Abstract
Coco Island, located 530 km off the Pacific coast of Costa Rica, experiences rainfall of over 7000 mm/year. This study aims to explore the distinctiveness of the growth ring boundaries and the dendrochronological potential (DP) of nine tree species found on Coco Island. The distinctiveness was classified into type 1: density variation; type 2: marginal parenchyma; type 3: fiber/parenchyma pattern; type 4: vessel distribution, and type 5: fiber band. Cecropia pittieri and Henriettella fascicularis lacked growth ring boundaries and, therefore, did not present any DP. Growth ring type 1 occurred in Ardisia compressa, Henriettea succosa, Henriettella odorata, and Psychotria cocosensis. Ocotea insularis and Sacoglottis holdridgei exhibited type 5 growth rings, with sufficient distinctiveness. Finally, Eugenia pacifica possessed growth rings of type 3 and type 4, but with limited distinctiveness in the growth ring boundary. In relation to DP, A. compressa, E. pacifica, and P. cocosensis presented low DP; meanwhile, H. succosa, H. odorata, O. insularis, and S. holdridgei demonstrated medium DP. If these species are found in abundance with a diameter exceeding 60 cm, a cross-dating technique like bomb-spike dating is established to confirm the annual growth of the ‘bands’. As these species exhibit some DP, they can be used to establish a chronology spanning 50–130 years, which could be utilized to create climate proxies or derive ecologically and climatically meaningful information. Additionally, the information presented could pave the way for future studies of sites that also have very high rainfall, where trees are presumed to lack annual growth rings.
1. Introduction
Located 530 km from the continental Pacific Coast of Costa Rica and 680 km northwest of the Galapagos Islands [1], Coco Island is the only island in the Tropical Pacific with precipitation levels above 7000 mm per year [2]. It is spread across a 24 km2 area, covered by protected marine ecosystems of 1980 km2 [1,2].
The growth of vascular plants has been governed by floristic links with continental areas lying northwards of Coco Island, including Central America, the Caribbean islands, and North America [3]. Perhaps the most prevalent dispersal mechanisms, including wind, birds, and ocean currents, carry propagules between Coco Island and continental areas [2]. Notably, the type of vegetation, as well as the climatic conditions of Coco Island and the Galapagos Islands, exhibit remarkable distinctions [1,2].
According to Bonilla-Mata and Acosta-Vargas [4] and Bonilla-Mata [5], Coco Island presents two life zones: the low-elevation humid forest and high-elevation cloud forest. Due to the presence of long-lived trees in these areas, there is promising potential for dendrochronological studies if annual rings are observed [6]. The low-elevation humid forest has relatively low tree diversity, with species including Sacoglottis holdridgei, Clusia rosea, Ocotea insularis, Henriettella fascicularis, Miconia dodecandra, Ficus pertusa, Eugenia cocosensis, and Brosimum sp. [3,7,8]. In contrast, the high-elevation cloud forest begins at an altitude of 450 m and covers the two highest peaks of the island, Cerro Yglesias and Cerro Pelón [3]. Though this forest mainly comprises S. holdridgei, with an understory largely dominated by tree ferns, but other species such as Psychotria cocosensis and Ocotea insularis are also found [7,8].
Despite the floral diversity observed in the vascular plants of Coco Island, there are limited studies that decipher the ecology and dynamics of the two types of forests found on this island [9], and there is a lack of knowledge surrounding the climatic conditions of the island [2].
However, with the presence of diverse forests and the distinct species found on this island, the trees and their possible growth ring formations could facilitate climate reconstruction and further comprehension of the ecology and dynamics of the island. Coco Island experiences a high rainfall of over 7000 mm per year and presents a low duration of daylight [9]; it is considered extremely humid, and the trees of some species could have limited growth rings [10], which provide the main anatomy for dendrochronology studies [11].
Dendrochronology is a technique for identifying and quantifying environmental processes (such as temperature and precipitation) through analysis of the growth rings of multiple vascular plants [12], mainly trees [6]. This technique is largely applied in climatology [13], ecology [14], and geomorphology [15]. In addition to precipitation and temperature, various other factors also impact the formation of growth rings [12,13], including the distribution of rain, the presence of cloud cover, and solar radiation, among many other factors [16].
Knowledge of the ecological tendencies of growth rings is limited [16]. Their significance has been discussed in comparative studies of anatomical features; however, their evolution has barely been mentioned in some studies [13,17]. Several recent studies have cataloged the different anatomical markers and the types of growth rings present across different species in tropical regions and Latin America [16]. Ecological studies on growth rings have been carried out in various regions including Mexico, where growth rings were correlated with variations in environmental controls [17]; Bolivia, where the influence of the total amount and seasonal distribution of rainfall on diameter growth were studied [18]; Costa Rica, where growth rings were examined in the historical vegetation of different forest zones [19]; Colombia, where tree rings were compared with annual rain levels [10], and Brazil, where growth rings in trees of the 22 most representative families of the Brazilian flora were analyzed, and it was revealed that the presence of growth rings was associated with seasonal environments [17].
Although different anatomical features are present in annual rings, there is limited information as to how ecological factors affect the formation of these differences [13,17]. Thus, studies on the distinctiveness of annual rings, and of the different types of rings, would lay the foundations for establishing an ecological trend of growth ring formation and a deeper comprehension of the ecological and climatic traits affecting tree growth [20].
On the other hand, prior to the application of techniques for ecological or climate studies on Coco Island, the presence of growth rings in species growing there must be established. Research on tree rings is fundamental for unraveling historical climate regimes in Central America and on different islands in this region, some of which experience limited tree growth [21]. In continental Costa Rica, the evaluation of growth rings has been limited to a few studies based on climatic reconstructions, ecological approaches, or geomorphic assessments [15,22]. However, a recent study by Moya et al. [19] reviewed and outlined distinctive types of tree growth rings among 320 tree species in correlation with families, genera, species type, growth conditions, altitudinal and life zones, and climate type, for the continental area of Costa Rica.
As this island is far from continental areas, it harbors a unique ecosystem, with limited or no exchange between species growing in this area and the continental landmass [1]. For example, Giraldo et al. [10] presented an interesting study on the formation of growth rings in the American region with the heaviest rainfall, examining a lowland, wet, and non-seasonal tropical forest, with a mean growth rainfall of >7200 mm. This study challenged the traditional assumption that only hydric seasonality, dry periods, or periodic floods are the dominant triggering variables for the formation of growth rings in tropical trees, and showed that other factors also intervene in the formation of growth rings. For example, high cloudiness limits radiation photosynthetically in non-seasonal tropical forests, making this factor crucial during the rainy season. Additionally, the availability of water was also revealed as a highly decisive determining factor for the seasonal variations in growth ring formation in trees, in contrast to the levels of precipitation. Considering the climatic similarities between Coco Island and the study site examined by Giraldo et al. [10], it is plausible that the trees on Coco Island may demonstrate distinct growth ring patterns, which could help to understand the island’s ecology and climatic conditions [23].
