1. Introduction
Larix gmelinii (Rupr.) Rupr. ex Kuzen is a widespread species in the boreal forests of the cryolithozone, playing a crucial role in maintaining ecosystem stability in the cryolithozone and global carbon balance while growing under extreme climatic conditions [
1,
2]. The boreal forests of the cryolithozone are characterized by long, frosty winters, short growing seasons, and low precipitation levels, making
L. gmelinii a key subject for studying adaptation mechanisms to stressful environments [
3,
4]. One of the factors influencing the growth and metabolism of the tree is its interaction with epiphytic lichens, which are widely present in boreal forests. Epiphytic lichens play an important role in water cycling [
5] and nutrient cycling [
6], and they serve as a vital component in forest ecosystem food chains [
7,
8,
9,
10]. However, the specific effects of these lichens on the host plant remain poorly understood [
11,
12].
By the 19th century, researchers had already discovered that mycobiont hyphae possess the ability to penetrate plant tissues. Frank [
13] and Bonnier [
14], in their studies on the structure of crustose lichens, observed that fungal hyphae, along with the photobiont (referred to at the time as gonidia), were immersed within the upper layers of the bark. Further studies have demonstrated that hyphae of epiphytic lichens can invade plant tissues and release metabolites capable of influencing the biochemical processes of phorophyte cells [
15,
16]. Previous research [
17] documents that hyphae of the epiphytic lichen
Evernia prunastri infiltrate
Fagus sylvatica L. branches, extending to the xylem and inducing notable deformation in the periderm’s peripheral layers. In the xylem sap of
Betula pendula branches colonized by
E. prunastri, evernic acid was detected, which can accumulate in buds, leading to chloroplast deformation and leaf senescence [
18]. Similarly,
E. prunastri affected
Quercus pyrenaica by inhibiting leaf and branch growth, which is most possibly related to evernic acid migration [
19]. Comparable effects were observed in
Hypogymnia physodes. The secondary metabolites produced by this lichen species led to the decreasing viability and withering of branches of
Picea abies. Spruce branch bark samples heavily colonized by
H. physodes exhibited an increased concentration of phenolic compounds and a decline in total protein content [
20]. Epiphytic lichens are an integral component of forest ecosystems, participating in nutrient and energy cycles. They are capable of synthesizing a wide range of secondary metabolites, including usnic acid, divaricatic acid, atranorin, and other lichen substances, which can exert allelopathic effects on the host plant [
21]. For instance, usnic acid, produced by lichens, can migrate into plant tissues, which in turn can disrupt photosynthesis, cellular respiration, and oxidative phosphorylation, ultimately leading to growth inhibition and even plant death [
22]. However, allelopathic interactions between epiphytic lichens and trees can also occur in the opposite direction. In vitro studies demonstrate that certain phenolic compounds of plants can inhibit lichen growth and development [
23]. Thus, trees and epiphytic lichens form a complex interaction system determined by both individual characteristics of the organisms and external conditions.
Physcia alnophila is an epiphytic lichen that is widely present in boreal and temperate vegetation zones [
24]. It grows on the bark of trunks and branches of deciduous trees in open habitats. In boreal forest ecosystems of the cryolithozone,
P. alnophila mainly grows with
L. gmelinii as a host tree. The primary lichen acid produced by
P. alnophila is atranorin [
25]. Atranorin, a depside, is one of the most common secondary metabolites of lichens and exhibits anti-inflammatory, analgesic, wound-healing, antibacterial, antifungal, cytotoxic, antioxidant, antiviral, and immunomodulatory properties [
26]. Another study [
27] showed that ATR inhibits growth processes, provokes oxidative stress, and increases chromosomal aberration rates in
Allium fistulosum tissues, arising the question whether
P. alnophila could have some similar effects on
L. gmelinii.
Currently, the usage of metabolomic methods in plant ecophysiology enhances the understanding of biochemical adaptation mechanisms to environmental stressors such as drought, pollutants, temperature extremes, and biotic interactions [
28]. By analyzing a broad spectrum of metabolites, researchers can identify key compounds that drive plant stress responses [
29]. While metabolomic approaches to biotic stress, such as resistance to insects, fungi, bacteria, and viruses, remain understudied, certain plant metabolites have been linked to enhanced resilience against these stressors [
30]. Notably, no prior metabolomic studies have focused on the effects of epiphytic lichens on trees. Such investigations could clarify how lichen colonization disrupts host plant processes, reveal defensive metabolites produced by the host, and trace the translocation pathways of lichen-derived compounds within plant tissues. Based on these gaps and the documented phytotoxicity of lichen metabolites, we hypothesize that
Physcia alnophila could exert a certain effect on
Larix gmelinii, possibly leading to some metabolic shifts, which could also reflect on host tree parameters like growth.
The aim of this study is to investigate the allelopathic effects of the epiphytic lichen Physcia alnophila on Larix gmelinii by analyzing the migration of lichen substances into the host plant’s tissues and their impact on biochemical and growth processes. Using a combination of dendrochronology, GC-MS metabolomic profiling, and HPLC–ESI–MS/MS in the cryolithozone, this work aims to elucidate interspecies interaction mechanisms in boreal ecosystems and predict their resilience in the context of climate change.
