
Microclimate Shifts and Leaf Miner Community Responses to Shelterwood Regeneration in Sessile Oak Forests


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Experimental Design
2.3. Environmental and Plant Characteristics
2.4. Characteristics of the Leaf Miner Community Analyzed
- Species richness (number of species identified) per tree, and the number of species per 100 leaves, calculated as:where si is the number of species found in one sample, li is the number of leaves per sample.
- Total species richness of leaf miners (number of species identified on all trees).
- The abundance of individual species (Ai) per tree, and the number of mines per 100 leaves, calculated as:where ni is the number of mines of one particular species found in one sample and li is the number of leaves per sample.
- The total abundance of leaf miners (Ab) is calculated as a sum of all individual species’ abundance.
- Shannon’s index of diversity (H′), calculated as:where Si is the species richness and pi is the proportion of individuals belonging to the i-th species in the dataset [61].
- Buzas and Gibson’s evenness index (Ev), calculated as:where H is Shannon’s index of diversity and Si is species richness [62].
2.5. Statistical Analysis
3. Results
3.1. The Influence of Changes in the Canopy Cover on the Forest Microclimate
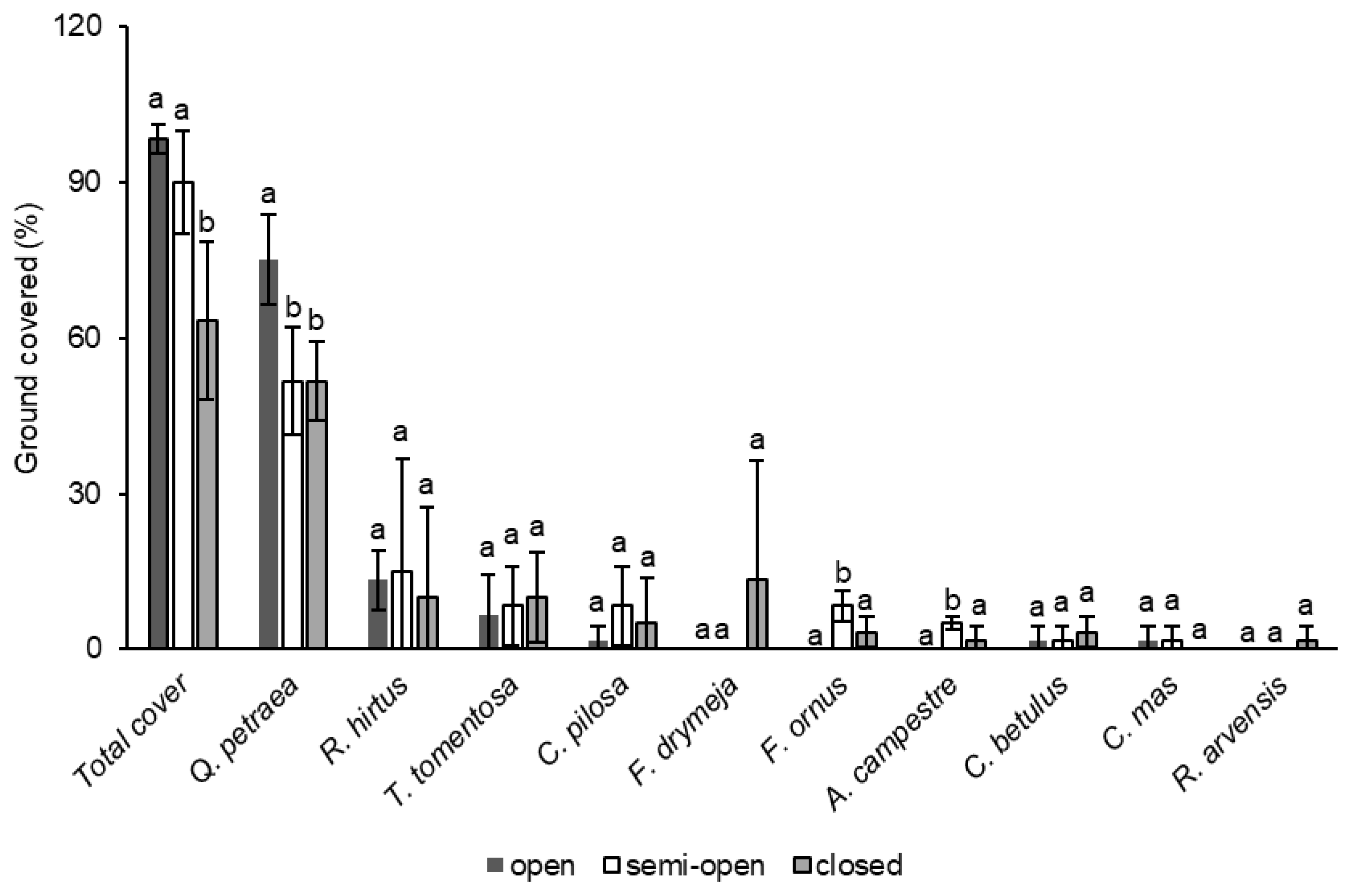
3.2. The Influence of Canopy Cover on Understory Vegetation
3.3. The Influence of Canopy Closure on the Leaf Miner Community on Young Sessile Oak Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fornoff, F.; Staab, M.; Zhu, C.D.; Klein, A.M. Multi-Trophic Communities Re-Establish with Canopy Cover and Microclimate in a Subtropical Forest Biodiversity Experiment. Oecologia 2021, 196, 289–301. [Google Scholar] [CrossRef] [PubMed]
- FAO; UNEP. The State of the World’s Forests 2020; De Gruyter: Rome, Italy, 2020; ISBN 978-92-5-132419-6. [Google Scholar]
- European Commission: Directorate-General for Environment. Guidelines on Closer-to-Nature Forest Management; Publications Office of the European Union: Luxembourg, 2023. [Google Scholar]
- Du Bus De Warnaffe, G.; Angerand, S. Forest Management and Climate Change: A New Approach to the French Strategy; Fern: Brussels, Belgium, 2020. [Google Scholar]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; López-Senespleda, E.; Bravo, F.; Del Río, M. Forest Management and Carbon Sequestration in the Mediterranean Region: A Review. For. Syst. 2017, 26, 10. [Google Scholar] [CrossRef]
- Hämäläinen, A.; Runnel, K.; Ranius, T.; Strengbom, J. Diversity of Forest Structures Important for Biodiversity Is Determined by the Combined Effects of Productivity, Stand Age, and Management. Ambio 2024, 53, 718–729. [Google Scholar] [CrossRef]
