

Species Differentiation of Two Endemic Montane Oaks in China: Population Genetics, Ecological Niche and Leaf Morphology Analyses


Abstract
1. Introduction
2. Materials and Methods
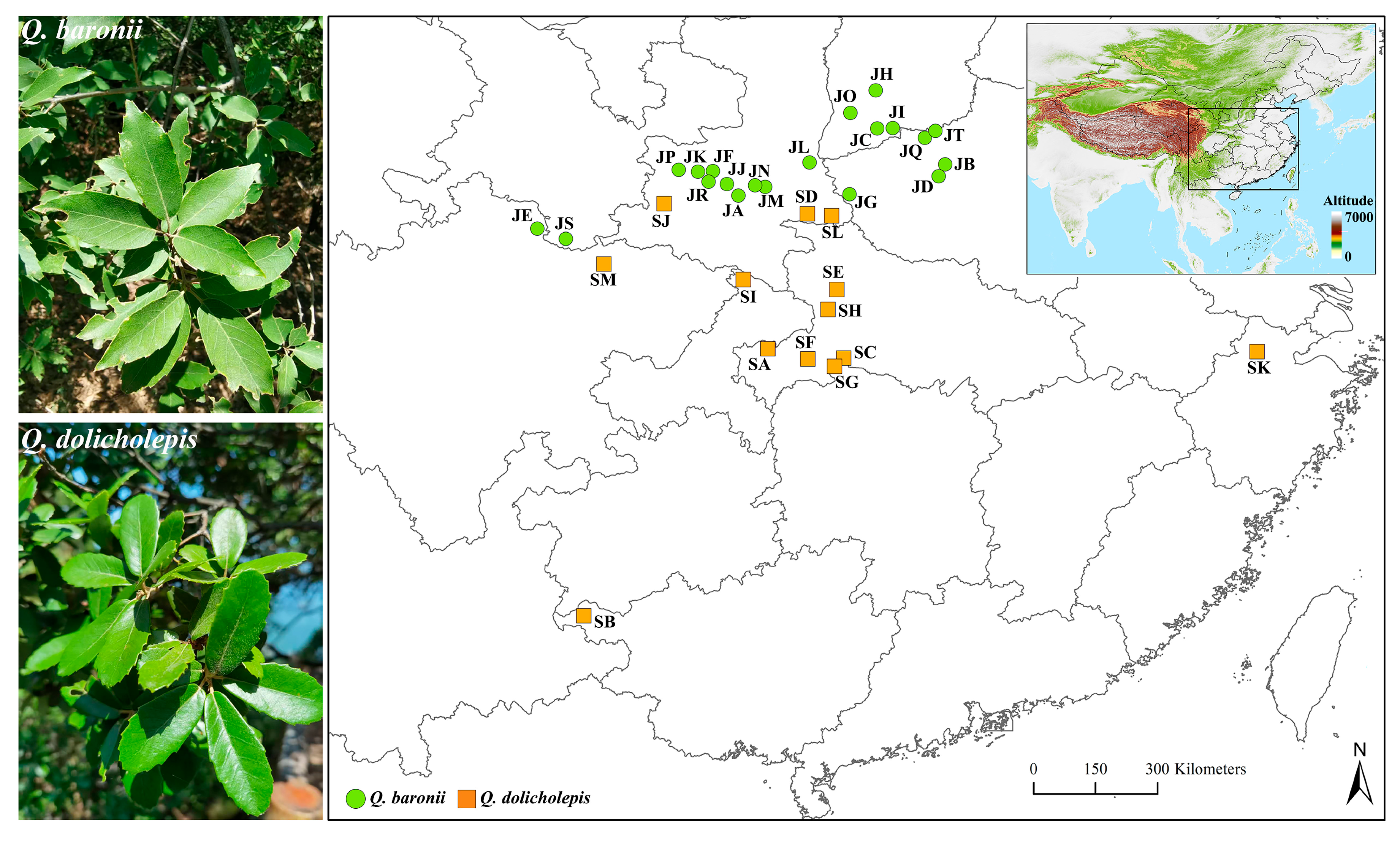
2.1. Sample Information and Microsatellite Genotyping
2.2. Genetic Variation and Population Structure
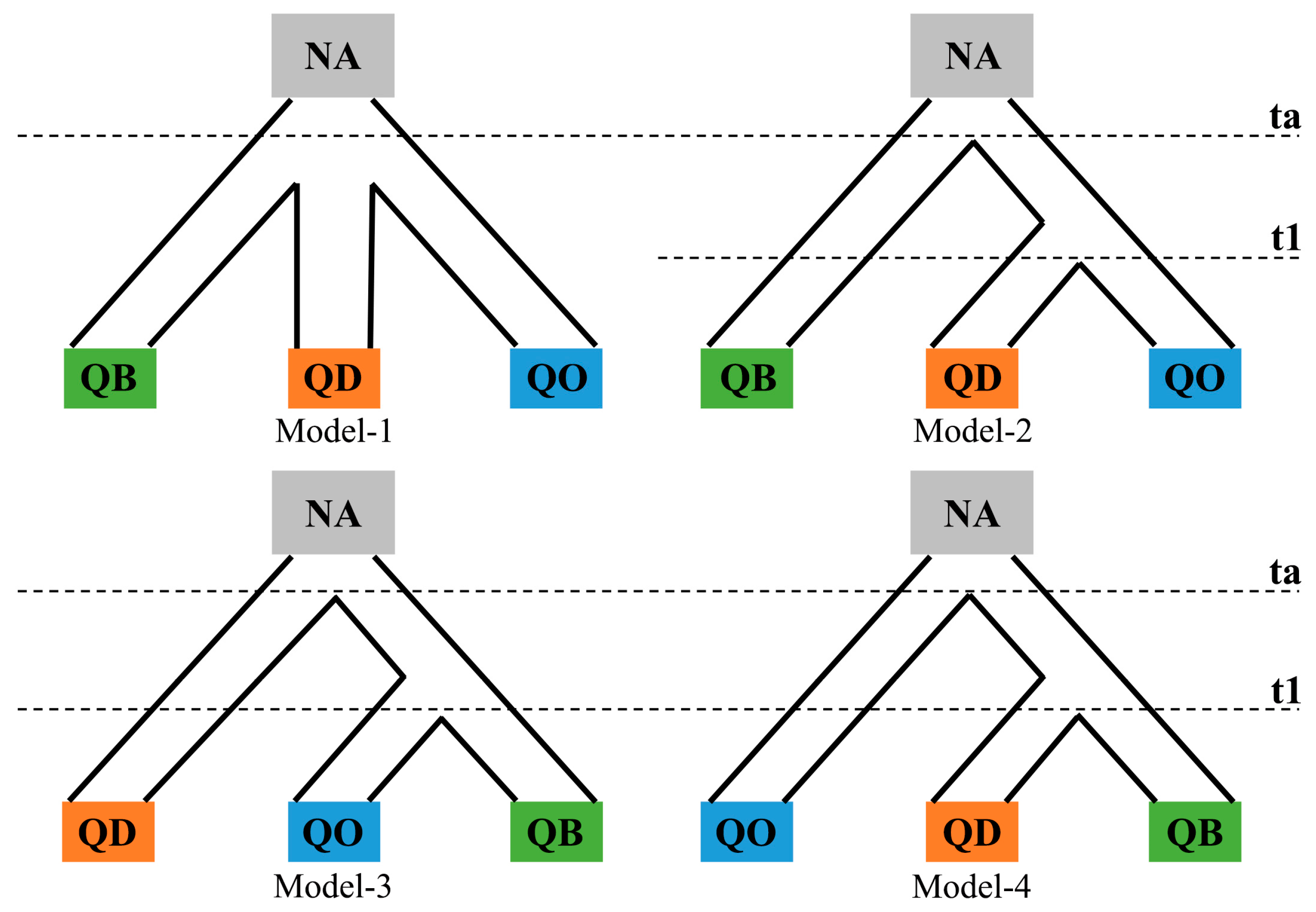
2.3. Demographic Models on Species Divergence
2.4. Ecological Differentiation Between Q. baronii and Q. dolicholepis
2.5. Morphological Analysis of Leaf Traits
3. Results
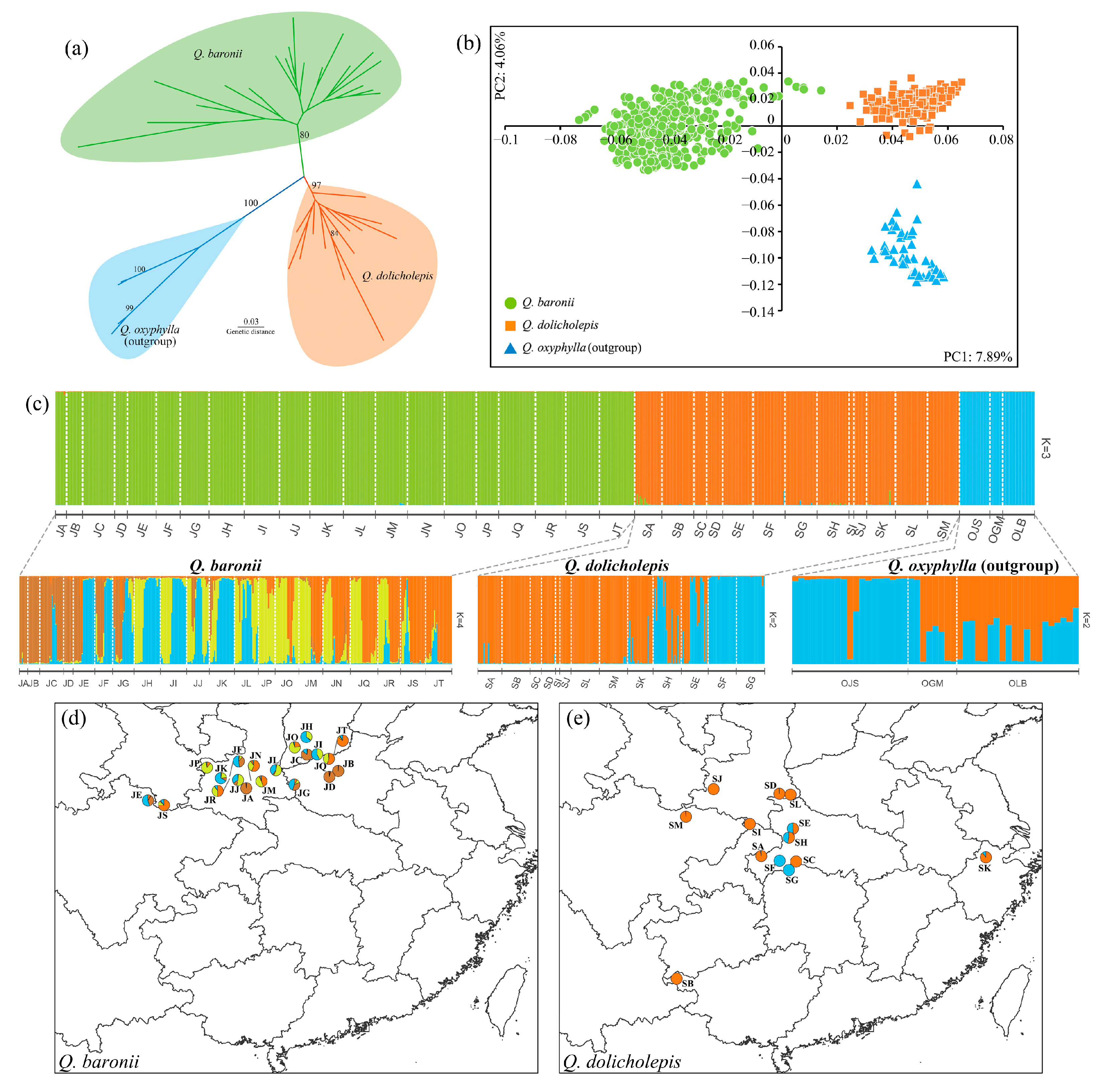
3.1. Genetic Diversity and Genetic Differentiation
3.2. Population Structure and Species Relationships
3.3. Models of Species Divergence, Species Demographic History and Gene Flow
3.4. Ecological Differentiation and Association Analysis
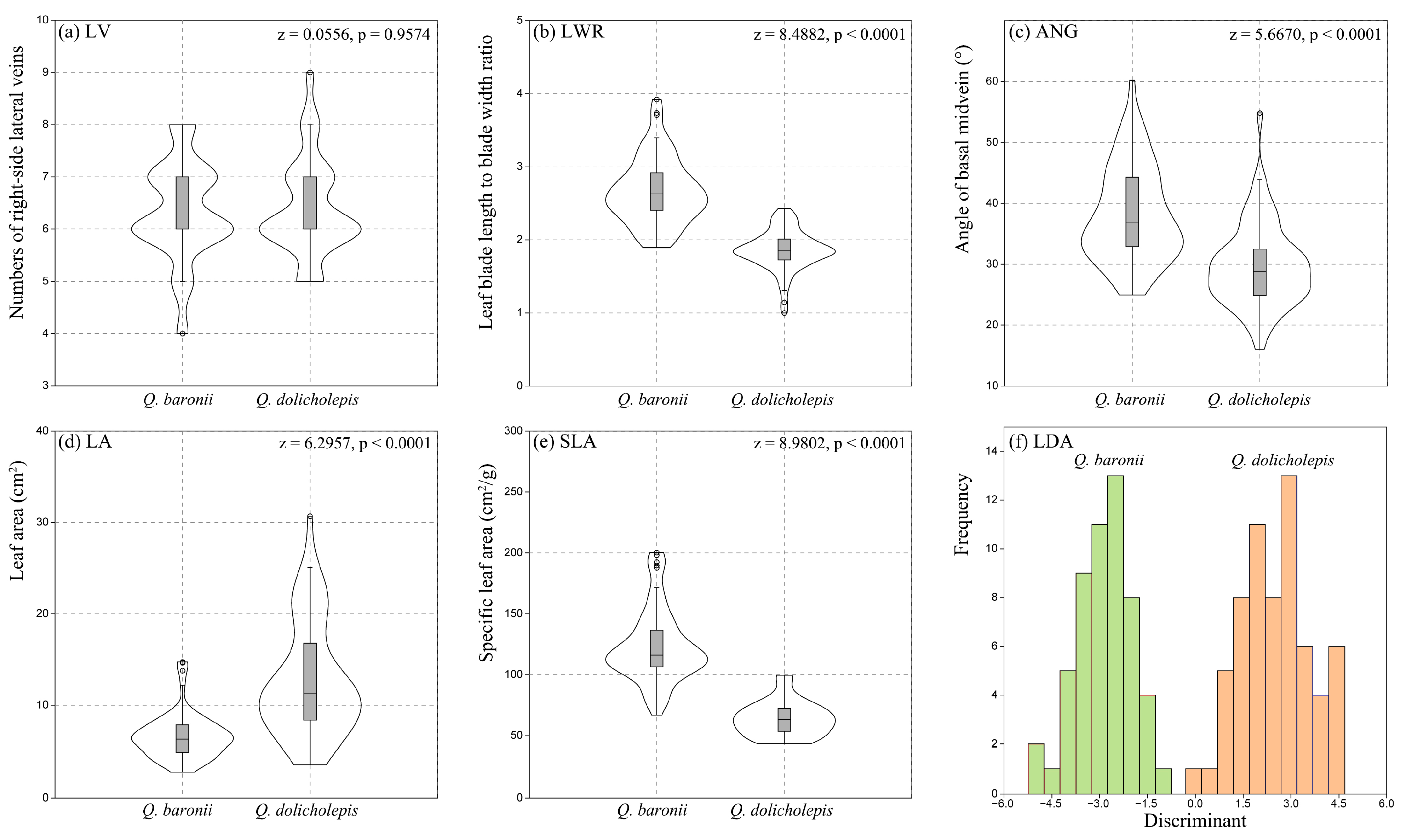
3.5. Variations of Leaf Morphology
4. Discussion
4.1. Species Differentiation Between Q. baronii and Q. dolicholepis
4.2. Evolutionary History and Potential Drivers on Species Divergence
4.3. Genetic Diversity and Implications on Species Conservation of the Two Endemic Oaks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nevado, B.; Wong, E.L.Y.; Osborne, O.G.; Filatov, D.A. Adaptive evolution is common in rapid evolutionary radiations. Curr. Biol. 2019, 29, 3081–3086. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.; Roddy, A.B.; DiVittorio, C.; Sanchez-Amaya, A.; Henriquez, C.L.; Brodersen, C.R.; Fehlberg, S.; Zapata, F. Diversification, disparification and hybridization in the desert shrubs Encelia. New Phytol. 2021, 230, 1228–1241. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.M.; Yang, M.Q.; Li, C.L.; Huang, S.X.; Jin, W.T.; Shen, T.T.; Wang, F.; Li, X.H.; Yoichi, W.; Zhang, L.H.; et al. Spatiotemporal evolution of the global species diversity of Rhododendron. Mol. Biol. Evol. 2022, 39, msab314. [Google Scholar] [CrossRef] [PubMed]
- Du, F.K.; Hou, M.; Wang, W.T.; Mao, K.S.; Hampe, A. Phylogeography of Quercus aquifolioides provides novel insights into the Neogene history of a major global hotspot of plant diversity in south-west China. J. Biogeogr. 2017, 44, 294–307. [Google Scholar] [CrossRef]
