
Adaptations of Pinus yunnanensis Seedlings to Simulated Light Patches: Growth Dynamics and C:N:P Stoichiometry

Abstract
1. Introduction
2. Material and Methods
2.1. Study Site and Soil Preparation
2.2. Experimental Design and Measurements
2.3. Indicator Measurements
2.4. Data Analysis
3. Results and Analysis
3.1. Plant Height and Basal Diameter Increment of P. yunnanensis Seedlings in Different Treatments
3.2. Effects of Treatments on P. yunnanensis Seedling Biomass
3.3. Treatment Effects on C, N, and P Contents and Stoichiometry of P. yunnanensis Seedlings
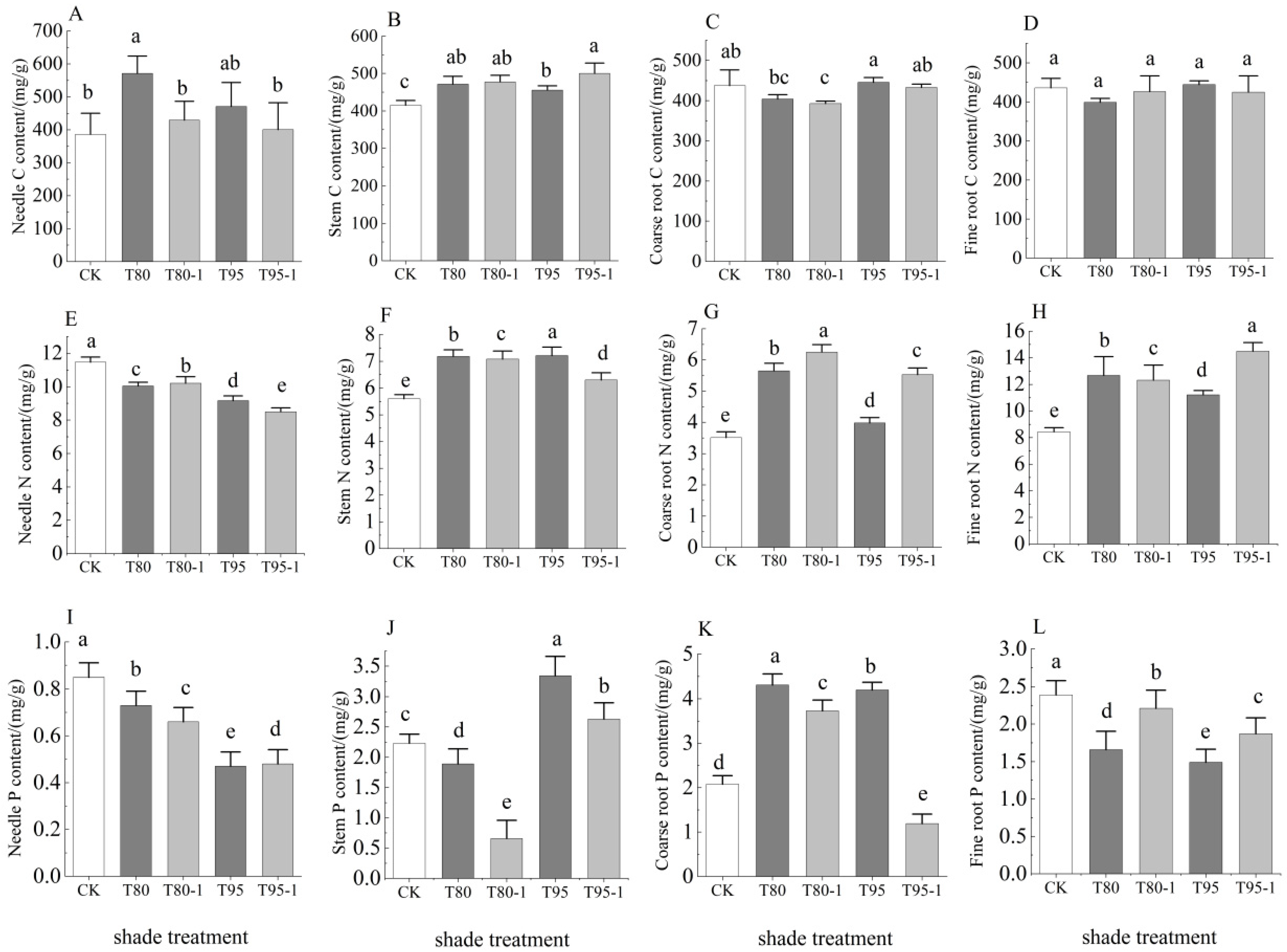
3.3.1. Treatment Effects on C, N, and P Contents of P. yunnanensis Seedlings
3.3.2. C, N, and P Stoichiometry of P. yunnanensis Seedlings Under Different Treatments
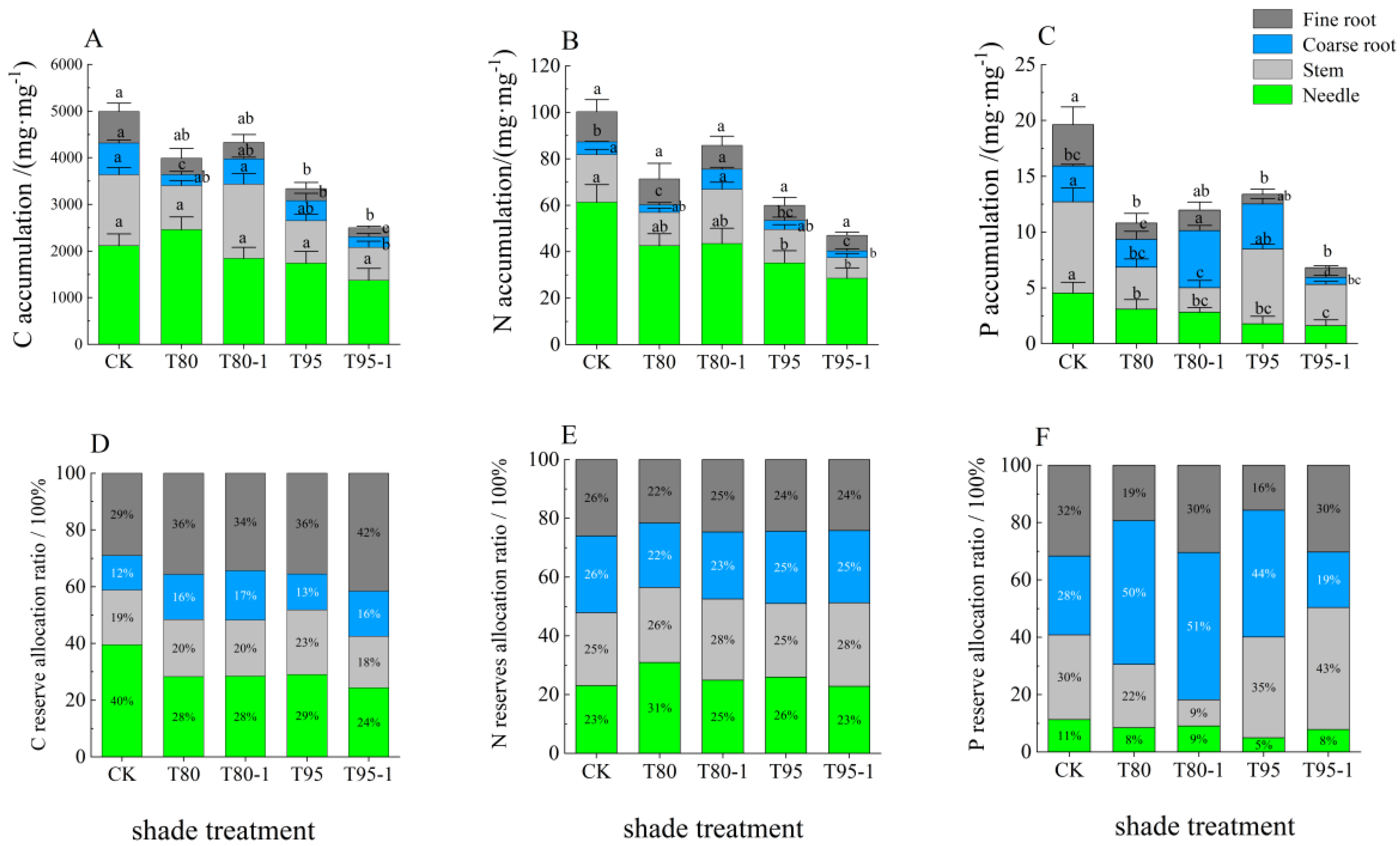
3.3.3. C, N, and P Allocation in the Organs of P. yunnanensis Seedlings Under Different Treatments
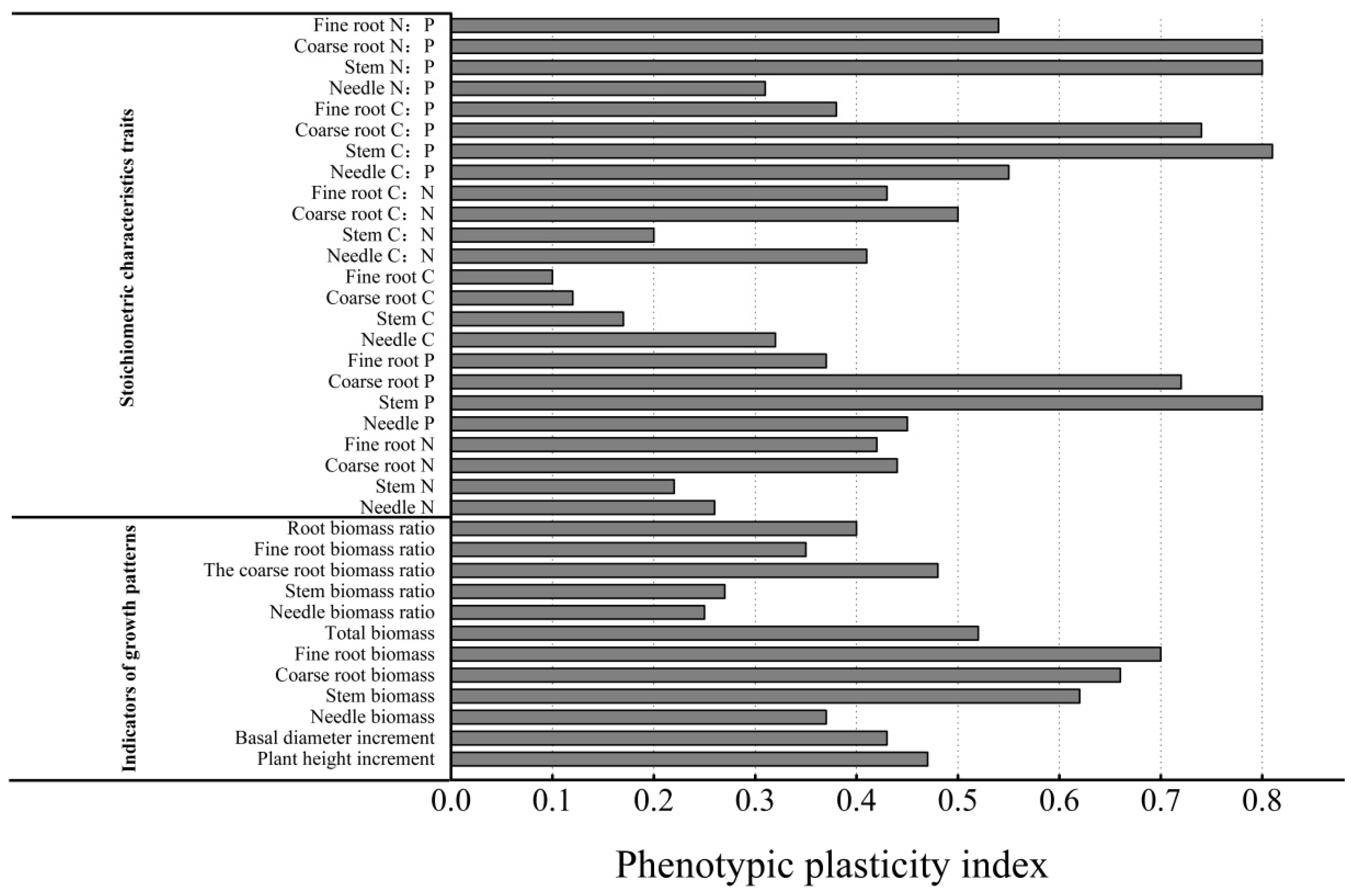
3.4. Phenotypic Plasticity of C, N, and P Stoichiometric Characteristics and Growth Patterns of P. yunnanensis Seedlings Under Different Light Conditions
3.5. Relationship Between C, N, P Stoichiometric and Morphological Characteristics of P. yunnanensis Seedlings
4. Discussion
4.1. Effect of Shade and Light Treatments on the Growth and Biomass Allocation of P. yunnanensis Seedlings
4.2. Effects of the Shade and Light Treatments on C, N, and P Accumulation and the Allocation Ratio of Different Organs of P. yunnanensis Seedlings
4.3. Effects of the Shade and Light Treatments on C:N, C:P, and N:P of Different Organs of P. yunnanensis Seedlings
4.4. Effect of C, N, and P Stoichiometry of Various Organs of P. yunnanensis on Morphological Traits
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rozendaal, D.M.A.; Hurtado, V.H.; Poorter, L. Plasticity in leaf traits of 38 tropical trees in response to light:relationships with light demand and adult stature. Funct. Ecol. 2006, 20, 207–216. [Google Scholar] [CrossRef]
