
Standardized Protocol for Somatic Embryogenesis from Vegetative Organs in Hybrid Sweetgum (L. styraciflua × L. formosana)


Abstract
1. Introduction
2. Materials and Methods
2.1. Development of In Vitro Plantlets from Mature Hybrid Sweetgum Leaves
2.2. Induction of Callus
2.3. Proliferation of Callus
2.4. Maturation of Somatic Embryos
2.5. Somatic Embryo Germination
2.6. Statistical Analysis
3. Results
3.1. Different Explant Types Induced Callus in Three Genotypes of Hybrid Sweetgum
3.2. Effect of PGRs on Callus Proliferation
3.3. Somatic Embryo Maturation
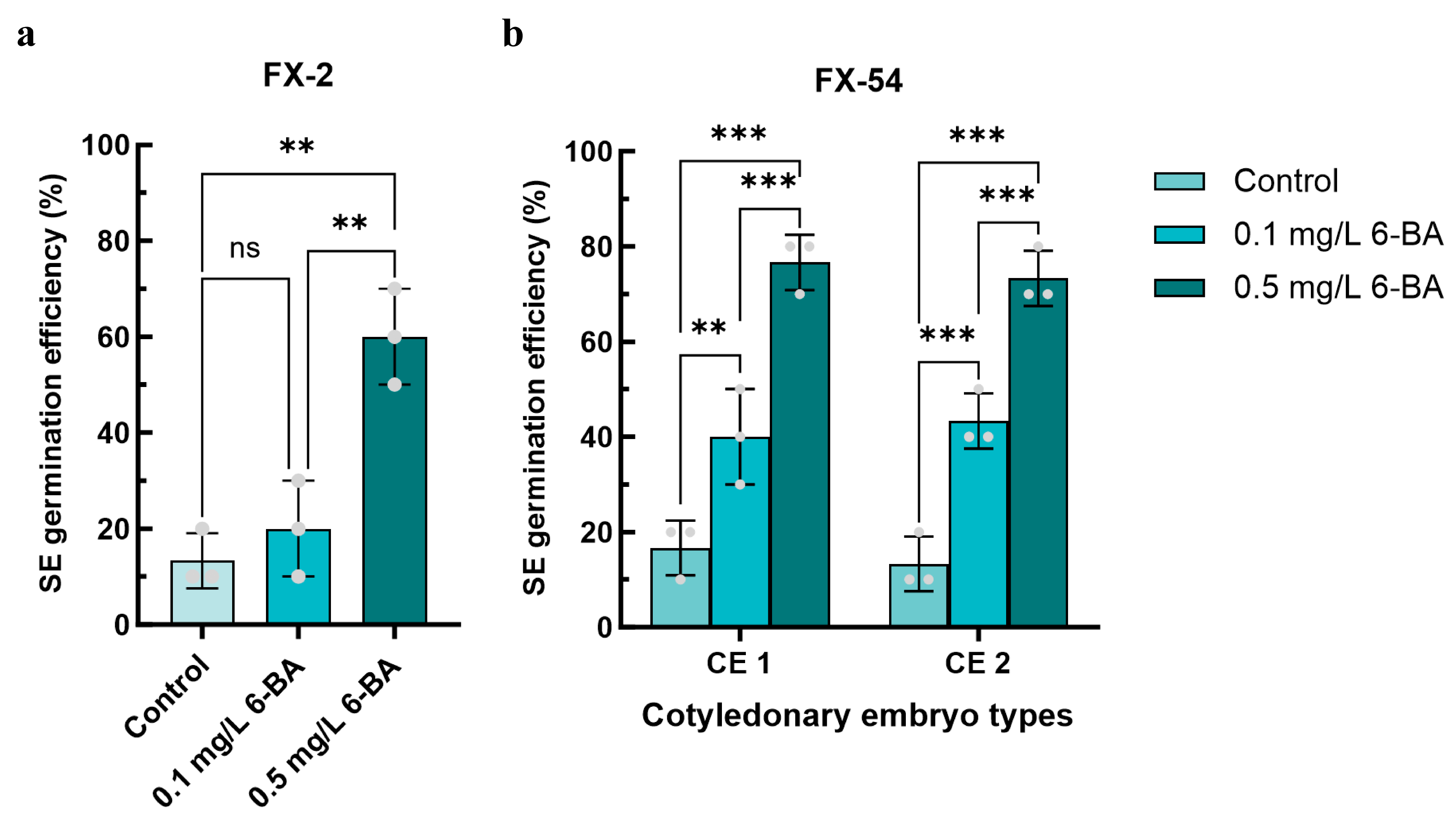
3.4. Effect of 6-BA on Somatic Embryo Germination into Seedlings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fan, Y.; Li, H.; Li, Y.; Bao, F.; Zhan, D.; Pang, Z.; Zhao, J.; Zhang, J. Construction of the First High-Density Genetic Linkage Map and QTL Mapping of Shikimic Acid Content in Liquidambar. Forests 2024, 15, 1662. [Google Scholar] [CrossRef]
- Lewis, J.; Will, R.; Zhai, L.; Murray, B.; Blazier, M.; Joshi, O. Productivity of hybrid sweetgum (Liquidambar formosana x styraciflua) plantations compared to native sweetgum (L. Styraciflua) in the Western Gulf Region of south-central U.S. New For. 2024, 55, 1753–1769. [Google Scholar] [CrossRef]
- Merkle, S. The ups and downs of developing hybrid sweetgum varieties for the U.S. bioenergy and pulp and paper industries: A 20-year case study. In The Fifth International Conference of the IUFRO Unit 2.09.02: Somatic Embryogenesis and Other Vegetative Propagation Technologies; University of Coimbra: Coimbra, Portugal, 2018. [Google Scholar]
- Lu, S.; Merkle, S.A. Enhancing hybrid Liquidambar somatic seedling production using a temporary immersion bioreactor. Trees 2021, 35, 503–512. [Google Scholar] [CrossRef]
- Kang, H.; Lee, C.; Kwon, S.; Park, J.; Kang, K.; Shim, D. Comparative transcriptome analysis during developmental stages of direct somatic embryogenesis in Tilia amurensis Rupr. Sci. Rep.-Uk 2021, 11, 6359. [Google Scholar] [CrossRef] [PubMed]
- Karami, O.; Philipsen, C.; Rahimi, A.; Nurillah, A.R.; Boutilier, K.; Offringa, R. Endogenous auxin maintains embryonic cell identity and promotes somatic embryo development in Arabidopsis. Plant J. 2023, 113, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Berenguer, E.; Carneros, E.; Pérez-Pérez, Y.; Gil, C.; Martínez, A.; Testillano, P.S. Small molecule inhibitors of mammalian GSK-3β promotein vitro plant cell reprogramming and somatic embryogenesis in crop and forest species. J. Exp. Bot. 2021, 72, 7808–7825. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hu, Y.; Wang, R.; Feng, K.; Di, J.; Feng, T.; Cao, F. Transcriptome and physiological analysis highlights the hormone, phenylpropanoid, and photosynthesis effects on early somatic embryogenesis in Ginkgo biloba. Ind. Crop. Prod. 2023, 203, 117176. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, D.; Shi, P.; Htwe, Y.M.; Yu, Q.; Huang, L.; Zhou, H.; Liu, L.; Wang, Y. Cell wall lignification may be necessary for somatic embryogenesis of areca palm (Areca catechu). Sci. Hortic.-Amst. 2023, 307, 111538. [Google Scholar] [CrossRef]
- Spinoso-Castillo, J.L.; Bello-Bello, J.J. In Vitro Stress-Mediated Somatic Embryogenesis in Plants; Springer: New York, NY, USA, 2022; Volume 2527, pp. 223–235. [Google Scholar]
- Su, Y.H.; Tang, L.P.; Zhao, X.Y.; Zhang, X.S. Plant cell totipotency: Insights into cellular reprogramming. J. Integr. Plant Biol. 2021, 63, 228–243. [Google Scholar] [CrossRef]
- Cai, Y.; Tang, L.; Chen, H.; Li, Y.; Liu, R.; Chen, J. Somatic embryogenesis in Rosa chinensis cv. ‘Old Blush’. Plant Cell Tissue Organ Cult. 2022, 149, 645–656. [Google Scholar] [CrossRef]
- Corredoira, E.; Ballester, A.; Ibarra, M.; Vieitez, A.M. Induction of somatic embryogenesis in explants of shoot cultures established from adult Eucalyptus globulus and E. saligna × E. maidenii trees. Tree Physiol. 2015, 35, 678–690. [Google Scholar] [CrossRef]
- San-José, M.C.; Corredoira, E.; Martínez, M.T.; Vidal, N.; Valladares, S.; Mallón, R.; Vieitez, A.M. Shoot apex explants for induction of somatic embryogenesis in mature Quercus robur L. trees. Plant Cell Rep. 2010, 29, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Lopes, M.L.; Canhoto, J.M. Somatic embryogenesis induction system for cloning an adult Cyphomandra betacea (Cav.) Sendt. (tamarillo). Trees 2011, 25, 1009–1020. [Google Scholar] [CrossRef]
- Vendrame, W.; Holliday, C.; Merkle, S. Clonal propagation of hybrid sweetgum (Liquidambar styraciflua × L. formosana) by somatic embryogenesis. Plant Cell Rep. 2001, 20, 691–695. [Google Scholar] [CrossRef]
- Dai, J.; Vendrame, W.A.; Merkle, S.A. Enhancing the productivity of hybrid yellow-poplar and hybrid sweetgum embryogenic cultures. Vitr. Cell. Dev. Biol.-Plant 2004, 40, 376–383. [Google Scholar] [CrossRef]
- Merkle, S.; Montello, P.; Kormanik, T.; Le, H. Propagation of novel hybrid sweetgum phenotypes for ornamental use via somatic embryogenesis. Propag. Ornam. Plants 2010, 10, 220–226. [Google Scholar]
- Qi, S.; Zhao, R.; Yan, J.; Fan, Y.; Huang, C.; Li, H.; Chen, S.; Zhang, T.; Kong, L.; Zhao, J.; et al. Global Transcriptome and Coexpression Network Analyses Reveal New Insights Into Somatic Embryogenesis in Hybrid Sweetgum (Liquidambar styraciflua × Liquidambar formosana). Front. Plant Sci. 2021, 12, 751866. [Google Scholar] [CrossRef]