- Kanjevac, B.; Krstić, M.; Babić, V.; Govedar, Z. Regeneration Dynamics and Development of Seedlings in Sessile Oak Forests in Relation to the Light Availability and Competing Vegetation. Forests 2021, 12, 384. [Google Scholar] [CrossRef]
- Máliš, F.; Ujházy, K.; Hederová, L.; Ujházyová, M.; Csölleová, L.; Coomes, D.A.; Zellweger, F. Microclimate Variation and Recovery Time in Managed and Old-Growth Temperate Forests. Agric. For. Meteorol. 2023, 342, 109722. [Google Scholar] [CrossRef]
- Horváth, C.V.; Kovács, B.; Tinya, F.; Schadeck Locatelli, J.; Németh, C.; Crecco, L.; Illés, G.; Csépányi, P.; Ódor, P. A Matter of Size and Shape: Microclimatic Changes Induced by Experimental Gap Openings in a Sessile Oak–Hornbeam Forest. Sci. Total Environ. 2023, 873, 162302. [Google Scholar] [CrossRef]
- De Frenne, P.; Zellweger, F.; Rodríguez-Sánchez, F.; Scheffers, B.R.; Hylander, K.; Luoto, M.; Vellend, M.; Verheyen, K.; Lenoir, J. Global Buffering of Temperatures Under Forest Canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef]
- Kanjevac, B.; Babić, V.; Stajić, S.; Martać, N.; Pavlović, B.; Furtula, D.; Čokeša, V. Key Drivers Affecting the Spatial Heterogeneity of the Regeneration Process in Old-Growth Beech Forests in Southeastern Europe. Front. For. Glob. Change 2023, 6, 1304037. [Google Scholar] [CrossRef]
- Bolton, N.W.; D’Amato, A.W. Herbaceous Vegetation Responses to Gap Size Within Natural Disturbance-Based Silvicultural Systems in Northeastern Minnesota, USA. Forests 2019, 10, 111. [Google Scholar] [CrossRef]
- Tinya, F.; Csépányi, P.; Horváth, C.V.; Kovács, B.; Németh, C.; Ódor, P. Fine-Scale Interventions Can Reinforce the Forest Character of the Understory Vegetation—The Effects of Different Artificial Gaps in an Oak-Dominated Forest. For. Ecol. Manag. 2025, 578, 122471. [Google Scholar] [CrossRef]
- Horák, J.; Marković, Č.; Dobrosavljević, J.; Rada, P.; Mladenović, S.; Kohutka, A.; Míkovcová, A.; Pech, P.; Drábek, O.; Tejnecký, V. Influence of Forest Structure and Soil Chemistry on Ants, Land Snails, and Beetles in Balkan Floodplain Forests: Examining Species Richness and Habitat Preferences. J. Soil Water Conserv. 2024, 79, 303–306. [Google Scholar] [CrossRef]
- Dobrosavljević, J.; Marković, Č.; Marjanović, M.; Milanović, S. Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat. Forests 2020, 11, 1300. [Google Scholar] [CrossRef]
- Marković, Č.; Dobrosavljević, J.; Vujičić, P.; Cebeci, H.H. Impact of Regeneration by Shelterwood Cutting on the Pedunculate Oak (Quercus robur) Leaf Mining Insect Community. Biologia 2021, 76, 1197–1203. [Google Scholar] [CrossRef]
- Valdés-Correcher, E.; Moreira, X.; Augusto, L.; Barbaro, L.; Bouget, C.; Bouriaud, O.; Branco, M.; Centenaro, G.; Csóka, G.; Damestoy, T.; et al. Search for Top-down and Bottom-up Drivers of Latitudinal Trends in Insect Herbivory in Oak Trees in Europe. Glob. Ecol. Biogeogr. 2021, 30, 651–665. [Google Scholar] [CrossRef]
