
Predicting the Potential Habitat Distribution of Relict Plant Davidia involucrata in China Based on the MaxEnt Model

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
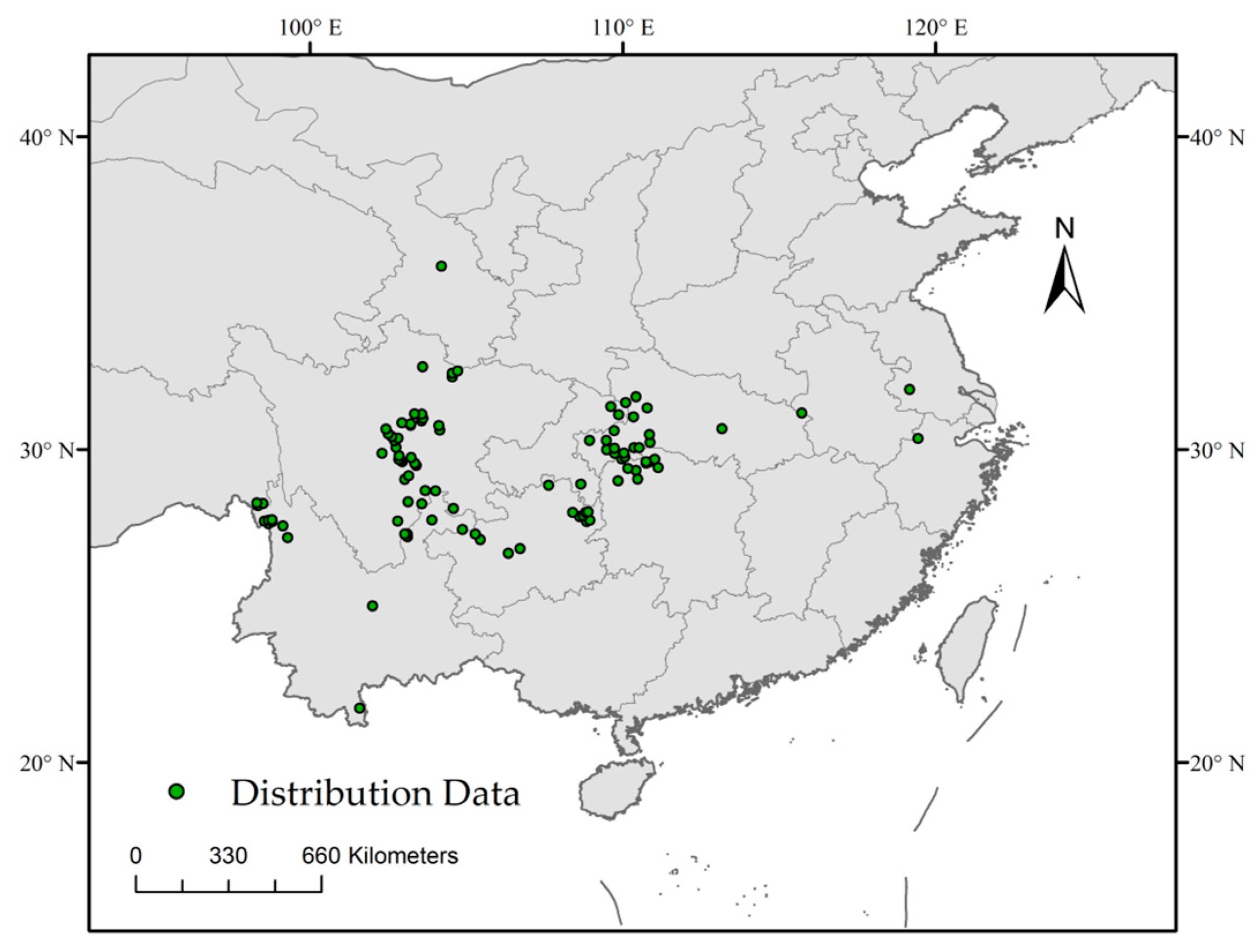
2.1. Species Distribution Data
2.2. Environmental Data
2.3. MaxEnt Modeling Process
2.4. The Classification and Spatial Pattern Changes of Suitable Areas of D. involucrata
3. Results
3.1. MaxEnt Model Prediction and Accuracy Evaluation for the Suitable Habitat of D. involucrata
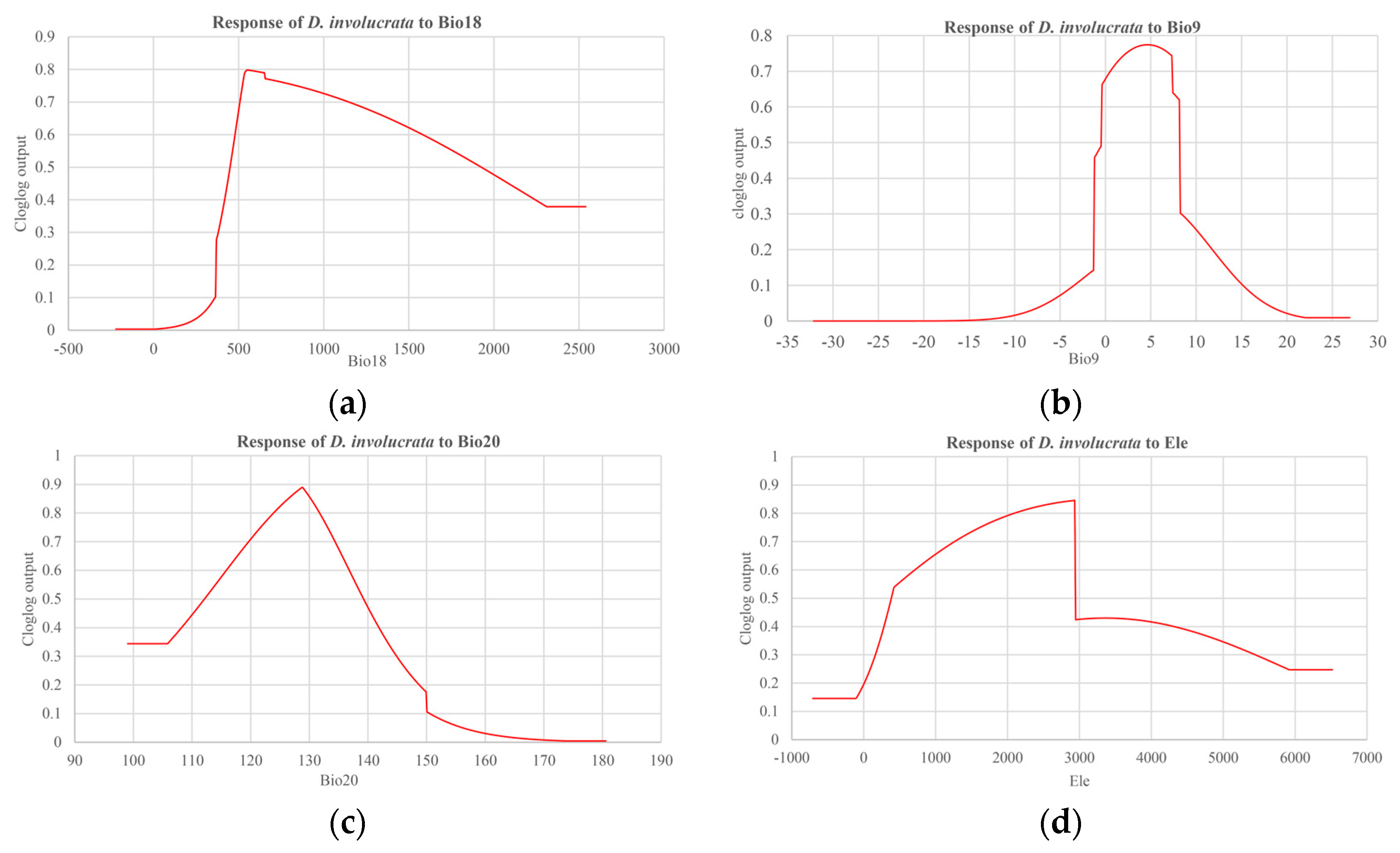
3.2. Contribution of Each Variable to the MaxEnt Prediction Model
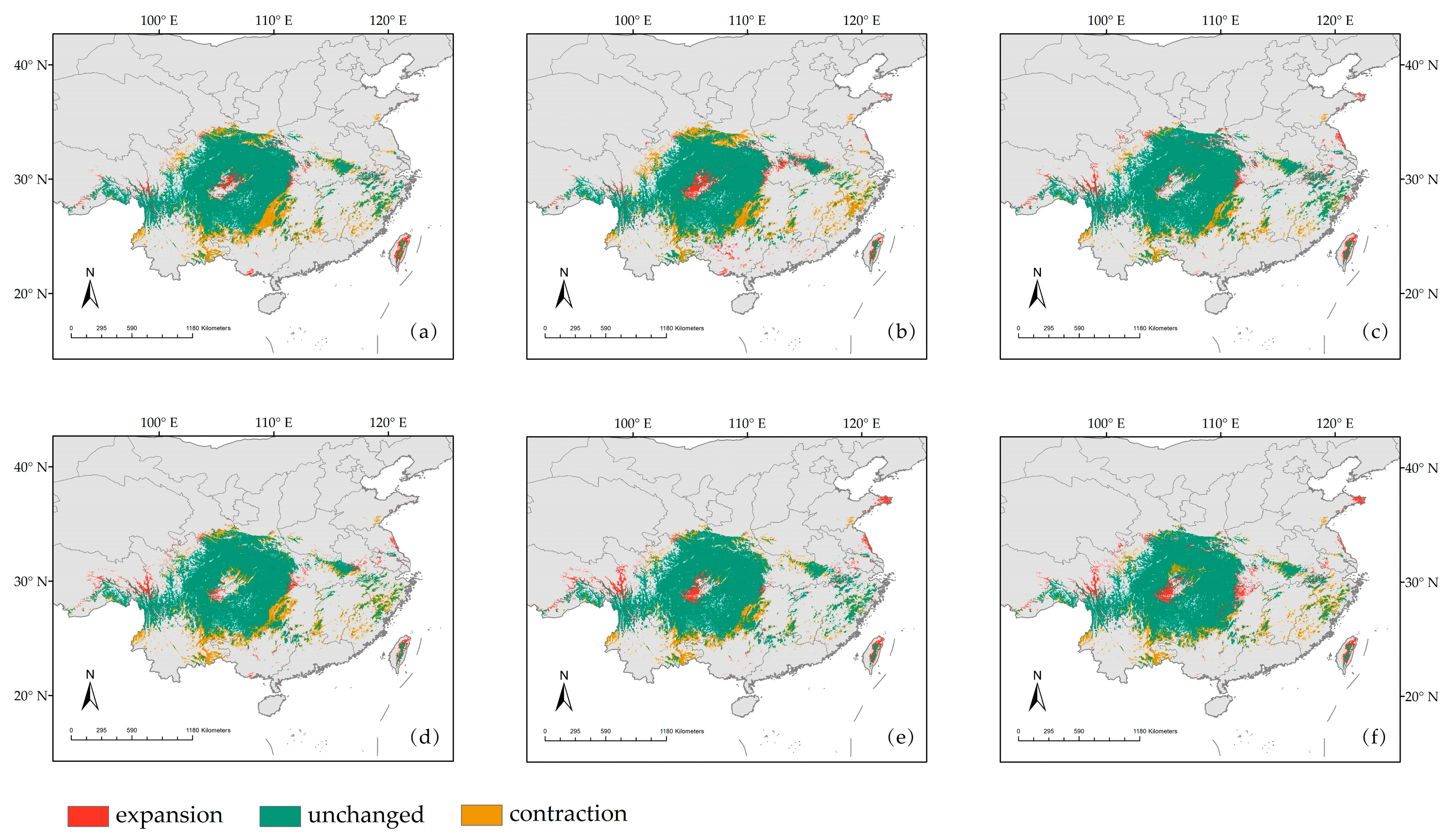
3.3. Potential Habitat Changes of D. involucrata under Current and Future Climate Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ginbo, T. Heterogeneous impacts of climate change on crop yields across altitudes in Ethiopia. Clim. Chang. 2022, 170, 12. [Google Scholar] [CrossRef]
- Schnitter, R.; Berry, P. The Climate Change, Food Security and Human Health Nexus in Canada: A Framework to Protect Population Health. Int. J. Environ. Res. Public Health 2019, 16, 2531. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.J.A.; Ehrlich, P.R.; Beattie, A.; Ceballos, G.; Crist, E.; Diamond, J.; Dirzo, R.; Ehrlich, A.H.; Harte, J.; Harte, M.E.; et al. Underestimating the challenges of avoiding a ghastly future. Front. Conserv. Sci. 2021, 1, 615419. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The human footprint and the last of the wild: The human footprint is a global map of human influence on the land surface, which suggests that human beings are stewards of nature, whether we like it or not. Bioscience 2002, 52, 891–904. [Google Scholar] [CrossRef]
- Wilson, R.J.; Gutiérrez, D.; Gutiérrez, J.; Martínez, D.; Agudo, R.; Monserrat, V.J. Changes to elevational limits and extent of species ranges associated with climate change. Ecol. Lett. 2005, 8, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.R.; Beißner, S.; Burga, C.A. Trends in the upward shift of alpine plants. J. Veg. Sci. 2005, 16, 541–548. [Google Scholar] [CrossRef]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef]
- Thomas, J.A.; Telfer, M.G.; Roy, D.B.; Preston, C.D.; Greenwood, J.J.D.; Asher, J.; Fox, R.; Clarke, R.T.; Lawton, J.H. Comparative losses of British butterflies, birds and plants and the global extinction crisis. Science 2004, 303, 1879–1881. [Google Scholar] [CrossRef]
- Zhang, G.; Zeng, G.; Yang, X.; Jiang, Z. Future Changes in Extreme High Temperature over China at 1.5 °C–5 °C Global Warming Based on CMIP6 Simulations. Adv. Atmos. Sci. 2021, 38, 253–267. [Google Scholar] [CrossRef]
- Moraitis, M.L.; Valavanis, V.D.; Karakassis, I. Modelling the effects of climate change on the distribution of benthic indicator species in the Eastern Mediterranean Sea. Sci. Total Environ. 2019, 667, 16–24. [Google Scholar] [CrossRef]
