
The Pulses of Soil CO2 Emission in Response to Rainfall Events in Central Siberia: Revisiting the Overall Frost-Free Season CO2 Flux

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Measurements
2.3. Data Analysis
3. Results
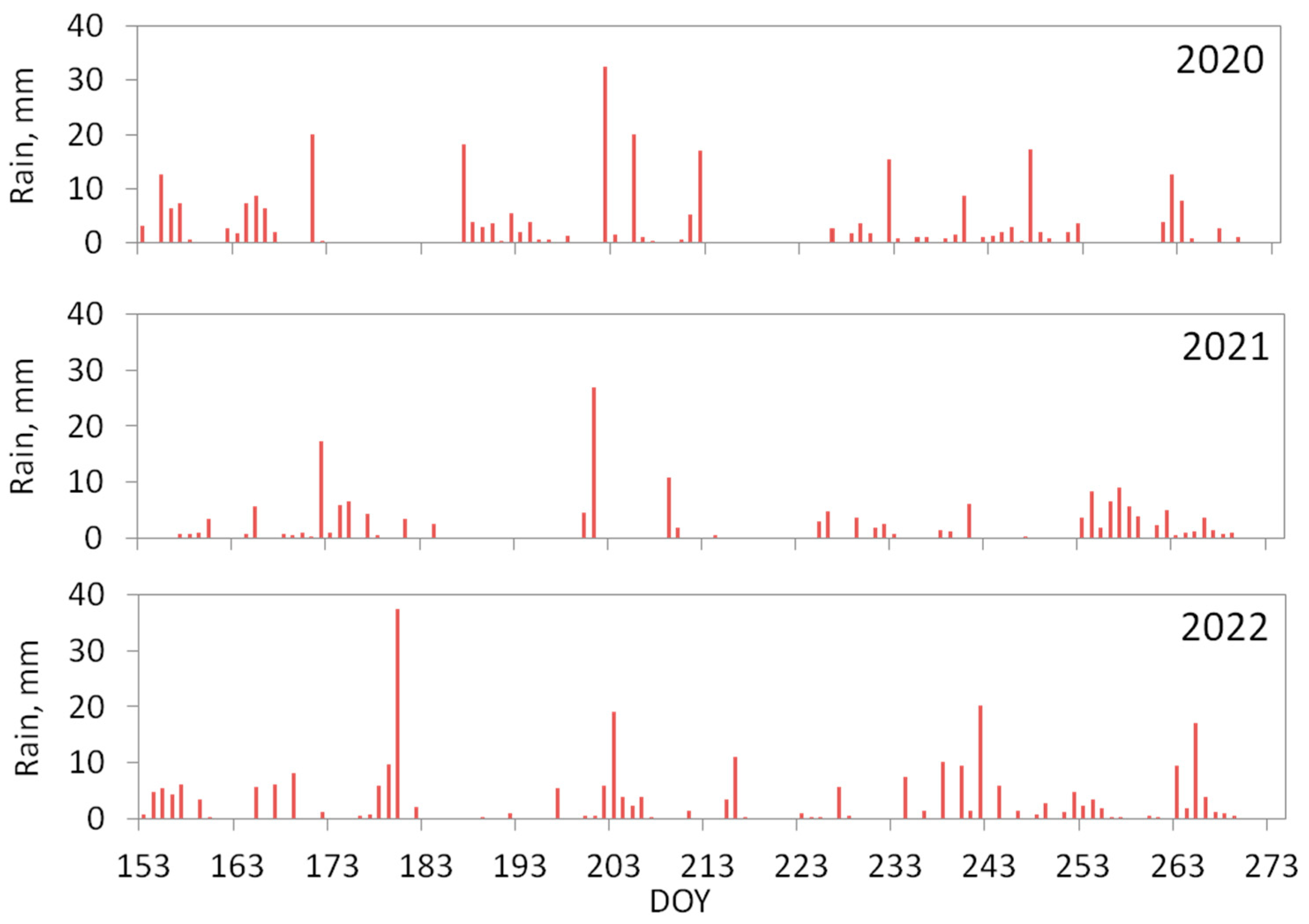
3.1. Precipitation Conditions of the Measurement Seasons
3.2. Seasonal Net Soil Exchange (NSE) and Soil Respiration (SR) Dynamics
3.3. NSE and SR Relationships with Soil Temperature and Soil Moisture
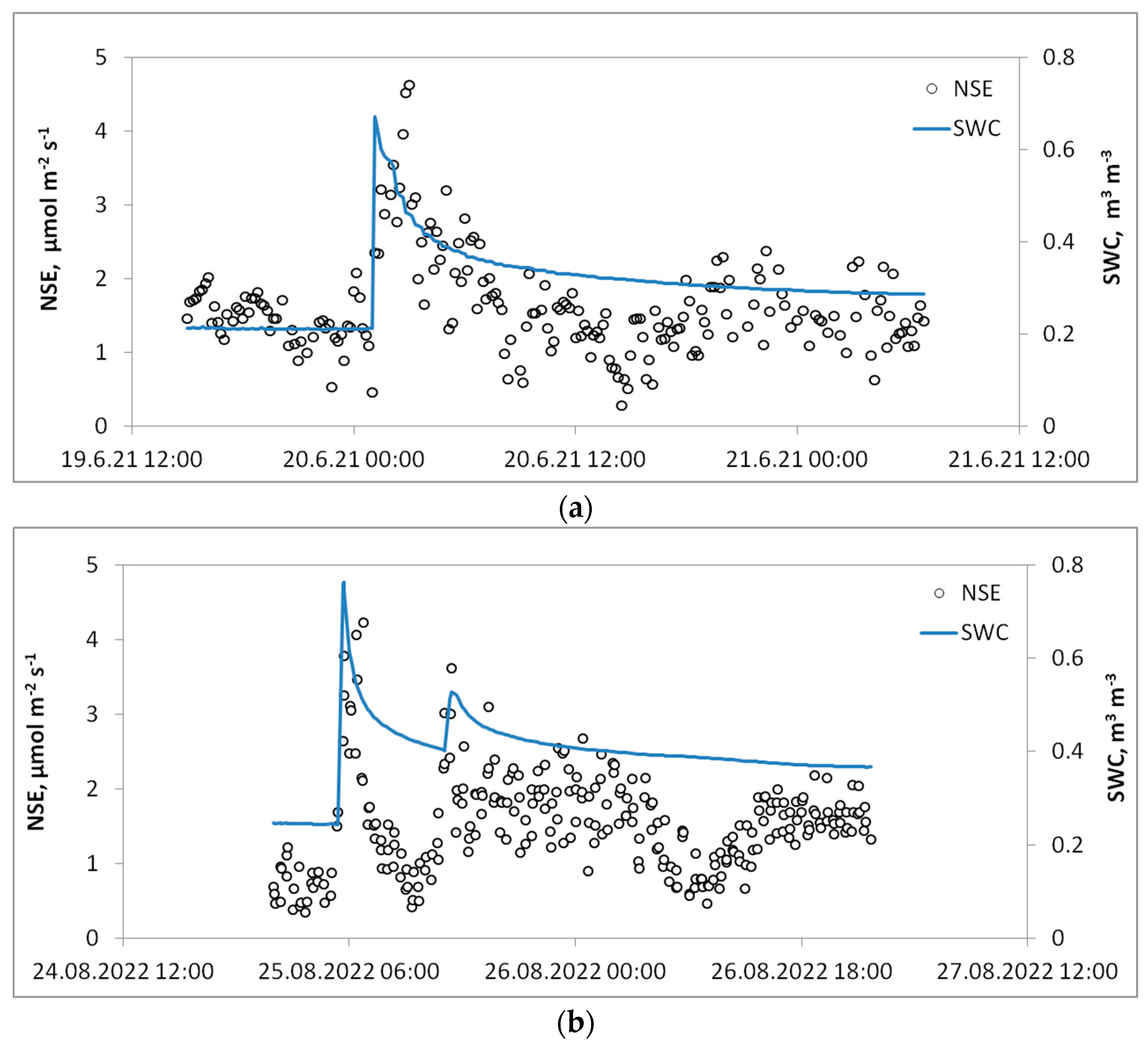
3.4. Rain-Induced CO2 Pulses
- -
- The first type was characterized by the descending behavior of CO2 efflux after a pulse (Figure 5a), i.e., after a heavy rain one observes a sharp increase in the CO2 flux rates, and then after the peak point it starts to decline;
- -
- The second type, the most common, is represented by multiple pulses following several consecutive rainfall events (Figure 5b). The first pulse is the strongest and the following pulses are usually less intense;
- -
- The third type of pulse has more complex behavior (Figure 5c). This type of pulse was observed in the 2020 season when after heavy rains, the sharp CO2 pulse lasted for a long period of time (up to 33 h), followed by a sharp decline of NSE rates. These pulses have the largest magnitude of NSE flux rates reaching 35–39 µmol m−2 s−1.