2. Materials and Methods
2.1. Study Area
Larix gmelinii var. gmelinii tissues were sampled in mid-August 2023 from a larch forest near Yakutsk. The sampling site’s forest stand is dominated by L. gmelinii, averaging 4–6 m in height, with a crown closure of 0.5 (proportion of ground area shaded by tree crowns) and classified as site index Va (lowest productivity category). The forest floor features a dense layer of fallen needles, while the undergrowth comprises Pleurozium schreberi, Polytrichum commune, and Vaccinium vitis-idaea. Situated on a Pleistocene paleoterrace of the Lena River, the study area is approximately 9 km from Yakutsk (62°06′21.0″ N 129°40′06.2″ E). The soil in this typical larch forest consists of clay interspersed with sand layers. The region exhibits a stark continental climate, including prolonged, severe winters; brief, arid summers; early seasonal frosts; and intense spring–summer insolation levels. Temperatures fluctuate between −60 °C (winter) and +35 °C (summer), averaging −8.8 °C annually. Precipitation totals 237 mm yearly, 30% of which falls as snow, with snow cover lasting 6–7 months (mid-October to late April).
2.2. Sample Collection and Preparation
Samples of
Larix gmelinii var.
gmelinii (
Pinaceae family) and
Physcia alnophila (
Physciaceae family) were collected and categorized into three groups based on the degree of epiphytic lichen infestation (
Table 1).
Fifteen trees per group (mean height: 2.2 m) were sampled within a uniform ecotope of a middle taiga subzone larch forest in Yakutia, confined to a 150 m radius. From each tree, P. alnophila thalli (for colonized groups) and L. gmelinii tissues (needles, stem cambium, stem base, and roots) were collected. The samples were freeze-dried (Jouan LP 3 laboratory freeze-dryer, Jouan SA, Saint-Nazaire, France) and stored under vacuum in sealed, airtight containers.
2.3. Determination of Annual Tree-Ring Growth
Cross-sectional samples from the stem base were analyzed to assess annual ring growth. The samples were randomized and sanded with fine sandpaper to reveal all rings, the thinnest of which often consist of only one or two cell layers. To enhance visual contrast, dry tooth powder was applied to the cross-section surface. Radial growth was measured along the stem cross-section’s mean radius using a BRESSER Advance ICD 10×–160× Zoom Stereo-Microscope (Bresser GmbH, Rhede, Germany) fitted with a ToupCam UCMOS08000KPB (ToupTek Photonics, Hangzhou, China) camera at 10×–20× magnification. Quantification was performed using ToupView software version 3.7.
2.4. Metabolomic Profiling
For metabolomic analysis, 10 mg of L. gmelinii needles, stem cambium, and roots were homogenized and extracted in 1 mL of methanol (LCMS grade, JT Baker). The extract was then gently evaporated at 40 °C using a rotary evaporator, and the residue was dissolved in 50 μL of pyridine. Trimethylsilyl (TMS) derivatization was performed by adding 50 μL of N,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA) with 1% trimethylchlorosilane (TMCS) and heating at 100 °C for 15 min. Samples were randomized prior to analysis to ensure double-blinded results.
Gas chromatography–mass spectrometry (GC–MS) analysis was conducted using an Agilent 7820A/5975 GC–MS system (Agilent Technologies, Lexington, MA, USA) equipped with an HP-5MS column (30 m × 0.25 mm × 0.25 μm). The injector temperature was set to 250 °C, and the column temperature was programmed from 70 to 320 °C at a heating rate of 4 °C min⁻
1. Helium was used as the carrier gas at a constant flow rate of 1 mL min⁻
1. Mass spectra were acquired in full scan mode (
m/
z 50–800) with electron ionization (70 eV) and an ion source temperature of 230 °C. Data were collected using Agilent ChemStation software (version E.02.02.1431) and analyzed using the NIST 2020 library. Quantitative interpretation was performed using the internal standardization method with tricosane (C23 hydrocarbon) as the internal standard [
20]. Principal component analysis (PCA) via the MetaboAnalyst platform (
www.metaboanalyst.ca (accessed on 27 January 2025)) was employed for the statistical evaluation of metabolomic profiles.
2.5. Isolation and Determination of Lichen Substances Using HPLC–ESI–MS/MS
Air-dried lichen thalli were ground in a mortar. Approximately 10 mg of the resulting powder underwent dual extraction with 1 mL of acetone (suitable for HPLC, Sigma-Aldrich), with constant stirring for 24 h at a temperature of 20–25 °C.
2.6. Tree Sample Preparation and Extraction
Lyophilized samples of needles, stem cambium, and roots of L. gmelinii were pulverized in an IKA M20 mill (IKA-Werke GmbH & Co. KG, Staufen, Germany). Then, 0.1 g of the grounded sample was extracted twice with 1 mL of acetone (suitable for HPLC, Sigma-Aldrich). The resulting extract was filtered, and then acetone was evaporated at a Labconco CentriVap (Labconco Corp., Kansas City, MO, USA). The residue was redissolved in 0.2 mL of acetone (suitable for HPLC, Sigma-Aldrich) via ultrasonification using Elma Elmasonic S30H ultrasonic bath (Elma Schmidbauer GmbH, Singen, Germany), centrifuged at 6000× g for 5 min, and then kept at 4 °C until analysis.