Based on the previous studies performed by Giraldo et al. [10], reflecting the development of growth rings even in sites with high rainfall, we hypothesize that trees on Coco Island, characterized by abundant precipitation and geographical isolation, might also exhibit similar patterns. The study aims to identify and categorize distinctive types of tree growth rings, in association with their dendrochronological potential (DP), among nine tree species on Coco Island, to assess their distinctiveness in contrast to the continental species of Central or South America. Thus, the determination of particular species’ growth rings could serve as a basis for performing chronological studies on temperature, precipitation, or other aspects related to dendrochronology. Subsequently, the chronology could be employed as climate proxies or derivatives of ecologically meaningful information. Hence, the growth rings in Coco Island’s trees could provide valuable contributions to our understanding of climate and ecology, thus allowing the development of future research programs concerning the ecology of and inferences about past environmental changes detected through tree rings.
2. Materials and Methods
2.1. Geographical and Climatological Characteristics of Coco Island
Coco Island, positioned 530 km southwest of the Pacific coast of Costa Rica (5°30′–5°34′ N and 87°01′–87°06′ W), presents a dramatic variation in topography, rising from sea level to a height of 575 m [24,25,26]. The island extends 7.6 km from east to west and 4.4 km from southeast to northwest. The average temperature recorded is 27 °C, fluctuating between 27 °C and 29 °C, with the highest temperatures occurring from February to April (Figure 1a). The average rainfall is 7000 mm/year, and its distribution is shown in Figure 1b. November to March is the dry season, with rainfall of 200 mm/month, whereas the rainy season falls between April and October, with precipitation over 500 mm/month [2]. There are two distinct ecological zones on Coco Island, as classified by the Holdridgei life zone system, which represents an ecological zoning system based on the identification of bioclimatic units (life zones); these include the variables of biotemperature, precipitation, potential evapotranspiration (EVP), the EVP/precipitation ratio, latitude, and altitude. This system is used to classify the different ecological zones in the tropical and subtropical regions of Latin America, the United States, China, Russia, and India [27]. The low-elevation humid forest thrives in areas from 0 to 300 m above sea level, whereas the high-elevation cloud forest is found 300 to 620 m above sea level [4]. Coco Island features acidic and clayey soil, with a highly developed organic layer, comprising alfisols and inceptisols [1].
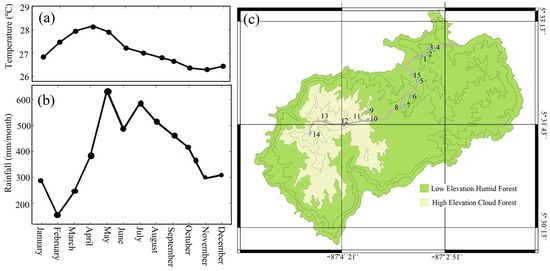
Figure 1.
Variation of temperature (a) and rainfall (b) and localization of 15 permanent plot in two Holdrige life zones present in Coco Island, Pacific Ocean (c). Source modified from Alfaro [8]. Note: the number from 1 to 14 represents permanent plot.
2.2. Tree Sample
The Technological Institute of Costa Rica established 15 permanent plots on Coco Island in 2010 (Figure 1c), with the objective of determining the floristic composition and tracking the growth of the natural forest [5]. These plots were established in areas ranging from sea level to Cerro Yglesias, located at a height of 575 m above sea level, in the two life zones: the low-elevation humid forest and high-elevation cloud forest (Figure 1c). The measurements of trees and floristic composition were carried out in 2017, for the trees exceeding 5 cm in diameter, specifying the most abundant tree species [3,5]. Due to their higher abundance, nine species were selected from this study (Table 1) for further anatomical analysis of the wood and to determine the presence of growth rings. It is important to mention that all species are classified as evergreen species. Two samples of different trees present in the permanent plots were collected for each selected species (Figure 1c). Four out of the nine selected species were endemic to the island, while the other five were native to Costa Rica, and some species were present in both life zones (Table 1). The samples consisted of two increment cores of 12 mm in diameter, with a depth or length of 50 mm, and were extracted using a 12 mm brush placed in a manual drill on two different sides of a tree. The number of growth rings varied from 10 to 15 years. The samples were extracted from the diameter at breast height (1.3 m above the ground) and transferred to the Biomaterials Laboratory of the School of Forestry Engineering in Cartago, Costa Rica. The samples were air-dried on the island, since their transport to Costa Rica’s continental areas could take several days. It was not possible to extract an increment core from bark to pith, due to the constraints linked to obtaining permission to gather a large number of samples. The island is subject to many restrictions set by the government, as Coco Island was declared a national park in 1978 [1,2]; hence, the government limits the studies carried out on this island.

Table 1.
Trees species studied and their distribution, forest, and code xylariorum of TECw from Coco Island of Costa Rica.
2.3. Wood Sample Preparation for Antomical Observation for Rings Boundary
First, the three samples were registered in the Xylariorum of the Technological Institute of Costa Rica (TECw), in accordance with their respective codes, detailed in Table 1. A block (1 cm3) was obtained from each increment core of 12 mm in diameter. It was extracted from an internal part of the core, or the deepest part of the core sample. The block was softened in hot water. Cross sections were cut (12–15 μm thick) and these sections were stained with safranin, then dehydrated with a series of alcohol (5 min each at 50%, 70%, and 95%). Finally, the sections were rinsed and mounted on microscope slides to analyze the growth ring boundary under microscopic conditions. The presence of false rings was not evaluated, due to the short length of the increment core samples.
2.4. Growth Ring Distinctiveness
The remaining increment cores were pasted on one wood support, with the cross-section upwards, and were conditioned until they reached a 12% moisture content (temperature at 22 °C and 66% relative humidity). After, they were sanded with 100, 200, 400, 800 m and 1200 grit sandpaper. Then the sample was observed for the annual rings boundary and its distinctiveness, using a light stereomicroscope with 1.5X to 7.5X magnification, which is appropriate for annual ring visualization [28]. The distinctiveness of the growth ring is associated with anatomical markers [12]. Although there are different types of classifications for growth rings [12], the classification proposed by Worbes et al. [29] and Worbes and Raschke [30] was used in the present research, because of its use in numerous dendrochronological studies [12]. The classification addresses two aspects: (i) distinctiveness is established in three categories: distinctive or boundary defined (+), indistinctive or boundary non-defined (−), and a third more or less distinctive category (+/−); (ii) type of growth ring, which is based on the structure of the growth zones and the anatomy. According to Worbes and Raschke [30], using an light stereomicroscope [28], there are five established categories:
- Type 1, density variation: the growth ring’s boundary is marked by several rows of fibers with a shortened radial diameter and thickened walls;
- Type 2, marginal parenchyma: the growth ring’s boundary is marked with a uniseriate or multiseriate marginal parenchyma band;
- Type 3, fiber/parenchyma pattern: the growth ring’s boundary is marked by periodically recurring patterns of alternating parenchyma and fiber bands;
- Type 4, vessel distribution: there is a variation in the diameter or frequency of vessel elements;
- Type 5, fiber band: the growth ring’s boundary is marked by a band of fibers.