- Smycka, J.; Roquet, C.; Boleda, M.; Alberti, A.; Boyer, F.; Douzet, R.; Perrier, C.; Rome, M.; Valay, J.-G.; Denoeud, F. Tempo and drivers of plant diversification in the European mountain system. Nat. Commun. 2022, 13, 2750. [Google Scholar] [CrossRef]
- Wang, X.Y.; Liang, D.; Wang, X.M.; Tang, M.K.; Liu, Y.; Liu, S.Y.; Zhang, P. Phylogenomics reveals the evolution, biogeography, and diversification history of voles in the Hengduan Mountains. Commun. Biol. 2022, 5, 1124. [Google Scholar] [CrossRef]
- Hughes, C.E.; Atchison, G.W. The ubiquity of alpine plant radiations: From the Andes to the Hengduan Mountains. New Phytol. 2015, 207, 275–282. [Google Scholar] [CrossRef]
- Bedoya, A.M.; Leache, A.D.; Olmstead, R.G. Andean uplift, drainage basin formation, and the evolution of plants living in fast-flowing aquatic ecosystems in northern South America. New Phytol. 2021, 232, 2175–2190. [Google Scholar] [CrossRef]
- Wu, S.D.; Wang, Y.; Wang, Z.F.; Shrestha, N.; Liu, J.Q. Species divergence with gene flow and hybrid speciation on the Qinghai-Tibet Plateau. New Phytol. 2022, 234, 392–404. [Google Scholar] [CrossRef]
- Kremer, A.; Hipp, A.L. Oaks: An evolutionary success story. New Phytol. 2020, 226, 987–1011. [Google Scholar] [CrossRef]
- Plomion, C.; Aury, J.M.; Amselem, J.; Alaeitabar, T.; Barbe, V.; Belser, C.; Berges, H.; Bodénès, C.; Boudet, N.; Boury, C.; et al. Decoding the oak genome: Public release of sequence data, assembly, annotation and publication strategies. Mol. Ecol. Resour. 2016, 16, 254–265. [Google Scholar] [CrossRef]
- Plomion, C.; Aury, J.M.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Le Provost, G.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 4, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Gugger, P.F.; Ikegami, M.; Sork, V.L. Influence of late Quaternary climate change on present patterns of genetic variation in valley oak, Quercus lobata Nee. Mol. Ecol. 2013, 22, 3598–3612. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.A.; Hipp, A.L.; Gonzalez-Rodriguez, A.; Cavender-Bares, J. Historical introgression among the American live oaks and the comparative nature of tests for introgression. Evolution 2015, 6, 2587–2601. [Google Scholar] [CrossRef] [PubMed]
- McVay, J.D.; Hipp, A.L.; Manos, P.S. A genetic legacy of introgression confounds phylogeny and biogeography in oaks. Proc. R. Soc. Lond. B. Biol. Sci. 2017, 284, 20170300. [Google Scholar] [CrossRef]
