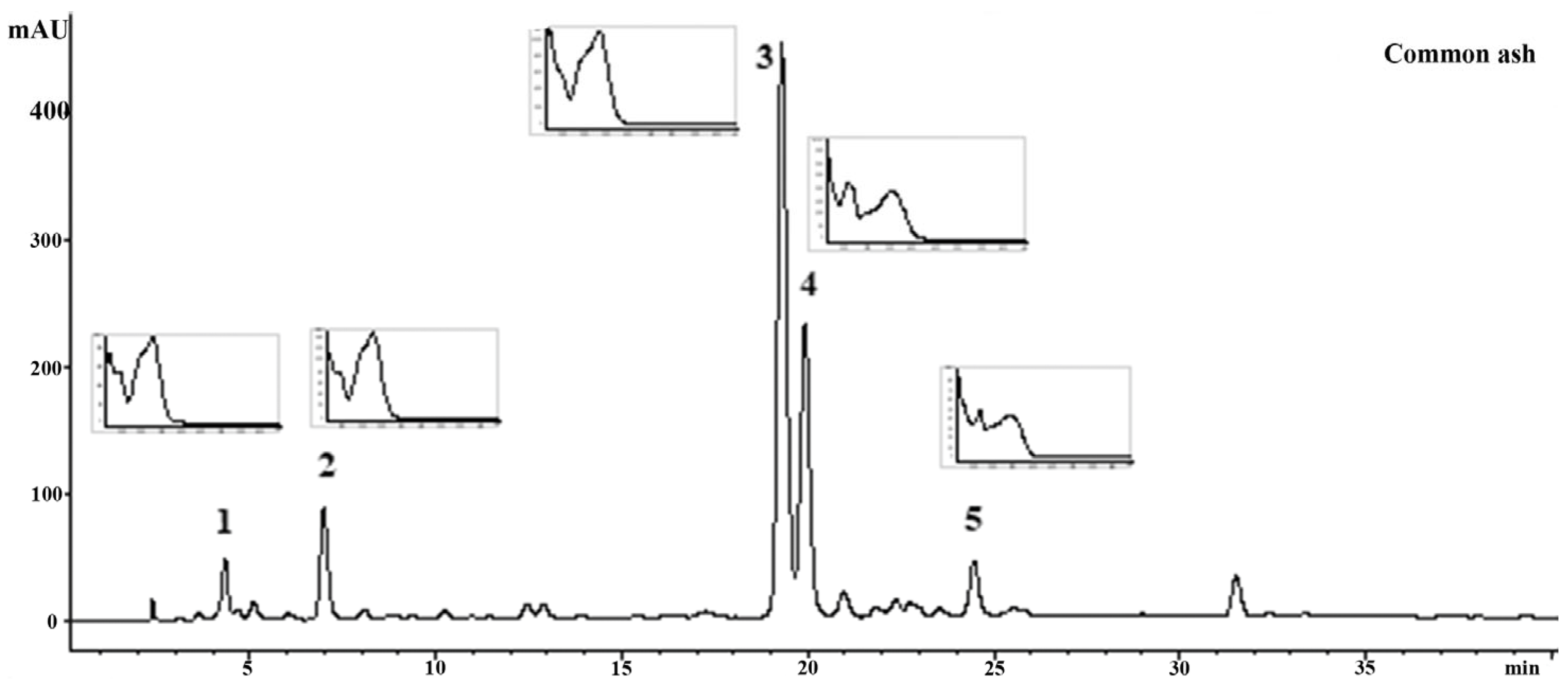
3.1. Chemical Composition of the Crude Leaf Extracts
3.2. Feeding Deterrence on the Spongy Moth Larvae
3.3. Nutritional Indices of the Spongy Moth Larvae
3.4. Duration of the Spongy Moth Third Larval Instar in the Toxicity Trials
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lewis, S.L. Tropical Forests and the Changing Earth System. Philos. Trans. R. Soc. B Biol. Sci. 2005, 361, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.L.L.; Arnell, A.; Maney, C.; Butchart, S.H.M.; Hilton-Taylor, C.; Ciciarelli, C.; Davis, C.; Dinerstein, E.; Purvis, A.; Burgess, N.D. Measuring Forest Biodiversity Status and Changes Globally. Front. For. Glob. Change 2019, 2, 464096. [Google Scholar] [CrossRef]
- Smith, P.; Ashmore, M.R.; Black, H.I.J.; Burgess, P.J.; Evans, C.D.; Quine, T.A.; Thomson, A.M.; Hicks, K.; Orr, H.G. REVIEW: The Role of Ecosystems and Their Management in Regulating Climate, and Soil, Water and Air Quality. J. Appl. Ecol. 2013, 50, 812–829. [Google Scholar] [CrossRef]
- Park, J.; Bui, H.T.; Lee, E.; Lim, H.S.; Lim, H.B.; Park, B.J. Accumulation of Particulate Matter, Heavy Metals, and Air Pollution Tolerance Index of 10 Species of Urban Forest Plants. Water Air Soil. Pollut. 2025, 236, 1–17. [Google Scholar] [CrossRef]
- Patacca, M.; Lindner, M.; Lucas-Borja, M.E.; Cordonnier, T.; Fidej, G.; Gardiner, B.; Hauf, Y.; Jasinevičius, G.; Labonne, S.; Linkevičius, E.; et al. Significant Increase in Natural Disturbance Impacts on European Forests since 1950. Glob. Change Biol. 2023, 29, 1359–1376. [Google Scholar] [CrossRef]
- Viana-Soto, A.; Senf, C. The European Forest Disturbance Atlas: A Forest Disturbance Monitoring System Using the Landsat Archive. Earth Syst. Sci. Data Discuss. 2024, 2024, 1–42. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.S.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten Golden Rules for Reforestation to Optimize Carbon Sequestration, Biodiversity Recovery and Livelihood Benefits. Glob. Change Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Hartmann, H.; Bastos, A.; Das, A.J.; Esquivel-Muelbert, A.; Hammond, W.M.; Martínez-Vilalta, J.; Mcdowell, N.G.; Powers, J.S.; Pugh, T.A.M.; Ruthrof, K.X.; et al. Climate Change Risks to Global Forest Health: Emergence of Unexpected Events of Elevated Tree Mortality Worldwide. Annu. Rev. Plant Biol. 2022, 73, 673–702. [Google Scholar] [CrossRef]
- Zanuncio, J.C.; Lemes, P.G.; Antunes, L.R.; Maia, J.L.S.; Mendes, J.E.P.; Tanganelli, K.M.; Salvador, J.F.; Serrão, J.E. The Impact of the Forest Stewardship Council (FSC) Pesticide Policy on the Management of Leaf-Cutting Ants and Termites in Certified Forests in Brazil. Ann. For. Sci. 2016, 73, 205–214. [Google Scholar] [CrossRef]
