
Rainfall Partitioning Dynamics in Xerophytic Shrubs: Interplays Between Self-Organization and Meteorological Drivers


 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
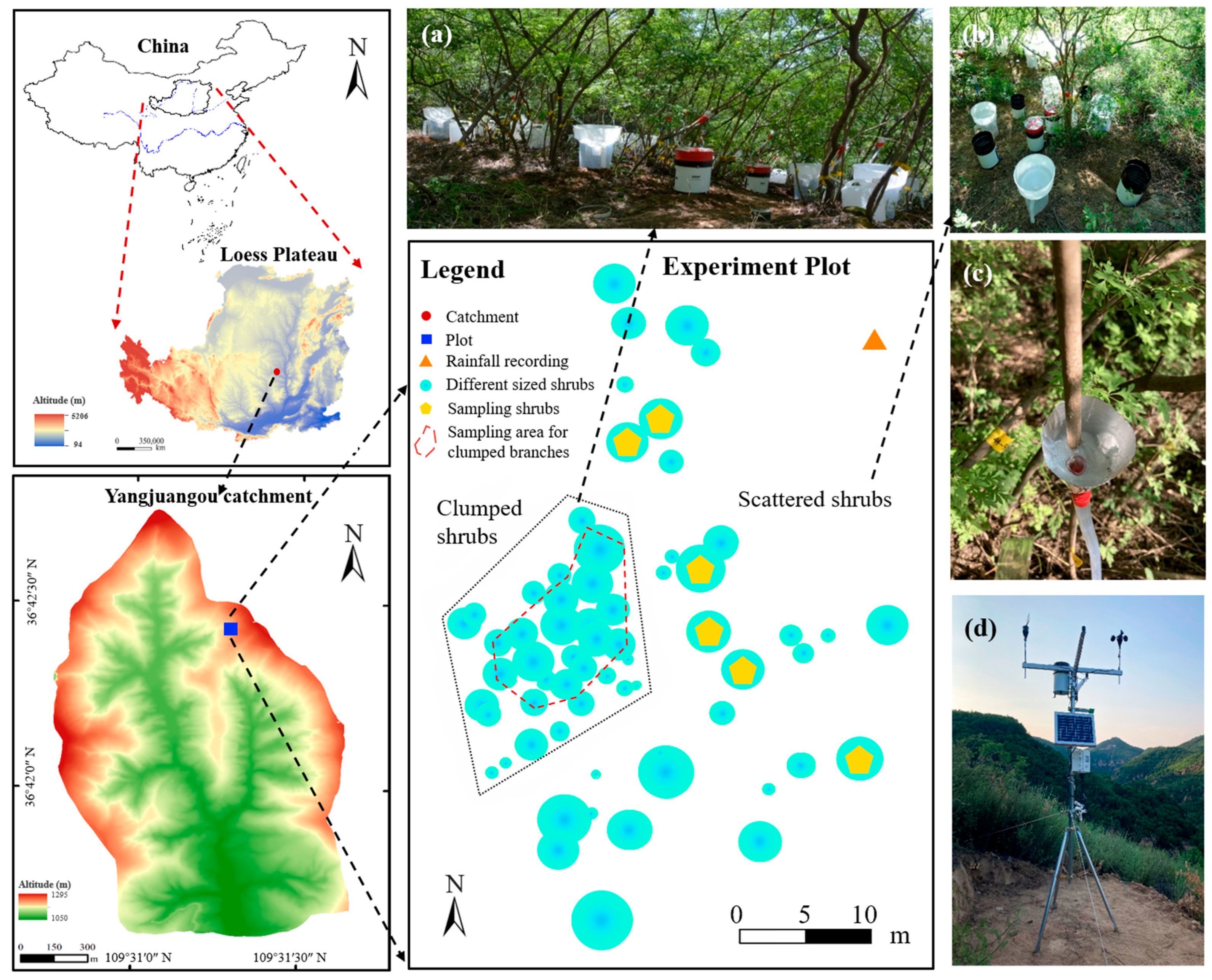
2.1. Study Sites
2.2. Meteorological Recordings
2.3. Plant Trait Measurements
2.4. Rainfall Partitioning Quantifications
2.5. Data Analysis
3. Results
3.1. Meteorological Conditions
3.2. Quantity and Efficiency of Rainfall Partitioning
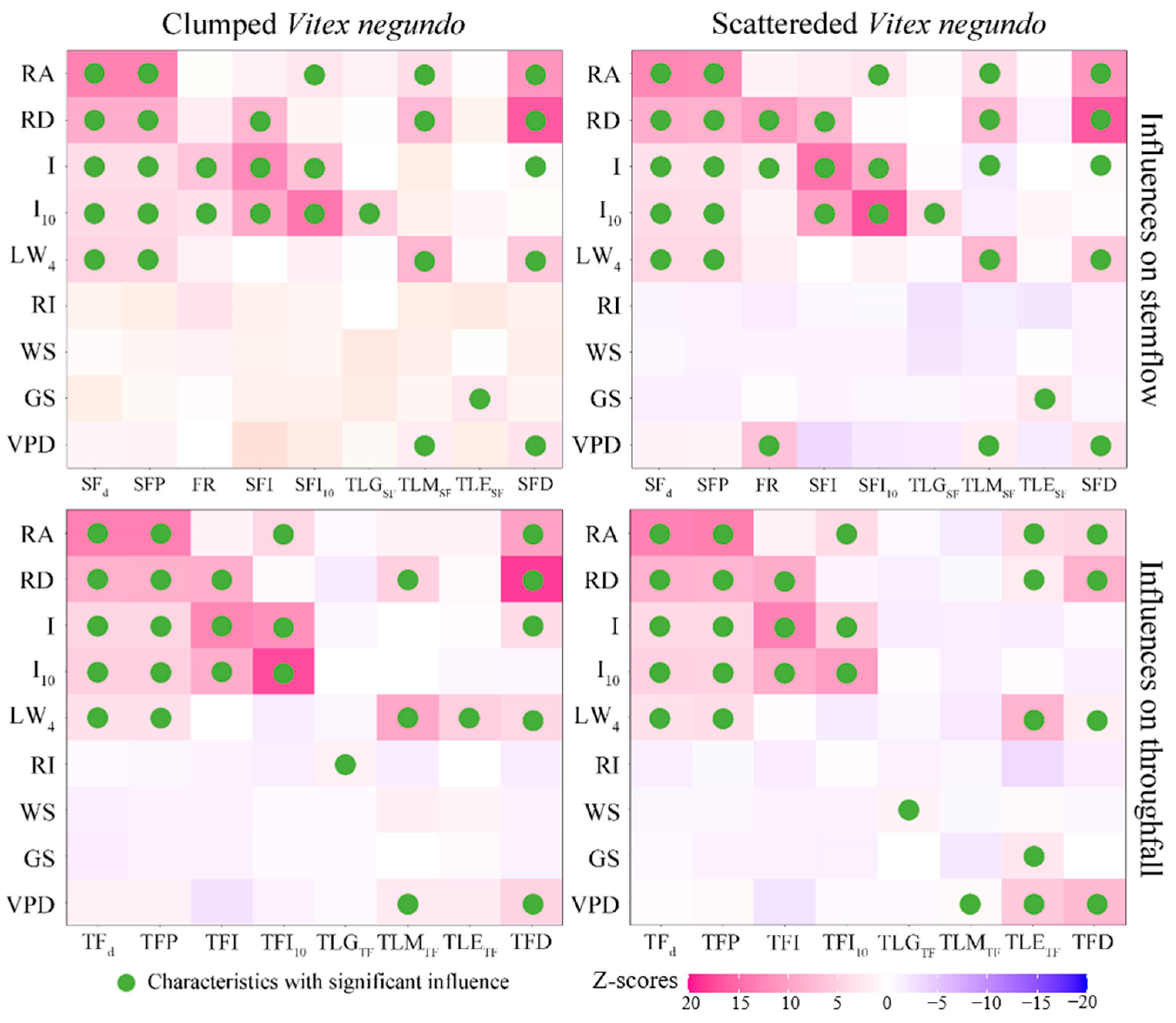
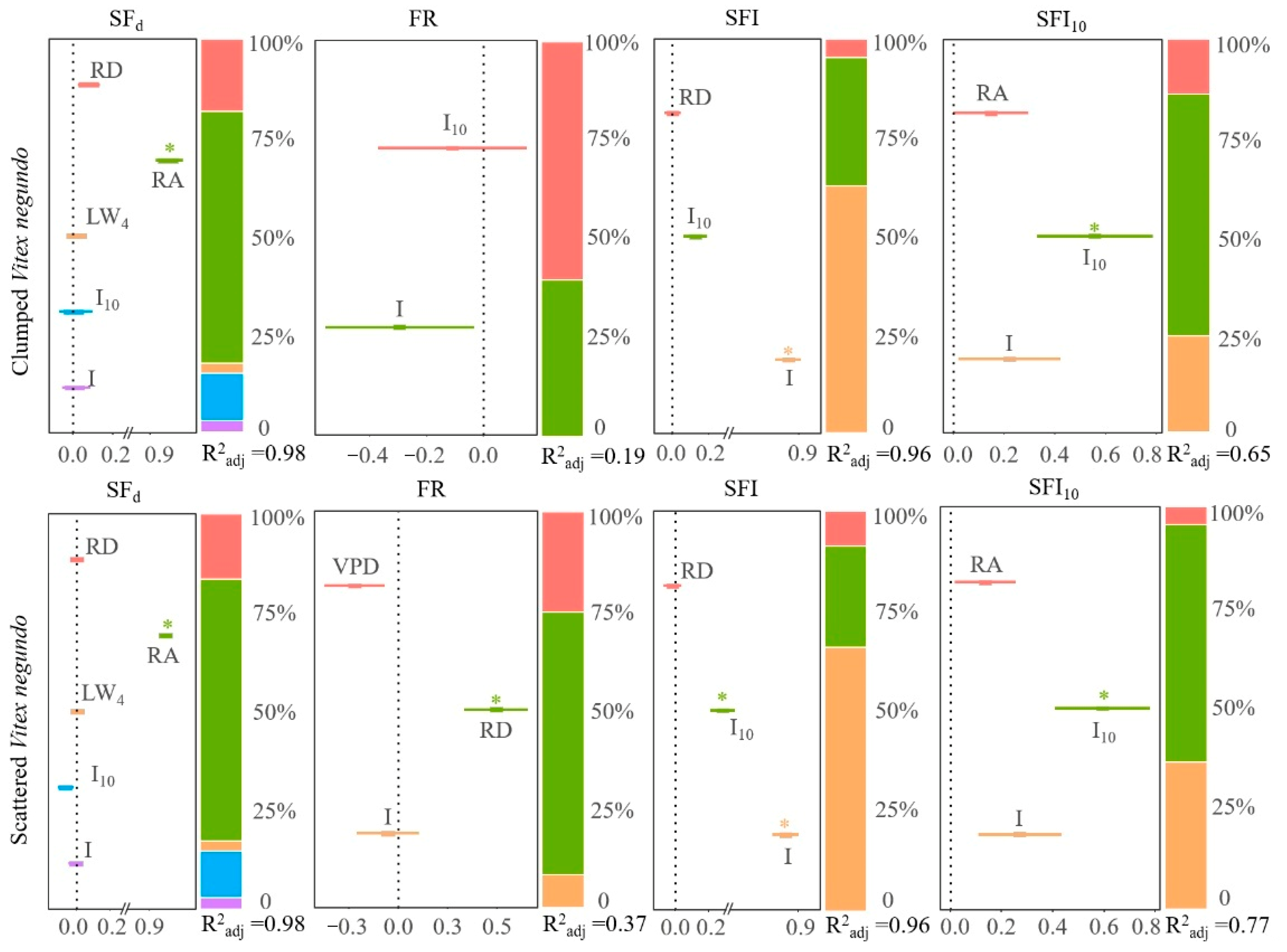
3.3. Meteorological Influences on Rainfall Partitioning
4. Discussion
4.1. Distinct Rainfall Harvesting Strategies Explained by Plant Traits
4.2. Meteorological Impacts on Rainfall Partitioning Strategy
4.3. Limitations and Future Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Symbols
| Abbreviation | Description |
| SFd | stemflow depth |
| TFd | throughfall depth |
| NPd | net precipitation depth |
| SF% | stemflow percentage |
| TF% | throughfall percentage |
| NP% | net precipitation percentage |
| SFV | stemflow volume of individual branch |
| SFI | average stemflow intensity |
| SFI10 | 10 min maximum stemflow intensity |
| TFI | average throughfall intensity |
| TFI10 | 10 min maximum throughfall intensity |
| FR | funneling ratio |
| SFP | stemflow productivity |
| TFP | throughfall productivity |
| TLG | time lag of stemflow generation |
| TLM | time lag of stemflow maximization |
| TLE | time lag of stemflow ending |
| SFD | stemflow duration |
| TFD | throughfall duration |
| RA | rainfall amount |
| RD | rainfall duration |
| RI | rainfall intervals |
| I | average rainfall intensity |
| I10 | 10 min maximum rainfall intensity |
| WS | wind speed |
| GS | gust speed |
| AH | air relative humidity |
| AT | air temperature |
| LW4 | relative humidity at leaf surface in 4 h prior to rain |
| VPD | vapor pressure deficit |
| SR | solar radiation |
| BD | branch basal diameter |
| BA | branch angle |
| BL | branch length |
| BMLC | leaf biomass of clumped shrubs |
| BMSC | stem biomass of clumped shrubs |
| BMLS | leaf biomass of scattered shrubs |
| BMSS | stem biomass of scattered shrubs |
| AGB | aboveground biomass |
| TLA | total leaf area |
| SLA | specific leaf area |
| SA | stem surface area of individual branch |
| CA | canopy area |
| LAI | leaf area index |
References
- Guo, L.; Mount, G.J.; Hudson, S.; Lin, H.; Levia, D. Pairing geophysical techniques improves understanding of the near-surface Critical Zone: Visualization of preferential routing of stemflow along coarse roots. Geoderma 2020, 357, 113953. [Google Scholar] [CrossRef]