High-performance liquid chromatography–electrospray ionization–mass spectrometry (HPLC–ESI–MS/MS) analysis was performed with a Shimadzu LC-30 Nexera chromatograph (Shimadzu Corp., Kyoto, Japan). For chromatographic separation, a Phenomenex Kinetex C18 column (100 × 2.1 mm, 1.7 µm) was used. The mobile phase consisted of (A) water with 0.1% (v/v) formic acid (hypergrade for LC-MS, Supelco) and (B) acetonitrile with 0.1% (v/v) formic acid (suitable for LC/MS, Supelco). Analyses were performed at 40 °C with a flow rate of 0.3 mL/min in the gradient elution mode; the percentage of B was programmed as follows: 30% (2 min)–80% (15 min)–100% (16 min)–100% (20 min)–30% (20.1 min)–30% (25 min, controller stop). The volume of the injected sample was 2 µL. Spectra of eluting substances were recorded in UV at 254 nm. After chromatographic separation, the samples were analyzed using a triple quadrupole mass-selective detector LCMS-8030 Shimadzu (Shimadzu Corp., Kyoto, Japan) with electrospray ionization (ESI) in the negative mode. The voltage on the capillary was 3.5 kV, and there was a nebulizing gas flow rate of 3 L/min, drying gas (nitrogen) temperature of 250 °C, and a drying gas flow rate of 15 L/min. Mass spectra were recorded in the range 100–800 m/z. MS/MS spectra were obtained by collision-induced dissociation of precursor ions with an energy of 30 eV. The resulting chromatograms were processed using the MZmine 4 v. 4.1.0 software. Lichen substances were identified by comparing their retention time (polarity dependent) and pseudo-molecular and fragment ion m/z values with authentic standards from the BIN RAS collection. The quantification of lichen substances was carried out using the calibration graphs, obtained by plotting peak area external standards versus the known concentration of each compound. The peak areas of lichen substances in plant extracts were determined using reconstructed ion chromatograms.
2.7. Statistical Procedures and Data Analysis
Results are expressed as arithmetic mean ± standard error of the mean (SEM). All analyses were conducted using 15 biological replicates (
n = 15) for each group. Normality was assessed using the Shapiro–Wilk test. Group comparisons were performed using the Kruskal–Wallis H-test (one-way ANOVA on ranks), with the post hoc Dunn’s test used for multiple comparisons (
p < 0.05, Bonferroni-adjusted). Data analysis was conducted in R 4.3.2 (“Eye Holes”). The Pearson linear correlation coefficient was calculated using the AnalystSoft, StatPlus software, version 2007. Principal component analysis (PCA) was performed on 45 observations for 118 metabolites in needles, 85 in cambium, and 111 in roots, using GC–MS data normalized to an internal standard (tricosane C23). Samples were normalized by median with a base-10 logarithmic transformation, followed by auto-scaling (mean-centering and division by the standard deviation of each variable) on the publicly available MetaboAnalyst resource (
www.metaboanalyst.ca (accessed on 27 January 2025)). Differential metabolite abundance between treatment groups was assessed using the non-parametric Kruskal–Wallis ANOVA test. Metabolites with a false discovery rate (FDR)-adjusted
p-value < 0.05 were considered statistically significant. To compare metabolomic profiles between treatment groups, we employed PERMANOVA (Permutational Multivariate Analysis of Variance), eigenvalues, and relative/cumulative variance calculated via MetaboAnalyst.ca. Loading plots and heatmaps were used to evaluate relationships between organism functioning and exposure factors. For heatmap generation and analysis, hierarchical clustering was performed using normalized data with auto scale features applied for standardization. The Euclidean distance measure and Ward’s clustering method were utilized.
3. Results
The collected samples of
L. gmelinii exhibited varying degrees of colonization by the epiphytic lichen
P. alnophila. The width of
P. alnophila thalli was used as the criterion for sample differentiation as follows: <1 cm (L1) and >1 cm (L2) (
Figure 1). Samples of
L. gmelinii without visible colonization by
P. alnophila or other lichens were considered the control group (L0). The morphological assessment of
L. gmelinii needles and wood samples showed no infection. All trees displayed comparable external traits (needles, bark, branches, height, crown structure), irrespective of lichen colonization. All
L. gmelinii specimens had comparable heights, with an average of 222 cm (
Figure 2a).
The average age of
L. gmelinii trees, determined through the tree-ring analysis, differed between groups, as follows: 22 ± 3 years (L0), 68 ± 14 years (L1), and 71 ± 17 years (L2) (
Figure 2b). This indicates that the average age of trees in lichen-colonized groups (L1,
p-value = 0.0001; L2,
p-value = 0.0001) was 46–49 years older, despite statistically similar average tree heights.
No significant differences in radial growth were observed among groups during the initial five years of tree development (L0; L1,
p-value = 0.18; L2,
p-value = 0.07), suggesting favorable growth conditions during early developmental stages for each group (
Figure 2c). However, in the last five years of tree life, despite identical weather and habitat conditions, a statistically significant reduction in average annual radial growth was observed in groups L1 and L2, with decreases of 27% (
p-value = 0.0463) and 51% (
p-value = 0.0001), respectively, compared to L0 (
Figure 2d).
The apical and radial growth of L. gmelinii trees colonized with P. alnophila was lower in experimental groups. This could be caused by the allelopathic effect of the epiphytic lichen, which disturbs biochemical processes in tree organisms.
