
Incredible Host Diversity and Regional Potential Distribution of an Oriental Parasitic Plant (Taxillus yadoriki)


Abstract
1. Introduction
2. Materials and Methods
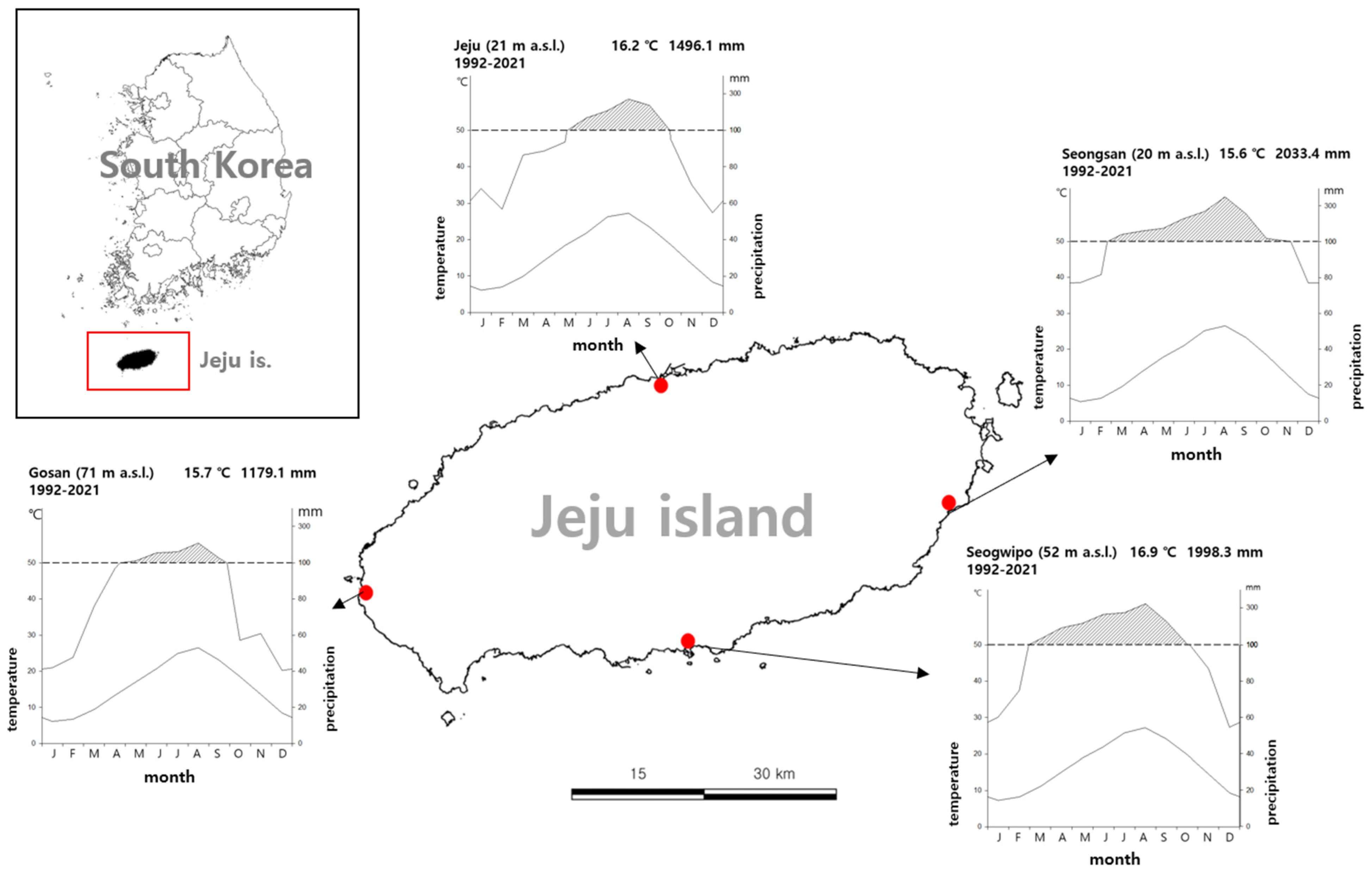
2.1. Field Research
2.2. Distribution Modeling
2.2.1. Data Collection
2.2.2. Data Processing and Selection
2.2.3. Modeling
3. Results and Discussion
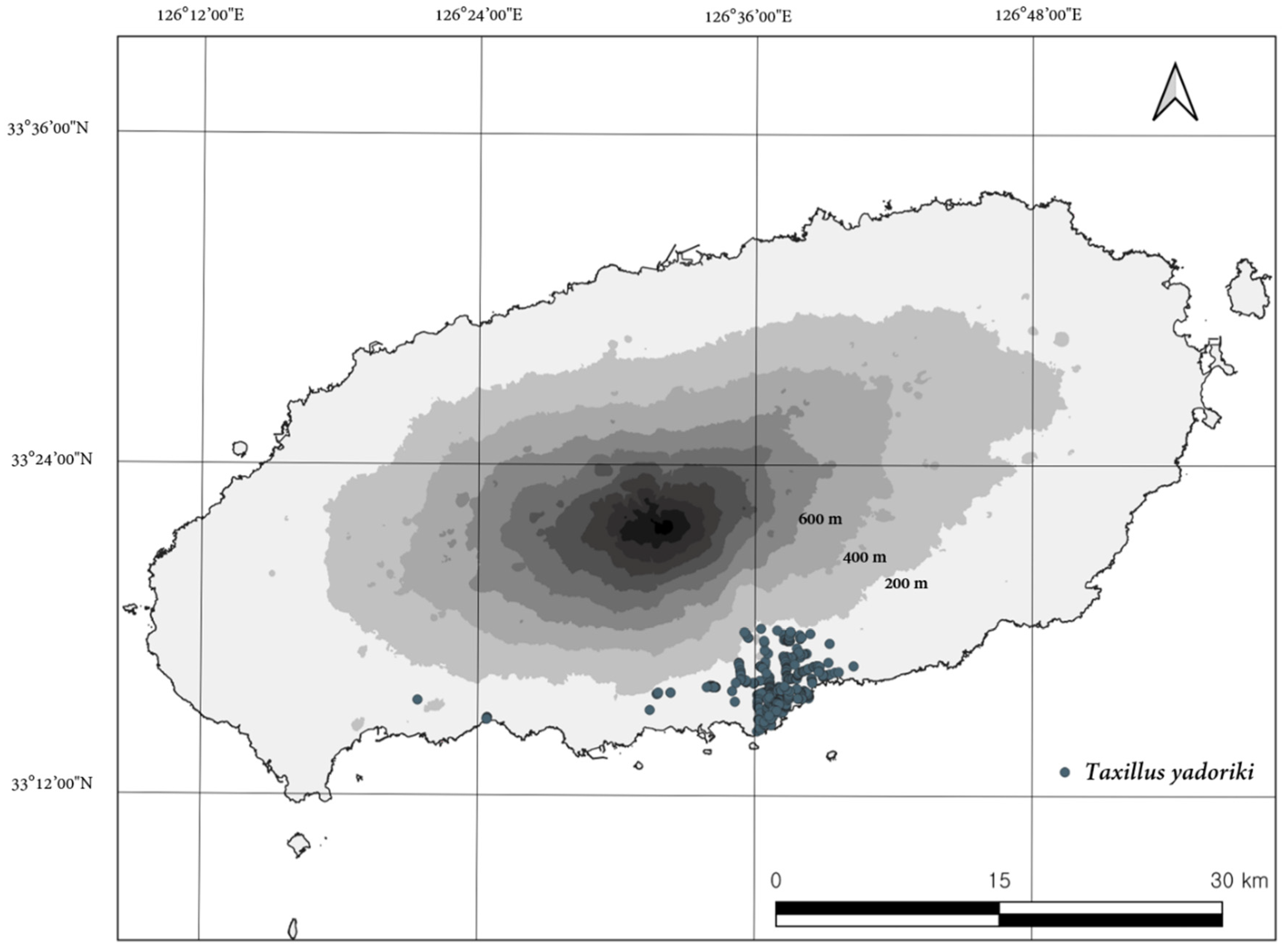
3.1. Distribution Range of Native Species
3.2. Host Plant Diversity and Species Characteristics
3.3. Attachment Characteristics within the Host Plant
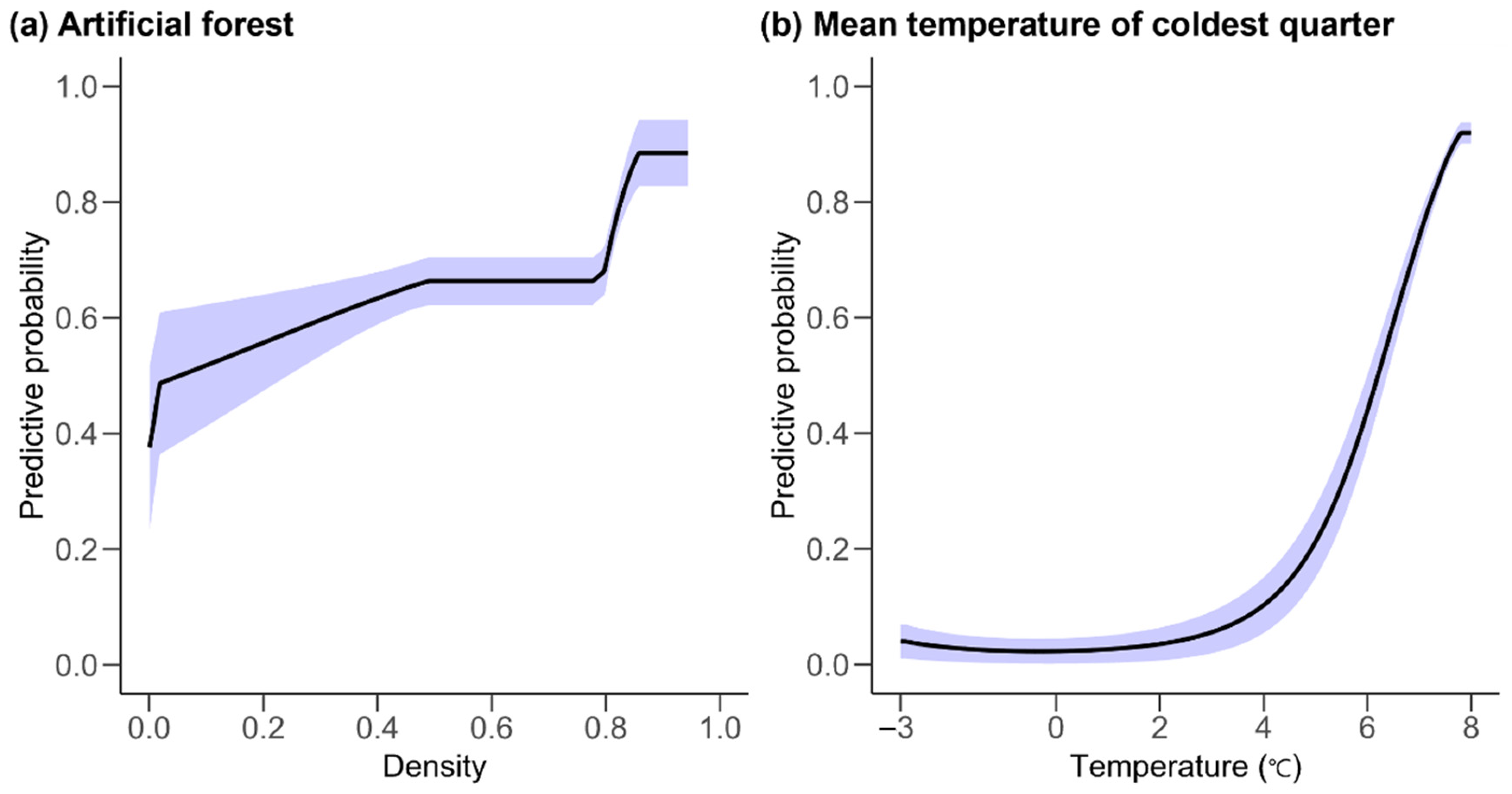
3.4. Species Distribution Model
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musselman, L.J.; Press, M.C. Introduction to parasitic plants. In Parasitic Plants; Pres, M.C., Graves, J.D., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1995; pp. 1–13. [Google Scholar]
- Nickrent, D.L. Parasitic plants of the world. In Parasitic Plants of the Iberian Peninsula and Balearic Islands; López-Sáez, J.A., Catalán, P., Sáez, L., Eds.; Mundi-Prensa Libros: Madrid, Spain, 2002; pp. 7–27. [Google Scholar]