- Wang, B.; Deveson, E.D.; Waters, C.; Spessa, A.; Lawton, D.; Feng, P.Y.; Liu, D.L. Future climate change likely to reduce the Australian plague locust (Chortoicetes terminifera) seasonal outbreaks. Sci. Total Environ. 2019, 668, 947–957. [Google Scholar] [CrossRef]
- Wilson, K.L.; Skinner, M.A.; Lotze, H.K. Projected 21st-century distribution of canopy-forming seaweeds in the Northwest Atlantic with climate change. Divers. Distrib. 2019, 25, 582–602. [Google Scholar] [CrossRef]
- Liu, M.; Wen, J.H.; Xu, W.J.; Chen, Y.M.; Ma, Z.L. The Impact of Nitrogen Application on Leaf and Root Functional Traits of Davidia involucrata Saplings. Forests 2023, 14, 1668. [Google Scholar] [CrossRef]
- Tang, C.; Dong, Y.F.; Herrando-Moraira, S.; Matsui, T.; Ohashi, H.; He, L.Y.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.S.; et al. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.G.; Jin, J.M. China Plant Red Data Book: Rare and Endangered Plants; Science Press: Beijing, China, 1992; Volume 1, pp. 350–351. [Google Scholar]
- Li, G.L.; Cao, C.X.; Yang, H.; Wang, J.H.; Wei, W.; Zhu, D.H.; Gao, P. Molecular cloning and potential role of DiSOC1s in flowering regulation in Davidia involucrata Baill. Plant Physiol. Biochem. 2020, 157, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Deng, H.Z.; Chao, Z.; Liu, C.M.; Zhao, H.W. Medicinal plant resources of the Chinese endemic genera of seed plants. China J. Chin. Mater. Med. 2004, 29, 123–129. [Google Scholar] [CrossRef]
- Xiang, G.Q.; Lu, F.S. Study on chemical components of Davidia involucrata Baill. native to China. J. Integr. Plant Biol. 1989, 31, 540–543. [Google Scholar]
- Fan, X.M. Extraction, Separation and Activity Study of Tannin of Patrinia villosa Juss; Zhejiang University: Hangzhou, China, 2014. [Google Scholar]
- Dai, Y. Effects of Plant Sterol on Plasma Lipids and Liver Lipids in High-Fat Diet Rat; Fourth Military Medical University: Xi’an, China, 2010. [Google Scholar]
- Zhou, M.; Zhang, R.H.; Wang, M.; Xu, G.B.; Liao, S.G. Prodrugs of triterpenoids and their derivatives. Eur. J. Med. Chem. 2017, 131, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.T.; Xu, G.B.; Wang, X.P. Literature Review of Research on Davidia involucrata Baill. Nonwood For. Res. 2006, 24, 92–94, 98. [Google Scholar] [CrossRef]
- Wu, Y.; Rong, R.; Chen, F.; Xu, Y. Effect of light quality on morphogenesis and photosynthetic characteristics of Davidia involucrata seedings. J. Sichuan Univ. Nat. Sci. Ed. 2020, 57, 804–810. [Google Scholar] [CrossRef]
- Xiong, Y.L.; Cao, F.X.; Liu, Z.M.; Dong, X.J.; Li, M. Cloning and expression analysis of Ces A genes involved in seed abortion in dove tree (Davidia involucrate). J. Plant Physiol. 2016, 52, 1481–1490. [Google Scholar] [CrossRef]
- Lei, N.; Peng, S.; Niu, B.; Chen, J.; Zhou, J.; Tang, L.; Xu, Y.; Wang, S.; Chen, F. Molecular cloning and characterization of a novel microsomal oleate desaturase gene DiFAD2 from Davidia involucrata Baill. Biol. Plant. 2010, 54, 41–46. [Google Scholar] [CrossRef]