The chromatographic analysis of
P. alnophila samples colonizing
L. gmelinii revealed that the primary secondary metabolite was atranorin (ATR). The atranorin content in the thalli of the studied samples ranged from 40 to 50 mg g
−1 of dry weight (DW) (
Figure 3). In contrast, the rhizines of the lichen contained 8–10 times less ATR than the thalli.
Figure 4 shows the ATR content in the stem cambium, roots, and needles of
L. gmelinii growing in the cryolithozone. In the control group (L0), trace amounts of ATR were detected in the stem cambium, roots, and needles. ATR was found in the stem cambium, roots, and needles of
L. gmelinii in both the lightly (L1) and heavily (L2) colonized groups. Notably, the highest concentrations of ATR in the needles and roots of
L. gmelinii were recorded in group L2, reaching 26.3 and 36.3 µg g
−1 DW, respectively. At the site of
P. alnophila colonization, no statistically significant changes in ATR content were observed in the stem cambium, which measured 20.6 and 26.3 µg g
−1 DW in groups L1 and L2, respectively. ATR content in the stem cambium (
r = −0.68;
p = 0.002), roots (
r = −0.62;
p = 0.004), and needles (
r = −0.40;
p = 0.08) of
L. gmelinii negatively correlated with radial growth over the last 5 years of life. Notably, the correlation between ATR content in the needles and radial growth was statistically insignificant, possibly due to the annual renewal of needles. In contrast, no correlation was observed between ATR content in the stem cambium (
r = −0.11;
p = 0.66), root (
r = −0.03;
p = 0.89), or needles (
r = −0.06;
p = 0.80) and radial growth during the first 5 years of the tree’s life.
This study demonstrates that the primary lichen substance of P. alnophila is ATR, which migrates into L. gmelinii tissues at the lichen colonization site. After penetrating the stem cambium, ATR translocates via conductive tissues to the root system, where it accumulates, followed by upward transport into the needles through the xylem flow. The systemic presence of ATR in all L. gmelinii organs likely disrupts biochemical processes, leading to the inhibition of radial and apical tree growth.
Stem cambium metabolomes from
L. gmelinii with varying degrees of colonization by epiphytic lichen
P. alnophila were analyzed. Score and loading plots (45 observations, 85 metabolites;
Figure 5) were constructed for statistical evaluation. The dataset was processed using principal component analysis (PCA).
The points corresponding to the metabolomes of
L. gmelinii stem cambium clustered based on the degree of lichen colonization (
Figure 5a), with variances of 28% and 12% for components 1 and 2, respectively. Eigenvalues, relative variance, and cumulative variance for the first four principal components (PC) for the stem cambium are provided in
Appendix A Table A2. This clustering suggests that the metabolism of
L. gmelinii is influenced by
P. alnophila, as confirmed by the significance level for experimental groups relative to the control (
p-value = 0.002;
Appendix A Table A1).
The loading plot (
Figure 5b) highlights mono- and disaccharides, polyols, fatty acids, phenolic/nitrogenous compounds, organic/inorganic acids, sterols, and terpenes as primary contributors to metabolome clustering. Control group samples exhibited higher concentrations of these metabolites, whereas colonized groups showed elevated polyols, sterols, terpenes, phenolics, and disaccharides.
The metabolites that were the most significant for this differentiation are shown in the heatmap (
Figure 6). The content of arabitol (a major lichen polyol) and trehalose (a lichen-specific disaccharide) in the stem cambium of
L. gmelinii increased in groups L1 and L2 compared to the control group, suggesting the migration of lichen metabolites into the plant’s conductive tissues. Additionally, increased concentrations of polyols, arabinose, disaccharides, sitosterol, and shikimic acid were observed in the stem cambium of
L. gmelinii in groups L1 and L2. However, the contents of monosaccharides, organic and inorganic acids, and chiro-inositol were lower in these groups compared to L0.
The metabolomic analysis of root samples from
L. gmelinii with varying degrees of
P. alnophila colonization was conducted. Score and loading plots were constructed for 45 observations of 111 metabolites (
Figure 7), and the dataset was processed using PCA.
The points corresponding to root metabolomes are clustered based on the degree of lichen colonization (
Figure 7a), with variances of 31% and 13% for components 1 and 2, respectively. Eigenvalues, relative variance, and cumulative variance for the first four PCs for the stem cambium are provided in
Appendix A Table A2. This clustering indicates that the metabolism of
L. gmelinii roots is influenced by
P. alnophila metabolites, as confirmed by the significance level for experimental groups relative to the control (
p-value = 0.001;
Appendix A Table A1).
The loading plot (
Figure 7b) identifies mono- and disaccharides, polyols, fatty acids, phenolic/nitrogenous compounds, organic/inorganic acids, sterols, and terpenes as key clustering factors. Control roots had higher metabolite levels, while colonized groups (L1/L2) showed increased polyols, sterols, terpenes, phenolics, fatty acids, and disaccharides.