- Nickrent, D.L. Parasitic angiosperms: How often and how many? Taxon 2020, 69, 5–27. [Google Scholar] [CrossRef]
- Douglas, A.E. Symbiotic Interactions; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Thompson, R.L.; Poindexterss, D.B. Host specificity of American mistletoe (Phoradendron leucarpum, Viscaceae) in Garrard County, Kentucky. J. Kentucky Acad. Sci. 2005, 66, 40–43. [Google Scholar] [CrossRef]
- Twyford, A.D. Parasitic plants. Curr. Biol. 2018, 28, 857–859. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.D. Santalaceae R.Br. In The Genera of vascular Plants of Korea; Park, C.W., Ed.; Academy Publishing, Co.: Seoul, Republic of Korea, 2007; pp. 653–655. [Google Scholar]
- Kim, Y.D. Loranthaceae Juss. In The Genera of vascular Plants of Korea; Park, C.W., Ed.; Academy Publishing Co.: Seoul, Republic of Korea, 2007; pp. 656–657. [Google Scholar]
- Těšitel, J. Functional biology of parasitic plants: A review. Plant Ecol. Evol. 2016, 149, 5–20. [Google Scholar] [CrossRef]
- Fukunaga, T.; Nishiya, K.; Kajikawa, I.; Takeya, K.; Itokawa, H. Studies on the constituents of Japanese mistletoes from different host trees, and their antimicrobial and hypotensive properties. Chem. Pharm. Bull. 1989, 37, 1543–1546. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kim, S.Y.; Sun, B.Y.; Yi, J.S. A review of the taxonomic and ecological characteristics of Korean mistletoe types (Viscum, Korthalsella, Loranthus and Taxillus). Korean J. Plant Tax. 2013, 43, 81–89. [Google Scholar] [CrossRef]
- Ishida, H.; Kuroda, A.; Iwakiri, K. Overgrowth of the hemiparasitic plant species Taxillus yadoriki (Loranthaceae) and the debilitation of its host trees in a park in Miyazaki City, Miyazaki Prefecture. Veg. Sci. 2016, 33, 15–32. [Google Scholar]
- GBIF (Global Biodiversity Information Facility). Biodiversity Research. 2022. Available online: https://www.gbif.org (accessed on 12 December 2022).