- Kim, B.Y.; Wei, X.; Fitz-Gibbon, S.; Lohmueller, K.E.; Ortego, J.; Gugger, P.F.; Sork, V.L. RADseq data reveal ancient, but not pervasive, introgression between Californian tree and scrub oak species (Quercus sect. Quercus: Fagaceae). Mol. Ecol. 2018, 27, 4556–4571. [Google Scholar] [CrossRef]
- Crowl, A.A.; Manos, P.S.; McVay, J.D.; Lemmon, A.R.; Lemmon, E.M.; Hipp, A.L. Uncovering the genomic signature of ancient introgression between white oak lineages (Quercus). New Phytol. 2020, 226, 1158–1170. [Google Scholar] [CrossRef]
- Hipp, A.L.; Manos, P.S.; Hahn, M.; Avishai, M.; Bodenes, C.; Cavender-Bares, J.; Crowl, A.A.; Deng, M.; Denk, T.; Fitz-Gibbon, S.; et al. Genomic landscape of the global oak phylogeny. New Phytol. 2020, 226, 1198–1212. [Google Scholar] [CrossRef]
- Yang, J.; Guo, Y.F.; Chen, X.D.; Zhang, X.; Ju, M.M.; Bai, G.Q.; Liu, Z.L.; Zhao, G.F. Framework phylogeny, evolution and complex diversification of Chinese oaks. Plants 2020, 9, 1024. [Google Scholar] [CrossRef]
- Jiang, X.L.; Hipp, A.L.; Deng, M.; Su, T.; Zhou, Z.K.; Yan, M.X. East Asian origins of European holly oaks (Quercus section Ilex Loudon) via the Tibet-Himalaya. J. Biogeogr. 2019, 46, 2203–2214. [Google Scholar] [CrossRef]
- Dow, B.D.; Ashley, M.V.; Howe, H.F. Characterization of highly variable (GA/CT)n microsatellites in the bur oak, Quercus macrocarpa. Theor. Appl. Genet. 1995, 91, 137–141. [Google Scholar] [CrossRef]
- Steinkellner, H.; Fluch, S.; Turetschek, E.; Lexer, C.; Streiff, R.; Kremer, A.; Burg, K.; Glossl, J. Identification and characterization of (GA/CT)n-microsatellite loci from Quercus petraea. Plant Mol. Biol. 1997, 33, 1093–1096. [Google Scholar] [CrossRef]
- Kampfer, S.; Lexer, C.; Glossl, J.; Steinkellner, H. Characterization of (GA)n microsatellite loci from Quercus robur. Hereditas 2004, 129, 183–186. [Google Scholar] [CrossRef]
- Sullivan, A.R.; Lind, J.F.; McCleary, T.S.; Romero-Severson, J.; Gailing, O. Development and characterization of genomic and gene-based microsatellite markers in north American red oak species. Plant Mol. Biol. Report. 2012, 31, 231–239. [Google Scholar] [CrossRef]