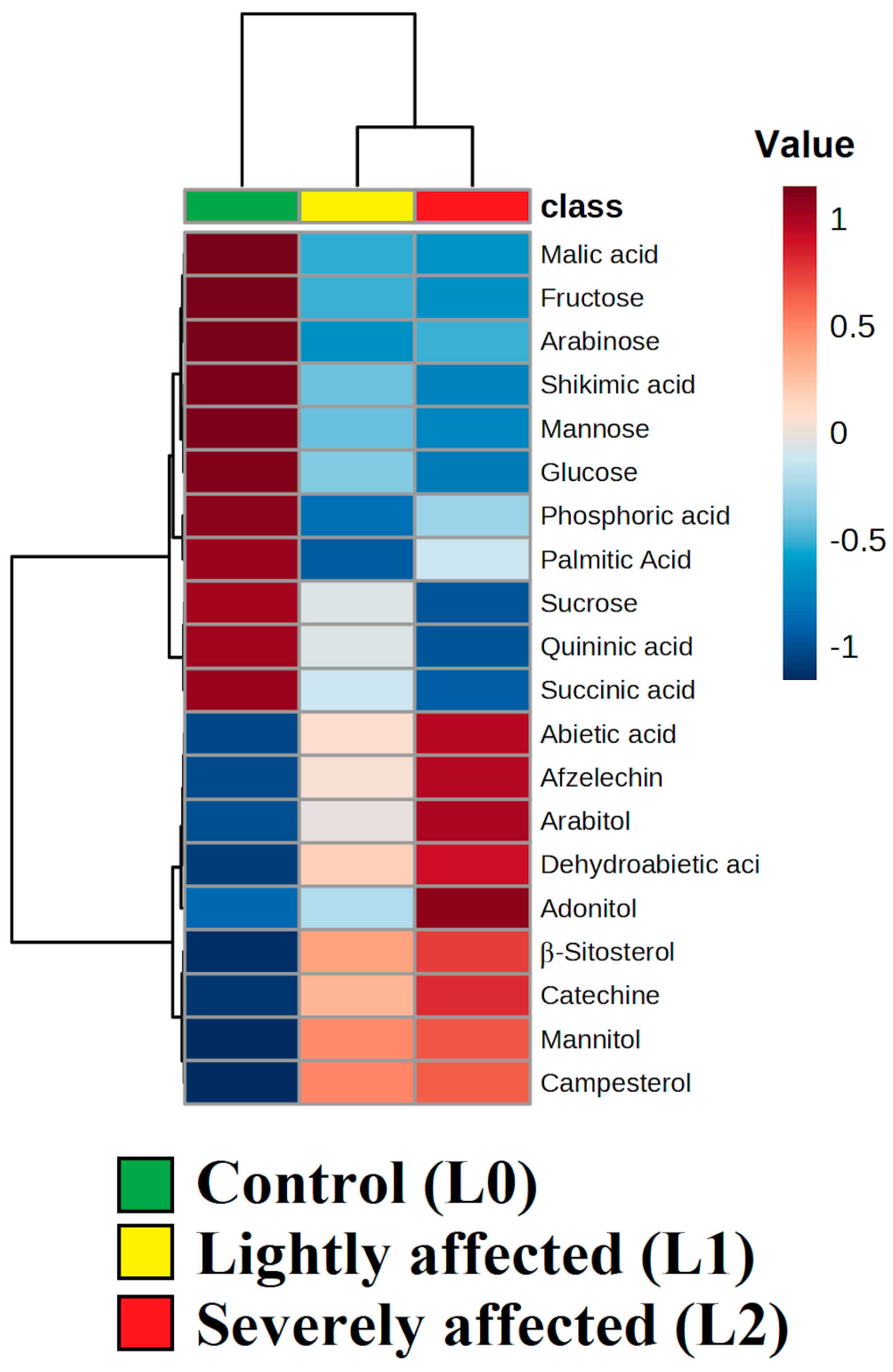
The heatmap (
Figure 8) highlights the most significant metabolites for this differentiation. The content of mannitol, adonitol, and arabitol (major lichen polyols) in the roots of
L. gmelinii increased in groups L1 and L2 compared to the control group, suggesting the migration of lichen metabolites into the root system. Additionally, increased concentrations of terpenes, sterols, and phenolic compounds were observed in the roots of
L. gmelinii in groups L1 and L2. However, the contents of monosaccharides, inorganic and organic acids, shikimate pathway products, sucrose, and palmitic acid were lower in these groups compared to L0.
The metabolomic analysis of needle samples from
L. gmelinii with varying degrees of
P. alnophila colonization was performed. Score and loading plots were constructed for 45 observations of 118 metabolites (
Figure 9), and the dataset was processed using PCA.
Needle metabolomes clustered by lichen colonization intensity (
Figure 9a), with principal components 1 and 2 explaining 16% and 8% variance, respectively. Eigenvalues, relative variance, and cumulative variance for the first four PCs for the stem cambium are provided in
Appendix A Table A2. This clustering suggests that the metabolism of
L. gmelinii needles is influenced by
P. alnophila metabolites, as confirmed by the significance level for experimental groups relative to the control (
p-value = 0.001;
Appendix A Table A1).
The loading plot (
Figure 9b) emphasizes mono- and disaccharides, polyols, fatty acids, phenolic/nitrogenous compounds, organic/inorganic acids, sterols, and terpenes as primary discriminants. Control group needles showed higher metabolite concentrations, while colonized groups displayed elevated polyols, sterols, terpenes, fatty acids, and disaccharides.
The heatmap (
Figure 10) highlights the most significant metabolites for this differentiation. The content of mannitol, arabitol (major lichen polyols), and trehalose (a lichen-specific disaccharide) in the needles of
L. gmelinii increased in groups L1 and L2 compared to the control group, suggesting the migration of lichen metabolites into the photosynthetic tissues. Additionally, increased concentrations of terpenes, organic acids involved in the tricarboxylic acid (TCA) cycle, fatty acids, and sucrose were observed in the needles of
L. gmelinii in groups L1 and L2. However, the contents of monosaccharides, polyols, taxifolin, and serine were lower in these groups compared to L0.