- Lemes, P.G.; Zanuncio, J.C.; Jacovine, L.A.G.; Wilcken, C.F.; Lawson, S.A. Forest Stewardship Council and Responsible Wood Certification in the Integrated Pest Management in Australian Forest Plantations. For. Policy Econ. 2021, 131, 102541. [Google Scholar] [CrossRef]
- Montgomery, M.E.; Wallner, W.E. The Gypsy Moth. In Dynamics of Forest Insect Populations; Springer: Boston, MA, USA, 1988; pp. 353–375. [Google Scholar] [CrossRef]
- Fajvan, M.A.; Wood, J.M. Stand Structure and Development after Gypsy Moth Defoliation in the Appalachian Plateau. For. Ecol. Manag. 1996, 89, 79–88. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Gottschalk, K.W.; Muzika, R.-M.; Montgomery, M.E.; Young, R.; O’Day, K.; Kelley, B. Suitability of North American Tree Species to Gypsy Moth: A Summary of Field and Laboratory Tests; General Technical Report NE-211; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1995; 34p. [Google Scholar] [CrossRef]
- Johnson, D.M.; Liebhold, A.M.; Bjørnstad, O.N. Geographical Variation in the Periodicity of Gypsy Moth Outbreaks. Ecography 2006, 29, 367–374. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.M.; Simard, F.; Courchamp, F. Massive yet Grossly Underestimated Global Costs of Invasive Insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef]
- Leroy, B.M.L. Global Insights on Insecticide Use in Forest Systems: Current Use, Impacts and Perspectives in a Changing World. Curr. For. Rep. 2024, 11, 1–30. [Google Scholar] [CrossRef]
- Rattan, R.S. Mechanism of Action of Insecticidal Secondary Metabolites of Plant Origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Peterson, J.A.; Ode, P.J.; Oliveira-Hofman, C.; Harwood, J.D. Integration of Plant Defense Traits with Biological Control of Arthropod Pests: Challenges and Opportunities. Front. Plant Sci. 2016, 7, 198096. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Pavela, R. History, Presence and Perspective of Using Plant Extracts as Commercial Botanical Insecticides and Farm Products for Protection against Insects—A Review. Plant Prot. Sci. 2016, 52, 229. [Google Scholar] [CrossRef]
- Devrnja, N.; Kostić, I.; Lazarević, J.; Savić, J.; Ćalić, D. Evaluation of Tansy Essential Oil as a Potential “Green” Alternative for Gypsy Moth Control. Environ. Sci. Pollut. Res. 2020, 27, 11958–11967. [Google Scholar] [CrossRef]
- Gvozdenac, S.; Indjic, D.; Vukovic, S.; Grahovac, M.; Tanaskovic, S. Antifeeding Activity of Several Plant Extracts against Lymantria dispar. L. (Lepidoptera: Lymantriidae) Larvae. Pesticidi i Fitomedicina 2012, 27, 305–311. [Google Scholar] [CrossRef]
- Kostić, I.; Petrović, O.; Milanović, S.; Popović, Z.; Stanković, S.; Todorović, G.; Kostić, M. Biological Activity of Essential Oils of Athamanta haynaldii and Myristica fragrans to Gypsy Moth Larvae. Ind. Crops Prod. 2013, 41, 17–20. [Google Scholar] [CrossRef]