- Achury, R.; Staab, M.; Blüthgen, N.; Weisser, W.W. Forest Gaps Increase True Bug Diversity by Recruiting Open Land Species. Oecologia 2023, 202, 299–312. [Google Scholar] [CrossRef]
- Preisser, E.; Smith, D.C.; Lowman, M.D. Canopy and Ground Level Insect Distribution in a Temperate Forest. Selbyana 1998, 19, 141–146. [Google Scholar]
- Thomson, L.J.; Macfadyen, S.; Hoffmann, A.A. Predicting the Effects of Climate Change on Natural Enemies of Agricultural Pests. Biol. Control 2010, 52, 296–306. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B. Canopy Structure and Vertical Patterns of Photosynthesis and Related Leaf Traits in a Deciduous Forest. Oecologia 1993, 96, 169–178. [Google Scholar] [CrossRef]
- Caldwell, E.; Read, J.; Sanson, G.D. Which Leaf Mechanical Traits Correlate with Insect Herbivory Among Feeding Guilds? Ann. Bot. 2016, 117, 349–361. [Google Scholar] [CrossRef]
- Klapwijk, M.J.; Bylund, H.; Schroeder, M.; Björkman, C. Forest Management and Natural Biocontrol of Insect Pests. Forestry 2016, 89, 253–262. [Google Scholar] [CrossRef]
- Schillé, L.; Valdés-Correcher, E.; Archaux, F.; Bălăcenoiu, F.; Bjørn, M.C.; Bogdziewicz, M.; Boivin, T.; Branco, M.; Damestoy, T.; de Groot, M.; et al. Decomposing Drivers in Avian Insectivory: Large-Scale Effects of Climate, Habitat and Bird Diversity. J. Biogeogr. 2024, 51, 1079–1094. [Google Scholar] [CrossRef]
- Dobrosavljević, J.; Marković, Č.; Marjanović, M. The Effect of Urban–Rural Gradient on Black Poplar Endophagous Herbivorous Insects. Arthropod-Plant Interact. 2023, 17, 341–350. [Google Scholar] [CrossRef]
- Lenk, A.; Richter, R.; Kretz, L.; Wirth, C. Effects of Canopy Gaps on Microclimate, Soil Biological Activity and Their Relationship in a European Mixed Floodplain Forest. Sci. Total Environ. 2024, 941, 173572. [Google Scholar] [CrossRef] [PubMed]
- Perlík, M.; Kraus, D.; Bußler, H.; Neudam, L.; Pietsch, S.; Mergner, U.; Seidel, D.; Sebek, P.; Thorn, S. Canopy Openness as the Main Driver of Aculeate Hymenoptera and Saproxylic Beetle Diversity Following Natural Disturbances and Salvage Logging. For. Ecol. Manag. 2023, 540, 121033. [Google Scholar] [CrossRef]
- Valdés-Correcher, E.; Popova, A.; Galmán, A.; Prinzing, A.; Selikhovkin, A.V.; Howe, A.G.; Mrazova, A.; Dulaurent, A.; Hampe, A.; Tack, A.J.M.; et al. Herbivory on the Pedunculate Oak along an Urbanization Gradient in Europe: Effects of Impervious Surface, Local Tree Cover, and Insect Feeding Guild. Ecol. Evol. 2022, 12, e8709. [Google Scholar] [CrossRef]
- Takafumi, H.; Kawase, S.; Nakamura, M.; Hiura, T. Herbivory in Canopy Gaps Created by a Typhoon Varies by Understory Plant Leaf Phenology. Ecol. Entomol. 2010, 35, 576–585. [Google Scholar] [CrossRef]