This study demonstrates that ATR penetration disrupts metabolic processes in L. gmelinii tissues across all examined organs, including impairment of the TCA cycle and oxidative phosphorylation. This is accompanied by elevated synthesis of protective compounds such as phenolic compounds, sterols, and terpenes, which enhance organismal stress tolerance and survival but fail to restore tree growth processes.
The allelopathic effect of P. alnophila on L. gmelinii involves the migration of ATR from lichen thalli through its rhizines into the tree’s cambium. ATR is subsequently transported through the phloem to the roots, where it accumulates and then is redistributed upward via the xylem to the needles. This systemic translocation of ATR disrupts key metabolic pathways in the host, including the TCA cycle and oxidative phosphorylation. In response, L. gmelinii exhibits a generalized defense response characterized by increased synthesis of terpenes, sterols, phenolic compounds, and fatty acids. These metabolic shifts correlate with reduced radial and apical growth in lichen-colonized trees, even in mature individuals, suggesting that chronic ATR exposure compromises growth efficiency despite the tree’s age.
4. Discussion
The epiphytic lichen
P. alnophila contains atranorin (ATR), a β-orcinol depside [
25]. The highest ATR content was detected in the thalli of
P. alnophila, with significantly lower levels observed in the rhizines, which directly contact the substrate. ATR synthesis via the acetyl–polymalonyl pathway occurs in the upper cortex of the lichen; thus, its highest concentration is localized in this part of the thallus. The biosynthesis pathway initiates with the carboxylation of acetyl-CoA by acetyl-CoA carboxylase, producing malonyl-CoA. This compound then serves as the substrate for polyketide synthase, a multienzyme complex that drives polyketide formation. This process advances through decarboxylative condensation, enabling the attachment of an acetyl-CoA unit to generate an eight-carbon precursor. Cyclization of this intermediate yields structural variants such as orsellinic acid or its methylated form, methyl-3-orsellinate. The latter acts as a pivotal building block for depsides, including the lichen metabolite atranorin. Studies reveal that atranorin synthesis is stimulated by oxygen and NADH, suggesting the involvement of oxidase and dehydrogenase enzymes. These enzymes are proposed to catalyze the conversion of intermediates into hemmatomyl aldehyde, a direct precursor in the atranorin pathway. This oxidative and reductive interplay highlights the regulatory role of environmental and enzymatic factors in shaping the final metabolite [
31,
32]. Despite this, it should be noted that the transport of ATR to the rhizines is observed, likely serving a protective role against biotic stressors [
33].
Analysis revealed ATR in the stem cambium tissues of
L. gmelinii beneath
P. alnophila thalli in groups L1 and L2, indicating the migration of lichen substances into the substrate. Rhizines of foliose epiphytic lichens, including
Physcia species, typically penetrate only the superficial layers without deep invasion into bark tissues [
34]. During substrate colonization, rhizine growth damages bark tissues, potentially enabling ATR diffusion into the cambium. This study used relatively young trees (with an average height of 2.2 m) with underdeveloped bark protective layers, which may facilitate lichen metabolite migration. ATR content in the stem cambium did not differ significantly between L1 and L2 groups, suggesting further downward transport of lichen substances via phloem to tree roots. Maximum ATR concentrations were detected in
L. gmelinii roots, indicating preferential accumulation in the root system. Subsequent upward xylem transport of ATR from roots to needles was observed, with ATR detected in needles, confirming systemic distribution. A similar migration mechanism has been proposed for
E. esorodiosa on
L. gmelinii [
22]. Lichen metabolites are known to inhibit growth and biochemical processes upon plant cell penetration [
12,
35]. ATR exhibits phytotoxic effects on
Allium fistulosum [
27], manifesting as growth inhibition, oxidative stress elevation, and chromosomal aberrations. ATR disrupts oxidative phosphorylation and inhibits respiration via mitochondrial membrane interactions in mouse hepatocytes [
36]. While ATR demonstrates low cytotoxicity in human and animal cells, it induces apoptosis in both healthy and cancerous cells [
26]. Thus, ATR may disrupt growth and biochemical processes in host organisms.
Some studies question the presence of the direct allelopathy of secondary lichen metabolites [
37] and, so, propose that the differences in lichen colonization and metabolites may stem from pre-existing variations in tree health or site conditions. Control trees (L0) might thrive in better microhabitats, resisting lichens [
38], while stressed trees (L1/L2) in harsher sites could accumulate metabolites passively due to higher lichen infestation. Selecting the trees for sampling within a single forest site with a 150 m radius as the base for experimental design mitigated the possible environmental differences between microhabitats. This study demonstrated a significant negative correlation between ATR content in the stem cambium (
r = −0.68;
p = 0.002) and roots (
r = −0.62;