- Yuan, C.; Yue, X.P.; Zhang, Y.F.; Zhang, Y.; Hu, Y.T.; Tang, Q.; Guo, L.; Wang, S.; Duan, X.W.; Xiang, W.H.; et al. Nutrient enrichment driven by canopy rainfall redistribution: Mechanism, quantification, and pattern. Sci. China Earth Sci. 2024, 67, 1529–1544. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Allen, S.T. What We Know About Stemflow's Infiltration Area. Front. For. Glob. Change 2020, 3, 61. [Google Scholar] [CrossRef]
- An, J.; Gao, G.; Yuan, C.; Pinos, J.; Fu, B. Inter- and intra-event rainfall partitioning dynamics of two typical xerophytic shrubs in the Loess Plateau of China. Hydrol. Earth Syst. Sci. 2022, 26, 3885–3900. [Google Scholar] [CrossRef]
- Herwitz, S.R. Interception storage capacities of tropical rainforest canopy trees. J. Hydrol. 1985, 77, 237–252. [Google Scholar] [CrossRef]
- Levia, D.F.; Hudson, S.A.; Llorens, P.; Nanko, K. Throughfall drop size distributions: A review and prospectus for future research. Wiley Interdiscip. Rev. Water 2017, 4, e1225. [Google Scholar] [CrossRef]
- Levia, D.F.; Germer, S. A review of stemflow generation dynamics and stemflow-environment interactions in forests and shrublands. Rev. Geophys. 2015, 53, 673–714. [Google Scholar] [CrossRef]
- Schume, H.; Jost, G.; Katzensteiner, K. Spatio-temporal analysis of the soil water content in a mixed Norway spruce (Picea abies (L.) Karst.)–European beech (Fagus sylvatica L.) stand. Geoderma 2003, 112, 273–287. [Google Scholar] [CrossRef]
- Molina, A.J.; Gonzalez-Sanchis, M.; Biel, C.; del Campo, A.D. Ecohydrological turnover in overstocked Aleppo pine plantations: Does the effect of thinning, in relation to water, persist at the mid-term? For. Ecol. Manag. 2021, 483, 118781. [Google Scholar] [CrossRef]
- Jochheim, H.; Luettschwager, D.; Riek, W. Stem distance as an explanatory variable for the spatial distribution and chemical conditions of stand precipitation and soil solution under beech (Fagus sylvatica L.) trees. J. Hydrol. 2022, 608, 127629. [Google Scholar] [CrossRef]
- Wang, Y.; Shao, M.A.; Zhu, Y.; Liu, Z. Impacts of land use and plant characteristics on dried soil layers in different climatic regions on the Loess Plateau of China. Agric. For. Meteorol. 2011, 151, 437–448. [Google Scholar]
- Yuan, C.; Guo, L.; Levia, D.F.; Rietkerk, M.; Fu, B.; Gao, G. Quantity or Efficiency: Strategies of Self-Organized Xerophytic Shrubs to Harvest Rain. Water Resour. Res. 2022, 58, e2022WR032008. [Google Scholar]
- Yuan, C.; Gao, G.Y.; Fu, B.J.; He, D.M.; Duan, X.W.; Wei, X.H. Temporally dependent effects of rainfall characteristics on inter- and intra-event branch-scale stemflow variability in two xerophytic shrubs. Hydrol. Earth Syst. Sci. 2019, 23, 4077–4095. [Google Scholar] [CrossRef]
- Rodrigues, A.F.; Terra, M.C.N.S.; Mantovani, V.A.; Cordeiro, N.G.; Ribeiro, J.P.C.; Guo, L.; Nehren, U.; Mello, J.M.; Mello, C.R. Throughfall spatial variability in a neotropical forest: Have we correctly accounted for time stability? J. Hydrol. 2022, 608, 127632. [Google Scholar]