- Chen, X.D.; Zhang, X.; Zhang, H.; Zhou, T.; Zhao, Y.M.; Yang, J.; Zhao, G.F. Genetic differentiation and demographic history of three Cerris oak species in China based on nuclear microsatellite makers. Forests 2021, 12, 1164. [Google Scholar] [CrossRef]
- Holland, M.M.; Parson, W. GeneMarker HID: A reliable software tool for the analysis of forensic STR data. J. Forensic Sci. 2011, 56, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Boyle, T.J.B. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot. 1997, 129, 157. [Google Scholar]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT (Version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Takezaki, N.; Nei, M.; Tamura, K. POPTREE2: Software for constructing population trees from allele frequency data and computing other population statistics with Windows interface. Mol. Biol. Evol. 2010, 27, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Francis, R.M. POPHELPER: An R package and web app to analyse and visualize population structure. Mol. Ecol. Resour. 2017, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F.; Beeravolu, C.R.; Leblois, R. Likelihood computation and inference of demographic and mutational parameters from population genetic data under coalescent approximations. J. Soc. Française Stat. 2018, 159, 142–166. [Google Scholar]
- Cornuet, J.M.; Pudlo, P.; Veyssier, J.; Dehne-Garcia, A.; Gautier, M.; Leblois, R.; Marin, J.M.; Estoup, A. DIYABC v2.0: A software to make approximate Bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics 2014, 30, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Rosenberg, M.S.; Anderson, C.D. PASSaGE: Pattern analysis, spatial statistics and geographic exegesis. version 2. Methods Ecol. Evol. 2011, 2, 229–232. [Google Scholar] [CrossRef]
- Yang, J.; Di, X.Y.; Meng, X.; Feng, L.; Liu, Z.L.; Zhao, G.F. Phylogeography and evolution of two closely related oak species (Quercus) from north and northeast China. Tree Genet. Genomes 2016, 12, 89. [Google Scholar] [CrossRef]
- Garcia-Nogales, A.; Linares, J.C.; Laureano, R.G.; Seco, J.I.; Merino, J. Range-wide variation in life-history phenotypes: Spatiotemporal plasticity across the latitudinal gradient of the evergreen oak Quercus ilex. J. Biogeogr. 2016, 43, 2366–2379. [Google Scholar] [CrossRef]