p = 0.004) of
L. gmelinii and radial growth over the last 5 years, suggesting an allelopathic effect of the epiphytic lichen
P. alnophila on tree growth processes.
Metabolomic analysis revealed altered biochemical profiles in stem cambium, roots, and needles of
L. gmelinii colonized by
P. alnophila. At the lichen attachment site, cambium metabolomes of L1 and L2 groups had clustered separately from the control group (L0). The lack of separation between L1 and L2 cambium metabolomes suggests similar impacts regardless of the colonization severity, likely due to comparable ATR levels. Prolonged colonization by the epiphytic lichen in group L2 does not result in greater ATR accumulation in the stem cambium, as ATR further migrates through vascular tissues into the root system and needles of
L. gmelinii. The annual renewal of cambium cells and continuous ATR migration via vascular tissues maintain similar ATR levels in the stem cambium of L1 and L2 groups, provoking a uniform response in
L. gmelinii. Loading plots highlight terpenes and sterols as major contributors to metabolome differentiation (
Figure 5b), indicative of non-specific biotic stress responses [
39] triggered by ATR migration. The activation of terpene synthesis in plant tissues, particularly in coniferous trees, is a primary response to biotic stress, where terpenes act as pathogen inhibitors [
40]. The authors of [
41] showed that terpenes and phenolic compounds play a key role in the defensive responses of
Pinus resinosa against insect and fungal infections. Transcriptomic, metabolomic, and proteomic studies revealed that
Bursaphelenchus xylophilus infection activates metabolic pathways linked to terpenoid, phenylpropanoid, flavonoid, and carbohydrate biosynthesis in
Pinus koraiensis [
42]. Plant sterols enhance stress resistance by strengthening cell membrane cohesion, altering membrane permeability, and serving as precursors to steroid hormones that regulate metabolic processes [
43,
44]. Elevated arabitol (a lichen polyol) and ATR in colonized cambium suggest metabolite transfer via rhizine penetration into periderm and subsequent vascular transport. Another study showed trehalose migration from the tissues of the arbuscular mycorrhizal fungus
Glomus intraradices into
Medicago truncatula roots [
45]. Increased disaccharide levels likely reflect protective responses to periderm damage [
46]. Reduced monosaccharides and phosphoric/organic acids in colonized cambium imply disrupted TCA cycle and oxidative phosphorylation, potentially due to ATR interference. Atranorin can penetrate cell membranes due to its chemical structure, likely disrupting mitochondrial function by uncoupling the mitochondrial membrane potential. This leads to impaired respiration, oxidative phosphorylation, reduced ATP synthesis, elevated reactive oxygen species (ROS) levels, and subsequent cell death. Notably, potato virus Y infection caused reduced carbohydrate synthesis in tissues of various potato cultivars [
47]. Mycorrhizal fungi were shown to reduce sugar synthesis in
Glycine max tissues [
48]. Additionally, mycorrhizal fungi induced the strong accumulation of free amino acids, the overexpression of enzymes/transporters linked to sugar/amino acid/fatty acid catabolism, and the activation of autophagy and biotic stress-related genes in
Epipactis helleborine [
49]. Conversely, increased arabinose levels with decreased fructose, glucose, mannose, xylose, and allose levels suggest enhanced arabinogalactan synthesis for cell wall reinforcement and cytokine-mediated tissue repair [
50,
51].
Further migration of ATR from the epiphytic lichen
P. alnophila downward through the phloem flow was observed in the root system of
L. gmelinii, where ATR accumulation occurs. According to the results of metabolomic analysis using the PCA of
L. gmelinii roots, the separation of the metabolomes of groups L1 and L2 from the control group (L0) was revealed. It should be noted that the metabolomes of roots of
L. gmelinii in groups L1 and L2 clustered according to the degree of colonization by the epiphytic lichen
P. alnophila, which may be associated with varying concentrations of ATR. An increase in lichen-specific metabolites, such as arabitol, adonitol, and mannitol, was detected in the root system of
L. gmelinii in groups L1 and L2 when colonized by
P. alnophila, indicating their migration through the phloem into the roots. In the root system, a similar response to that observed in the stem cambium of
L. gmelinii during colonization by the epiphytic lichen was noted, but it was more pronounced, as evident in the loading plot. A significant increase in the content of terpenes, sterols, and phenolic compounds was observed in the roots of
L. gmelinii in groups L1 and L2, likely due to a non-specific response aimed at membrane stabilization and the suppression of oxidative stress [
52,
53], both resulting from ATR migration. Additionally, disruptions in metabolic processes such as the TCA cycle and oxidative phosphorylation were recorded in the root system, contributing to the slowdown of tree growth processes [
22]. Biotic stress in plants induces ROS accumulation [
54], which can disrupt redox balance and impair the TCA cycle. Caterpillar herbivory on
Glycine max increased lipid peroxidation and hydroxyl anion formation, depleting antioxidants and disrupting energy metabolism [
55]. Oxidative stress and ROS generation reduced glycolysis and TCA cycle activity in heterotrophic
Arabidopsis cells [
56].