- NIBR (National Institute of Biological Resources). Species Information Search. 2022. Available online: https://www.nibr.go.kr (accessed on 12 December 2022).
- Satake, Y.; Hara, H.; Watari, S.; Tominari, T. Wild Flowers of Japan (Woody Plants); Heibonsha: Tokyo, Japan, 1989; pp. 100–102. [Google Scholar]
- KNA (Korea National Arboretum). Rare Plants Data Book in Korea; Korea National Arboretum: Seoul, Republic of Korea, 2008. [Google Scholar]
- KBIS (Korea Biodiversity Information System). Species Information Search. 2022. Available online: http://www.nature.go.kr (accessed on 12 December 2022).
- Griebel, A.; Watson, D.; Pendall, E. Mistletoe, friend and foe: Synthesizing ecosystem implications of mistletoe infection. Environ. Res. Lett. 2017, 12, 115012. [Google Scholar] [CrossRef]
- Bell, D.M.; Pabst, R.J.; Shaw, D.C. Tree growth declines and mortality were associated with a parasitic plant during warm and dry climatic conditions in a temperate coniferous forest ecosystem. Glob. Chang. Biol. 2020, 26, 1714–1724. [Google Scholar] [CrossRef]
- Park, S.B.; Park, G.H.; Kim, H.N.; Son, H.J.; Song, H.M.; Kim, H.S.; Jeong, J.B. Anti-inflammatory effect of the extracts from the branch of Taxillus yadoriki being parasitic in Neolitsea sericea in LPS-stimulated RAW264. 7 cells. Biomed. Pharmacother. 2018, 104, 1–7. [Google Scholar] [CrossRef]
- Park, S.B.; Park, G.H.; Kim, H.N.; Song, H.M.; Song, H.J.; Park, J.A.; Kim, H.S.; Jeong, J.B. Ethanol extracts from the branch of Taxillus yadoriki parasitic to Neolitsea sericea induces cyclin D1 proteasomal degradation through cyclin D1 nuclear export. BMC Complement. Altern Med. 2018, 18, 189. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Kim, H.N.; Kim, J.D.; Park, G.H.; Eo, H.J.; An, M.H.; Jeong, J.B. Inhibitory effect of the branch extracts from Taxillus yadoriki parasitic to Neolitsea sericea against the cell proliferation in human lung cancer cells, A549. Korean J. Plant Res. 2019, 32, 109–115. [Google Scholar]
- Lee, T.B. Illustrated Flora of Korea; Hyangmunsa: Seoul, Republic of Korea, 2004. [Google Scholar]
- Lee, W.T. Lineamenta Flora Korea; Academy Press: Seongnam, Republic of Korea, 1996. [Google Scholar]
- Mucina, L.; Rutherford, M.C.; Powrie, L.W. The logic of the map: Approaches and procedures. In The Vegetation of South Africa, Lesotho and Swaziland; Mucina, L., Rutherford, M.C., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2006; pp. 13–30. [Google Scholar]
- Numata, M. Ecological judgement of grassland condition and trend: I. Judgement by biological spectra. J. Jpn. Soc. Grassl. Sci. 1965, 11, 20–33. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- MoE (Ministry of Environment) Land Cover Map. 2022. Available online: https://egis.me.go.kr (accessed on 12 December 2022).
- QGIS.org. QGIS Geographic Information System. Available online: http://www.qgis.org (accessed on 1 March 2024).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org (accessed on 1 March 2024).