4. Discussion
4.1. Seasonal Dynamic of Soil CO2 Emission (SR) and Net Soil Exchange (NSE)
4.2. CO2 Pulses in NSE Seasonal Dynamics
4.3. The Role of CO2 Pulses in the Overall Frost-Free Season CO2 Flux
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 33, 988–993. [Google Scholar] [CrossRef]
- Harel, A.; Sylvain, J.-D.; Drolet, G.; Thiffault, E.; Thiffault, N.; Tremblay, S. Fine scale assessment of seasonal, intra-seasonal and spatial dynamics of soil CO2 effluxes over a balsam fir-dominated perhumid boreal landscape. Agric. For. Meteorol. 2023, 335, 109469. [Google Scholar] [CrossRef]
- Mukhortova, L.; Schepaschenko, D.; Shvidenko, A.; McCallum, I.; Kraxner, F. Soil contribution to carbon budget of Russian forests. Agric. For. Meteorol. 2015, 200, 97–108. [Google Scholar] [CrossRef]
- Abbott, B.W.; Jones, J.B. Permafrost collapse alters soil carbon stocks, respiration, CH4, and N2O in upland tundra. Glob. Chang. Biol. 2015, 21, 4570–4587. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Bailey, V.L.; Chen, M.; Gough, C.M.; Vargas, R. Globally rising soil heterotrophic respiration over recent decades. Nature 2018, 560, 80–83. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; McGuire, A.D.; Schädel, C.; Grosse, G.; Harden, J.W.; Hayes, D.J.; Hugelius, G.; Koven, C.D.; Kuhry, P.; Lawrence, D.M.; et al. Climate change and the permafrost carbon feedback. Nature 2015, 520, 171–179. [Google Scholar] [CrossRef]
- Kurganova, I.N.; Kudeyarov, V.N. Is a significant positive imbalance in the carbon cycle (sink) possible in Russia? Environ. Dyn. Glob. Clim. Chang. 2015, 6, 32–35. [Google Scholar] [CrossRef][Green Version]
- Darrouzet-Nardi, A.; Reed, S.C.; Grote, E.E.; Belnap, J. Observations of net soil exchange of CO2 in a dryland show experimental warming increases carbon losses in biocrust soils. Biogeochemistry 2015, 126, 363–378. [Google Scholar] [CrossRef]
- Fernandez, D.; Neff, J.; Belnap, J.; Reynolds, R. Soil respiration in the cold desert environment of the Colorado Plateau (USA): Abiotic regulators and thresholds. Biogeochemistry 2006, 78, 247–265. [Google Scholar] [CrossRef]
- Grote, E.E.; Belnap, J.; Housman, D.C.; Sparks, J.P. Carbon exchange in biological soil crust communities under differential temperatures and soil water contents: Implications for global change. Glob. Chang. Biol. 2010, 16, 2763–2774. [Google Scholar] [CrossRef]
- Lee, X.; Wu, H.J.; Sigler, J.; Oishi, C.; Siccama, T. Rapid and transient response of soil respiration to rain. Glob. Chang. Biol. 2004, 10, 1017–1026. [Google Scholar] [CrossRef]
- Prudhomme, C.; Giuntoli, I.; Robinson, E.L.; Clark, D.B.; Arnell, N.W.; Dankers, R.; Fekete, B.M.; Franssen, W.; Gerten, D.; Gosling, S.N.; et al. Hydrological droughts in the 21st century, hotspots and uncertainties from a global multimodel ensemble experiment. Proc. Natl. Acad. Sci. USA 2014, 111, 3262–3267. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar] [CrossRef]
- Birch, H.F. The effect of soil drying on humus decomposition and nitrogen availability. Plant Soil 1958, 10, 9–31. [Google Scholar] [CrossRef]
- Jiang, Z.; Yang, S.; Ding, J.; Sun, X.; Chen, X.; Liu, X.; Xu, J. Modeling climate change effects on rice yield and soil carbon under variable water and nutrient management. Sustainability 2021, 13, 568. [Google Scholar] [CrossRef]