- Markovic, I.; Norris, D.M.; Cekic, M. Some Chemical Bases for Gypsy Moth, Lymantria dispar, Larval Rejection of Green Ash, Fraxinus pennsylvanica, Foliage as Food. J. Chem. Ecol. 1996, 22, 2283–2298. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, B.M.; Milanović, S.D.; Milenković, I.L.; Todosijević, M.M.; Đorđević, I.; Brkić, M.Z.; Mitić, Z.S.; Marin, P.D.; Tešević, V.V. Bioactivity of Chamaecyparis lawsoniana (A. Murray) Parl. and Thuja plicata Donn Ex D. Don Essential Oils on Lymantria dispar. (Linnaeus, 1758) (Lepidoptera: Erebidae) Larvae and Phytophthora de Bary 1876 Root Pathogens. Ind. Crops Prod. 2022, 178, 114550. [Google Scholar] [CrossRef]
- Milanović, S.D.; Milenković, I.L.; Lazarević, J.M.; Todosijević, M.M.; Ljujić, J.P.; Mitić, Z.S.; Nikolić, B.M.; Marin, P.D.; Tešević, V.V. Biological Activity of Essential Oils of Calocedrus decurrens and Cupressus arizonica on Lymantria dispar. Larvae and Phytophthora Root Pathogens. Ind. Crops Prod. 2024, 215, 118602. [Google Scholar] [CrossRef]
- Tanasković, S.; Gvozdenac, S.; Kolarov, R.; Bursić, V.; Konstantinović, B.; Prvulović, D. Antifeeding and Insecticidal Activity of Ailanthus altissima and Morus alba Extracts Against Gipsy Moth (Lymantria dispar. (L.), Lepidoptera, Lymantridae) Larvae Under Laboratory Conditions. J. Entomol. Res. Soc. 2021, 23, 197–212. [Google Scholar] [CrossRef]
- Kaur, S.; Samota, M.K.; Choudhary, M.; Choudhary, M.; Pandey, A.K.; Sharma, A.; Thakur, J. How Do Plants Defend Themselves against Pathogens-Biochemical Mechanisms and Genetic Interventions. Physiol. Mol. Biol. Plants 2022, 28, 485–504. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of Plant Defense against Insect Herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef]
- Isman, M.B.; Akhtar, Y. Plant Natural Products as a Source for Developing Environmentally Acceptable Insecticides. In Insecticides Design Using Advanced Technologies; Springer: Berlin/Heidelberg, Germany, 2007; pp. 235–248. ISBN 9783540469070. [Google Scholar] [CrossRef]
- Khursheed, A.; Rather, M.A.; Jain, V.; Wani, A.R.; Rasool, S.; Nazir, R.; Malik, N.A.; Majid, S.A. Plant Based Natural Products as Potential Ecofriendly and Safer Biopesticides: A Comprehensive Overview of Their Advantages over Conventional Pesticides, Limitations and Regulatory Aspects. Microb. Pathog. 2022, 173, 105854. [Google Scholar] [CrossRef]
- Turchen, L.M.; Cosme-Júnior, L.; Guedes, R.N.C. Plant-Derived Insecticides Under Meta-Analyses: Status, Biases, and Knowledge Gaps. Insects 2020, 11, 532. [Google Scholar] [CrossRef]