- Leroy, T.; Louvet, J.M.; Lalanne, C.; Provost, G.L.; Labadie, K.; Aury, J.-M.; Delzon, S.; Plomion, C.; Kremer, A. Adaptive introgression as a driver of local adaptation to climate in European white oaks. New Phytol. 2020, 226, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.R.; Zhu, Y.X.; Liu, Y.; Feng, Y.; Lu, R.S.; Li, Y.; Li, P.; Kremer, A.; Lascoux, M.; Chen, J. Genome-wide analyses of introgression between two sympatric Asian oak species. Nat. Ecol. Evol. 2022, 6, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Leroy, T.; Roux, C.; Villate, L.; Bodenes, C.; Romiguier, J.; Paiva, J.A.; Dossat, C.; Aury, J.M.; Plomion, C.; Kremer, A. Extensive recent secondary contacts between four European white oak species. New Phytol. 2017, 214, 865–878. [Google Scholar] [CrossRef]
- Feng, L.; Zheng, Q.J.; Qian, Z.Q.; Yang, J.; Zhang, Y.P.; Li, Z.H.; Zhao, G.F. Genetic structure and evolutionary history of three alpine sclerophyllous oaks in east Himalaya-Hengduan Mountains and adjacent regions. Front. Plant. Sci. 2016, 7, 1688. [Google Scholar] [CrossRef]
- Meng, H.H.; Su, T.; Gao, X.Y.; Li, J.; Jiang, X.L.; Sun, H.; Zhou, Z.K. Warm-cold colonization: Response of oaks to uplift of the Himalaya-Hengduan Mountains. Mol. Ecol. 2017, 26, 3276–3294. [Google Scholar] [CrossRef]
- Ju, M.M.; Yang, J.; Yue, M.; Zhao, G.F. Speciation patterns of related species under the hybrid zone: A case study of three sclerophyllous oaks in the east Himalaya-Hengduan Mountains. Mol. Ecol. 2023, 32, 4610–4626. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Liao, W.J.; Petit, R.J.; Zhang, D.Y. Geographic variation in the structure of oak hybrid zones provides insights into the dynamics of speciation. Mol. Ecol. 2011, 20, 4995–5011. [Google Scholar] [CrossRef]
- An, M.; Deng, M.; Zheng, S.S.; Jiang, X.L.; Song, Y.G. Introgression threatens the genetic diversity of Quercus austrocochinchinensis (Fagaceae), an endangered oak: A case inferred by molecular markers. Front. Plant. Sci. 2017, 8, 229. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Gonzalez-Rodriguez, A.; Pahlich, A.; Koehler, K.; Deacon, N. Phylogeography and climatic niche evolution in live oaks (Quercus series Virentes) from the tropics to the temperate zone. J. Biogeogr. 2011, 38, 962–981. [Google Scholar] [CrossRef]