- Patrick, C.J.; McGarvey, D.J.; Larson, J.H.; Cross, W.F.; Allen, D.C.; Benke, A.C.; Brey, T.; Huryn, A.D.; Jones, J.; Murphy, C.A.; et al. Precipitation and temperature drive continental-scale patterns in stream invertebrate production. Sci. Adv. 2019, 5, eaav2348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yu, Z.; Lin, J.; Zhu, B. Responses of soil carbon decomposition to drying-rewetting cycles: A meta-analysis. Geoderma 2020, 361, 114069. [Google Scholar] [CrossRef]
- Denef, K.; Six, J.; Bossuyt, H.; Frey, S.D.; Elliott, E.T.; Merckx, R.; Paustian, K. Influence of dry-wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial activity dynamics. Soil Biol. Biochem. 2001, 33, 1599–1611. [Google Scholar] [CrossRef]
- Bottner, P. Response of microbial biomass to alternate moist and dry conditions in a soil incubated with 14C- and 15N-labelled plant material. Soil Biol. Biochem. 1985, 17, 329–337. [Google Scholar] [CrossRef]
- Griffiths, E.; Birch, H.F. Microbiological changes in freshly moistened soil. Nature 1961, 189, 424. [Google Scholar] [CrossRef]
- Orchard, V.A.; Cook, F.J. Relationship between soil respiration and soil moisture. Soil Biol. Biochem. 1983, 15, 447–453. [Google Scholar] [CrossRef]
- Schimel, J.P. Life in Dry Soils: Effects of drought on soil microbial communities and processes. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 409–432. [Google Scholar] [CrossRef]
- Sun, S.; Lei, H.; Chang, S.X. Drought differentially affects autotrophic and heterotrophic soil respiration rates and their temperature sensitivity. Biol. Fertil. Soils 2019, 55, 275–283. [Google Scholar] [CrossRef]
- Yang, W.; Cao, J.; Wu, Y.; Kong, F.; Li, L. Review on plant terpenoid emissions worldwide and in China. Sci. Total Environ. 2021, 787, 147454. [Google Scholar] [CrossRef] [PubMed]
- Kieft, T.L.; Soroker, E.; Firestone, M.K. Microbial biomass response to a rapid increase in water potential when dry soil is wetted. Soil Biol. Biochem. 1987, 19, 119–126. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil. Soil Sci. Soc. Am. J. 2003, 67, 798–805. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Niklaus, P.A.; Hungate, B.A. A meta-analysis of responses of soil biota to global change. Oecologia 2011, 165, 553–565. [Google Scholar] [CrossRef]
- Sun, D.; Li, K.; Bi, Q.; Zhu, J.; Zhang, Q.; Jin, C.; Lu, L.; Lin, X. Effects of organic amendment on soil aggregation and microbial community composition during drying-rewetting alternation. Sci. Total Environ. 2017, 574, 735–743. [Google Scholar] [CrossRef]
- Kim, D.G.; Vargas, R.; Bond-Lamberty, B.; Turetsky, M.R. Effects of soil rewetting and thawing on soil gas fluxes: A review of current literature and suggestions for future research. Biogeosciences 2012, 9, 2459–2483. [Google Scholar] [CrossRef]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Inglima, I.; Alberti, G.; Bertolini, T.; Vaccari, F.P.; Gioli, B.; Miglietta, F.; Peressotti, A. Precipitation pulses enhance respiration of Mediterranean ecosystems: The balance between organic and inorganic components of increased soil CO2 efflux. Glob. Chang. Biol. 2009, 15, 1289–1301. [Google Scholar] [CrossRef]