- Liu, Y.X.; Wu, J.W.; Jing, H.Q. Non-structural carbohydrate (NSC) content and C:N:P stoichiometry of Pinus yunnanensis seedling needles in response to shade treatment. Ind. Crops Prod. 2024, 210, 118138. [Google Scholar] [CrossRef]
- Lee, G.B.; Lee, J.E.; Je, B.I.; Lee, Y.J.; Park, Y.H.; Choi, Y.W.; Son, B.G.; Kang, N.J.; Kang, J.S. Effect of low-light intensity on growth, yield and quality of strawberries. J. Environ. Sci. Int. 2020, 29, 167–175. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, L.; Wei, X.; Zhou, H.; Chen, X. Physiological, morphological, and anatomical changes in Rhododendron agastum in response to shading. Plant Growth Regul. 2017, 81, 23–30. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Change Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Ali, A.A.; Xu, C.; Rogers, A.; McDowell, N.G.; Medlyn, B.E.; Fisher, R.A.; Wullschleger, S.D.; Reich, P.B.; Vrugt, J.A.; Bauerle, W.L.; et al. Global-scale environmental control of plant photosynthetic capacity. Ecol. Appl. 2015, 25, 2349–2365. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, G.; Fang, Y.; Wang, T.; Wu, Y.; Wu, Y.; Liu, X.; Jiang, B. Hydrogen peroxide as a systemic messenger in the photosynthetic induction of mulberry leaves. J. For. Res. 2021, 32, 945–952. [Google Scholar] [CrossRef]
- Fan, D.Y.; Xie, Z.Q.; Wang, Q.; Zhang, Q.D. Photosynthetic response of a shrub (Pachysandra terminalis) in a subtropical evergreen broad-leaved forest (English). Chin. J. Plant Ecol. 2002, 4, 447–453. [Google Scholar]
- Ma, Z.L.; Yang, W.Q.; Wu, F.Z.; Gao, S. Effects of shading on the aboveground biomass and stiochiometry characteristics of Medicago sativa. Chin. J. Appl. Ecol. 2014, 25, 3139–3144. [Google Scholar]
- Han, Y.H.; Zhao, Q.L.; Zhang, J.; Sha, H. Effects of Shading on Seedling Growth and C, N, P Stoichiometry Characteristics of Cercidiphyllum japonicum. For. Eng. 2024, 40, 36–46. [Google Scholar]