- Wei, X.R.; Ding, H.F.; Fan, Y.L.; Wu, X.M.; Liu, X.D.; Niu, J.; Cao, F.X.; Li, M. Overexpression of a laccase gene, DiLAC17, from Davidia involucrata causes severe seed abortion in Arabidopsis. Plant Physiol. Biochem. 2023, 202, 107956. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Jin, X.L.; Shen, S.Y.; Zhang, R.Q. Population quantitative characteristics and dynamics of rare and endangered plant Davidia involucrata in Hunan Province. Acta Ecol. Sin. 2012, 32, 7738–7746+5. [Google Scholar] [CrossRef]
- Su, R.J.; Wu, Q.G.; Yang, Y.L.; Hu, T.X. Relationship between Diameter at Breast Height and Tree Age in Populations of a Rare and Endangered Plant, Davidia involucrata. Pol. J. Ecol. 2021, 69, 84–95. [Google Scholar] [CrossRef]
- Yang, S.X.; Zhang, J. The Study on Davidia involucrata Community in the Seven-Sisters Mountain Nature Reserve. J. Shandong For. Sci. Technol. 2015, 45, 69–71+65. [Google Scholar] [CrossRef]
- Chen, L.X.; He, M.X.; Wang, B.; Tan, J.B.; Huang, Y.H.; Ran, J.H. Analysis of suitable habitat distribution and its influence factors of Davidia involucrata in Liangshan Mountains based on Maxent model. J. Sichuan Univ. Nat. Sci. Ed. 2018, 55, 873–880. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Zhou, R.H.; Yu, F.Y.; Ye, X.; Wang, M.; Qi, J.Q.; Hao, J.F. Flora, structure, and dynamics of Davidia involucrata community in Daxiangling Nature Reserve. Chin. J. Ecol. 2020, 39, 1509–1517. [Google Scholar] [CrossRef]
- Stockwell, D.R.; Peterson, A. Effects of sample size on accuracy of species distribution models. Ecol. Model. 2002, 148, 1–13. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Hausser, J.; Perrin, D.C. Ecological-Niche Factor Analysis: How to compute habitat-suitability maps without absence data? Ecology 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Haase, C.G.; Yang, A.N.; McNyset, K.M.; Blackburn, J.K. GARPTools: R software for data preparation and model evaluation of GARP models. Ecography 2021, 44, 1790–1796. [Google Scholar] [CrossRef]
- Semwal, D.P.; Pandey, A.; Gore, P.G.; Ahlawat, S.P.; Yadav, S.K.; Kumar, A. Habitat prediction mapping using BioClim model for prioritizing germplasm collection and conservation of an aquatic cash crop ‘makhana’ (Euryale ferox Salisb.) in India. Genet. Resour. Crop. Evol. 2021, 68, 3445–3456. [Google Scholar] [CrossRef]
- Ma, R.M.; Ban, J.; Wang, Q.; Zhang, Y.Y.; Yang, Y.; Li, S.S.; Shi, W.J.; Zhou, Z.; Zang, J.W.; Li, T.T. Full-coverage 1km daily ambient PM2.5 and O3 concentration of China in 2005-2017 based on a multi-variable random forest model. Earth Syst. Sci. Data 2022, 14, 943–954. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen–Geiger climate classifcation maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Y.; Milne, R.I.; Fan, X.Y.; Xie, S.Y.; Lei, Z.; Zheng, H.L.; Fan, L.Q.; Chung, J.M.; Chuang, M.G.; Ma, T.; et al. Blow to the Northeast? Intraspecifc differentiation of Populus davidiana suggests a north-eastward skew of a phylogeographic break in East Asia. J. Biogeogr. 2020, 48, 187–201. [Google Scholar] [CrossRef]