- Zhao, R.; Qi, S.; Cui, Y.; Gao, Y.; Jiang, S.; Zhao, J.; Zhang, J.; Kong, L. Transcriptomic and physiological analysis identifies a gene network module highly associated with brassinosteroid regulation in hybrid sweetgum tissues differing in the capability of somatic embryogenesis. Hortic. Res.-Engl. 2022, 9, uhab047. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Qi, S.; Wang, X.; Zhao, J.; Zhang, J.; Li, B.; Zhang, Y.; Liu, X.; Yuan, W. In Vitro Tetraploid Induction from Leaf and Petiole Explants of Hybrid Sweetgum (Liquidambar styraciflua × Liquidambar formosana). Forests 2017, 8, 264. [Google Scholar] [CrossRef]
- Witham, F.H.; Blaydes, B.F.; Devlin, R.M. Experiments in Plant Physiology; Van Nostrand Reinhold: New York, NY, USA, 1971. [Google Scholar]
- Gresshoff, P.M.; Doy, C.H. Development and differentiation of haploid Lycopersicon esculentum (tomato). Planta 1972, 107, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Sommer, H.E.; Brown, C.L. Embryogenesis in Tissue Cultures of Sweetgum. For. Sci. 1980, 26, 257–260. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Agsays with Tohaoco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Merkle, S.A.; Battle, P.J. Enhancement of embryogenic culture initiation from tissues of mature sweetgum trees. Plant Cell Rep. 2000, 19, 268–273. [Google Scholar] [CrossRef]
- Hazubska-Przybył, T.; Ratajczak, E.; Obarska, A.; Pers-Kamczyc, E. Different Roles of Auxins in Somatic Embryogenesis Efficiency in Two Picea Species. Int. J. Mol. Sci. 2020, 21, 3394. [Google Scholar] [CrossRef]
- Martínez, M.; Corredoira, E. Recent Advances in Plant Somatic Embryogenesis: Where We Stand and Where to Go. Int. J. Mol. Sci. 2024, 25, 8912. [Google Scholar] [CrossRef]
- Yang, J.; Yang, D.; Lü, W.; Zhang, X.; Ma, M.; Liu, G.; Jiang, J.; Li, C. Somatic embryogenesis and plant regeneration in Betula platyphalla. J. For. Res. 2021, 32, 937–944. [Google Scholar] [CrossRef]
- Zou, S.; Yao, X.; Zhong, C.; Li, D.; Wang, Z.; Huang, H. Recurrent somatic embryogenesis and development of somatic embryos in Akebia trifoliata (Thunb.) Koidz (Lardizabalaceae). Plant Cell Tissue Organ Cult. (PCTOC) 2019, 139, 493–504. [Google Scholar] [CrossRef]
- Yuan, J.; Chao, Y.; Han, L. Uncovering a Phenomenon of Active Hormone Transcriptional Regulation during Early Somatic Embryogenesis in Medicago sativa. Int. J. Mol. Sci. 2022, 23, 8633. [Google Scholar] [CrossRef]
- Wójcik, A.M.; Wójcikowska, B.; Gaj, M.D. Current Perspectives on the Auxin-Mediated Genetic Network that Controls the Induction of Somatic Embryogenesis in Plants. Int. J. Mol. Sci. 2020, 21, 1333. [Google Scholar] [CrossRef]