- Barnard, R.L.; Blazewicz, S.J.; Mary, K. Firestone rewetting of soil: Revisiting the origin of soil CO2 emissions. Soil Biol. Biochem. 2020, 147, 107819. [Google Scholar] [CrossRef]
- Xu, L.; Baldocchi, D.D.; Tang, J. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Glob. Biogeochem. Cycles 2004, 18, GB4002. [Google Scholar] [CrossRef]
- Tiemann, L.K.; Billings, S.A. Changes in variability of soil moisture alter microbial community C and N resource use. Soil Biol. Biochem. 2011, 43, 1837–1847. [Google Scholar] [CrossRef]
- Dai, A.; Zhao, T.; Chen, J. Climate change and drought: A precipitation and evaporation perspective. Curr. Clim. Chang. Rep. 2018, 4, 301–312. [Google Scholar] [CrossRef]
- Fan, Z.; Neff, J.C.; Hanan, N.P. Modeling pulsed soil respiration in an African savanna ecosystem. Agric. For. Meteorol. 2015, 200, 282–292. [Google Scholar] [CrossRef]
- Sang, J.; Lakshani, M.M.T.; Deepagoda, C.T.K.K.; Shen, Y.; Li, Y. Drying and rewetting cycles increased soil carbon dioxide rather than nitrous oxide emissions: A meta-analysis. J. Environ. Manage. 2022, 324, 116391. [Google Scholar] [CrossRef]
- Klimchenko, A.V.; Verkhovets, S.V.; Slinkina, O.A.; Koshurnikova, N.N. Reserves of coarse woody debris in middle-taiga ecosystems of Yenisei Siberia. Geogr. Environ. Resour. 2011, 2, 91–97. (In Russian) [Google Scholar]
- Dymov, A.A.; Startsev, V.V.; Yakovleva, E.V.; Dubrovskiy, Y.A.; Milanovsky, E.Y.; Severgina, D.A.; Panov, A.V.; Prokushkin, A.S. Fire-induced alterations of soil properties in albic podzols developed under pine forests (middle taiga, Krasnoyarsky Kray). Fire 2023, 6, 67. [Google Scholar] [CrossRef]
- Polosukhina, D.A.; Prokushkin, A.S. Comparative characteristics of reserves and isotopic composition of soil organic matter in forest biogeocenoses in the footprint zone of the ZOTTO tall tower. In Proceedings of the Lomonosov Readings in Altai: Fundamental Problems of Science and Education, Barnaul, Russia, 14–17 November 2017. (In Russian). [Google Scholar]
- Makhnykina, A.V. The influence of temperature and humidity on CO2 emission fluxes from the soil surface in pine forests of the middle taiga subzone of Central Siberia. Diss. Cand. Biol. Sci. 2020. (In Russian) [Google Scholar]
- Makhnykina, A.V.; Prokushkin, A.S.; Vaganov, E.A.; Verkhovets, S.V.; Rubtsov, A.V. Dynamics of the CO2 fluxes from the soil surface in pine forests in Central Siberia. J. Sib. Fed. Univ. Biol. 2016, 3, 338–357. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.r-project.org (accessed on 15 May 2023).
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. R Package Version 0.8.4.2020. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 10 May 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; Mc-Gowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Grolemund, G.; Wickham, H. Dates and times made easy with lubridate. J. Open Source Softw. 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Wickham, H.; Çetinkaya-Rundel, M.; Grolemund, G. R for Data Science, 2nd ed.; O’Reilly Media: Sebastopol, CA, USA, 2016; Available online: https://r4ds.hadley.nz (accessed on 6 April 2023).
- Wickham, H. Mastering Shiny; O’Reilly Media: Sebastopol, CA, USA, 2016; Available online: https://github.com/rstudio/shiny. (accessed on 16 April 2023).