- Anand, V.; Oinam, B.; Singh, I.H. Predicting the current and future potential spatial distribution of endangered Rucervus eldii eldii (Sangai) using Maxent model. Environ. Monit. Assess. 2021, 193, 147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, J.S.; Ren, G.; Zhao, K.X.; Wang, X.F. Global potential distribution prediction of Xanthium italicum based on Maxent model. Sci. Rep. 2021, 11, 16545. [Google Scholar] [CrossRef]
- Liu, J.M.; Xu, Y.Y.; Sun, C.W.; Wang, X.; Zheng, Y.L.; Shi, S.L.; Chen, Z.; He, Q.Y.; Weng, X.H.; Jia, L.M. Distinct ecological habits and habitat responses to future climate change in three east and southeast Asian Sapindus species. For. Ecol. Manag. 2022, 507, 119982. [Google Scholar] [CrossRef]
- Karuppaiah, V.; Maruthadurai, R.; Das, B.; Soumia, P.S.; Gadge, A.S.; Thangasamy, A.; Ramesh, S.V.; Shirsat, D.V.; Mahajan, V.; Krishna, H.; et al. Predicting the potential geographical distribution of onion thrips, Thrips tabaci in India based on climate change projections using MaxEnt. Sci. Rep. 2023, 13, 7934. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Berger, U.; Cao, M.; Zhang, Y.; He, J.; Pan, L.; Jiang, J. Conservation and Restoration of Mangroves in Response to Invasion of Spartina alterniflora Based on the MaxEnt Model: A Case Study in China. Forests 2023, 14, 1220. [Google Scholar] [CrossRef]
- Wang, X.F.; Duan, Y.X.; Jin, L.L.; Wang, C.Y.; Peng, M.C.; Li, Y.; Wang, X.H.; Ma, Y.F. Prediction of historical, present and future distribution of Quercus sect. Heterobalanus based on the optimized MaxEnt model in China. Acta Ecol. Sin. 2023, 43, 6590–6604. [Google Scholar]
- Wu, C.; Chen, D.; Shen, J.; Sun, X.; Zhang, S. Estimating the distribution and productivity characters of Larix kaempferi in response to climate change. J. Environ. Manag. 2021, 280, 111633. [Google Scholar] [CrossRef]
- Ma, Y.F.; Li, J.Q. Population structure of Davidia involucrata in Mt. Seven-sister Nature Reserve of central China’s Hubei Province. J. Beijing For. Univ. 2005, 27, 12–16. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, G.X.; Liu, G.H.; Liu, X. Population structure and distribution pattern of Davidia involucrata at Labahe Nature Reserve of Sichuan Province. J. Zhejiang For. Coll. 2008, 25, 451–457. [Google Scholar] [CrossRef]
- Wu, M.K.; Shen, Z.J.; Liu, H.; Wu, Y.Y.; He, R.T.; Qiu, Y.; Yang, N.; Yang, N. Life table and survival analysis of nature Davidia involucrata population in Fanjing Mountain Nature Reserve, Guizhou Province of Southwest China. Chin. J. Ecol. 2012, 31, 1419–1424. [Google Scholar] [CrossRef]