2.5. Dendrochronological Potential
In addition to the previous classification (type of growth rings) of each species, another classification of the growth rings was carried out to provide detailed information about the species in Costa Rica, considering their dendrochronological potential. For this purpose, we referred to Giraldo et al. [10], who proposed four degrees:
- (1)
- Null: growth rings are non-defined;
- (2)
- Low: growth rings are slightly defined; it is difficult to observe growth ring boundaries, there are false growth rings, growth rings are too close together, or they have frequent discontinuity;
- (3)
- Medium: there are false growth rings or few frequent, more or less distinctive, or poorly defined growth rings;
- (4)
- High: growth rings are clearly defined in tangential orientation, and there is no presence of false growth rings.
3. Results
The different types of growth rings found in the nine tree species on Coco Island are detailed in Table 2. In two species (C. pittieri and H. fascicularis), no growth ring boundary was observed under both macroscopic and microscopic conditions (Table 2, Figure 2a,b); the transversal section (macro and microscopic) showed a uniform distribution of vessels, fibers, rays, and parenchyma tissue (Figure 2a,b).
Table 2.
Species studied in relation growth ring type: references, growing condition, altitudinal and life ecology zone, distinctiveness of growth rings, and potential in dendrochronology for different tree species in Costa Rica.

Figure 2.
Macro and microscopic cross-sections in two species growing in Coco Island species with any growth ring boundary.
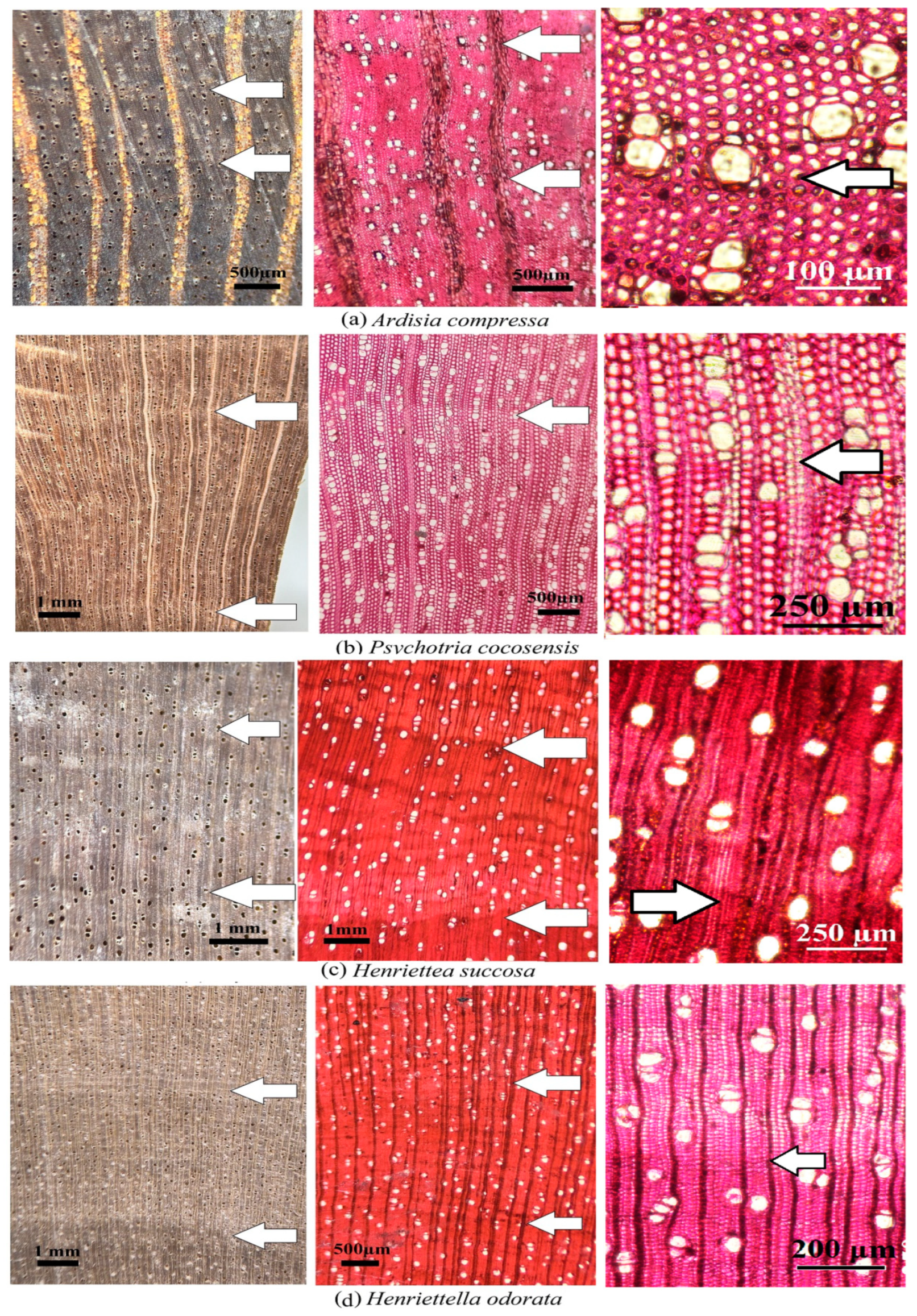
Growth ring type 1 (density variation) was presented in A. compressa, H. succosa, H. odorata, and P. cocosensis (Table 2). Despite the existing density variation (Type 1) in these four species, A. compressa and P. cocosensis exhibited limited density variation due to the presence of only a few rows of thick-walled fibers, which was more easily observed in the microscopic section than in the macroscopic section in four species (Figure 3a–d). However, this density variation was well-defined in H. odorata and H. succosa. The growth ring boundary was easily observed at the microscopic level in H. odorata (Figure 3d); however, at the macroscopic level, it was difficult to observe the annual ring boundaries. Likewise, the vessel diameter tended to be reduced in this species; however, it did not match the growth ring seen in type 4 (Figure 3d). The growth ring boundary in H. succosa was difficult to observe at both microscopic and macroscopic levels (Figure 3c).

Figure 3.
Macro- and microscopic cross-sections in four species (a–d) from Coco Island with growth ring boundary type 1 (density variation). Legend: Arrow represents annual ring boundary.