- Padalia, H.; Srivastava, V.; Kushwaha, S.P.S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 2014, 22, 36–43. [Google Scholar] [CrossRef]
- Negrini, M.; Fidelis, E.G.; Picanço, M.C.; Ramos, R.S. Mapping of the Steneotarsonemus spinki invasion risk in suitable areas for rice (Oryza sativa) cultivation using MaxEnt. Exp. Appl. Acarol. 2020, 80, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.E.; Zimmermann, N. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Ngarega, B.K.; Chaibva, P.; Masocha, V.F.; Saina, J.K.; Khine, P.K.; Schneider, H. Application of MaxEnt modeling to evaluate the climate change effects on the geographic distribution of Lippia javanica (Burm. f.) Spreng in Africa. Environ. Monit. Assess. 2024, 196, 62. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J. A Brief Tutorial on Maxent. Network of Conservation Educators and Practitioners, Center for Biodiversity and Conservation, American Museum of Natural History. Lessons Conserv. 2009, 3, 108–135. [Google Scholar]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Shi, J.; Xia, M.; He, G.; Gonzalez, N.C.; Zhou, S.; Lan, K.; Ouyang, L.; Shen, X.; Jiang, X.; Cao, F.; et al. Predicting Quercus gilva distribution dynamics and its response to climate change induced by GHGs emission through MaxEnt modeling. J. Environ. Manag. 2024, 357, 120841. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Hernández, M.J.P.; Báez, W.L.; Ramos, J.H.; Flores, H.J.M.; Uicab, J.V.C.; Santos, M.D.R. The ecological niche as a tool for predicting potential areas of two pine species. Mex. J. For. Sci. 2018, 9, 47–68. [Google Scholar]
- KMA (Korea Meteorological Administration). Climate Data. 2022. Available online: https://www.weather.go.kr (accessed on 12 December 2022).
- Lee, S.G. Distribution, habitat characteristics and assessment of the conservation status of a rare mistletoe species, Loranthus tanakae (Loranthaceae) in Korea. J. Korean Forest Soc. 2013, 102, 428–436. [Google Scholar]
- Lee, B.D. Ecological environment of native habitats and host plant in mistletoe (Viscum album var. coloratum). Korean J. Plant Res. 2009, 22, 389–393. [Google Scholar]
- Venturelli, M. Estudos sobre Struthanthus vulgaris Mart.: Anatomia do fruto e semente e aspectos de germinac¸ a. o, crescimento e desenvolvimento. Rev. Bras. Bot. 1981, 4, 131–147. [Google Scholar]
- Kim, T.Y.; Kim, J.S. Woody Plants of the Korean Peninsula; Dolbegae: Paju, Republic of Korea, 2018. [Google Scholar]
- Choi, K.; Park, K.W.; Kim, H.J.; Lee, J.D.; Koo, J.C.; Whang, S.S. Anatomy of the Korean mistletoe and their haustorial features in host plants. Korean J. Plant Taxon. 2009, 39, 4–11. [Google Scholar] [CrossRef]
- Yoshikawa, E.; Yoneda, M.; Miyamoto, K.; Iwanaga, F.; Yamamoto, F. Leaf water relations and assimilation in Scurruta yadoriki and its host species. Tree For. Hearth 2009, 13, 58–66. [Google Scholar]
- Makino, T. Makino’s Illustrated Pocket Book of Japanese in Colour; Hokuryukan Co.: Tokyo, Japan, 1985. [Google Scholar]
- Guerra, T.J.; Pizo, M.A.; Silva, W.R. Host specificity and aggregation for a widespread mistletoe in Campo Rupestre vegetation. Flora 2018, 238, 148–154. [Google Scholar] [CrossRef]
- Okubamichael, D.Y.; Griffiths, M.E.; Ward, D. Host specificity in parasitic plants—Perspectives from mistletoes. AoB Plants 2016, 8, 69. [Google Scholar] [CrossRef]
- Norton, D.A.; Carpenter, M.A. Mistletoes as parasites: Host specificity and speciation. Trends Ecol. Evol. 1998, 13, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Tworek, S. Different bird strategies and their responses to habitat changes in an agricultural landscape. Ecol. Res. 2002, 17, 339–359. [Google Scholar] [CrossRef]