- Wang, D.S.; Zhao, W.; Cheng, B.B.; Zhang, J.J. Potential Suitable Areas of Crataegus pinnatifida in China based on MaxEnt Modeling. Sci. Silvae Sin. 2022, 58, 43–50. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Peerj 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fang, Y.; Zhang, Y.; Wang, Z.M.; Ma, J.; Gao, J.; Yang, D.G.; Wei, Y.D.; Kang, F.F. Prediction of the Potential Geographical Distribution of Solanum sisymbriifolium in China. Plant Quar. 2023, 37, 52–56. [Google Scholar] [CrossRef]
- Wei, J.F.; Li, X.Z.; Lu, Y.Y.; Zhao, L.; Zhang, H.F.; Zhao, Q. Modeling the potential global distribution of Phenacoccus madeirensis Green under Various Climate Change Scenarios. Forests 2019, 10, 773. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Chen, S.T.; Gao, Y.; Yang, L.; Yu, H. Prediction of global potential suitable habitats of Nicotiana alata Link et Otto based on MaxEnt model. Sci. Rep. 2023, 13, 4851. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Zhang, W.P.; Hu, Y.Y.; Li, Z.H.; Feng, X.P.; Li, D.W. Predicting suitable distribution areas of Juniperus przewalskii in Qinghai Province under climate change scenarios. J. Appl. Ecol. 2021, 32, 2514–2524. [Google Scholar] [CrossRef]
- Jiang, C.X.; Zhang, X.Y.; Xie, W.Q.; Wang, R.L.; Feng, C.H.; Ma, L.; Li, Q.; Yang, Q.F.; Wang, H.J. Predicting the potential distribution of the fall armyworm Spodoptera frugiperda (J.E. Smith) under climate change in China. Glob. Ecol. Conserv. 2022, 33, e01994. [Google Scholar] [CrossRef]
- Hausfather, Z. Explainer: How ‘Shared Socioeconomic Pathways’ Explore Future Climate Change. Carbon Brief (19 April 2018). Available online: https://www.carbonbrief.org/explainer-how-shared-socioeconomic-pathways-explore-future-climate-change/ (accessed on 8 September 2023).
- Wu, Y.X.; Zhang, M.; Yang, Y.; Lyu, Z.M.; Zhang, X.Q.; Wang, L.S. Effects of Climate Changes on the Distribution of Osmanthus fragrans. J. Northwest For. Univ. 2022, 37, 129–134. [Google Scholar] [CrossRef]
- Teng, J.; Li, H.; Lu, S.F.; Yin, X.J.; Li, G.; Chen, Z.; Wang, Y. Responses of Cold-Temperate Coniferous Forest to Climate Change in Southwestern China. J. Northwest For. Univ. 2023, 38, 33–44. [Google Scholar]
- Fischer, G.; Nachtergaele, F.O.; Prieler, S.; Teixeira, E.; van Velthuizen, H.; Verelst, L.; Wiberg, D. Global Agro-Ecological Zones Assessment for Agriculture (GAEZ 2008); IIASA: Laxenburg, Austria; FAO: Rome, Italy, 2008. [Google Scholar]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2011, 3, 53–64. [Google Scholar] [CrossRef]
- Guo, Y.L.; Li, X.; Zhao, Z.F.; Wei, H.Y.; Gao, B.; Gu, W. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci. Rep. 2017, 7, 46221. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Jin, J.W.; Cheng, J.M. Predicting the potential geographic distribution and habitat suitability of two economic forest trees on the Loess Plateau, China. Forests 2021, 12, 747. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Guevara, L.; Gerstner, B.E.; Kass, J.M.; Anderson, R.P. Toward ecologically realistic predictions of species distributions: A cross-time example from tropical montane cloud forests. Glob. Chang. Biol. 2018, 24, 1511–1522. [Google Scholar] [CrossRef]