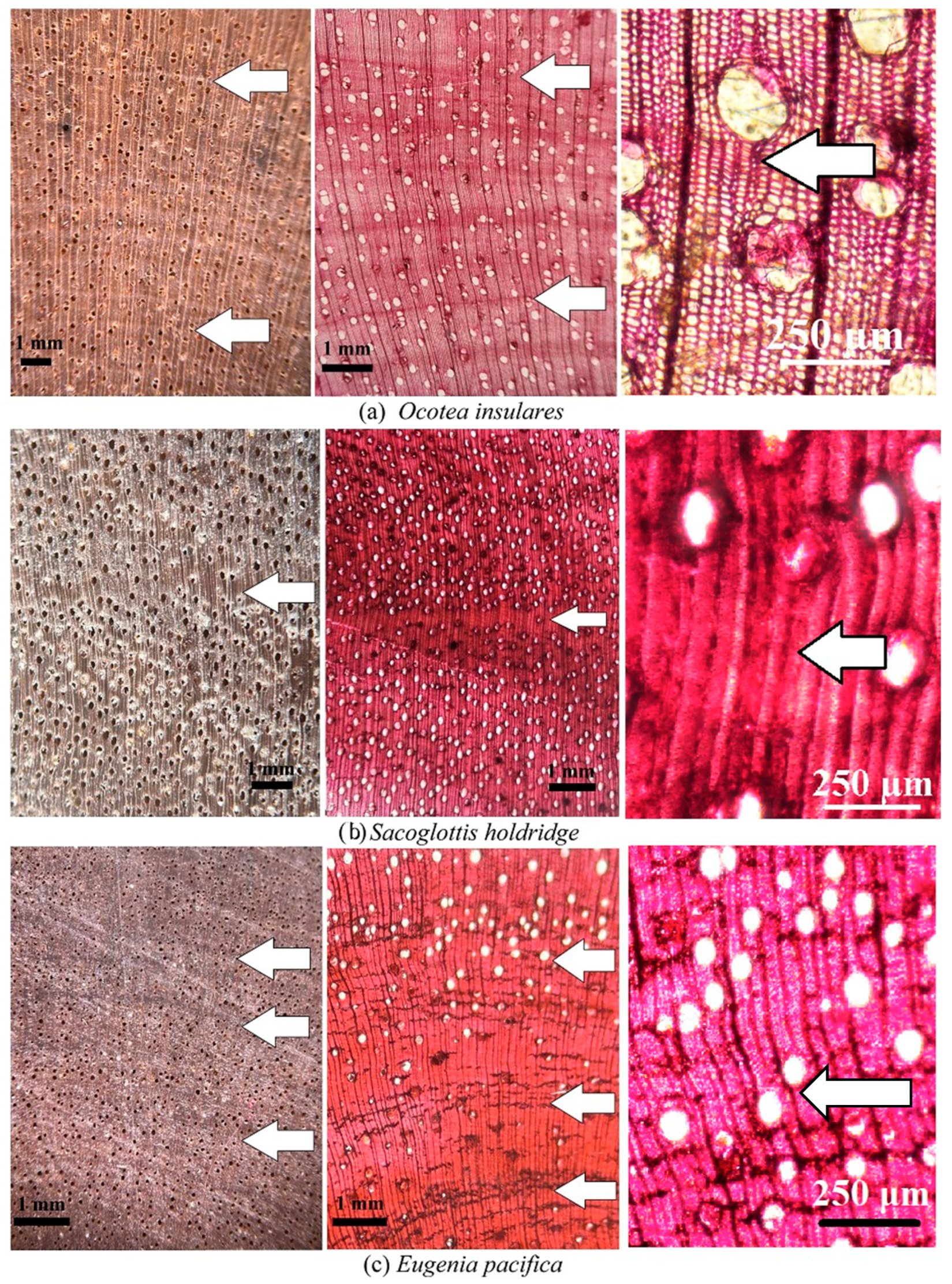
O. insularis and S. holdridgei demonstrated growth ring boundaries of type 5 (fiber band), which were easily observed at microscopic and macroscopic levels (Table 2, Figure 4a,b), with the formation of fiber bands more evident in S. holdridgei (Figure 4b) than in O. insularis (Table 2, Figure 4a). Finally, E. pacifica possessed type 3 (fiber/parenchyma pattern) and type 4 (vessels distribution) growth rings, exhibiting medium distinctiveness at the macroscopic and microscopic levels (Figure 4c). Another important observation was the presence of incomplete fiber bands, forming false growth rings, highlighting the need for further evaluation or careful observation of the growth rings in H. odorata (Figure 3d) and Ocotea insularis (Figure 4a).
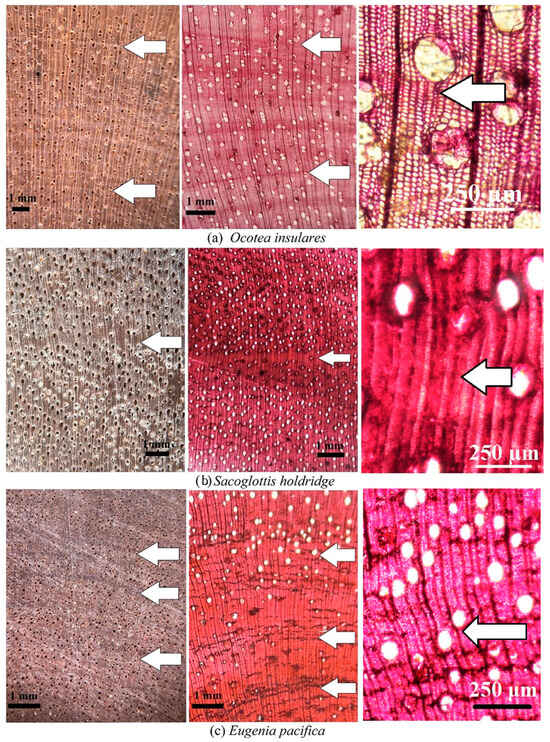
Figure 4.
Macro- and microscopic cross-sections showing growth ring boundary type 5 (fiber band) presents in two species (a,b) and growth ring boundary type 4 (pattern de fiber/parenchyma) in one species (c) from Coco Island; and. Legend: Arrow represents annual ring boundary.
Considering the absence of well-defined growth ring types in any of the species (Table 2), none of the nine tree species found in Coco Island presented high dendrochronological potential. Moreover, two out of the nine studied species presented null dendrochronological potential (C. pittieri and H. fascicularis), whereas the remaining seven species from Coco Island presented growth rings with more or less distinctiveness (+/−). Three of the species (A. compressa, E. pacifica, and P. cocosensis) presented low dendrochronological potential, due to the low distinctiveness of the growth ring boundary. Meanwhile, H. succosa, H. odorata, O. insularis, and S. holdridgei were cataloged with medium dendrochronological potential (Table 2).