- Giunti, G.; Campolo, O.; Laudani, F.; Palmeri, V.; Spinozzi, E.; Bonacucina, G.; Maggi, F.; Pavela, R.; Canale, A.; Lucchi, A.; et al. Essential Oil-Based Nanoinsecticides: Ecological Costs and Commercial Potential. In Development and Commercialization of Biopesticides: Costs and Benefits; Academic Press: Cambridge, MA, USA, 2023; pp. 375–402. [Google Scholar] [CrossRef]
- Reddy, C.K.; Agarwal, R.K.; Shah, M.A.; Suriya, M. Encapsulation Techniques for Plant Extracts. In Plant Extracts: Applications in the Food Industry; Academic Press: Cambridge, MA, USA, 2022; pp. 75–88. [Google Scholar] [CrossRef]
- Detsi, A.; Kavetsou, E.; Kostopoulou, I.; Pitterou, I.; Pontillo, A.R.N.; Tzani, A.; Christodoulou, P.; Siliachli, A.; Zoumpoulakis, P. Nanosystems for the Encapsulation of Natural Products: The Case of Chitosan Biopolymer as a Matrix. Pharmaceutics 2020, 12, 669. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Abou-Elseoud, W.S.; Elbehery, H.H.; Hassan, M.L. Chitosan-Cellulose Nanoencapsulation Systems for Enhancing the Insecticidal Activity of Citronella Essential Oil against the Cotton Leafworm Spodoptera Littoralis. Ind. Crops Prod. 2022, 184, 115089. [Google Scholar] [CrossRef]
- Beck, P.; Caudullo, G.; Tinner, W.; de Rigo, D. Fraxinus excelsior in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Caudullo, G.; Rigo, D. Fraxinus ornus in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Milanović, S.D.; Popović, M.M.; Dobrosavljević, J.N.; Kostić, I.M.; Lazarević, J.M. Desperate Times Call for Desperate Measures: Short-Term Use of the Common Ash Tree by Gypsy Moth Larvae (Lepidoptera: Erebidae) under Density and Starvation Stress. Arch. Biol. Sci. 2020, 72, 63–69. [Google Scholar] [CrossRef]
- Lechowicz, M.J.; Jobin, L. Estimating the Susceptibility of Tree Species to Attack by the Gypsy Moth, Lymantria Dispar. Ecol. Entomol. 1983, 8, 171–183. [Google Scholar] [CrossRef]
- Sarfraz, I.; Rasul, A.; Jabeen, F.; Younis, T.; Zahoor, M.K.; Arshad, M.; Ali, M. Fraxinus: A Plant with Versatile Pharmacological and Biological Activities. Evid.-Based Complement. Altern. Med. 2017, 2017, 4269868. [Google Scholar] [CrossRef]
- Kołtun-Jasion, M.; Sawulska, P.; Patyra, A.; Woźniak, M.; Dudek, M.K.; Filipek, A.; Kiss, A.K. Bio-Guided Isolation of Compounds from Fraxinus excelsior Leaves with Anti-Inflammatory Activity. Int. J. Mol. Sci. 2023, 24, 3750. [Google Scholar] [CrossRef]
- Eruygur, N. Fraxiınus Ornus L. In Novel Drug Targets with Traditional Herbal Medicines; Springer International Publishing: Cham, Switzerland, 2022; pp. 289–300. ISBN 978-3-031-07753-1. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- European Directorate for the Quality of Medicines & HealthCare. European Pharmacopoeia (Ph. Eur.) 11th Edition . Available online: https://www.edqm.eu/en/european-pharmacopoeia-ph.-eur.-11th-edition#{%22468369%22:[0]} (accessed on 20 December 2024).