- Wildermuth, B.; Penanhoat, A.; Sennhenn-Reulen, H.; Matevski, D.; Drescher, J.; Aubry-Kientz, M.; Seidel, D.; Schuldt, A. Canopy Structure Influences Arthropod Communities Within and Beyond Tree Identity Effects: Insights from Combining LiDAR Data, Insecticidal Fogging and Machine Learning Regression Modelling. Ecol. Indic. 2024, 160, 111901. [Google Scholar] [CrossRef]
- Parker, G.G.; Fitzjarrald, D.R.; Gonçalves Sampaio, I.C. Consequences of Environmental Heterogeneity for the Photosynthetic Light Environment of a Tropical Forest. Agric. For. Meteorol. 2019, 278, 107661. [Google Scholar] [CrossRef]
- Parker, G.G.; Lowman, M.D.; Nadkarni, N.M. Structure and Microclimate of Forest Canopies. In Forest Canopies; Lowman, M.D., Nadkari, N.M., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 73–106. [Google Scholar]
- Holuša, J.; Fiala, T.; Foit, J. Ambrosia Beetles Prefer Closed Canopies: A Case Study in Oak Forests in Central Europe. Forests 2021, 12, 1223. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Ina, A.B.; Diaci, J.; Anić, I.; Boydak, M.; Curovic, M.; Govedar, Z.; Grigoriadis, N.; Ivojevic, S.; Keren, S.; et al. Culture and Silviculture: Origins and Evolution of Silviculture in Southeast Europe. Int. For. Rev. 2018, 20, 130–143. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W. Influence of Selection Systems and Shelterwood Methods on Understory Plant Communities of Longleaf Pine Forests in Flatwoods and Uplands. For. Ecol. Manag. 2015, 357, 138–150. [Google Scholar] [CrossRef]
- Day, K.; Koot, C.; Wiensczyk, A. The Shelterwood Silvicultural System in British Columbia—A Practitioner’s Guide Part 3: Operational Implementation. J. Ecosyst. Manag. 2011, 12, 95–106. [Google Scholar] [CrossRef]
- Agestam, E.; Ekö, P.M.; Nilsson, U.; Welander, N.T. The Effects of Shelterwood Density and Site Preparation on Natural Regeneration of Fagus Sylvatica in Southern Sweden. For. Ecol. Manag. 2003, 176, 61–73. [Google Scholar] [CrossRef]
- Nolet, P.; Kneeshaw, D.; Messier, C.; Béland, M. Comparing the Effects of Even- and Uneven-aged Silviculture on Ecological Diversity and Processes: A Review. Ecol. Evol. 2018, 8, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Nyland, R.D. The Shelterwood Method: Adapting to Diverse Management Objectives. J. For. 2010, 108, 419–420. [Google Scholar] [CrossRef]
- Atkinson, B.; Bailey, S.; Vaughan, I.P.; Memmott, J. A Comparison of Clearfelling and Gradual Thinning of Plantations for the Restoration of Insect Herbivores and Woodland Plants. J. Appl. Ecol. 2015, 52, 1538–1546. [Google Scholar] [CrossRef]
- Hering, E.M. Biology of the Leaf Miners; Dr. W. Junk: Hague, The Netherlands, 1951. [Google Scholar]
- Ellis, W.N. Leafminers and Plant Galls of Europe. Available online: http://www.bladmineerders.nl/ (accessed on 17 October 2023).