- Zhou, Z.K. Origin, phylogeny and dispersal of Quercus from China. Acta Bot. Yunnanica 1992, 14, 227–236. [Google Scholar]
- Liu, J.Q.; Sun, Y.S.; Ge, X.J.; Gao, L.M.; Qiu, Y.X. Phylogeographic studies of plants in China: Advances in the past and directions in the future. J. Syst. Evol. 2012, 50, 267–275. [Google Scholar] [CrossRef]
- Yuan, J.H.; Cheng, F.Y.; Zhou, S.L. Genetic structure of the Tree peony (Paeonia rockii) and the Qinling Mountains as a geographic barrier driving the fragmentation of a large population. PLoS ONE 2012, 7, e34955. [Google Scholar] [CrossRef] [PubMed]
- Ortego, J.; Gugger, P.F.; Sork, V.L. Impacts of human-induced environmental disturbances on hybridization between two ecologically differentiated Californian oak species. New Phytol. 2017, 213, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Truffaut, L.; Chancerel, E.; Ducousso, A.; Dupouey, J.L.; Badeau, V.; Ehrenmann, F.; Kremer, A. Fine-scale species distribution changes in a mixed oak stand over two successive generations. New Phytol. 2017, 215, 126–139. [Google Scholar] [CrossRef]
- Saleh, D.; Chen, J.; Leple, J.C.; Leroy, T.; Truffaut, L.; Dencausse, B.; Lalanne, C.; Labadie, K.; Lesur, I.; Bert, D.; et al. Genome-wide evolutionary response of European oaks during the Anthropocene. Evol. Lett. 2022, 6, 4–20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample Information | Genetic Diversity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Population | Longitude (E) | Latitude (N) | Samples | Na | Ne | Ho | He | Ar | F | |
| Q. baronii | JA | 108.53 | 33.80 | 7 | 2.8000 | 2.2498 | 0.0857 | 0.4857 | 2.8000 | 0.8266 |
| JB | 113.03 | 34.48 | 10 | 2.8000 | 2.5366 | 0.3300 | 0.5775 | 2.7894 | 0.4569 | |
| JC | 111.55 | 35.26 | 20 | 4.0000 | 3.2049 | 0.3450 | 0.6433 | 3.6221 | 0.4195 | |
| JD | 112.89 | 34.22 | 8 | 2.6000 | 2.2819 | 0.2875 | 0.5328 | 2.5742 | 0.4647 | |
| JE | 104.15 | 33.08 | 18 | 3.7000 | 2.9046 | 0.2222 | 0.6133 | 3.4407 | 0.6581 | |
| JF | 107.89 | 34.23 | 15 | 3.8000 | 2.8833 | 0.3333 | 0.6302 | 3.4571 | 0.4731 | |
| JG | 110.95 | 33.83 | 18 | 4.0000 | 3.1851 | 0.3389 | 0.6650 | 3.6451 | 0.4811 | |
| JH | 111.52 | 36.09 | 22 | 3.9000 | 3.2214 | 0.4500 | 0.6687 | 3.6154 | 0.3023 | |
| JI | 111.89 | 35.27 | 22 | 4.3000 | 3.3087 | 0.4318 | 0.6711 | 3.8975 | 0.3350 | |