- Yackuic, C.B.; Chandler, R.; Zipkin, E.F.; Royle, J.A.; Nichols, J.D.; Campbell Grant, E.H.; Veran, S. Presence-only modelling using MAXENT: When can we trust the inferences? Methods Ecol. Evol. 2013, 4, 236–243. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Wang, R.; Jiang, C.; Liu, L.; Shen, Z.; Yang, J.; Wang, Y.; Hu, J.; Wang, M.; Hu, J.; Lu, X.; et al. Prediction of the potential distribution of the predatory mite Neoseiulus californicus McGregor in China using MaxEnt. Glob. Ecol. Conserv. 2021, 29, e01733. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.B.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; MacKey, B. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [PubMed]
- Somodi, I.; Lepesi, N.; Botta-Dukát, Z. Prevalence dependence in model goodness measures with special emphasis on true skill statistics. Ecol. Evol. 2017, 7, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Sánchez-Mercado, A.Y.; Ferrer-Paris, J.R. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2011; Volume 86, pp. 219–220. [Google Scholar] [CrossRef]
- Thapa, A.; Wu, R.D.; Hu, Y.B.; Nie, Y.G.; Singh, P.B.; Khatiwada, J.R.; Yan, L.; Gu, X.D.; Wei, F.W. Predicting the potential distribution of the endangered red panda across its entire range using MaxEnt modeling. Ecol. Evol. 2018, 8, 10542–10554. [Google Scholar] [CrossRef] [PubMed]
- Zarzo-Arias, A.; Penteriani, V.; Delgado, M.D.M.; Peón Torre, P.; García-González, R.; Mateo-Sánchez, M.C.; Vázquez García, P.; Dalerum, F. Identifying potential areas of expansion for the endangered brown bear (Ursus arctos) population in the Cantabrian Mountains (NW Spain). PLoS ONE 2019, 14, e0209972. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. MaxEnt modeling for predicting the potential geographical distribution of two peony species under climate change. Sci Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Leitão, R.P.; Zuanon, J.; Villéger, S.; Williams, S.; Baraloto, C.; Fortunel, C.; Mendonça, F.; Mouillot, D. Rare species contribute disproportionately to the functional structure of species assemblages. Proc. Biol. Sci. 2016, 283, 20160084. [Google Scholar] [CrossRef] [PubMed]
- Dee, L.E.; Cowles, J.; Isbell, F.; Pau, S.; Gaines, S.D.; Reich, P.B. When do ecosystem services depend on rare species? Trends Ecol. Evol. 2019, 34, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Girona, M.M.; Aakala, T.; Aquilué, N.; Bélisle, A.C.; Chaste, E.; Danneyrolles, V.; Díaz-Yáñez, O.; D’Orangeville, L.; Grosbois, G.; Gauthier, S.; et al. Challenges for the Sustainable Management of the Boreal Forest Under Climate Change. In Boreal Forests in the Face of Climate Change: Sustainable Management; Springer International Publishing: Cham, Switzerland, 2023; pp. 773–837. [Google Scholar] [CrossRef]