- Tanaka, N.; Levia, D.F.; Igarashi, Y.; Yoshifuji, N.; Tanaka, K.; Tantasirin, C.; Nanko, K.; Suzuki, M.; Kumagai, T.O. What factors are most influential in governing stemflow production from plantation-grown teak trees? J. Hydrol. 2017, 544, 10–20. [Google Scholar]
- González, J.C.M.; Lopez-Gonzalez, C.; Marquez-Linares, M.A.; Perez-Verdin, G. Response of water yield to silvicultural treatments in a temperate forest in northern Mexico. PLoS ONE 2023, 18, e0291094. [Google Scholar]
- Exler, J.L.; Moore, R.D. Quantifying throughfall, stemflow and interception loss in five vegetation communities in a maritime raised bog. Agric. For. Meteorol. 2022, 327, 109202. [Google Scholar]
- Yue, K.; De Frenne, P.; Fornara, D.A.; Van Meerbeek, K.; Li, W.; Peng, X.; Ni, X.; Peng, Y.; Wu, F.; Yang, Y.; et al. Global patterns and drivers of rainfall partitioning by trees and shrubs. Glob. Change Biol. 2021, 27, 3350–3357. [Google Scholar]
- Zhang, Y.; Yuan, C.; Chen, N.; Levia, D.F. Rainfall partitioning by vegetation in China: A quantitative synthesis. J. Hydrol. 2023, 617, 128946. [Google Scholar] [CrossRef]
- Yuan, C.; Gao, G.; Fu, B. Stemflow of a xerophytic shrub (Salix psammophila) in northern China: Implication for beneficial branch architecture to produce stemflow. J. Hydrol. 2016, 539, 577–588. [Google Scholar]
- Whitworth-Hulse, J.I.; Magliano, P.N.; Zeballos, S.R.; Gurvich, D.E.; Spalazzi, F.; Kowaljow, E. Advantages of rainfall partitioning by the global invader Ligustrum lucidum over the dominant native Lithraea molleoides in a dry forest. Agric. For. Meteorol. 2020, 290, 108013. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Wang, X.P.; Hu, R.; Pan, Y.X. Meteorological influences on process-based spatial-temporal pattern of throughfall of a xerophytic shrub in arid lands of northern China. Sci. Total Environ. 2018, 619–620, 1003–1013. [Google Scholar]
- Yang, X.G.; Chen, L.; Wang, L.; Wang, X.; Gu, J.L.; Qu, W.J.; Song, N.P. Dynamic rainfall-partitioning relationships among throughfall, stemflow, and interception loss by Caragana intermedia. J. Hydrol. 2019, 574, 980–989. [Google Scholar] [CrossRef]
- Zheng, J.; Fan, J.; Zhang, F.; Yan, S.; Xiang, Y. Rainfall partitioning into throughfall, stemflow and interception loss by maize canopy on the semi-arid Loess Plateau of China. Agric. Water Manag. 2018, 195, 25–36. [Google Scholar] [CrossRef]
- Zhao, W.; Ji, X.; Jin, B.; Du, Z.; Zhang, J.; Jiao, D.; Zhao, L. Experimental partitioning of rainfall into throughfall, stemflow and interception loss by Haloxylon ammodendron, a dominant sand-stabilizing shrub in northwestern China. Sci. Total Environ. 2023, 858, 159928. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Siegert, C.M.; Levia, D.F.; Scheick, C.E. Effects of wind-driven rainfall on stemflow generation between codominant tree species with differing crown characteristics. Agric. For. Meteorol. 2011, 151, 1277–1286. [Google Scholar] [CrossRef]