- Milanović, S.; Milenković, I.; Dobrosavljević, J.; Popović, M.; Solla, A.; Tomšovský, M.; Jankovský, L. Growth Rates of Lymantria dispar Larvae and Quercus robur Seedlings at Elevated CO2 Concentration and Phytophthora plurivora Infection. Forests 2020, 11, 1059. [Google Scholar] [CrossRef]
- Odell, T.M.; Rollinson, W.D. A Technique for Rearing the Gypsy Moth, Porthetria dispar (L.), on an Artificial Diet. J. Econ. Entomol. 1966, 59, 741–742. [Google Scholar] [CrossRef]
- Milanović, S.; Janković-Tomanić, M.; Kostić, I.; Kostić, M.; Morina, F.; Živanović, B.; Lazarević, J. Behavioural and Physiological Plasticity of Gypsy Moth Larvae to Host Plant Switching. Entomol. Exp. Appl. 2016, 158, 152–162. [Google Scholar] [CrossRef]
- Szczepanik, M.; Dams, I.; Wawrzeńczyk, C. Feeding Deterrent Activity of Terpenoid Lactones with the P-Menthane System Against the Colorado Potato Beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 2005, 34, 1433–1440. [Google Scholar] [CrossRef]
- Szczepanik, M.; Szumny, A.; Wawrzeńczyk, C. The Effect of α-Methylenelactone Group on the Feeding Deterrent Activity of Natural and Synthetic Alkenes Against Colorado Potato, Leptinotarsa decemlineata Say. Pol. J. Environ. Stud. 2009, 18, 1107–1112. [Google Scholar]
- Waldbauer, G.P. The Consumption and Utilization of Food by Insects. Adv. Insect Phys. 1968, 5, 229–288. [Google Scholar] [CrossRef]
- Jovanović, J.; Ćirković, J.; Radojković, A.; Tasić, N.; Mutavdžić, D.; Branković, G.; Branković, Z. Enhanced Stability of Encapsulated Lemongrass Essential Oil in Chitosan-Gelatin and Pectin-Gelatin Biopolymer Matrices Containing ZnO Nanoparticles. Int. J. Biol. Macromol. 2024, 275, 133335. [Google Scholar] [CrossRef]
- Ćirković, J.; Radojković, A.M.; Jovanović, J.; Perać, S.; Branković, Z.M.; Milenković, I.; Milanović, S.D.; Dobrosavljević, J.N.; Tadić, V.M.; Žugić, A.R.; et al. Encapsulated Thuja plicata Essential Oil into Biopolymer Matrix as a Potential Pesticide against Phytophthora Root Pathogens. Int. J. Biol. Macromol. 2024, 278, 134684. [Google Scholar] [CrossRef]
- Milanović, S.D.; Simović, N.; Dobrosavljević, J.; Milenković, I.L.; Branković, Z.; Ćirković, J.; Radojković, A.; Perać, S.; Jovanović, J.; Tadić, V.; et al. Bioactivity of the Tree of Heaven Leaf Extracts Incorporated into Biopolymer Matrix Against Spongy Moth Larvae. Forests 2025, 16, 375. [Google Scholar] [CrossRef]
- Čulum, D.; Vidic, D.; Topčagić, A.; Klepo, L.; Čopra-Janićijević, A. Chemical Composition and Antioxidant Activity of Fraxinus ornus L. and Fraxinus excelsior L. Kem. Ind. 2024, 73, 19–25. [Google Scholar] [CrossRef]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A Review of Its Occurrence, (Bio)Synthesis and Pharmacological Significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef]