- Dobrosavljević, J.; Marković, Č. First Findings of Deciduous Woody Plant Leaf Miners in Serbia. J. Entomol. Res. Soc. 2024, 26, 183–207. [Google Scholar] [CrossRef]
- Dobrosavljević, J.; Marković, Č.; Bojić, S. Overview of Leaf Miner Fauna in Serbia. In Proceedings of the VIII International Agriculture Symposium “AGROSYM 2017”, Jahorina, Bosnia and Herzegovina, 5–8 October 2017; pp. 1490–1498. [Google Scholar]
- Jones, E.L.; Leather, S.R. Invertebrates in Urban Areas: A Review. Eur. J. Entomol. 2012, 109, 463–478. [Google Scholar] [CrossRef]
- Maldonado-López, Y.; Cuevas-Reyes, P.; González-Rodríguez, A.; Pérez-López, G.; Acosta-Gómez, C.; Oyama, K. Relationships among Plant Genetics, Phytochemistry and Herbivory Patterns in Quercus castanea Across a Fragmented Landscape. Ecol. Res. 2015, 30, 133–143. [Google Scholar] [CrossRef]
- Rickman, J.K.; Connor, E.F. The Effect of Urbanization on the Quality of Remnant Habitats for Leaf-Mining Lepidoptera on Quercus agrifolia. Ecography 2003, 26, 777–787. [Google Scholar] [CrossRef]
- Grabenweger, G.; Kehrli, P.; Schlick-Steiner, B.; Steiner, F.; Stolz, M.; Bacher, S. Predator Complex of the Horse Chestnut Leafminer Cameraria ohridella: Identification and Impact Assessment. J. Appl. Entomol. 2005, 129, 353–362. [Google Scholar] [CrossRef]
- Cebeci, H.H.; Markovic, C.; Grabenweger, G.; Ayberk, H.; Dobrosavljevic, J.; Goltas, M.; Stojanovic, A.; Ale, A.; Bacchetta, C.; Cazenave, J. Preliminary Notes on Pupal Parasitism Rates of the Horse Chestnut Leafminer, Cameraria ohridella (Lepidoptera gracillariidae) in Belgrade and Istanbul. Fresen. Environ. Bull. 2018, 27, 7122–7124. [Google Scholar]
- Černý, J.; Špulák, O.; Kománek, M.; Žižková, E.; Sýkora, P. Sessile Oak (Quercus petraea [Matt.] Liebl.) and Its Adaptation Strategies in the Context of Global Climate Change: A Review. Cent. Eur. For. J. 2024, 70, 77–94. [Google Scholar] [CrossRef]
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus Robur and Quercus Petraea in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 160–163. ISBN 978-92-79-52833-0. [Google Scholar]
- Marković, Č.; Dobrosavljević, J. Review of Scolytinae (Coleoptera, Curculionidae) of Serbia. J. Entomol. Res. Soc. 2023, 25, 545–561. [Google Scholar] [CrossRef]
- Yu, X.-D.; Luo, T.-H.; Zhou, H.-Z. Distribution of Carabid Beetles among Regenerating and Natural Forest Types in Southwestern China. For. Ecol. Manag. 2006, 231, 169–177. [Google Scholar] [CrossRef]
- Forrester, G.J. The Population Ecology of Acorn Weevils and Their Influence on Natural Regeneration of Oak. Ph.D. Thesis, University of London, Imperial College, London, UK, 1990. [Google Scholar]
- Siira-Pietikäinen, A.; Haimi, J.; Siitonen, J. Short-Term Responses of Soil Macroarthropod Community to Clear Felling and Alternative Forest Regeneration Methods. For. Ecol. Manag. 2003, 172, 339–353. [Google Scholar] [CrossRef]
- Hering, E.M. Bestimmungstabellen der Blattminen von Europa Einschliesslich des Mittelmeerbeckens und Der Kanarishen Inseln, Band I, II und III.; Dr. W. Junk: Hague, The Netherlands, 1957; ISBN 9789061939818. [Google Scholar]
- Patočka, J.; Turčani, M. Lepidoptera Pupae. Central European Species; BRILL: Stenstrup, The Netherlands, 2005; ISBN 9788788757477. [Google Scholar]
- Laštůvka, A.; Zdeněk, L.; Liška, J.; Šumpich, J. Motýli a Housenky Střední Evropy V., Drobní Motýli I.; Academia: Prague, Czech Republic, 2018; ISBN 978-80-200-2852-5. [Google Scholar]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Version 2.0: Imaging to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye, Users Manual and Program Documentation; SCIRP: Wuhan, China, 1999. [Google Scholar]