- Karelin, D.V.; Pochikalov, A.V.; Zamolodchikov, D.G.; Gitarskii, M.L. Factors of spatiotemporal variability of CO2 fluxes from soils of southern taiga spruce forests of Valdai. Contemp. Probl. Ecol. 2014, 7, 743–751. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bergeron, Y.; Chen, H.Y.H. The effect of boreal forest composition on soil respiration is mediated through variations in soil temperature and C quality. Soil Biol. Biochem. 2012, 53, 18–27. [Google Scholar] [CrossRef]
- Raich, J.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B Chem. Phys. Meteorol. 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thompson, A. Temperature-associated increases in the global soil respiration record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Roby, M.C.; Scott, R.L.; Biederman, J.A.; Smith, W.K.; Moore, D.J.P. Response of soil carbon dioxide efflux to temporal repackaging of rainfall into fewer, larger events in a semiarid grassland. Front. Environ. Sci. 2022, 10, 940943. [Google Scholar] [CrossRef]
- Savage, K.; Davidson, E.A.; Richardson, A.D.; Hollinger, D.Y. Three scales of temporal resolution from automated soil respi-ration measurements. Agric. For. Meteorol. 2009, 149, 2012–2021. [Google Scholar] [CrossRef]
- Sun, J.; Yu, K.; Chen, N.; Munson, S.M.; Li, X.; Jia, R. Biocrusts modulate carbon losses under warming across global drylands: A bayesian meta-analysis. Soil Biol. Biochem. 2013, 188, 109214. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Subke, J.-A.; Reichstein, M.; Tenhunen, J.D. Explaining temporal variation in soil CO2 efflux in a mature spruce forest in Southern Germany. Soil Biol. Biochem. 2003, 35, 1467–1483. [Google Scholar] [CrossRef]
- Kolari, P.; Kulmala, L.; Pumpanen, J.; Launiainen, S.; Ilvesniemi, H.; Hari, P.; Nikinmaa, E. CO2 exchange and component CO2 fluxes of a boreal Scots pine forest. Boreal Environ. Res. 2009, 14, 761–783. [Google Scholar]
- Niinistö, S.M.; Kellomäki, S.; Silvola, J. Seasonality in a boreal forest ecosystem affects the use of soil temperature and moisture as predictors of soil CO2 efflux. Biogeosciences 2011, 8, 3169–3186. [Google Scholar] [CrossRef]
- Shibistova, O.; Lloyd, J.; Evgrafova, S.; Savushkina, N.; Zrazhevskaya, G.; Arneth, A.; Knohl, A.; Kolle, O.; Schulze, E.-D. Seasonal and spatial variability in soil CO2 efflux rates for a Central Siberian Pinus sylvestris forest. Tellus B 2002, 54, 552–567. [Google Scholar] [CrossRef]
- Makhnykina, A.; Panov, A.; Prokushkin, A. The impact of wildfires on soil CO2 emission in middle taiga forests in Central Siberia. Land 2023, 12, 1544. [Google Scholar] [CrossRef]
- Masyagina, O.V.; Menyailo, O.V.; Prokushkin, A.S.; Matvienko, A.I.; Makhnykina, A.V.; Evgrafova, S.Y.; Mori, S.; Koike, T.; Prokushkin, S.G. Soil respiration in larch and pine ecosystems of the Krasnoyarsk region (Russian Federation): A latitudinal comparative study. Arab. J. Geosci. 2020, 13, 954. [Google Scholar] [CrossRef]
- Makhnykina, A.V.; Prokushkin, A.S.; Menyailo, O.V.; Verkhovets, S.V.; Tychkov, I.I.; Urban, A.V.; Rubtsov, A.V.; Koshurnikova, N.N.; Vaganov, E.A. The impact of climatic factors on CO2 emissions from soils of middle-taiga forests in Central Siberia: Emission as a function of soil temperature and moisture. Russ. J. Ecol. 2020, 51, 46–56. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Braun, M.; Tataurov, V.A. Seasonal and daily dynamics of the CO2 emission from soils of Pinus koraiensis forests in the south of the Sikhote-Alin Range. Eurasian Soil Sci. 2018, 51, 290–295. [Google Scholar] [CrossRef]