- Liu, Y.; Xiao, J.; Sun, J.; Zhao, Z.; Deng, X.; Wu, J.; Zhang, D.; Bao, Y. Seasonal variation in C:N:P stoichiometry, nonstructural carbohydrates, and carbon isotopes of two coniferous pioneer tree species in subtropical China. Front. Plant Sci. 2023, 27, 1225436. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, Z.; Gao, C.; Li, S.; Huang, X.; Lang, X.; Su, J. Radial growth response of Pinus yunnanensis to rising temperature and drought stress on the Yunnan Plateau, southwestern China. Forest Ecol. Manag. 2020, 474, 118357. [Google Scholar] [CrossRef]
- Su, W.; Yu, J.; Zhang, G.; Shi, Z.; Wang, L.; Zhao, G.; Zhou, R. Comparison of the canopy and soil seed banks of Pinus yunnanensis in central Yunnan, China. Forest Ecol. Manag. 2019, 437, 41–48. [Google Scholar] [CrossRef]
- Xu, Y.; Woeste, K.; Cai, N.; Kang, X.; Li, G.; Chen, S.; Duan, A. Variation in needle and cone traits in natural populations of Pinus yunnanensis. J. For. Res. 2016, 27, 41–49. [Google Scholar] [CrossRef]
- Wang, X.Z.; Wang, L.F.; Wang, Y.; Huang, Y.; Ding, Z.; Zhou, J.; Gou, D. Identification and genetic analysis of the pinewood nematode Bursaphelenchus xylophilus from Pinus yunnanensis. Forest Pathol. 2015, 45, 388–399. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, Y. Carbon: Nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar]
- Frey, B.R.; Ashton, M.S. Growth, survival and sunfleck response of underplanted red oaks (Quercus spp., section Erythrobalanus) along a topographic gradient in southern New England. For. Ecol. Manag. 2018, 419–420, 179–186. [Google Scholar] [CrossRef]
- Li, K.; Li, Z.J. Soil Agrochemical Analysis Method; China Agricultural Science and Technology Press: Beijing, China, 2019. [Google Scholar]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar]
- Yin, D.; Shen, H.; Wei, X. Effects of Shading on Physocarpus amurensis Seedlings Photosynthetic Ability and Carbohydrate Accumulation. Bull. Bot. Res. 2017, 37, 841–847. [Google Scholar]