- Fan, J.; Oestergaard, K.T.; Guyot, A.; Lockington, D.A. Measuring and modeling rainfall interception losses by a native Banksia woodland and an exotic pine plantation in subtropical coastal Australia. J. Hydrol. 2014, 515, 156–165. [Google Scholar] [CrossRef]
- Nanko, K.; Hotta, N.; Suzuki, M. Evaluating the influence of canopy species and meteorological factors on throughfall drop size distribution. J. Hydrol. 2006, 329, 422–431. [Google Scholar] [CrossRef]
- Yan, T.; Wang, Z.; Liao, C.; Xu, W.; Wan, L. Effects of the morphological characteristics of plants on rainfall interception and kinetic energy. J. Hydrol. 2021, 592, 125807. [Google Scholar] [CrossRef]
- Klaassen, W. Evaporation From rain-wetted forest in relation to canopy wetness, canopy cover, and net radiation. Water Resour. Res. 2001, 37, 3227–3236. [Google Scholar] [CrossRef]
- Levia, D.F.; Keim, R.F.; Carlyle-Moses, D.E.; Frost, E.E. Throughfall and Stemflow in Wooded Ecosystems. In Forest Hydrology and Biogeochemistry: Synthesis of Past Research and Future Directions; Levia, D.F., Carlyle-Moses, D., Tanaka, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 425–443. [Google Scholar]
- Levia, D.F.; Herwitz, S.R. Interspecific variation of bark water storage capacity of three deciduous tree species in relation to stemflow yield and solute flux to forest soils. CATENA 2005, 64, 117–137. [Google Scholar] [CrossRef]
- Murakami, S. Reduction in the ratio of stemflow to rainfall during heavy rain in two Japanese cedar stands and the influence on rainfall partitioning. J. Hydrol. 2024, 634, 131100. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Gay, T.E.; Lewis, E.S. Use of multiple correspondence analysis (MCA) to identify interactive meteorological conditions affecting relative throughfall. J. Hydrol. 2016, 533, 452–460. [Google Scholar] [CrossRef]
- Iida, S.; Levia, D.F.; Shimizu, A.; Shimizu, T.; Tamai, K.; Nobuhiro, T.; Kabeya, N.; Noguchi, S.; Sawano, S.; Araki, M. Intrastorm scale rainfall interception dynamics in a mature coniferous forest stand. J. Hydrol. 2017, 548, 770–783. [Google Scholar] [CrossRef]
- Nakai, T.; Lai, Y.-J. Micrometeorological estimation of wet canopy evaporation from a cloud forest in central Taiwan. Agric. For. Meteorol. 2024, 350, 109997. [Google Scholar] [CrossRef]