4. Discussion
4.1. Growth Ring Presence
The lack of growth ring boundaries (low and null intensity of expression) (Table 2) in five species is attributed to the climatic conditions prevalent on the island and to the origin of the species. The trees on Coco Island limit the possiblility of studies based on dendrochronology, especially for ecological or climatic research endeavors. The species studied in Coco Island, as indicated, are classified as evergreen, and almost all of the species showed limited distinctiveness in the annual ring. These results agreed with the studies carried out on this type of species in Neotropical regions. Marcelo-Peña et al. [31] indicated that evergreen species presented poor visibility of annual rings in countries of the Neotropical region of America, specifically in Peru, Brazil, Colombia, Ecuador, Costa Rica, and some regions of Mexico.
While analyzing the impact of climatic conditions, Giraldo et al. [10] mentioned that among other conditions, the growth rings’ formation in sites with precipitation over 7000 mm/year (considered as tropical hyper-humid conditions) is influenced by seasonal variations in irradiance, the time of sunrise or sunset, and the sunlight intensity. The precipitation and cloudiness values in Coco Island are the highest between November and March [8], and the solar radiation and duration of daylight hours are at their lowest levels in these months [2,8]. These conditions on Coco Island impact the formation of growth rings in tree species, such that high cloudiness limits the availability of photosynthetically active radiation [32]. This factor induces a reduction in flux photosynthetic photon depletion (PPFD), implying that the greater the cloud cover, the greater the reduction in PPFD [10]. Thus, excess water in the soil, cloudiness, and low solar radiation between November and March could result in a reduction in photosynthesis, along with seasonal variations in the vascular cambium and other aspects of tree growth [16], leading to the formation of growth rings in seven of the nine species studied on Coco Island. According to Giraldo et al. [33], annual formation of growth rings in unexplored tropical forests in rainy areas is governed by seasonal growth that occurs due to high soil moisture and hypoxia, or light limitations due to cloudiness and limitations of photosynthesis.
While comparing the findings regarding the presence of growth rings in this study with other studies, surprisingly, 77.8% of the species of Coco Island presented the same values of growth ring distinctiveness (well-defined or more or less defined) as in the continental parts of Costa Rica [19], despite the unfavorable conditions for growth ring formation. Therefore, both regions (island and continental area) shared a similar 22.2% of the species lacking growth ring boundaries. When comparing these percentages with other studies of ring formation in tropical areas with high precipitation over 3000 mm/years (Table 3), Coco Island presented large variety in distinctive and anatomical features in growth ring boundaries. For example, a similar percentage was observed to those found in the rainy regions of Choco in Colombia, which reported 18% of species having no growth ring boundaries, but the authors concluded that without any hydric seasonality, tropical species that produced annual ring totalled 82% [10], a percentage similar to the species from Coco Islands. The author explained that this annual ring formation is probably due to the growing season occurring during the least rainy months [34], as determined by the tree-ring intra-annual stable isotopes (δ13C and δ18O) [35]. Our results, presented in Table 3, show that sites with high precipitation, or prevailing droughts or floods, are not conditions for growth ring formation in tropical trees.
Table 3.
Studies carried out in relation to growth ring formation in sites with high precipitation in tropical areas.
Density variation (type 1) was the predominant growth ring type displayed in four out of nine species of trees on Coco Island (A. compressa, H. succosa, H. odorata, and P. cocosensis, Figure 3). This type of growth ring is more frequent in tropical species [31]. The variation in density can be attributed to the variations in the shape and dimensions of the fiber, especially those featuring radially-flattened and thick-walled latewood fibers, where there is gradual variation in the thick-walled fibers of earlywood transforming into latewood due, to the fluctuation in cambial activity [12].
The marking of the growth ring boundary with marginal parenchyma is typical in many tropical species [31]; however, this growth ring boundary marker was not present in any of the studied species. The marginal parenchyma in tropical species is associated with cambial activity; in some genera, such as Copaifera, it appears before cambial dormancy [21], whereas in other tropical species, such as Tectona or Cedrela species, marginal parenchyma is observed after cambial activity [31]. The climatic conditions on Coco Island are not favorable for the formation of this growth ring boundary.
Although the lack of growth rings in H. fascicularis (Figure 2b) observed in this study agrees with previous studies of H. fascicularis performed in Peru [31], conflicting findings exist for several species of Henriettella trees growing in continental Central America. While a few studies reported the presence of growth ring boundaries, other species lacked growth ring boundaries [39], although the climatic growth conditions were not mentioned in this particular study.
Although Giraldo et al. [10] did not study C. pittieri explicitly, they reported the difference between the growth rings of C. pittieri and C. virgusa, which grows in over-humid tropical conditions in Colombia. Nevertheless, Brienen et al. [24] mentioned that the ambiguity demonstrated in the growth rings of C. pelata and other species of this genus, which is considered a typical Neotropical pioneer taxon, poses too many issues for its straightforward evaluation in growth ring studies. This remains true regardless of the observation of growth rings in Cecropia in more seasonal climates, as evidenced in the floodplain forests of Brazil [25] and Colombia [10]. Thus, the lack of growth ring boundaries in C. pittieri growing on Coco Island (Figure 2a) confirms the low dendrochronological potential.
The comparison of the findings of this study pertaining to the presence of growth rings with other studies showed that no species of the genus Ardisia have been reported to have growth rings. Welle and Koek-Noorman [39] reported that H. succosa trees growing in different parts of continental areas of Central America do not produce growth rings, which contradicts the findings of our study (Figure 3a, Table 2). The existence of growth rings in H. odorata (Figure 3d) agrees with the fact that several species of Henriettea grow in continental Central America, where the trees exhibit the presence of growth rings.
The presence of growth rings in P. cocosensis aligns with the presence of growth rings in several species of Psychotria in Brazil [26,27], which is characterized by thick-walled and radially-flattened latewood fibers, versus thin-walled earlywood fibers (Figure 3b).
This study unveils the fiber band marking (type 5) observed in two species, O. insularis and S. holdridgei (Table 2). Although the causes of the formation of this type of growth ring are not fully understood, Silva et al. [12] mentioned that the fiber band is produced by the seasonal dynamics of the vascular cambium, and its reaction to various environmental, physiological, and genetic factors [16]. According to the literature, the majority of Lauraceae species occur in the Ocotea, Nectandra, Cinnamomum, and Persea genera [40,41,42]. Thus, O. insularis displayed normal behavior in relation to the other genus of the Laureceae family, S. holdridgei, with growth ring type 5. S. trichogyna also presents a growth ring due to the gradual variation in the thickening of the fibers from earlywood to latewood, found in the continental zone of Costa Rica [19]. S. guianensis and Sacoglottis sp., growing in the Pantanal region of Brazil [16] and the Chocó Biogeographic Region of Colombia [10], respectively, also present this growth ring boundary.