| JJ | 108.28 | 34.05 | 19 | 3.8000 | 2.8488 | 0.4158 | 0.6324 | 3.3966 | 0.3502 | |
| JK | 107.75 | 34.28 | 21 | 4.7000 | 3.3107 | 0.4286 | 0.6762 | 4.0139 | 0.3563 | |
| JL | 110.08 | 34.52 | 20 | 4.5000 | 3.5972 | 0.3500 | 0.6844 | 4.0612 | 0.4740 | |
| JM | 108.97 | 33.99 | 20 | 4.6000 | 3.7120 | 0.3650 | 0.7116 | 4.1735 | 0.4819 | |
| JN | 108.89 | 34.02 | 23 | 4.3000 | 3.2665 | 0.3739 | 0.6630 | 3.8094 | 0.4244 | |
| JO | 110.97 | 35.60 | 20 | 4.8000 | 3.8100 | 0.3400 | 0.6648 | 4.1449 | 0.4736 | |
| JP | 107.23 | 34.36 | 14 | 3.4000 | 2.8825 | 0.2214 | 0.6355 | 3.3094 | 0.6560 | |
| JQ | 112.59 | 35.06 | 23 | 4.1000 | 3.1536 | 0.3304 | 0.6382 | 3.6204 | 0.4872 | |
| JR | 107.88 | 34.10 | 19 | 3.7000 | 3.0285 | 0.2842 | 0.6440 | 3.3737 | 0.5743 | |
| JS | 104.77 | 32.86 | 21 | 3.9000 | 3.0389 | 0.3429 | 0.6608 | 3.4964 | 0.4908 | |
| JT | 112.82 | 35.21 | 22 | 3.7000 | 3.0814 | 0.4000 | 0.6546 | 3.5397 | 0.3915 | |
| Q. dolicholepis | SA | 109.17 | 30.46 | 17 | 8.2000 | 6.1017 | 0.1294 | 0.8066 | 4.1213 | 0.8503 |
| SB | 105.16 | 24.65 | 20 | 7.8000 | 5.2612 | 0.0450 | 0.7923 | 3.9311 | 0.9438 | |
| SC | 110.68 | 30.20 | 8 | 5.1000 | 4.1974 | 0.0375 | 0.7273 | 3.6758 | 0.9398 | |
| SD | 110.03 | 33.40 | 10 | 6.3000 | 4.7984 | 0.1500 | 0.7660 | 3.9306 | 0.7801 | |
| SE | 110.67 | 31.75 | 19 | 11.0000 | 8.0059 | 0.2000 | 0.8519 | 4.5399 | 0.7429 | |
| SF | 110.04 | 30.24 | 20 | 9.9000 | 6.0517 | 0.1600 | 0.8145 | 4.2018 | 0.7986 | |
| SG | 110.62 | 30.08 | 20 | 8.6000 | 5.7415 | 0.0650 | 0.7893 | 3.9813 | 0.9257 | |
| SH | 110.48 | 31.32 | 20 | 9.3000 | 6.9327 | 0.0450 | 0.8395 | 4.3558 | 0.9480 | |
| SI | 108.63 | 31.97 | 3 | 2.6000 | 2.5400 | 0.0333 | 0.5167 | 2.6000 | 0.9423 | |
| SJ | 106.91 | 33.62 | 8 | 5.0000 | 3.8381 | 0.1125 | 0.7211 | 3.6012 | 0.8485 | |
| SK | 119.82 | 30.40 | 18 | 9.1000 | 6.8007 | 0.1611 | 0.8228 | 4.2382 | 0.8104 | |
| SL | 110.56 | 33.36 | 20 | 9.1000 | 6.8931 | 0.1200 | 0.8378 | 4.3056 | 0.8587 | |
| SM | 105.60 | 32.31 | 20 | 8.0000 | 5.8406 | 0.0800 | 0.8051 | 4.0901 | 0.9048 | |
| Q. baronii | Q. dolicholepis | |||
|---|---|---|---|---|
| Mean | SE | Mean | SE | |
| Na | 8.7000 | 0.9434 | 33.2000 | 2.7439 |
| Ne | 5.0625 | 0.4542 | 15.2271 | 1.6077 |
| Np | 4.4000 | 1.0975 | 28.3000 | 3.2628 |