- Predojević, D.Z.; Vukajlović, F.N.; Mihailović, V.B.; Tanasković, S.T.; Pešić, S.B. Larvicidal Efficacy of Verbascum Spp. Methanol Extracts against Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae). Kragujev. J. Sci. 2020, 42, 167–175. [Google Scholar] [CrossRef]
- Zaynab, M.; Khan, J.; Al-Yahyai, R.; Sadder, M.; Li, S. Toxicity of Coumarins in Plant Defense against Pathogens. Toxicon 2024, 250, 108118. [Google Scholar] [CrossRef]
- Mohan, K.; Kandasamy, S.; Rajarajeswaran, J.; Sundaram, T.; Bjeljac, M.; Surendran, R.P.; Ganesan, A.R. Chitosan-Based Insecticide Formulations for Insect Pest Control Management: A Review of Current Trends and Challenges. Int. J. Biol. Macromol. 2024, 280, 135937. [Google Scholar] [CrossRef]
- Beaumelle, L.; Tison, L.; Eisenhauer, N.; Hines, J.; Malladi, S.; Pelosi, C.; Thouvenot, L.; Phillips, H.R.P. Pesticide Effects on Soil Fauna Communities—A Meta-Analysis. J. Appl. Ecol. 2023, 60, 1239–1253. [Google Scholar] [CrossRef]
- Popović, Z.; Kostić, M.; Stanković, S.; Milanović, S.; Sivčev, I.; Kostić, I.; Kljajić, P. Ecologically Acceptable Usage of Derivatives of Essential Oil of Sweet Basil, Ocimum basilicum, as Antifeedants against Larvae of the Gypsy Moth, Lymantria dispar. J. Insect Sci. 2013, 13, 161. [Google Scholar] [CrossRef] [PubMed]
- Kostić, I.; Lazarević, J.; Jovanović, D.Š.; Kostić, M.; Marković, T.; Milanović, S. Potential of Essential Oils from Anise, Dill and Fennel Seeds for the Gypsy Moth Control. Plants 2021, 10, 2194. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.L.; Card, J.A. Ailanthus altissima Aqueous Extract Deters Spodoptera frugiperda Oviposition. Great Lakes Entomol. 2020, 53, 11. [Google Scholar] [CrossRef]
- Wagner, R.L.; Leach, M.E.; Wallace, J.R. Leaf Extract from Ailanthus altissima Negatively Impacts Life History Aspects in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Kans. Entomol. Soc. 2021, 93, 140–152. [Google Scholar] [CrossRef]
- Stoyenoff, J.L.; Witter, J.A.; Montgomery, M.E. Nutritional Indices in the Gypsy Moth (Lymantria dispar (L.)) under Field Conditions and Host Switching Situations. Oecologia 1994, 97, 158–170. [Google Scholar] [CrossRef]
- Milanović, S.; Mihajlović, L.; Karadžić, D.; Jankovsky, L.; Aleksić, P.; Janković-Tomanić, M.; Lazarević, J. Effects of Pedunculate Oak Tree Vitality on Gypsy Moth Preference and Performance. Arch. Biol. Sci. 2014, 66, 1659–1672. [Google Scholar] [CrossRef]
- Bölöni, J.; Aszalós, R.; Frank, T.; Ódor, P. Forest Type Matters: Global Review about the Structure of Oak Dominated Old-Growth Temperate Forests. For. Ecol. Manag. 2021, 500, 119629. [Google Scholar] [CrossRef]
- Dobrosavljević, J.; Marković, Č.; Marjanović, M.; Milanović, S. Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat. Forests 2020, 11, 1300. [Google Scholar] [CrossRef]
- Pereira, J.F.; Oliveira, A.L.M.; Sartori, D.; Yamashita, F.; Mali, S. Perspectives on the Use of Biopolymeric Matrices as Carriers for Plant-Growth Promoting Bacteria in Agricultural Systems. Microorganisms 2023, 11, 467. [Google Scholar] [CrossRef]