- Gao, G.Y.; Fu, B.J.; Lu, Y.H.; Liu, Y.; Wang, S.; Zhou, J. Coupling the modified SCS-CN and RUSLE models to simulate hydrological effects of restoring vegetation in the Loess Plateau of China. Hydrol. Earth Syst. Sci. 2012, 16, 2347–2364. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, L.; Gao, Y.; Chen, S.; Chen, W.; Hao, Z.; Jia, J.; Han, N. Geochemical isotopic composition in the Loess Plateau and corresponding source analyses: A case study of China’s Yangjuangou catchment. Sci. Total Environ. 2017, 581, 794–800. [Google Scholar] [CrossRef]
- Chen, W.L.; Li, Z.S.; Jiao, L.; Wang, C.; Gao, G.Y.; Fu, B.J. Response of Soil Moisture to Rainfall Event in Black Locust Plantations at Different Stages of Restoration in Hilly-gully Area of the Loess Plateau, China. Chin. Geogr. Sci. 2020, 30, 427–445. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Chen, L. Effects of non-native tree plantations on the diversity of understory plants and soil macroinvertebrates in the Loess Plateau of China. Plant Soil. 2020, 446, 357–368. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, B. Vitex negundo. In Flora of China; 65, Part 1: Angiospermae, Dicotyledoneae, Verbenaceae; Science Press: Beijing, China, 1982. [Google Scholar]
- Chen, J.; Wang, J.; Wang, Q.; Jia, C.; Kou, Y. Analysis of leaf area index and transmittance of isolated Vitex negundo canopy measured by plant canopy analyzer. Chin. J. Agrometeorol. 2014, 35, 373–379. [Google Scholar]
- Abtew, W.; Melesse, A. (Eds.) Evaporation and Evapotranspiration Estimation Methods. In Evaporation and Evapotranspiration: Measurements and Estimations; Springer: Dordrecht, The Netherlands, 2013; pp. 63–91. [Google Scholar]
- Herwitz, S.R.J.E.S.P. Landforms, Infiltration-excess caused by Stemflow in a cyclone-prone tropical rainforest. Earth Surf. Process. Landf. 1986, 11, 401–412. [Google Scholar] [CrossRef]
- Rodríguez-Iturbe, I. Hydrologic dynamics and ecosystem structure. Water Sci. Technol. 2003, 476, 18–24. [Google Scholar] [CrossRef]
- Gholami, H.; Mohammadifar, A.; Golzari, S.; Kaskaoutis, D.G.; Collins, A.L. Using the Boruta algorithm and deep learning models for mapping land susceptibility to atmospheric dust emissions in Iran. Aeolian Res. 2021, 50, 100682. [Google Scholar]
- Barton, K. Package MuMIn: Multi-Model Inference, Version 1.48.4. Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 22 June 2024).
- Fávero, L.P.; Belfiore, P.; de Freitas Souza, R. Chapter 7-Data visualization and multivariate graphs. In Data Science, Analytics and Machine Learning with R; Fávero, L.P., Belfiore, P., de Freitas Souza, R., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 89–125. [Google Scholar]
- Arnold, J.B. Package Ggthemes: Extra Themes, Scales and Geoms for ‘ggplot2’, Version 5.1.0. Available online: https://cran.r-project.org/web/packages/ggthemes/index.html (accessed on 10 February 2024).