- Katahata, S.; Naramoto, M.; Kakubari, Y.; Mukai, Y. Photosynthetic capacity and nitrogen partitioning in foliage of the evergreen shrub Daphniphyllum humile along a natural light gradient. Tree Physiol. 2007, 27, 199–208. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Q.; Daryanto, S.; Guo, S.; Huang, Z.; Wang, Z.; Wang, L.; Ma, X. Responses of Chinese fir and Schima superba seedlings to light gradients: Implications for the restoration of mixed broadleaf-conifer forests from Chinese fir monocultures. For. Ecol. Manag. 2018, 419–420, 51–57. [Google Scholar] [CrossRef]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Pires, M.V.; Almeida, A.A.F.; Figueiredo, A.L.; Gomes, F.P.; Souza, M.M. Photosynthetic characteristics of ornamental passion flowers grown under different light intensities. Photosynthetica 2011, 49, 593–602. [Google Scholar] [CrossRef]
- Poorter, L.; Hayashida-Oliver, Y. Effects of seasonal drought on gap and understorey seedlings in Bolivian moistforest. J. Trop. Ecol. 2000, 16, 481–498. [Google Scholar] [CrossRef]
- Mediavilla, S.; Escudero, A. Differences in biomass allo-cation patterns between saplings of two co-occurring Mediterranean oaks as reflecting different strategies in the use of light and water. Eur. J. For. Res. 2010, 129, 697–706. [Google Scholar] [CrossRef]
- Portsmuth, A.; Niinemets, Ü. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct. Ecol. 2007, 21, 61–77. [Google Scholar] [CrossRef]
- McConnaughay, K.D.M.; Coleman, J.S. Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients. Ecology 1999, 80, 2581–2593. [Google Scholar] [CrossRef]
- Lin, J.; Zhang, R.; Hu, Y.; Song, Y.; Hänninen, H.; Wu, J. Interactive effects of drought and shading on Torreya grandis seedlings: Physiological and growth responses. Trees 2019, 33, 951–961. [Google Scholar] [CrossRef]
- Dickman, E.M.; Newell, J.M.; González, M.J.; Vanni, M.J. Light, nutrients, and food chain length constrain planktonic energy transfer efficiency across multiple trophic levels. Proc. Natl. Acad. Sci. USA 2008, 105, 18408–18412. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, Z.; Ma, X.; Wang, Z.; Xing, X.; Liu, B. Changes of seedling growth and C, N, P stoichiometric characteristics in Chinese fir under shading. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 46, 74–82. [Google Scholar]