- Hu, J.; Feng, Y.; Zhong, H.; Liu, W.; Tian, X.; Wang, Y.; Tan, T.; Hu, Z.; Liu, Y. Impact of climate change on the geographical distribution and niche dynamics of Gastrodia elata. PeerJ 2023, 11, e15741. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.X. Response of Potential Geographical Distribution of Eight China’s First-Class Rare and Endangered Plants to Climate Change and Analysis of GAP; Northwest Normal University: Lanzhou, China, 2021. [Google Scholar] [CrossRef]
- Liu, H.Y.; Jin, X.L.; Xue, H.W.; Luo, X.M.; Zhang, R.Q. Research Progress on Community Characteristics and Population Ecology of Davidia involucrata Baill. Chin. Agric. Sci. Bull. 2012, 28, 1–4. [Google Scholar] [CrossRef]
- Su, Z.X.; Zhang, S.L. The reproductive Phenology and the Influencing Factors of Davidia involucrata Population. J. China West Norm. Univ. Nat. Sci. 1999, 20, 313–318. [Google Scholar] [CrossRef]
- Li, S.; Mo, S.; Hu, X.H.; Deng, T. Prediction of potential suitable areas of endangered plant Abies ziyuanensis based on MaxEnt and ArcGIS. Chin. J. Ecol. 2023, 1–11. Available online: http://kns.cnki.net/kcms/detail/21.1148.Q.20230311.1445.004.html (accessed on 20 October 2023).
- Xu, Y. Predicting the Suitable Habitats of Relic Plants Ginkgo biloba and Davidia involucrate; North China Electric Power University: Beijing, China, 2019. [Google Scholar] [CrossRef]
- Sun, Y.A.; Wang, C.T.; Chen, H.Y.H.; Ruan, H.H. Response of Plants to Water Stress: A Meta-Analysis. Front. Plant Sci. 2020, 11, 978. [Google Scholar] [CrossRef]
- Jiang, R.F.; Liu, Y.H. Effects of Soil Matrix and Moisture on the Growth of Davidia involucrata Seedlings. J. Northwest For. Univ. 2016, 31, 134–139+164. [Google Scholar] [CrossRef]
- Ding, K.Y.; Liu, Y.H. Effects of Nitrogen and Continuous Ngihttime Low Temperature on Photosynthetic Characteristics of Davidia involucrata Leaves. J. Northeast. For. Univ. 2015, 43, 56–61. [Google Scholar] [CrossRef]
- Peng, H.L.; Su, Z.X. Effects of Low Temperature Stress on the Activity of Perox Dase, the Content of Chlorophyll and Free Proline in Davidia involucrata Seedling. J. Hanzhong Teach. Coll. Nat. Sci. 2004, 22, 50–53. [Google Scholar] [CrossRef]
- Zeng, R.; Ma, L.; Wu, J.Y.; Yang, G.; Zhang, N.; Xu, J.; Zhu, M.C.; Ma, M.; Tao, X.L.; Li, X.C.; et al. Relationship between withered leaf stage and cold resistance of winter Brassica rapa under low temperature stress. Chin. J. Oil Crop Sci. 2023, 45, 766–775. [Google Scholar] [CrossRef]
- Li, L.B.; Huang, G.Y.; Wu, D.; Zhang, H.B.; Wang, X.Y.; Wang, L.; Wu, J.H. Responses of Davidia involucrata Leaves to Different Elevations on Physiological Indices and Photosynthetic Characteristics. Mol. Plant Breed. 2023, 1–10. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20230228.0938.006.html (accessed on 20 October 2023).