- Groemping, U. Relative Importance for Linear Regression in R: The Package relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, D.; Yang, Z.; Seki, K.; Singh, M.; Wang, L. Changes in rainfall partitioning and its effect on soil water replenishment after the conversion of croplands into apple orchards on the Loess Plateau. Agric. Ecosyst. Environ. 2021, 312, 107342. [Google Scholar]
- Sadeghi, S.M.M.; Epstein, J.M.; Deljouei, A.; Gorora, F.J.; Cohen, M.J. Stand age controls canopy and soil rainfall partitioning in slash pine forests. For. Ecol. Manag. 2024, 572, 122307. [Google Scholar]
- Magliano, P.N.; Whitworth-Hulse, J.I.; Cid, F.D.; Leporati, J.L.; Van Stan, J.T.; Jobbágy, E.G. Global rainfall partitioning by dryland vegetation: Developing general empirical models. J. Hydrol. 2022, 607, 127540. [Google Scholar]
- Zhang, Y.; Wang, X.; Hu, R.; Pan, Y. Stemflow volume per unit rainfall as a good variable to determine the relationship between stemflow amount and morphological metrics of shrubs. J. Arid Environ. 2017, 141, 1–6. [Google Scholar]
- Magliano, P.N.; Whitworth-Hulse, J.I.; Baldi, G. Interception, throughfall and stemflow partition in drylands: Global synthesis and meta-analysis. J. Hydrol. 2019, 568, 638–645. [Google Scholar]
- Zhu, Z.; Shao, M.A.; Jia, X.; Zhao, C. Rainfall partitioning characteristics and simulation of typical shelter forest in Chinese Mu Us Sandy Land. Sci. Total Environ. 2024, 945, 174091. [Google Scholar]
- Rietkerk, M.; van de Koppel, J.; Kumar, L.; Herbert, H.T.; Prins, L. The ecology of scale. Ecol. Model. 2002, 149, 1–4. [Google Scholar] [CrossRef]
- Hatfield, J.; Dold, C. Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar]
- Gash, J.H.C.; Lloyd, C.R.; Lachaud, G. Estimating sparse forest rainfall interception with an analytical model. J. Hydrol. 1995, 170, 79–86. [Google Scholar]
- Herbst, M.; Rosier, P.T.W.; McNeil, D.D.; Harding, R.J.; Gowing, D.J. Seasonal variability of interception evaporation from the canopy of a mixed deciduous forest. Agric. For. Meteorol. 2008, 148, 1655–1667. [Google Scholar]
- Jeong, S.; Otsuki, K. Effects of tree mortality on the estimation of stemflow yield in a self-thinning coniferous plantation. Ecohydrology 2021, 14, e2327. [Google Scholar] [CrossRef]
- Levia, D.F.; Van Stan, J.T.; Mage, S.M.; Kelley-Hauske, P.W. Temporal variability of stemflow volume in a beech-yellow poplar forest in relation to tree species and size. J. Hydrol. 2010, 380, 112–120. [Google Scholar] [CrossRef]
- Berland, A.; Shiflett, S.A.; Shuster, W.D.; Garmestani, A.S.; Goddard, H.C.; Herrmann, D.L.; Hopton, M.E. The role of trees in urban stormwater management. Landsc. Urban. Plan. 2017, 162, 167–177. [Google Scholar]
- Chen, S.; Cao, R.; Yoshitake, S.; Ohtsuka, T. Stemflow hydrology and DOM flux in relation to tree size and rainfall event characteristics. Agric. For. Meteorol. 2019, 279, 107753. [Google Scholar]