- Sun, H.; Zhang, Y.P.; Wu, J.C.; Peng, X.; Zheng, Y. Effect of Drought Stress and Shading on Growth and Carbon-nitrogen Metabolism of Azadirachta indica Seedlings. Acta Bot. Boreali-Occident. Sin. 2020, 40, 463–470. [Google Scholar]
- Gong, J.; Zhang, Z.; Zhang, C.; Zhang, J.; Ran, A. Ecophysiological responses of three tree species to a high altitude environment in the southeastern Tibetan plateau. Forests 2018, 9, 48. [Google Scholar] [CrossRef]
- Sardans, J.; Grau, O.; Chen, H.Y.; Janssens, I.A.; Ciais, P.; Piao, S.; Peñuelas, J. Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Change Biol. 2017, 23, 3849–3856. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Chen, H.Y.; Reich, P.B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat. Commun. 2011, 2, 344. [Google Scholar] [CrossRef] [PubMed]
- Donovan, L.A.; Maherali, H.; Caruso, C.M.; Huber, H.; de Kroon, H. The evolution of the worldwide leaf economics spectrum. Trends Ecol. Evol. 2011, 26, 88–95. [Google Scholar] [CrossRef]
- Ågren, G.I. The C: N: P stoichiometry of autotrophs theory and observations. Ecol. Lett. 2004, 7, 185–191. [Google Scholar] [CrossRef]
- Güsewell, S.; Koerselman, W.; Verhoeven, J.T. BiomassN: P ratios as indicators of nutrient limitation for plant populations in wetlands. Ecol. Appl. 2003, 13, 372–384. [Google Scholar] [CrossRef]
- Klausmeier, C.A.; Litchman, E.; Daufresne, T.; Levin, S.A. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 2004, 429, 171–174. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Unravelling phenotypic plasticity why should we bother. New Phytol. 2006, 170, 644–648. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Biomass/g | ||||
|---|---|---|---|---|---|
| Needles | Stems | Coarse Roots | Fine Roots | Total Biomass | |