- Nobis, M.; Hunziker, U. Automatic Thresholding for Hemispherical Canopy-Photographs Based on Edge Detection. Agric. For. Meteorol. 2005, 128, 243–250. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.H.; Jost, L. Unifying Species Diversity, Phylogenetic Diversity, Functional Diversity, and Related Similarity and Differentiation Measures Through Hill Numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef]
- Buzas, M.A.; Hayek, L.-A.C. Biodiversity Resolution: An Integrated Approach. Biodivers. Lett. 1996, 3, 40–43. [Google Scholar] [CrossRef]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows; Version 26.0; IBM Corp: New York, NY, USA, 2019. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to Shade, Drought, and Waterlogging of Temperate Northern Hemisphere Trees and Shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Babani, F.; Langsdorf, G. Chlorophyll Fluorescence Imaging of Photosynthetic Activity in Sun and Shade Leaves of Trees. Photosynth. Res. 2007, 93, 235–244. [Google Scholar] [CrossRef]
- Huber, A.E.; Bauerle, T.L. Long-Distance Plant Signaling Pathways in Response to Multiple Stressors: The Gap in Knowledge. J. Exp. Bot. 2016, 67, 2063–2079. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Rashmi, R.; Toppo, V.; Chole, P.B.; Banadka, A.; Sudheer, W.N.; Nagella, P.; Shehata, W.F.; Al-Mssallem, M.Q.; Alessa, F.M.; et al. Plant Secondary Metabolites: The Weapons for Biotic Stress Management. Metabolites 2023, 13, 716. [Google Scholar] [CrossRef]
- Pereira, A. Plant Abiotic Stress Challenges from the Changing Environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [PubMed]
- Keppel, G.; Anderson, S.; Williams, C.; Kleindorfer, S.; O’Connell, C. Microhabitats and Canopy Cover Moderate High Summer Temperatures in a Fragmented Mediterranean Landscape. PLoS ONE 2017, 12, e0183106. [Google Scholar] [CrossRef]
- Richter, R.; Ballasus, H.; Engelmann, R.A.; Zielhofer, C.; Sanaei, A.; Wirth, C. Tree Species Matter for Forest Microclimate Regulation During the Drought Year 2018: Disentangling Environmental Drivers and Biotic Drivers. Sci. Rep. 2022, 12, 17559. [Google Scholar] [CrossRef]
- Wolf, J.; Asch, J.; Tian, F.; Georgiou, K.; Ahlström, A. Canopy Responses of Swedish Primary and Secondary Forests to the 2018 Drought. Environ. Res. Lett. 2023, 18, 064044. [Google Scholar] [CrossRef]
- Kohler, M.; Pyttel, P.; Kuehne, C.; Modrow, T.; Bauhus, J. On the Knowns and Unknowns of Natural Regeneration of Silviculturally Managed Sessile Oak (Quercus petraea (Matt.) Liebl.) Forests—A Literature Review. Ann. Sci. 2020, 77, 101. [Google Scholar] [CrossRef]
- Modrow, T.; Kuehne, C.; Saha, S.; Bauhus, J.; Pyttel, P.L. Photosynthetic Performance, Height Growth, and Dominance of Naturally Regenerated Sessile Oak (Quercus petraea [Mattuschka] Liebl.) Seedlings in Small-Scale Canopy Openings of Varying Sizes. Eur. J. For. Res. 2020, 139, 41–52. [Google Scholar] [CrossRef]
- Jarvis, P.G. The Adaptability to Light Intensity of Seedlings of Quercus petraea (Matt.) Liebl. J. Ecol. 1964, 52, 545–571. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-43040-9. [Google Scholar]
- Kingsolver, J.G.; Huey, R.B. Size, Temperature, and Fitness: Three Rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in Global Climate Change Research: Direct Effects of Rising Temperature on Insect Herbivores. Glob. Change Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Altermatt, F. Climatic Warming Increases Voltinism in European Butterflies and Moths. Proc. R. Soc. B Biol. Sci. 2010, 277, 1281–1287. [Google Scholar] [CrossRef]
- Jaworski, T.; Hilszczański, J. The Effect of Temperature and Humidity Changes on Insects Development Their Impact on Forest Ecosystems in the Expected Climate Change. For. Res. Pap. 2013, 74, 345–355. [Google Scholar] [CrossRef]
- Rasmann, S.; Erwin, A.C.; Halitschke, R.; Agrawal, A.A. Direct and Indirect Root Defences of Milkweed (Asclepias syriaca): Trophic Cascades, Trade-Offs and Novel Methods for Studying Subterranean Herbivory. J. Ecol. 2011, 99, 16–25. [Google Scholar] [CrossRef]