- Wu, M.-D.; Jiang, Z.Y.; Yang, X.; Yeerbolati, B.; Huang, L.; Zhong, Y.Y.; Zhang, H.-W.; Li, X.Y. Global quantitative synthesis on the patterns and drivers of funneling ratio and enrichment ratio for stemflow. Catena 2024, 244, 108253. [Google Scholar] [CrossRef]
- Sun, X.; Onda, Y.; Kato, H.; Gomi, T.; Komatsu, H. Effect of strip thinning on rainfall interception in a Japanese cypress plantation. J. Hydrol. 2015, 525, 607–618. [Google Scholar]
- Alejandro, G.O.; Alexia, S.; Slobodan, B.M. A novel framework to study the effect of tree architectural traits on stemflow yield and its consequences for soil-water dynamics. J. Hydrol. 2020, 582, 124448. [Google Scholar]
- Herwitz, S.; Slye, R. Three-dimensional modeling of canopy tree interception of wind-driven rainfall. J. Hydrol. 1995, 168, 205–226. [Google Scholar] [CrossRef]
- Fan, J.; Oestergaard, K.; Guyot, A.; Jensen, D.; Lockington, D. Spatial variability of throughfall and stemflow in an exotic pine plantation of subtropical coastal Australia. Hydrol. Process 2015, 29, 793–804. [Google Scholar] [CrossRef]
- Zabret, K.; Šraj, M. How Characteristics of a Rainfall Event and the Meteorological Conditions Determine the Development of Stemflow: A Case Study of a Birch Tree. Front. For. Glob. Change 2021, 4, 663100. [Google Scholar] [CrossRef]
- Zhang, H.X.; Fu, C.S.; Liao, A.M.; Zhang, C.; Liu, J.F.; Wang, N.; He, B. Exploring the stemflow dynamics and driving factors at both inter- and intra-event scales in a typical subtropical deciduous forest. Hydrol. Process. 2021, 3, e14091. [Google Scholar] [CrossRef]
- Yang, X.L.; Shao, M.A.; Wei, X.R. Stemflow production differ significantly among tree and shrub species on the Chinese Loess Plateau. J. Hydrol. 2019, 568, 427–436. [Google Scholar] [CrossRef]
- Niu, X.T.; Fan, J.; Du, M.G.; Dai, Z.J.; Luo, R.H.; Yuan, H.Y.; Zhang, S.G. Changes of Rainfall Partitioning and Canopy Interception Modeling after Progressive Thinning in Two Shrub Plantations on the Chinese Loess Plateau. J. Hydrol. 2023, 619, 129299. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Self-Organized Patterns | Total Canopy Areas (m2) | Average Canopy Height (m) | BD Categories (mm) | Branch Amount | Average BD (mm) | Average BL (cm) | Average BA (°) | Average TLA (cm2) |
|---|---|---|---|---|---|---|---|---|
| Clumped | 31.5 | 2.2 ± 0.2 | <10 | 120 | 6.3 ± 2.0 | 131.9 ± 49.0 | 52.6 ± 18.0 | 846.7 ± 522.5 |
| 10–15 | 69 | 12.1 ± 1.4 | 202.9 ± 72.6 | 52.6 ± 16.3 | 3086.6 ± 794.7 | |||
| 15–20 | 28 | 17.7 ± 1.3 | 225.6 ± 42.3 | 52.5 ± 12.2 | 6867.0 ± 1043.6 | |||
| >20 | 7 | 21.9 ± 1.0 | 245.2 ± 69.9 | 51.6 ± 13.1 | 10,654.8 ± 988.5 | |||
| Scattered | 33.8 | 2.6 ± 0.5 | <10 | 13 | 7.4 ± 1.7 | 158.2 ± 49.3 | 61.7 ± 15.8 | 1130.8 ± 460.8 |
| 10–15 | 13 | 13.2 ± 1.4 | 237.6 ± 30.2 | 50.6 ± 12.4 | 3665.8 ± 789.2 | |||