- Kotani, A.; Saito, A.; Kononov, A.V.; Petrov, R.E.; Maximov, T.C.; Iijima, Y.; Ohta, T. Impact of unusually wet permafrost soil on understory vegetation and CO2 exchange in a larch forest in eastern Siberia. Agric. For. Meteorol. 2019, 265, 295–309. [Google Scholar] [CrossRef]
- Maximov, T.C.; Maksimov, A.; Kononov, A.; Kotani, A.; Dolman, J. Carbon cycles in forest. In Water-Carbon Dynamics in Eastern Siberia; Ohta, T., Hiyama, T., Iijima, Y., Kotani, A., Maximov, T.C., Eds.; Springer: Tokyo, Japan, 2019; pp. 69–100. [Google Scholar] [CrossRef]
- Kim, Y. Effect of thaw depth on fluxes of CO2 and CH4 in manipulated Arctic coastal tundra of Barrow. Sci. Total Environ. 2015, 505, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Osipov, A.F. Carbon dioxide emission from the soil surface in mature bilberry pine forest in middle taiga (Republic of Komi). Eurasian Soil Sci. 2016, 49, 926–933. [Google Scholar] [CrossRef]
- Makhnykina, A.V.; Tychkov, I.I.; Prokushkin, A.S.; Pyzhev, A.I.; Vaganov, E.A. Factors of soil CO2 emission in boreal forests: Evidence from Central Siberia. iForest 2023, 16, 86–94. [Google Scholar] [CrossRef]
- Henskens, F.L.; Allan Green, T.G.; Wilkins, A. Cyanolichens can have both cyanobacteria and green algae in a common layer as major contributors to photosynthesis. Ann. Bot. 2012, 110, 555–563. [Google Scholar] [CrossRef]
- Savage, K.; Davidson, E.A.; Tang, J. Diel patterns of autotrophic and heterotrophic respiration among phenological stages. Glob. Chang. Biol. 2013, 19, 1151–1159. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Zhang, C.; Wu, X.; Du, E.; Wu, H.; Yang, X.; Wang, P.; Bai, Y.; Wu, Y.; et al. Responses of soil respiration to rainfall addition in a desert ecosystem: Linking physiological activities and rainfall pattern. Sci. Total Environ. 2019, 650, 3007–3016. [Google Scholar] [CrossRef]
- Chapin, F.S.; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science & Business Media: New York, NY, USA, 2011; p. 546. [Google Scholar] [CrossRef]
- Darrouzet-Nardi, A.; Reed, S.C.; Grote, E.E.; Belnap, J. Patterns of longer-term climate change effects on CO2 efflux from biocrusted soils differ from those observed in the short term. Biogeosciences 2018, 15, 4561–4573. [Google Scholar] [CrossRef]
- Dyukarev, E.A.; Golovatskaya, E.A.; Lapshina, E.D.; Filippova, N.V.; Zarov, E.A.; Filippov, I.V. Modeling of the net ecosystem exchange, gross primary production, and ecosystem respiration for peatland ecosystems of western Siberia. IOP Conf. Ser. Earth Environ. Sci. 2018, 211, 012028. [Google Scholar] [CrossRef]
- Schwinning, S.; Sala, O.E. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef]
- Scott, R.L.; Serrano-Ortiz, P.; Domingo, F.; Hamerlynck, E.P.; Kowalski, A.S. Commonalities of carbon dioxide exchange in semiarid regions with monsoon and Mediterranean climates. J. Arid Environ. 2012, 84, 71–79. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Scott, R.L.; Huxman, T.E. Whole ecosystem metabolic pulses following precipitation events. Funct. Ecol. 2008, 22, 924–930. [Google Scholar] [CrossRef]
- Sponseller, R.A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Glob. Chang. Biol. 2007, 13, 426–436. [Google Scholar] [CrossRef]
- Barron-Gafford, G.A.; Scott, R.L.; Jenerette, G.D.; Huxman, T.E. The relative controls of temperature, soil moisture, and plant functional group on soil CO2 efflux at diel, seasonal, and annual scales. J. Geophys. Res. 2011, 116, G01023. [Google Scholar] [CrossRef]