| Ho | 0.3489 | 0.0347 | 0.1084 | 0.0428 |
| He | 0.7888 | 0.0182 | 0.9273 | 0.0080 |
| Dest | 0.4066 | 0.0384 | 0.5783 | 0.0550 |
| HS | 0.6377 | 0.0164 | 0.7762 | 0.0135 |
| HT | 0.7952 | 0.0169 | 0.9231 | 0.0076 |
| F | 0.5527 | 0.0480 | 0.8826 | 0.0462 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Variation (%) | RST | |
|---|---|---|---|---|---|---|
| Q. baronii-Q. dolicholepis | Among species | 1 | 6757820.992 | 12945.24002 Va | 94.27 | 0.9880 *** |
| Among populations | 31 | 662266.789 | 622.43719 Vb | 4.53 | ||
| Within populations | 1097 | 181012.571 | 165.00690 Vc | 1.20 | ||
| Q. baronii | Among populations | 19 | 39386.212 | 55.71205 Va | 46.78 | 0.4678 *** |
| Within populations | 704 | 44613.705 | 63.37174 Vb | 53.22 | ||
| Q. dolicholepis | Among populations | 12 | 622880.577 | 1670.29220 Va | 82.80 | 0.8279 *** |
| Within populations | 393 | 136398.866 | 347.07091 Vb | 17.20 |
| Likelihood Ratio Tests | ||||
|---|---|---|---|---|
| Demographic Models | Log-Likelihood | Divergence | Migration | Divergence-with-Migration |
| Divergence | −1939.2 | - | 343.4 *** | 458.8 *** |
| Migration | −2110.4 | - | - | 116.4 *** |
| Isolation-Migration | −2168.6 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Z.-M.; Zhang, L.-L.; Huang, Y.-J.; Chen, X.-D.; Yang, J. Species Differentiation of Two Endemic Montane Oaks in China: Population Genetics, Ecological Niche and Leaf Morphology Analyses. Forests 2025, 16, 549. https://doi.org/10.3390/f16030549
Chang Z-M, Zhang L-L, Huang Y-J, Chen X-D, Yang J. Species Differentiation of Two Endemic Montane Oaks in China: Population Genetics, Ecological Niche and Leaf Morphology Analyses. Forests. 2025; 16(3):549. https://doi.org/10.3390/f16030549
Chicago/Turabian StyleChang, Zhi-Mei, Lu-Lu Zhang, Yun-Ju Huang, Xiao-Dan Chen, and Jia Yang. 2025. "Species Differentiation of Two Endemic Montane Oaks in China: Population Genetics, Ecological Niche and Leaf Morphology Analyses" Forests 16, no. 3: 549. https://doi.org/10.3390/f16030549
APA StyleChang, Z.-M., Zhang, L.-L., Huang, Y.-J., Chen, X.-D., & Yang, J. (2025). Species Differentiation of Two Endemic Montane Oaks in China: Population Genetics, Ecological Niche and Leaf Morphology Analyses. Forests, 16(3), 549. https://doi.org/10.3390/f16030549