An identical growth ring structure was observed in E. pacifica and other species of this genus, for example, E. monticola and E. salamansis, growing in the continental parts of Costa Rica, with similar rainfall levels around 4000 mm/year [39]. However, unlike in the study of E. pacifica, Moya et al. [19] did not mention the presence of parenchyma bands in these two species, despite the absence of growth rings. Two species (E. sonderiana and E. uniflora) growing in Brazil [43,44], in sites with precipitation between 1400 and 2400 mm/year, exhibited a similar type of growth ring boundary pattern and fiber/parenchyma (growth ring type 3). Another important aspect of E. pacifica, is that this species reflects the type of growth ring found in the Myristicaeae family, which poses difficulties in identifying the rings due to the fiber/parenchyma combination and renders it a species with low dendrochronological potential [3,24].
4.2. The Dendrochronological Potential for Climate and Ecology Studies
None of the species on Coco Island presented high dendrochronological potential, however, seven species presented a limited degree of dendrochronological potential (Table 2). Considering the species with medium dendrochronological potential, H. succosa and H. odorata are classified as native, and the other two species (O. insularis and S. holdridgei) are referred to as endemic to the island (Table 1); hence, the dendrochronological potential of these four species makes them suitable for dendrochronological studies. Another important consideration is that O. insularis and S. holdridgei grow in two different altitudinal zones and Holdridgei life zones on the island, unlike the other two species (H. succosa and H. odorata) that are only present in the low-elevation humid forest (Table 1). Therefore, the studies related to ecology and climate in these two conditions can be implemented or utilized as either climate proxies, or sources of ecologically meaningful information, as tree rings in these Holdridgei life zones make valuable contributions to our understanding of the climate and ecology.
Although Coco Island has limited species of long-lived trees, displaying limited tree diversity in general [2,3,4,5,7], four species were found to have growth rings, which makes them suitable for dendrochronology studies applicable to ecological, climatic, morphological, climatic change, and other research domains. However, this was a preliminary study to evaluate the dendrochronological potential of the species growing on this island; additional studies must be performed, such as the bomb-spike dating technique, to confirm that the growth rings are annual in nature, which might enhance the applicability of findings on growth ring formation in future research endeavors.
Recently, Porras-Jimenez et al. [7], Bonilla-Mata [5], and Bonilla-Mata and Acosta-Vargas [4] reported the population status of the trees in Coco Island and determined that S. holdridgei was the most abundant species with 160 trees ha−1. A higher abundance was observed in trees with a diameter less than 40 cm, however, trees with a diameter close to 60 cm can be found in some parts of the island. For O. insularis, 70 trees ha−1, less than the number of S. holdridgei, have been reported; however, the advantage is that the trees of this species are found to have a diameter between 60 and 70 cm. These data and the fact that these two species presented growth ring boundaries easily observed at microscopic and macroscopic levels (Figure 4a,b), suggests that S. holdridgei and O. insularis are two species with high dendrochronological potential. In addition, the growth rings must be confirmed in the two species in trees with a larger diameter, as the sampled trees in this study were small (Table 1).
According to Bonilla-Mata and Acosta-Vargas [4], H. succosa presented a growth rate of 0.19–0.26 cm/year, H. odorata presented 0.25 cm/year, O. insularis presented 0.25–0.31 cm/year, and S. holdridgei presented 0.19–0.26 cm/year. Based on these growth rates, and assuming a minimum diameter of each species [4,7,9], H. succosa trees with a diameter of 50 cm are suggested to have a varying chronology between 75 and 100 years, from 77 to 100 years for H. odorata, from 50 to 75 years for O. insularis, and from 80 to 130 years for S. holdridgei. Then, this chronology can be increased using data reported on temperature and precipitation for Coco Island, which has a reported chronology from 1958, by Quiros-Badilla and Alfaro [44]. The predictions of precipitation and temperature were calculated from time series of the grid points closest to the island, obtained from different data bases based on and compiled from surface gauge stations [44]. In this way, we can see the influence of climate variables on annual rings and are able to study the role of climate and disturbances on forest dynamics [45].
The modeling of diameter growth of trees using annual rings is a technique frequently used for the planning and management of forests. because it can determine life periods, sivicultural treatment in cases of forest management, and the dynamics of tree growing, among many other applications [46]. Thus, the two species with annual rings that are distinctive (S. holdridgei and O. insularis) may have long chronologies. In this way, it is possible to study the dynamics of tree growth on the island using indirect methods, with permanent plots established [3,5], and decrease the economic costs of measuring these plots, which demands more time and cost, because the island is located so far from the continent [47].
Recently, two important topics have been governed for tropical forests: one related to biomass accumulation in trees and another one in relation to climate change, so annual rings are an important tool for these studies [48]. The determination of volume increment by mathematical models, as previously detailed, for shorter or longer periods, allows the calculation of stored biomass or sequestered carbon [48]. The results of this study of growth rings distinctions will allow us to know the function of captured carbon and its fixation in regions located so far from the content, such in as Coco Island, which has the function of a carbon sink, changing the pH conditions of the oceans [49].
5. Conclusions
While Coco Island has a low abundance of species, specifically with limited diversity in the upper canopy, four species have been predominantly identified to grow in the two altitudinal and/or life zones present on the island. The abundance of these species provides them with strong dendrochronological potential. H. succosa, H. odorata, O. insularis, and S. holdridgei present a more or less distinctive growth ring boundary, however, the annual development pattern of growth rings still remains to be determined. If the growth rings are successfully defined for species with DP, they could serve either as a climate proxy, or as a source of ecologically meaningful information, thus emphasizing the valuable contributions made by tree rings in Coco Island to a better understanding of the climate and ecology. Additionally, the findings of this study could lay the foundation for reviewing sites with very high rainfall, that were assumed to lack annual rings in the past.
Author Contributions
Conceptualization, R.M. and C.T.; methodology, R.M., L.A.-V. and C.T.; validation, R.M. and L.A.-V.; formal analysis, R.M. and C.T.; investigation, R.M. and C.T.; resources, R.M., L.A.-V. and C.T.; writing—original draft preparation, R.M. and C.T.; writing—review and editing, R.M., L.A.-V. and C.T.; availability, L.A.-V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data measured for this article are few and can requested from corresponding authors.