- Brambilla, M.; Assandri, G.; Martino, G.; Bogliani, G.; Pedrini, P. The importance of residual habitats and crop management for the conservation of birds breeding in intensive orchards. Ecol. Res. 2015, 30, 597–604. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | Climate Zone | Host Diversity |
|---|---|---|---|---|
| Santalaceae | Viscum | V. album var. lutescens | Cool-temperate zone | Fagaceae, Betulaceae, Aceraceae etc. |
| V. album f. rubroauranticum | Cool-temperate zone | Fagaceae, Betulaceae etc. | ||
| Korthalsella | K. japonica | Warm-temperate and subtropical zone | Theaceae, Oleaceae, Lauraceae, etc. | |
| Loranthaceae | Loranthus | L. tanakae | Cool-temperate zone | Fagaceae, Betulaceae etc. |
| Taxillus | T. yadoriki | Warm-temperate and subtropical zone | Lauraceae, Taxodiaceae, Fagaceae etc. |
| Order | Family | Scientific Name | Newly Discovered |
|---|---|---|---|
| Ginkgoales | Ginkgoaceae | Ginkgo biloba L. | ○ |
| Cupressales | Cupressaceae | Cryptomeria japonica (Thunb. ex L.f.) D.Don | |
| Pinales | Taxaceae | Torreya nucifera (L.) Siebold & Zucc. | ○ |
| Fagales | Fagaceae | Castanopsis sieboldii (Makino) Hatus. | |
| Quercus acutissima Carruth. | |||
| Quercus glauca Thunb. | |||
| Quercus mongolica var. crispula H.Ohashi | ○ | ||
| Quercus serrata Murray | |||
| Magnoliales | Ulmaceae | Aphananthe aspera (Thunb.) Planch. | |
| Celtis sinensis Pers. | |||
| Ulmus parvifolia Jacq. | ○ | ||
| Moraceae | Ficus erecta Thunb. | ○ | |
| Ficus erecta var. sieboldii | ○ | ||
| Magnoliaceae | Magnolia kobus DC. | ||
| Magnolia obovata Thunb. | ○ | ||
| Illiciaceae | Illicium anisatum L. | ○ | |
| Lauraceae | Actinodaphne lancifolia Meisn. | ||
| Cinnamomum camphora (L.) J.Presl | |||
| Cinnamomum yabunikkei H.Ohba | |||
| Litsea japonica (Thunb.) Juss. | |||
| Machilus thunbergii Siebold & Zucc. ex Meisn. | |||
| Neolitsea aciculata (Blume) Koidz. | |||
| Neolitsea sericea (Blume) Koidz. | |||
| Rosales | Theaceae | Camellia japonica L. | |
| Camellia sasanqua Thunb. | |||
| Camellia × williamsii W.W.Sm. | ○ | ||
| Eurya emarginata (Thunb.) Makino | ○ | ||
| Eurya japonica Thunb. | |||
| Hamamelidaceae | Distylium racemosum Siebold & Zucc. | ||
| Rosaceae | Eriobotrya japonica (Thunb.) Lindl. | ||
| Prunus mume (Siebold) Siebold & Zucc. | |||
| Prunus persica (L.) Stokes | ○ | ||
| Prunus salicina Lindl. | ○ | ||
| Prunus serrulata f. spontanea Chin S.Chang | |||
| Prunus spachiana f. ascendens Kitam. | ○ | ||
| Prunus × yedoensis Matsum. | |||
| Pseudocydonia sinensis C.K.Schneid. | ○ | ||
| Pyrus pyrifolia var. culta Nakai | ○ | ||
| Geraniales | Rutaceae | Citrus natsudaidai Hayata | ○ |
| Citrus unshiu Marcow. | ○ | ||
| Sapindales | Aceraceae | Acer palmatum Thunb. | |
| Acer palmatum var. sanguineum Nakai | ○ | ||
| Myrtales | Myrtaceae | Acca sellowiana Burret | ○ |
| Elaeagnaceae | Elaeagnus umbellata Thunb. | ○ | |
| Cornales | Cornaceae | Cornus kousa F.Buerger ex Hance | ○ |
| Ericales | Ericaceae | Rhododendron yedoense f. poukhanense Sugim. ex T.Yamaz. | ○ |
| Ebenaceae | Diospyros kaki Thunb. | ||
| Gentianales | Styracaceae | Styrax japonicus Siebold & Zucc. | |
| Apocynaceae | Nerium oleander L. | ○ |
| Family | Scientific Name | Korea | Others a |
|---|---|---|---|
| Ginkgoaceae | Ginkgo biloba L. | + | · |
| Cryptomeria japonica (Thunb. ex L.f.) D.Don | + | + | |
| Cupressaceae | Metasequoia glyptostroboides Hu et W.C.Cheng | · | + |
| Taxaceae | Torreya nucifera (L.) Siebold & Zucc. | + | · |
| Salicaceae | Populus angulata Aiton | · | + |
| Betulaceae | Carpinus tschonoskii Maxim. | · | + |