- Ni, X.; Liao, S.; Wu, F.; Groffman, P.M. Short-term precipitation pulses stimulate soil CO2 emission but do not alter CH4 and N2O fluxes in a northern hardwood forest. Soil Biol. Biochem. 2019, 130, 8–11. [Google Scholar] [CrossRef]
- Xiang, S.-R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Stepanova, O.G.; Trunova, V.A.; Osipov, E.Y.; Kononov, E.E.; Vorobyeva, S.S.; Parkhomchuk, E.V.; Kalinkin, P.N.; Vorobyeva, E.E.; Vershinin, K.E.; Rastigeev, S.A.; et al. Glacier dynamics in the southern part of East Siberia (Russia) from the final part of the LGM to the present based on from biogeochemical proxies from bottom sediments of proglacial lakes. Quat. Int. 2019, 524, 4–12. [Google Scholar] [CrossRef]
- Kharanzhevskaya, Y.A.; Voistinova, E.S.; Sinyutkina, A.A. Spatial and temporal variations in mire surface water chemistry as a function of geology, atmospheric circulation and zonal features in the south-eastern part of western Siberia. Sci. Total Environ. 2020, 733, 139343. [Google Scholar] [CrossRef] [PubMed]
- Bormann, H.; Klaassen, K. Seasonal and land use dependent variability of soil hydraulic and soil hydrological properties of two northern German soils. Geoderma 2008, 145, 295–302. [Google Scholar] [CrossRef]
- Zavalishin, A.A. Soils of the Forest Zone, Their Formation and Properties; Publishing House of the USSR Academy of Sciences: Leningrag, Russia, 1939; p. 111. [Google Scholar]
- Sanborn, P.; Lamontagne, L.; Hendershot, W. Podzolic soils of Canada: Genesis, distribution, and classification. Can. J. Soil Sci. 2011, 91, 843880. [Google Scholar] [CrossRef]
- Trefilova, O.V. Intensity of heterotrophic respiration in pine forest of middle taiga: Comparative analysis of assessment methods. Conifers Boreal Zone 2007, 24, 467–473. (In Russian) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Mean Season NSE, µmol m−2 s−1 ± SD | Pulse NSE (Rain > 5 mm), µmol m−2 s−1 ± SD | Sum Mean NSE Per Season, g C m−2 | Sum Pulse NSE Per Season, g C m−2 | Pulse NSE Input (Rain > 5 mm) Per Season, % |
|---|---|---|---|---|---|
| 2020 | 3.7 ± 5.1 | 18.0 ± 8.2 | 394.9 | 154.5 | 39% |
| 2021 | 1.6 ± 0.7 | 1.8 ± 1.3 | 156.8 | 3.8 | 2% |
| 2022 * | 1.4 ± 1.2 | 4.5 ± 2.9 | 34.8 | 5.4 | 15% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makhnykina, A.V.; Vaganov, E.A.; Panov, A.V.; Koshurnikova, N.N.; Prokushkin, A.S. The Pulses of Soil CO2 Emission in Response to Rainfall Events in Central Siberia: Revisiting the Overall Frost-Free Season CO2 Flux. Forests 2024, 15, 355. https://doi.org/10.3390/f15020355
Makhnykina AV, Vaganov EA, Panov AV, Koshurnikova NN, Prokushkin AS. The Pulses of Soil CO2 Emission in Response to Rainfall Events in Central Siberia: Revisiting the Overall Frost-Free Season CO2 Flux. Forests. 2024; 15(2):355. https://doi.org/10.3390/f15020355
Chicago/Turabian StyleMakhnykina, Anastasia V., Eugene A. Vaganov, Alexey V. Panov, Nataly N. Koshurnikova, and Anatoly S. Prokushkin. 2024. "The Pulses of Soil CO2 Emission in Response to Rainfall Events in Central Siberia: Revisiting the Overall Frost-Free Season CO2 Flux" Forests 15, no. 2: 355. https://doi.org/10.3390/f15020355
APA StyleMakhnykina, A. V., Vaganov, E. A., Panov, A. V., Koshurnikova, N. N., & Prokushkin, A. S. (2024). The Pulses of Soil CO2 Emission in Response to Rainfall Events in Central Siberia: Revisiting the Overall Frost-Free Season CO2 Flux. Forests, 15(2), 355. https://doi.org/10.3390/f15020355