| CK | 5.34 ± 0.182 a | 3.67 ± 0.199 a | 1.55 ± 0.023 a | 1.55 ± 0.021 a | 12.10 ± 1.554 a |
| T80 | 4.24 ± 0.123 a | 1.99 ± 0.115 bc | 0.58 ± 0.019 c | 0.88 ± 0.011 b | 7.69 ± 2.082 bc |
| T80-1 | 4.26 ± 0.231 a | 3.31 ± 0.221 ab | 1.38 ± 0.014 a | 0.83 ± 0.013 b | 9.77 ± 1.477 ab |
| T95 | 3.83 ± 0.221 a | 2.00 ± 0.098 bc | 0.96 ± 0.02 b | 0.58 ± 0.012 c | 7.38 ± 2.418 bc |
| T95-1 | 3.36 ± 0.213 a | 1.41 ± 0.068 c | 0.53 ± 0.032 c | 0.46 ± 0.011 c | 5.76 ± 1.717 c |
| Treatment | Biomass Proportion (%) | ||||
|---|---|---|---|---|---|
| Needles | Stems | Coarse Roots | Fine Roots | Roots Crown Ratio | |
| CK | 43.96 | 30.28 | 12.97 | 12.79 | 35.12 |
| T80 | 55.00 | 26.36 | 7.49 | 11.15 | 23.14 |
| T80-1 | 43.59 | 33.46 | 14.33 | 8.61 | 30.36 |
| T95 | 51.31 | 26.92 | 13.25 | 8.52 | 28.05 |
| T95-1 | 58.01 | 24.59 | 9.08 | 8.32 | 21.08 |
| Stoichiometric Characteristics | Order of Importance | Explanatory Rate/% | F | p |
|---|---|---|---|---|
| Fine root N:P | 1 | 40.3 | 8.8 | 0.002 |
| Needle N:P | 2 | 10.0 | 2.4 | 0.018 |
| Coarse root C | 3 | 8.8 | 2.3 | 0.042 |
| Stem C:P | 4 | 8.4 | 2.6 | 0.044 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Liu, Y.; Wu, J.; Li, C. Adaptations of Pinus yunnanensis Seedlings to Simulated Light Patches: Growth Dynamics and C:N:P Stoichiometry. Forests 2025, 16, 517. https://doi.org/10.3390/f16030517
Zhu W, Liu Y, Wu J, Li C. Adaptations of Pinus yunnanensis Seedlings to Simulated Light Patches: Growth Dynamics and C:N:P Stoichiometry. Forests. 2025; 16(3):517. https://doi.org/10.3390/f16030517
Chicago/Turabian StyleZhu, Weisong, Yuanxi Liu, Junwen Wu, and Chaojun Li. 2025. "Adaptations of Pinus yunnanensis Seedlings to Simulated Light Patches: Growth Dynamics and C:N:P Stoichiometry" Forests 16, no. 3: 517. https://doi.org/10.3390/f16030517
APA StyleZhu, W., Liu, Y., Wu, J., & Li, C. (2025). Adaptations of Pinus yunnanensis Seedlings to Simulated Light Patches: Growth Dynamics and C:N:P Stoichiometry. Forests, 16(3), 517. https://doi.org/10.3390/f16030517