| Fagaceae | Castanea crenata Siebold et Zucc. | · | + |
| Castanopsis cuspidata (Thunb.) Schottky | · | + | |
| Castanopsis sieboldii (Makino) Hatus. | + | + | |
| Lithocarpus edulis (Makino) Nakai | · | + | |
| Quercus acuta Thunb. | · | + | |
| Quercus acutissima Carruth. | + | + | |
| Quercus gilva Blume | · | + | |
| Quercus glauca Thunb. | + | + | |
| Quercus mongolica var. crispula (Blume) H.Ohashi | + | · | |
| Quercus phillyraeoides A. Gray | · | + | |
| Quercus salicina Blume | · | + | |
| Quercus serrata Murray | + | + | |
| Ulmaceae | Aphananthe aspera (Thunb.) Planch. | + | + |
| Celtis sinensis Pers. | + | + | |
| Ulmus parvifolia Jacq. | + | · | |
| Zelkova serrata (Thunb.) Makino | · | + | |
| Moraceae | Ficus erecta Thunb. | + | · |
| Ficus erecta var. sieboldii | + | · | |
| Magnoliaceae | Magnolia kobus DC. | + | + |
| Magnolia obovata Thunb. | + | · | |
| Illiciaceae | Illicium anisatum L. | + | · |
| Lauraceae | Actinodaphne acuminata (Blume) Meisn. | · | + |
| Actinodaphne lancifolia (Siebold & Zucc.) Meisn. | + | + | |
| Cinnamomum camphora (L.) J.Presl | + | + | |
| Cinnamomum yabunikkei H.Ohba | + | + | |
| Litsea japonica (Thunb.) Juss. | + | · | |
| Machilus japonica Siebold et Zucc. ex Blume | · | + | |
| Machilus thunbergii Siebold & Zucc. ex Meisn. | + | + | |
| Neolitsea aciculata (Blume) Koidz. | + | + | |
| Neolitsea sericea (Blume) Koidz. | + | + | |
| Theaceae | Camellia japonica L. | + | + |
| Camellia japonica var. macrocarpa Masam. | · | + | |
| Camellia sasanqua Thunb. | + | + | |
| Camellia × williamsii W.W.Sm. | + | · | |
| Cleyera japonica Thunb. | · | + | |
| Eurya emarginata (Thunb.) Makino | + | · | |
| Eurya japonica Thunb. | + | + | |
| Ternstroemia gymnanthera (Wight et Arn.) Bedd. | · | + | |
| Hamamelidaceae | Distylium racemosum Siebold & Zucc. | + | + |
| Pittosporaceae | Pittosporum tobira (Thunb.) W.T.Aiton | · | + |
| Rosaceae | Eriobotrya japonica (Thunb.) Lindl. | + | · |
| Prunus mume (Siebold) Siebold & Zucc. | + | + | |
| Prunus persica (L.) Stokes | + | · | |
| Prunus salicina Lindl. | + | · | |
| Prunus serrulata Lindl. f. spontanea (E.H.Wilson) Chin S.Chang | + | + | |
| Prunus spachiana (Lavallée ex Ed.Otto) Kitam. f. ascendens (Makino) Kitam. | + | · | |
| Prunus × yedoensis Matsum. | + | + | |
| Pseudocydonia sinensis (Thouin) C.K.Schneid. | + | · | |
| Pyrus pyrifolia var. culta (Makino) Nakai | + | · | |
| Euphorbiaceae | Mallotus japonicus (L.f.) Müll.Arg. | · | + |
| Rutaceae | Citrus natsudaidai Hayata | + | · |
| Citrus unshiu (Yu.Tanaka ex Swingle) Marcow. | + | · | |
| Anacardiaceae | Toxicodendron succedaneum (L.) Kuntze | · | + |
| Aceraceae | Acer morifolium Koidz. | · | + |
| Acer palmatum Thunb. | + | + | |
| Acer palmatum var. matsumurae (Koidz.) Makino | · | + | |
| Acer palmatum var. sanguineum Nakai | + | · | |
| Aquifoliaceae | Ilex integra Thunb. | · | + |
| Celastraceae | Euonymus japonicus Thunb. | · | + |
| Elaeocarpaceae | Elaeocarpus sylvestris var. ellipticus (Thunb.) H. Hara | · | + |
| Elaeagnaceae | Elaeagnus pungens Thunb. | · | + |
| Elaeagnaceae | Elaeagnus umbellata Thunb. | + | · |
| Myrtaceae | Acca sellowiana (O.Berg) Burret | + | · |
| Cornaceae | Cornus kousa F.Buerger ex Hance | + | · |
| Cornus macrophylla Wall. | · | + | |
| Araliaceae | Schefflera heptaphylla (L.) Frodin | · | + |
| Ericaceae | Rhododendron kaempferi Planch. | · | + |
| Rhododendron tashiroi Maxim. | · | + | |
| Rhododendron yedoense Maxim. f. poukhanense (H.Lév.) Sugim. ex T.Yamaz. | + | · | |
| Myrsinaceae | Ardisia sieboldii Miq. | · | + |
| Primulaceae | Myrsine seguinii H.Lév. | · | + |
| Ebenaceae | Diospyros kaki Thunb. | + | + |