- Miresmailli, S.; Isman, M.B. Botanical Insecticides Inspired by Plant-Herbivore Chemical Interactions. Trends Plant Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Christopher Cutler, G.; Scott-Dupree, C.D.; Sultan, M.; McFarlane, A.D.; Brewer, L. A Large-Scale Field Study Examining Effects of Exposure to Clothianidin Seed-Treated Canola on Honey Bee Colony Health Development, and Overwintering Success. PeerJ 2014, 2014, e652. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number Assigned to the Respective Peak in Figure 1 and Figure 2 | The Identified Compound | Content in the Investigated Extract (mg/g DW Extract) Common Ash | Content in the Investigated Extract (mg/g DW Extract) Manna Ash |
|---|---|---|---|
| 1 | neochlorogenic acid | 12.78 ± 0.09 | |
| 2 | chlorogenic acid | 18.34 ± 0.11 | |
| 3 | verbascoside | 144.79 ± 0.31 | |
| 4 | rutin | 38.69 ± 0.03 | 3.95 ± 0.03 |
| 5 | kaempferol-3-O-glucoside | 2.17 ± 0.02 | |
| 6 | aesculin | 20.41 ± 0.10 | |
| 7 | aesculetin | 92.18 ± 0.13 | |
| 8 | isoscopoletin | 2.83 ± 0.03 | |
| 9 | scopoletin | 2.91 ± 0.03 | |
| 10 | myricetin | 1.26 ± 0.03 | |
| 11 | hyperoside | 1.39 ± 0.05 | |
| 12 | isoquercetin | 1.09 ± 0.01 | |
| 13 | naringin | 4.09 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simović, N.; Dobrosavljević, J.; Milenković, I.L.; Branković, Z.; Ćirković, J.; Radojković, A.; Perać, S.; Jovanović, J.; Tadić, V.; Žugić, A.; et al. Enhancement of Bioactivity of Common Ash and Manna Ash Leaf Extracts Against Spongy Moth Larvae Using a Chitosan–Gelatin Biopolymer Matrix. Forests 2025, 16, 774. https://doi.org/10.3390/f16050774
Simović N, Dobrosavljević J, Milenković IL, Branković Z, Ćirković J, Radojković A, Perać S, Jovanović J, Tadić V, Žugić A, et al. Enhancement of Bioactivity of Common Ash and Manna Ash Leaf Extracts Against Spongy Moth Larvae Using a Chitosan–Gelatin Biopolymer Matrix. Forests. 2025; 16(5):774. https://doi.org/10.3390/f16050774
Chicago/Turabian StyleSimović, Nemanja, Jovan Dobrosavljević, Ivan Lj. Milenković, Zorica Branković, Jovana Ćirković, Aleksandar Radojković, Sanja Perać, Jelena Jovanović, Vanja Tadić, Ana Žugić, and et al. 2025. "Enhancement of Bioactivity of Common Ash and Manna Ash Leaf Extracts Against Spongy Moth Larvae Using a Chitosan–Gelatin Biopolymer Matrix" Forests 16, no. 5: 774. https://doi.org/10.3390/f16050774
APA StyleSimović, N., Dobrosavljević, J., Milenković, I. L., Branković, Z., Ćirković, J., Radojković, A., Perać, S., Jovanović, J., Tadić, V., Žugić, A., Branković, G., & Milanović, S. D. (2025). Enhancement of Bioactivity of Common Ash and Manna Ash Leaf Extracts Against Spongy Moth Larvae Using a Chitosan–Gelatin Biopolymer Matrix. Forests, 16(5), 774. https://doi.org/10.3390/f16050774