Acknowledgments
The authors wish to thank the Vicerrectoría de Investigación y Extensión of the Instituto Tecnológico de Costa Rica (ITCR) for the economic support given to this research. Many thanks also to Juan Carlos Castanedo and Personal of Parque Nacional Isla del Coco for providing support in sampling recollection.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Caludino-Sales, V. Cocos Island National Park, Costa Rica. In Coastal World Heritage Sites. Coastal Research Library; Springer: Dordrecht, The Netherlands, 2019; Volume 28. [Google Scholar] [CrossRef]
- Cortes, J. Isla del Coco: Coastal and Marine Ecosystems. In Costa Rica Ecosystem; Kappelle, M., Ed.; The University of Chicago Press: Chicago, IL, USA, 2016; pp. 97–191, Chapter 7. [Google Scholar]
- Estrada-Chavarría, A.; Sánchez-González, J.; Rodríguez-González, A. Catálogo actualizado de las plantas vasculares del Parque Nacional Isla del Coco, Costa Rica. Rev. Biol. Trop. 2020, 68, 73–88. [Google Scholar] [CrossRef]
- Bonilla-Mata, R.; Acosta-Vargas, L.G. Dynamic and growth of the forests of the Isla del Coco National Park, Costa Rica. Rev. Biol. Trop. 2020, 68, 89–102. [Google Scholar] [CrossRef]
- Bonilla-Mata, R. Evaluación de los bosques del Parque Nacional Isla del Coco, Costa Rica. Bachelor’s Thesis, Escuela de Ingeniería Forestal, Instituto Tecnológico de Costa Rica, Cartago, Costa Rica, 2017; 145p. [Google Scholar]
- Pearl, J.K.; Keck, J.R.; Tintor, W.; Siekacz, L.; Herrick, H.M.; Meko, M.D.; Pearson, C.L. New frontiers in tree-ring research. Holocene 2020, 30, 923–941. [Google Scholar] [CrossRef]
- Porras-Jiménez, M.A.; Acosta-Vargas, L.G.; Castillo-Ugalde, M.; Quesada-Monge, R. Estructura y composición florística del bosque nuboso de la Isla del Coco. Rev. Tecnol. Marcha 2014, 27, 22–36. [Google Scholar] [CrossRef]
- Alfaro, E.J. Ciclo diario y anual de variables troposfericas y oceanicas en la Isla del Coco, Costa Rica (Pacifico Oriental). Rev. Biologia Trop. 2008, 56 (Suppl. 2), 19–29. [Google Scholar]
- Vargas, J.L.; Alfaro, E.J. Radiación ultravioleta (UV) en el Parque Nacional Isla del Coco, Costa Rica. Rev. Biol. Trop. 2016, 64, 75–86. [Google Scholar] [CrossRef]
- Giraldo, J.A.; Valle, J.I.; Sierra, C.A.; Melo, O. Dendrochronological potential of trees from America’s rainiest region. In Latin American Dendroecology: Combining Tree-Ring Sciences and Ecology in a Megadiverse Territory; Pompa-García, M., Camarero, J.J., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 79–119. [Google Scholar]
- Esper, J.; Krusic, P.J.; Ljungqvist, F.C.; Luterbacher, J.; Carrer, M.; Cook, E.; Davi, N.K.; Hartl-Meier, C.; Kirdyanov, A.; Konter, O.; et al. Ranking of tree-ring based temperature reconstructions of the past millennium. Quat. Sci. Rev. 2016, 145, 134–151. [Google Scholar] [CrossRef]
- Silva, M.; Funch, L.S.; da Silva, L.B. The growth ring concept: Seeking a broader and unambiguous approach covering tropical species. Biol. Rev. 2019, 94, 1161–1178. [Google Scholar] [CrossRef]
- Locosselli, G.M.; Brienen, R.J.; Leite, M.D.S.; Gloor, M.; Krottenthaler, S.; Oliveira, A.A.D.; Barichivich, J.; Anhuf, D.; Ceccantini, G.; Schöngart, H.; et al. Global tree-ring analysis reveals rapid decrease in tropical tree longevity with temperature. Proc. Natl. Acad. Sci. USA 2020, 117, 33358–33364. [Google Scholar] [CrossRef]
- Quesada-Román, A.; Ballesteros-Cánovas, J.A.; George, S.S.; Stoffel, M. Tropical and subtropical dendrochronology: Approaches, applications, and prospects. Ecol. Indic. 2022, 144, 109506. [Google Scholar] [CrossRef]
- Quesada-Román, A.; Ballesteros-Cánovas, J.A.; Granados-Bolaños, S.; Birkel, C.; Stoffel, M. Improving regional flood risk assessment using flood frequency and dendrogeomorphic analyses in mountain catchments impacted by tropical cyclones. Geomorphology 2022, 396, 108000. [Google Scholar] [CrossRef]
- Alves, E.S.; Angyalossy-Alfonso, V. Ecological trends in the wood anatomy of some Brazilian species. 1. Growth rings and vessels. IAWA J. 2000, 21, 3–30. [Google Scholar] [CrossRef]
- Sánchez-Calderón, O.D.; Carlón-Allende, T.; Mendoza, M.E.; Villanueva-Díaz, J. Dendroclimatology in Latin America: A Review of the State of the Art. Atmosphere 2022, 13, 748. [Google Scholar] [CrossRef]
- Pacheco-Solana, A.; Oelkers, R.; D’Arrigo, R.; Santos, G.M.; Rodriguez-Caton, M.; Tejedor, E.; Ferrero, E.; Fuentes, A.F.; Maldonado, C.; Andreu-Hayles, L. Radiocarbon and wood anatomy as complementary tools for generating tree-ring records in Bolivia. Front. Plant Sci. 2023, 14, 1135480. [Google Scholar] [CrossRef]
- Moya, R.; Quesada-Román, A.; Quesada-Monge, R. Potential use of Costa Rican wood data for neotropical dendrochronology. New For. 2024; submitted. [Google Scholar]
- Aragão, J.R.V.; Groenendijk, P.; Lisi, C.S. Dendrochronological potential of four neotropical dry-forest tree species: Climate-growth correlations in northeast Brazil. Dendrochronologia 2019, 53, 5–16. [Google Scholar] [CrossRef]
- Fontana, C.; Pérez-de-Lis, G.; Santini Junior, L.; Botosso, P.C.; Nabais, C.; Tomazello-Filho, M.; Lousada, J.L.P.C. Wood anatomy and growth ring boundaries of Copaifera lucens (Fabaceae). IAWA J. 2018, 39, 395–405. [Google Scholar] [CrossRef]
- Quesada-Román, A.; Ballesteros-Cánovas, J.A.; Guillet, S.; Madrigal-González, J.; Stoffel, M. Neotropical Hypericum irazuense shrury reveal recent ENSO variability in Costa Rican páramo. Dendrochronologia 2020, 61, 125704. [Google Scholar] [CrossRef]
- López, R.; Cano, F.J.; Rodríguez-Calcerrada, J.; Sangüesa-Barreda, G.; Gazol, A.; Camarero, J.J.; Rozenberg, P.; Gil, L. Tree-ring density and carbon isotope composition are early-warning signals of drought-induced mortality in the drought tolerant Canary Island pine. Agricul. For. Meteorol. 2021, 310, 108634. [Google Scholar] [CrossRef]