| Diospyros morrisiana Hance | · | + | |
| Styracaceae | Styrax japonicus Siebold & Zucc. | + | + |
| Symplocaceae | Symplocos prunifolia Siebold et Zucc. | · | + |
| Apocynaceae | Nerium oleander L. | + | · |
| Character | Type | Proportion (%) | |
|---|---|---|---|
| Host character | Life form | Tree | 82.0 |
| Shrub | 18.0 | ||
| Leaf functional group | Deciduous broad-leaved tree | 50.0 | |
| Evergreen broad-leaved tree | 44.0 | ||
| Deciduous, coniferous tree | 2.0 | ||
| Evergreen coniferous tree | 4.0 | ||
| Distribution | Native | 66.0 | |
| Non-native | 34.0 | ||
| Parasitic character | Position adhered to | Top | 31.7 |
| Middle | 62.7 | ||
| Bottom | 5.5 | ||
| Type of branch adhered to | Trunk | 21.3 | |
| Branch | 58.1 | ||
| Twig | 20.7 | ||
| Sp. Code | Freq | L/F | L/T | Seas | Nat | Sp. Code | Freq | L/F | L/T | Seas | Nat | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | CrJa | 189 | T | Co | EG | E | 26 | IlAn * | 2 | T | Br | EG | N |
| 2 | LiJa | 109 | S | Br | EG | N | 27 | NeAc | 2 | T | Br | EG | N |
| 3 | PrxYe | 88 | T | Br | SG | N | 28 | PrPe * | 2 | T | Br | SG | P |
| 4 | MaTh | 65 | T | Br | EG | N | 29 | QuSe | 2 | T | Br | SG | N |
| 5 | NeSe | 61 | T | Br | EG | N | 30 | UlPa * | 2 | T | Br | SG | N |
| 6 | CaJa | 23 | T | Br | EG | N | 31 | AcSe * | 1 | S | Br | EG | P |
| 7 | MaKo | 22 | T | Br | SG | N | 32 | AcPav.s * | 1 | T | Br | SG | P |
| 8 | AcPa | 19 | T | Br | SG | E | 33 | ApAs | 1 | T | Br | SG | N |
| 9 | CeSi | 17 | T | Br | SG | N | 34 | CaxWi * | 1 | T | Br | EG | P |
| 10 | DiKa | 15 | T | Br | SG | P | 35 | CaSa | 1 | T | Br | EG | P |
| 11 | QuQl | 11 | T | Br | EG | N | 36 | CiCa | 1 | T | Br | EG | N |
| 12 | DiRa | 6 | T | Br | EG | N | 37 | CiNa * | 1 | T | Br | EG | P |
| 13 | PrSpf.a * | 6 | T | Br | SG | N | 38 | CoKo * | 1 | T | Br | SG | N |
| 14 | CiYa | 5 | T | Br | EG | N | 39 | ElUm * | 1 | S | Br | SG | N |
| 15 | PsSi * | 5 | T | Br | SG | P | 40 | EurJa | 1 | S | Br | EG | N |
| 16 | PrSa * | 4 | T | Br | SG | P | 41 | FiEr * | 1 | S | Br | SG | N |
| 17 | StJa | 4 | T | Br | SG | N | 42 | FiErv.s * | 1 | S | Br | SG | N |
| 18 | ErJa | 3 | T | Co | EG | P | 43 | MaOb * | 1 | T | Br | SG | P |
| 19 | NeOl * | 3 | S | Br | EG | P | 44 | PrMu | 1 | T | Br | SG | P |
| 20 | RhYef.p * | 3 | S | Br | EG | E | 45 | PrSef.s | 1 | T | Br | SG | N |
| 21 | AcLa | 2 | T | Br | EG | N | 46 | PyPyv.c * | 1 | T | Br | SG | P |
| 22 | CaSi | 2 | T | Br | EG | N | 47 | QueAc | 1 | T | Br | SG | N |
| 23 | CiUn * | 2 | T | Br | EG | P | 48 | QuMov.c * | 1 | T | Br | SG | N |
| 24 | EuEm * | 2 | S | Br | EG | N | 49 | ToNu * | 1 | T | Co | EG | N |
| 25 | GiBi * | 2 | T | Co | SG | P | Total | 697 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.; Choi, B.; Park, E.; Yim, E. Incredible Host Diversity and Regional Potential Distribution of an Oriental Parasitic Plant (Taxillus yadoriki). Forests 2024, 15, 799. https://doi.org/10.3390/f15050799
Lee C, Choi B, Park E, Yim E. Incredible Host Diversity and Regional Potential Distribution of an Oriental Parasitic Plant (Taxillus yadoriki). Forests. 2024; 15(5):799. https://doi.org/10.3390/f15050799
Chicago/Turabian StyleLee, Cheolho, Byoungki Choi, Eunha Park, and Eunyoung Yim. 2024. "Incredible Host Diversity and Regional Potential Distribution of an Oriental Parasitic Plant (Taxillus yadoriki)" Forests 15, no. 5: 799. https://doi.org/10.3390/f15050799
APA StyleLee, C., Choi, B., Park, E., & Yim, E. (2024). Incredible Host Diversity and Regional Potential Distribution of an Oriental Parasitic Plant (Taxillus yadoriki). Forests, 15(5), 799. https://doi.org/10.3390/f15050799