From the root system, where the accumulation of lichen substances was observed in groups L1 and L2, migration occurred upwards with xylem flow into the needles of
L. gmelinii, where ATR was detected. On the score plot constructed from the metabolomic analysis of
L. gmelinii needles, a more distinct separation of groups based on the degree of colonization by the epiphytic lichen
P. alnophila was observed. This is likely due to the higher sensitivity of photosynthetic organs to the ATR influence. In the needle tissues of
L. gmelinii, an increase in lichen-specific metabolites such as arabitol, mannitol, and trehalose was also detected in groups L1 and L2, indicating their migration through the xylem. The response of
L. gmelinii needles to
P. alnophila colonization was characterized by an increase in terpenes and sterols, the synthesis of which is a primary defense mechanism against biotic stress in coniferous trees [
53]. A distinctive feature of the metabolic response in
L. gmelinii needles to
P. alnophila colonization was the significant increase in fatty acids (FA), such as linolenic, linoleic, and stearic acids. That increase in FA contents in needles may be associated with a compensatory response to oxidative stress caused by ATR [
57]. The increase in FA may also be attributed to the signaling function of these substances, which serve as a substrate for lipoxygenase and the subsequent octadecanoid (oxylipin) pathways, leading to the synthesis of phytohormones such as jasmonic acid and methyl jasmonates [
58]. These phytohormones are involved in regulating numerous processes in plants, including wound healing and defense against pathogens [
59]. The reduction in monosaccharide content was observed in the cambium, roots, and needles of
L. gmelinii. Such effects may be caused by impaired photosynthetic activity due to ATR migration into the photosynthetic organs during colonization by the epiphytic lichen
P. alnophila, leading to a slowdown in tree growth processes. At the same time, in the needles of
L. gmelinii colonized by the epiphytic lichen, activation of the synthesis of organic acids (citric, malic, and succinic acids) involved in the TCA cycle was observed. This may indicate an intensification of respiratory processes in the organism to produce ATP, which is necessary for the synthesis of protective metabolites and compensation for disrupted biochemical processes. The decrease in monosaccharide concentration in the needles of
L. gmelinii may also be caused by the increased intensification of glycolytic processes for pyruvate synthesis and its oxidation to acetyl-CoA, which is the main substrate for the TCA cycle. This could also lead to the increased synthesis of organic acids such as citric, malic, and succinic acids. Studies on
Pinus radiata infected by
Fusarium circinatum showed sustained TCA activity via the activation of energy-producing pathways, namely, fermentation and fatty acid β-oxidation [
60].
The migration of atranorin (ATR) from the epiphytic lichen Physcia alnophila into Larix gmelinii tissues and its accumulation in the stem cambium, roots, and needles lead to slowed apical and radial tree growth due to disruptions in metabolic processes, while simultaneously activating non-specific defense responses. Thus, the epiphytic lichen P. alnophila exerts direct allelopathic effects on its host, L. gmelinii, growing in the boreal forests of the cryolithozone. These findings deepen our understanding of epiphytic lichen–plant interactions, demonstrating that lichen metabolites can directly influence host physiology through allelopathy. This study underscores the role of epiphytic lichens as potential bioindicators of host plant stress and ecosystem health, particularly in young or vulnerable tree populations, which could enhance ecological monitoring efforts in the cryolithozone’s boreal forests.
5. Conclusions
The present study demonstrates the allelopathic impact of the epiphytic lichen P. alnophila on Larix gmelinii, a keystone species and the primary forest-forming tree in the boreal forests of the cryolithozone. It was established that the main secondary metabolite of the epiphytic lichen P. alnophila is atranorin (ATR). ATR was found to migrate into the tissues of the host plant L. gmelinii, penetrating the cambium and subsequently moving downward through the phloem flow into the root system, where its accumulation was observed. From there, it was transported upward through the xylem flow into the needles of L. gmelinii. The migration of ATR into the tissues of L. gmelinii led to a significant reduction in the radial growth of trees during the last five years of their life (by 27%–51% in groups L1 and L2) and a slowdown in apical growth, despite statistically similar tree heights. Trees colonized by the epiphytic lichen were 46–49 years older, which may indicate a phytotoxic effect as the most likely cause of the diminished L. gmelinii apical growth in experimental groups.
Metabolomic analysis revealed profound changes in the biochemical profiles of the cambium, roots, and needles of L. gmelinii. Disruptions in the tricarboxylic acid cycle and oxidative phosphorylation, as well as increased concentrations of protective metabolites (terpenes, sterols, and phenolic compounds), confirm the plant’s adaptive response to biotic stress. The migration of lichen polyols (arabitol, mannitol, and adonitol) into larch tissues indicates the systemic distribution of allelopathic substances through the plant’s vascular systems.
These findings highlight the significant role of epiphytic lichens as important ecological mediators in boreal forests, modulating the physiology of woody plants under the extreme conditions of the cryolithozone. Their pervasive influence on tree growth, nutrient cycling, and stress adaptation underscores their importance in maintaining the structural and functional integrity of these ecosystems. It was established that lichen-derived allelochemicals like atranorin not only induce phytotoxic effects but also trigger systemic biochemical responses in host plants, reshaping metabolic networks critical for survival. These results could provide a better understanding of the mechanisms of interspecies interactions in boreal ecosystems, particularly the dual role of epiphytic lichens as both stressors and regulators of forest health, and could predict their resilience in the context of climate change.
By bridging symbiotic and antagonistic interactions, epiphytic lichens emerge as dynamic components of boreal biodiversity, driving evolutionary adaptations while potentially exacerbating vulnerabilities to environmental shifts. This study underscores how lichen-mediated biochemical disruptions, such as TCA cycle inhibition and oxidative stress, may alter carbon allocation patterns in trees, a critical factor for boreal forests facing permafrost thaw and climate change disturbances. These insights provide a framework to predict how lichen–plant interactions may amplify or buffer climate-driven impacts on boreal carbon sinks, suggesting the urgent need to integrate epiphytic communities into ecosystem resilience models.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}