- Brienen, R.J.; Lebrija-Trejos, E.; Van Breugel, M.; Pérez-García, E.A.; Bongers, F.; Meave, J.A.; Martínez-Ramos, M. The potential of tree rings for the study of forest succession in southern Mexico. Biotropica 2009, 41, 186–195. [Google Scholar] [CrossRef]
- Marques, J.B.; Callado, C.H.; Rabelo, G.R.; Silva Neto, S.J.D.; Cunha, M.D. Comparative wood anatomy of species of Psychotria L. (Rubiaceae) in Atlantic Rainforest remnants of Rio de Janeiro State, Brazil. Acta Bot. Brasilica 2015, 29, 433–444. [Google Scholar] [CrossRef]
- Campbell, G.; Mielke, M.S.; Rabelo, G.R.; Da Cunha, M. Key anatomical attributes for occurrence of Psychotria schlechtendaliana (Müll. Arg.) Müll. Arg. (Rubiaceae) in different successional stages of a tropical moist forest. Flora 2018, 246, 33–41. [Google Scholar] [CrossRef]
- Reis-Avila, G.; Oliveira, J.M. Lauraceae: A promising family for the advance of neotropical dendrochronology. Dendrochronologia 2017, 44, 103–116. [Google Scholar] [CrossRef]
- Worbes, M. One hundred years of tree-ring research in the tropics—A brief history and an outlook to future challenges. Dendrochronologia 2002, 20, 217–231. [Google Scholar] [CrossRef]
- Godoy-Veiga, M.; Slotta, F.; Alecio, P.C.; Ceccantini, G.; Buckeridge, M.S.; Locosselli, G.M. Improved tree-ring visualization using autofluorescence. Dendrochronologia 2019, 55, 33–42. [Google Scholar] [CrossRef]
- Worbes, M.; Raschke, N. Carbon allocation in a Costa Rican dry forest derived from tree ring analysis. Dendrochronologia 2012, 30, 231–238. [Google Scholar] [CrossRef]
- Marcelo-Peña, J.L.; Roig, F.A.; Goodwin, Z.A.; Tomazello-Filho, M. Characterizing growth rings in the trees of Perú: A wood anatomical overview for potential applications in dendroecological-related fields. Dendrochronologia 2020, 62, 125728. [Google Scholar] [CrossRef]
- Quintilhan, M.T.; Santini, L., Jr.; Rodriguez, D.R.O.; Guillemot, J.; Cesilio, G.H.M.; Chambi-Legoas, R.; Nouvellon, Y.; Tomazello-Filho, M. Growth-ring boundaries of tropical tree species: Aiding delimitation by long histological sections and wood density profiles. Dendrochronologia 2021, 69, 125878. [Google Scholar] [CrossRef]
- Giraldo, J.A.; del Valle, J.I.; González-Caro, S.; David, D.A.; Taylor, T.; Tobón, C.; Sierra, C.A. Tree growth periodicity in the ever-wet tropical forest of the Americas. J. Ecol. 2023, 111, 889–902. [Google Scholar] [CrossRef]
- Giraldo, J.A.; del Valle, J.I.; González-Caro, S.; Sierrra, C.A. Intra-annual isotope variations in tree rings reveal growth rhythms within the least rainy season of an ever-wet tropical forest. Trees 2022, 36, 1039–1052. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Hu, J.; Dorado-Liñán, I.; Gagen, M.; Szejner, P.; Chen, T.; Trouet, V. Intra-annual tree-ring δ18O and δ13C reveal a trade-off between isotopic source and humidity in moist environments. Tree Physiol. 2022, 42, 2203–2223. [Google Scholar] [CrossRef]
- Vetter, R.E.; Botosso, P.C. Remarks on age and growth rate determination of Amazonian trees. IAWA J. 1989, 10, 133–145. [Google Scholar] [CrossRef]
- Groenendijk, P.; Sass-Klaassen, U.; Bongers, F.; Zuidema, P.A. Potential of tree-ring analysis in a wet tropical forest: A case study on 22 commercial tree species in Central Africa. For. Ecol. Manag. 2014, 323, 65–78. [Google Scholar] [CrossRef]
- Azim, A.A.; Okada, N. Occurrence and anatomical features of growth rings in tropical rainforest trees in Peninsular Malaysia: A preliminary study. Tropics 2014, 23, 15–31. [Google Scholar] [CrossRef]
- Welle, B.J.H.; Jifke Koek-Noorman, J. Wood anatomy of the Neotropical Melastomataceae. Blumea 1981, 27, 335–394. [Google Scholar]
- León-Hernández, W.J. Anatomía xilemática caulinar de 14 especies de la familia Lauraceae. Rev. For. Venez. 2002, 46, 15–25. [Google Scholar]
- León, H.W. Anatomía de la madera de 31 especies de la subfamilia mimosaceae (leguminaceae) en Venezuela. Rev. Colombia For. 2008, 11, 113–135. [Google Scholar]
- Marqués, P.A.; Callado, C.H.; Barros, C.F.; Costa, C.G. Variação intraespecífica do lenho de Eugenia uniflora L. em duas diferentes fitofisionomias do complexo vegetacional atlântico. Floresta Ambiente 2012, 19, 483–496. [Google Scholar] [CrossRef]
- Beltrán, L.A.; Valencia, G.M. Anatomía de anillos de crecimiento de 80 especies arbóreas potenciales para estudios dendrocronológicos en la Selva Central, Perú. Rev. Biología Trop. 2013, 61, 1025–1037. [Google Scholar]
- Quirós-Badilla, E.; Alfaro, E. Algunos aspectos relacionados con la variabilidad climática en la Isla del Coco, Costa Rica. Rev. Climatol. 2009, 9, 33–44. [Google Scholar]
- Bahtiar, E.T.; Iswanto, A.H. Annual tree-ring curve-fitting for graphing the growth curve and determining the increment and cutting cycle period of sungkai (Peronema canescens). Forests 2023, 14, 1643. [Google Scholar] [CrossRef]
- Andreu-Hayles, L.; Tejedor, E.; D’arrigo, R.; Locosselli, G.M.; Rodríguez-Catón, M.; Daux, V.; Oelkers, R.; Pacheco-Solana, A.; Paredes-Villanueva, K.; Rodríguez-Morata, C. Dendrochronological advances in the tropical and subtropical Americas: Research priorities and future directions. Dendrochronologia 2023, 81, 126124. [Google Scholar] [CrossRef]
- Moreno, M.L.; Jiménez, K.; Villalobos, C. Approximation of the benefits of socioeconomic activities in Cocos Island National Park and the effects of climate change. Rev. Interamer. Amb. Tur. 2021, 17, 14–26. [Google Scholar] [CrossRef]
- Cabon, A.; Kannenberg, S.A.; Arain, A.; Babst, F.; Baldocchi, D.; Belmecheri, S.; Delpierre, N.; Guerrieri, R.; Maxwell, J.T.; McKenzie, S.; et al. Cross-biome synthesis of source versus sink limits to tree growth. Science 2022, 376, 758–761. [Google Scholar] [CrossRef]
- Gruber, N.; Bakker, D.C.; DeVries, T.; Gregor, L.; Hauck, J.; Landschützer, P.; McKinley, G.A.; Müller, J.D. Trends and variability in the ocean carbon sink. Nat. Rev. Earth Environ. 2023, 4, 119–134. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).