- Yarnes, C.T.; Boecklen, W.J. Abiotic Mosaics Affect Seasonal Variation of Plant Resources and Influence the Performance and Mortality of a Leaf-Miner in Gambel’s Oak (Quercus gambelii, Nutt.). Ecol. Res. 2006, 21, 157–163. [Google Scholar] [CrossRef]
- Close, D.C.; McArthur, C. Rethinking the Role of Many Plant Phenolics: Protection from Photodamage Not Herbivores? Oikos 2002, 99, 166–172. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Constabel, C.P. Tannins in Plant–Herbivore Interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef]
- Barton, K.E.; Hanley, M.E. Seedling-Herbivore Interactions: Insights into Plant Defence and Regeneration Patterns. Ann. Bot. 2013, 112, 643–650. [Google Scholar] [CrossRef]
- Forkner, R.E.; Marquis, R.J.; Lill, J.T. Feeny Revisited: Condensed Tannins as Anti-Herbivore Defences in Leaf-Chewing Herbivore Communities of Quercus. Ecol. Entomol. 2004, 29, 174–187. [Google Scholar] [CrossRef]
- Ermolaev, I.V.; Zorin, D.A. Distribution of the Lime Leafminer Phyllonorycter issikii (Lepidoptera, Gracillariidae) in Natural Stands. Entomol. Rev. 2011, 91, 1088–1091. [Google Scholar] [CrossRef]
- Gripenberg, S.; Ovaskainen, O.; Elly, M.; Roslin, T. Spatial Population Structure of a Specialist Leaf-Mining Moth. J. Anim. Ecol. 2008, 77, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity Increases the Resistance of Ecosystem Productivity to Climate Extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Vasiliev, D. The Role of Biodiversity in Ecosystem Resilience. IOP Conf. Ser. Earth Environ. Sci. 2022, 1072, 012012. [Google Scholar] [CrossRef]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and Resilience of Ecosystem Functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Goodale, E.; Lalbhai, P.; Goodale, U.M.; Ashton, P.M.S. The Relationship Between Shelterwood Cuts and Crown Thinnings and the Abundance and Distribution of Birds in a Southern New England Forest. For. Ecol. Manag. 2009, 258, 314–322. [Google Scholar] [CrossRef]
- Koivula, M.; Felton, A.; Jönsson, M.; Löfroth, T.; Schei, F.H.; Siitonen, J.; Sjögren, J. Biodiversity. In Continuous Cover Forestry in Boreal Nordic Countries; Springer: Cham, Switzerland, 2025; pp. 195–220. [Google Scholar]
- Pourmajidian, M.R.; Jalilvand, H.; Fallah, A.; Hosseini, S.A.; Parsakhoo, A.; Vosoughian, A.; Rahmani, A. Effect of Shelterwood Cutting Method on Forest Regeneration and Stand Structure in a Hyrcanian Forest Ecosystem. J. For. Res. 2010, 21, 265–272. [Google Scholar] [CrossRef]
- Munck, I.A.; Yamasaki, M.; Janelle, J. Silvicultural Treatments Improve Pest and Disease Conditions of White Pine (Pinus strobus) Residual Trees and Regeneration. Front. For. Glob. Change 2023, 6, 1239835. [Google Scholar] [CrossRef]
- Leidinger, J.; Blaschke, M.; Ehrhardt, M.; Fischer, A.; Gossner, M.M.; Jung, K.; Kienlein, S.; Kózak, J.; Michler, B.; Mosandl, R.; et al. Shifting Tree Species Composition Affects Biodiversity of Multiple Taxa in Central European Forests. For. Ecol. Manag. 2021, 498, 119552. [Google Scholar] [CrossRef]
- Hannerz, M.; Hånell, B. Effects on the Flora in Norway Spruce Forests Following Clearcutting and Shelterwood Cutting. Ecol. Manag. 1997, 90, 29–49. [Google Scholar] [CrossRef]
- Nasiri, M.; Parsakhoo, A. Shelterwood Cutting System for Forest Management. J. Appl. Biol. Sci. 2012, 6, 57–60. [Google Scholar]
- Uhl, B.; Schall, P.; Bässler, C. Achieving Structural Heterogeneity and High Multi-Taxon Biodiversity in Managed Forest Ecosystems: A European Review. Biodivers. Conserv. 2024, 1–32. [Google Scholar] [CrossRef]