- Solórzano-Cascante, P.; Sánchez-Chiang, N.; Jiménez, V.M. Explant Type, Culture System, 6-Benzyladenine, Meta-Topolin and Encapsulation Affect Indirect Somatic Embryogenesis and Regeneration in Carica papaya L. Front. Plant Sci. 2018, 9, 1769. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | 2,4-D(mg/L) | 6-BA(mg/L) |
|---|---|---|
| 1 | 1.0 | 0.25 |
| 2 | 1.0 | 0.50 |
| 3 | 2.0 | 0.25 |
| 4 | 2.0 | 0.50 |
| 5 | 3.0 | 0.25 |
| 6 | 3.0 | 0.50 |
| Treatment | 2,4-D(mg/L) | NAA(mg/L) | 6-BA(mg/L) |
|---|---|---|---|
| 1 | 0.5 | - | 0.25 |
| 2 | 1.0 | - | 0.5 |
| 3 | - | 0.5 | 0.25 |
| 4 | - | 1.0 | 0.5 |
| Genotype | Treatment | Callus Induction Efficiency (%) | ||
|---|---|---|---|---|
| Leaf | Petiole | Stem Segment | ||
| FX-2 | 1.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 80 ± 10 bcdeBCD | 86.67 ± 5.77 bcABC | 86.67 ± 5.77 bcABC |
| 1.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 90 ± 10 abAB | 96.67 ± 5.77 aA | 96.67 ± 5.77 aA | |
| 2.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 86.67 ± 5.77 bcABC | 80 ± 10 bcdeBCD | 86.67 ± 5.77 bcABC | |
| 2.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 83.33 ± 5.77 bcdBC | 76.67 ± 5.77 cdefBCD | 86.67 ± 5.77 bcABC | |
| 3.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 70 ± 10 cdefCDE | 66.67 ± 5.77 defgCDE | 63.33 ± 5.77 efgCDE | |
| 3.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 56.67 ± 5.77 fgDE | 46.67 ± 5.77 gE | 56.67 ± 5.77 fgDE | |
| FX-12 | 1.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 53.33 ± 5.77 deC | 50 ± 10 eC | 53.33 ± 5.77 deC |
| 1.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 80 ± 10 abAB | 86.67 ± 5.77 aA | 70 ± 10 abcdeABC | |
| 2.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 76.67 ± 15.28 abcABC | 63.33 ± 15.28 bcdeABC | 66.67 ± 5.77 bcdeABC | |
| 2.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 70 ± 10 abcdeABC | 60 ± 10 cdeBC | 63.33 ± 5.77 bcdeABC | |
| 3.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 73.33 ± 5.77 abcdABC | 50 ± 10 eC | 60 ± 10 cdeBC | |
| 3.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 63.33 ± 11.55 bcdeABC | 66.67 ± 15.28 bcdeABC | 56.67 ± 15.28 deBC | |
| FX-54 | 1.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 66.67 ± 5.77 bcdCD | 76.67 ± 5.77 bcBC | 60 ± 10 cdCD |
| 1.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 93.33 ± 5.77 aA | 90 ± 10 aAB | 80 ± 10 bBC | |
| 2.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 76.67 ± 5.77 bcBC | 73.33 ± 5.77 bcCD | 70 ± 10 bcCD | |
| 2.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 66.67 ± 5.77 bcdCD | 76.67 ± 5.77 bcBC | 73.33 ± 5.77 bcCD | |