| 15–20 | 12 | 17.2 ± 1.7 | 275.2 ± 44.6 | 58.4 ± 13.3 | 6444.7 ± 1378.2 | |||
| >20 | 21 | 22.5 ± 2.5 | 266.8 ± 69.0 | 53.3 ± 20.1 | 11,425.2 ± 2957.0 |
| Rainfall Partitioning Components | Indicators | Clumped V. negundo | Scattered V. negundo |
|---|---|---|---|
| Net precipitation | Depth (NPd, mm) | 25.7 ± 33.3 a | 25.4 ± 32.6 a |
| Percentage (NP%, %) | 83.5 ± 11.2 a | 84.2 ± 8.2 a | |
| Stemflow | Depth (SFd, mm) | 2.7 ± 3.6 a | 1.7 ± 2.4 b |
| Percentage (SF%, %) | 8.6 ± 2.0 a | 4.7 ± 1.2 b | |
| Average intensity (SFI, mm·h–1) | 651.0 ± 962.1 a | 346.9 ± 649.1 b | |
| 10 min maximum intensity (SFI10, mm·h–1) | 2386.6 ± 2990.3 a | 2623.9 ± 3135.8 a | |
| Funneling ratio (FR, unitless) | 154.4 ± 51.7 a | 134.8 ± 46.1 b | |
| Productivity (SFP, mm·kg–1) | 3.3 ± 4.8 a | 2.2 ± 2.1 b | |
| Time lag for generation (TLGSF, min) | 27.2 ± 29.8 a | 23.8 ± 26.1 a | |
| Time lag for maximization (TLMSF, min) | 119.3 ± 221.9 a | 92.3 ± 161.0 a | |
| Time lag for ending (TLESF, min) | 161.0 ± 137.0 a | 137.0 ± 112.6 a | |
| Duration (SFD, h) | 11.9 ± 8.0 a | 11.6 ± 8.0 a | |
| Throughfall | Depth (TFd, mm) | 23.0 ± 29.3 a | 23.8 ± 29.8 a |
| Percentage (TF%, %) | 74.9 ± 10.2 a | 79.0 ± 7.9 b | |
| Average intensity (TFI, mm·h–1) | 5.0 ± 8.0 a | 5.8 ± 10.5 a | |
| 10 min maximum intensity (TFI10, mm·h–1) | 20.3 ± 19.2 a | 16.4 ± 16.3 b | |
| Productivity (TFP, mm·kg–1) | 6.1 ± 7.6 a | 9.4 ± 12.2 b | |
| Time lag for generation (TLGTF, min) | 17.8 ± 18.0 a | 17.6 ± 23.7 a | |
| Time lag for maximization (TLMTF, min) | 61.5 ± 101.2 a | 138.4 ± 142.5 b | |
| Time lag for ending (TLETF, min) | 102.2 ± 218.1 a | 216.1 ± 270.7 b | |
| Duration (TFD, h) | 11.1 ± 14.7 a | 14.2 ± 9.4 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Yuan, C.; Zhang, Y.; Hu, Y.; Guo, L.; Jiang, Z.; Wang, S.; Wang, C. Rainfall Partitioning Dynamics in Xerophytic Shrubs: Interplays Between Self-Organization and Meteorological Drivers. Forests 2025, 16, 605. https://doi.org/10.3390/f16040605
Gao Y, Yuan C, Zhang Y, Hu Y, Guo L, Jiang Z, Wang S, Wang C. Rainfall Partitioning Dynamics in Xerophytic Shrubs: Interplays Between Self-Organization and Meteorological Drivers. Forests. 2025; 16(4):605. https://doi.org/10.3390/f16040605
Chicago/Turabian StyleGao, Yinghao, Chuan Yuan, Yafeng Zhang, Yanting Hu, Li Guo, Zhiyun Jiang, Sheng Wang, and Cong Wang. 2025. "Rainfall Partitioning Dynamics in Xerophytic Shrubs: Interplays Between Self-Organization and Meteorological Drivers" Forests 16, no. 4: 605. https://doi.org/10.3390/f16040605
APA StyleGao, Y., Yuan, C., Zhang, Y., Hu, Y., Guo, L., Jiang, Z., Wang, S., & Wang, C. (2025). Rainfall Partitioning Dynamics in Xerophytic Shrubs: Interplays Between Self-Organization and Meteorological Drivers. Forests, 16(4), 605. https://doi.org/10.3390/f16040605