- Chen, K.; Li, T.; Yang, M.; Zhou, X.; Peng, C. The Effects of Environmental Factors and Plant Diversity on Forest Carbon Sequestration Vary Between Eastern and Western Regions of China. J. Clean. Prod. 2024, 437, 140371. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.L.; He, J.K.; Kong, F.M.; Yu, J.H.; Jiang, H.S. Climate and Soils Determine Aboveground Biomass Indirectly via Species Diversity and Stand Structural Complexity in Tropical Forests. For. Ecol. Manag. 2019, 432, 823–831. [Google Scholar] [CrossRef]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of Forest Management on Species Richness: Global Meta-Analysis and Economic Trade-Offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef]
- Lapin, K.; Oettel, J.; Steiner, H.; Langmaier, M.; Sustic, D.; Starlinger, F.; Kindermann, G.; Frank, G. Invasive Alien Plant Species in Unmanaged Forest Reserves, Austria. NeoBiota 2019, 48, 71–96. [Google Scholar] [CrossRef]
- Bobiec, A.; Jaszcz, E.; Wojtunik, K. Oak (Quercus Robur L.) Regeneration as a Response to Natural Dynamics of Stands in European Hemiboreal Zone. Eur. J. For. Res. 2011, 130, 785–797. [Google Scholar] [CrossRef]
- Povak, N.A.; Lorimer, C.G.; Guries, R.P. Altering Successional Trends in Oak Forests: 19 Year Experimental Results of Low- and Moderate-Intensity Silvicultural Treatments. Can. J. For. Res. 2008, 38, 2880–2895. [Google Scholar] [CrossRef]
- Maleki, K.; Zeller, L.; Pretzsch, H. Oak Often Needs to Be Promoted in Mixed Beech-Oak Stands—The Structural Processes behind Competition and Silvicultural Management in Mixed Stands of European Beech and Sessile Oak. iForest 2020, 13, 80–88. [Google Scholar] [CrossRef]
- Mölder, A.; Sennhenn-Reulen, H.; Fischer, C.; Rumpf, H.; Schönfelder, E.; Stockmann, J.; Nagel, R.-V. Success Factors for High-Quality Oak Forest (Quercus robur, Q. petraea) Regeneration. For. Ecosyst. 2019, 6, 49. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | p | |
|---|---|---|
| Total cover | 16.710 | 0.012 |
| Q. petraea | 6.759 | 0.029 |
| R. hirtus | 0.072 | 0.931 |
| T. tomentosa | 2.736 | 0.267 |
| C. pilosa | 1.079 | 0.464 |
| F. drymeia | 1.000 | 0.422 |
| F. ornus | 9.500 | 0.014 |
| A. campestre | 7.000 | 0.027 |
| C. betulus | 1.000 | 0.444 |
| C. mas | 1.000 | 0.444 |
| R. arvensis | 1.000 | 0.422 |
| Species | Canopy Cover Class | ||
|---|---|---|---|
| Open | Semi-Open | Closed | |
| Bucculatrix ulmella (Zeller, 1848) | ✓ | ✓ | |
| Caloptilia alchimiella (Scopoli, 1763) | ✓ | ✓ | ✓ |
| Coleophora flavipennella (Duponchel, 1843) | ✓ | ||
| Ectoedemia albifasciella (Heinemann, 1871) | ✓ | ✓ | |
| Phyllonorycter harrisella (Linnaeus, 1761) | ✓ | ✓ | |
| Phyllonorycter quercifoliella (Zeller, 1839) | ✓ | ✓ | ✓ |
| Phyllonorycter roboris (Zeller, 1839) | ✓ | ✓ | ✓ |
| Profenusa pygmaea (Klug, 1816) | ✓ | ✓ | ✓ |
| Stigmella basiguttella (Heinemann, 1862) | ✓ | ✓ | ✓ |
| Stigmella roborella (Johansson, 1971) | ✓ | ✓ | ✓ |
| Tischeria decidua (Wocke, 1876) | ✓ | ||
| Tischeria ekebladella (Bjerkander, 1795) | ✓ | ✓ | ✓ |
| Σ | 11 | 10 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobrosavljević, J.; Kanjevac, B.; Marković, Č. Microclimate Shifts and Leaf Miner Community Responses to Shelterwood Regeneration in Sessile Oak Forests. Forests 2025, 16, 739. https://doi.org/10.3390/f16050739
Dobrosavljević J, Kanjevac B, Marković Č. Microclimate Shifts and Leaf Miner Community Responses to Shelterwood Regeneration in Sessile Oak Forests. Forests. 2025; 16(5):739. https://doi.org/10.3390/f16050739
Chicago/Turabian StyleDobrosavljević, Jovan, Branko Kanjevac, and Čedomir Marković. 2025. "Microclimate Shifts and Leaf Miner Community Responses to Shelterwood Regeneration in Sessile Oak Forests" Forests 16, no. 5: 739. https://doi.org/10.3390/f16050739
APA StyleDobrosavljević, J., Kanjevac, B., & Marković, Č. (2025). Microclimate Shifts and Leaf Miner Community Responses to Shelterwood Regeneration in Sessile Oak Forests. Forests, 16(5), 739. https://doi.org/10.3390/f16050739