| 3.0 mg/L 2,4-D + 0.25 mg/L 6-BA | 63.33 ± 5.77 bcdCD | 66.67 ± 5.77 bcdCD | 60 ± 10 cdCD | |
| 3.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 50 ± 10 dD | 60 ± 10 cdCD | 63.33 ± 5.77 bcdCD | |
| Genotype | Treatment | Callus Proliferation Coefficient | |
|---|---|---|---|
| Callus I | Callus II | ||
| FX-2 | 0.5 mg/L 2,4-D + 0.25 mg/L 6-BA | 1.88 ± 0.06 bB | 1.84 ± 0.05 bBC |
| 1.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 2.12 ± 0.01 aA | 2.10 ± 0.13 aA | |
| 0.5 mg/L NAA + 0.25 mg/L 6-BA | 1.58 ± 0.07 cD | 1.63 ± 0.04 cD | |
| 1.0 mg/L NAA + 0.5 mg/L 6-BA | 1.69 ± 0.10 cCD | 1.65 ± 0.03 cCD | |
| FX-12 | 0.5 mg/L 2,4-D + 0.25 mg/L 6-BA | 1.92 ± 0.04 bcBC | 1.88 ± 0.03 cdBC |
| 1.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 2.31 ± 0.19 aA | 2.06 ± 0.06 bB | |
| 0.5 mg/L NAA + 0.25 mg/L 6-BA | 1.75 ± 0.02 dCD | 1.60 ± 0.04 eD | |
| 1.0 mg/L NAA + 0.5 mg/L 6-BA | 1.92 ± 0.07 bcBC | 1.81 ± 0.04 cdCD | |
| FX-54 | 0.5 mg/L 2,4-D + 0.25 mg/L 6-BA | 1.93 ± 0.04 cC | 1.84 ± 0.07 dCD |
| 1.0 mg/L 2,4-D + 0.5 mg/L 6-BA | 2.58 ± 0.04 aA | 2.38 ± 0.05 bB | |
| 0.5 mg/L NAA + 0.25 mg/L 6-BA | 1.66 ± 0.04 eE | 1.45 ± 0.05 fF | |
| 1.0 mg/L NAA + 0.5 mg/L 6-BA | 1.82 ± 0.04 dCD | 1.78 ± 0.07 dD | |
| Callus Type | The Number of Mature Embryos/g | ||
|---|---|---|---|
| Genotype | Callus I | Callus II | |
| FX-2 | 0 ± 0 cC | 22 ± 5 aA | |
| FX-12 | 0 ± 0 cC | 0 ± 0 cC | |
| FX-54 | 17 ± 3 bAB | 13 ± 5 bB | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Fan, Y.; Kang, J.; Qi, S.; Bao, F.; Li, Y.; Cheng, L.; Zhan, D.; Pang, Z.; Zhao, J.; et al. Standardized Protocol for Somatic Embryogenesis from Vegetative Organs in Hybrid Sweetgum (L. styraciflua × L. formosana). Forests 2025, 16, 670. https://doi.org/10.3390/f16040670
Li H, Fan Y, Kang J, Qi S, Bao F, Li Y, Cheng L, Zhan D, Pang Z, Zhao J, et al. Standardized Protocol for Somatic Embryogenesis from Vegetative Organs in Hybrid Sweetgum (L. styraciflua × L. formosana). Forests. 2025; 16(4):670. https://doi.org/10.3390/f16040670
Chicago/Turabian StyleLi, Hongxuan, Yingming Fan, Jindian Kang, Shuaizheng Qi, Fen Bao, Ying Li, Long Cheng, Dingju Zhan, Zhenwu Pang, Jian Zhao, and et al. 2025. "Standardized Protocol for Somatic Embryogenesis from Vegetative Organs in Hybrid Sweetgum (L. styraciflua × L. formosana)" Forests 16, no. 4: 670. https://doi.org/10.3390/f16040670
APA StyleLi, H., Fan, Y., Kang, J., Qi, S., Bao, F., Li, Y., Cheng, L., Zhan, D., Pang, Z., Zhao, J., & Zhang, J. (2025). Standardized Protocol for Somatic Embryogenesis from Vegetative Organs in Hybrid Sweetgum (L. styraciflua × L. formosana). Forests, 16(4), 670. https://doi.org/10.3390/f16040670