- Wang, N.N. Responses of Photosynthetic Characteristics of Davidia involucrata Baill. Seedlings to Drought Stress, Shading and Carbon Dioxide Elevation; Beijing Forestry University: Beijing, China, 2010. [Google Scholar]
- Liu, Y.; Wang, Y.R.; Hou, G.W.; Liu, X.A.; Chen, C.; Xiao, X.Y.; Wang, H.Q. Analysis of Natural Population Dynamics of Davidia involucrata in Bayuelin Nature Reserve. J. Sichuan For. Sci. Technol. 2018, 39, 87–90. [Google Scholar] [CrossRef]
- Yin, L.M.; Dijkstra, F.A.; Wang, P.; Zhu, B.; Cheng, W. Rhizosphere priming effects on soil carbon and nitrogen dynamics among tree species with and without intraspecific competition. New Phytol. 2018, 218, 1036–1048. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Tian, L.; Huang, Y.; Shao, C. The Ginkgo biloba L. in China: Current Distribution and Possible Future Habitat. Forests 2023, 14, 2284. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Environmental Variables | Percent Contribution (%) | Permutation Importance (%) |
|---|---|---|---|
| Bio18 | Precipitation of Warmest Quarter | 30 | 28 |
| Bio9 | Mean Temperature of Driest Quarter | 24.4 | 51 |
| Bio20 | Annual mean radiation | 14.6 | 15.3 |
| Bio2 | Mean Diurnal Range | 12.7 | 1.1 |
| ele | Elevation | 12.6 | 0.7 |
| Bio10 | Mean Temperature of Warmest Quarter | 1.7 | 0.7 |
| T_USDA | Topsoil USDA Texture Classification | 1.6 | 1.2 |
| T_CEC_clay | Topsoil CEC (clay) | 1.1 | 0.1 |
| T_CAC | Topsoil Calcium Carbonate | 0.4 | 0.9 |
| T_pH | Topsoil pH (H2O) | 0.4 | 0.1 |
| Bio15 | Precipitation Seasonality | 0.3 | 0.5 |
| T_gravel | Topsoil gravel content | 0.2 | 0.2 |
| Period | Current | 2050s (2041–2060) | 2070s (2061–2080) | ||||
|---|---|---|---|---|---|---|---|
| ssp126 | ssp370 | ssp585 | ssp126 | ssp370 | ssp585 | ||
| Low-suitable area | 37.41 | 29.01 | 32.41 | 32.29 | 29.78 | 34.50 | 33.98 |
| Medium-suitable area | 25.27 | 21.08 | 21.07 | 23.13 | 20.23 | 22.99 | 20.26 |
| High-suitable area | 35.34 | 34.06 | 31.94 | 36.84 | 34.00 | 34.42 | 36.93 |
| Total | 98.02 | 84.15 | 85.42 | 92.26 | 84.01 | 91.91 | 91.17 |
| Period | Area/(×104 km2) | Change Rate/(%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Expansion | Unchanged | Contraction | Change | Increment | Unchanged | Loss | Change | |
| 2050s ssp126 | 6.47 | 77.27 | 20.54 | 14.07 | 6.60 | 78.83 | 20.95 | 14.35 |
| 2050s ssp370 | 8.78 | 76.27 | 21.54 | 12.76 | 8.96 | 77.81 | 21.98 | 13.02 |
| 2050s ssp585 | 8.43 | 83.51 | 14.31 | 5.88 | 8.60 | 85.20 | 14.60 | 6.00 |
| 2070s ssp126 | 6.28 | 77.34 | 20.47 | 14.19 | 6.40 | 78.90 | 20.88 | 14.48 |
| 2070s ssp370 | 8.93 | 82.67 | 15.14 | 6.21 | 9.11 | 84.34 | 15.45 | 6.34 |
| 2070s ssp585 | 9.62 | 81.23 | 16.58 | 6.96 | 9.81 | 82.87 | 16.91 | 7.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Li, W.; Cui, H.; Song, Y.; Liu, C.; Yan, Q.; Wu, Y.; Jia, Y.; Fang, L.; Qi, L. Predicting the Potential Habitat Distribution of Relict Plant Davidia involucrata in China Based on the MaxEnt Model. Forests 2024, 15, 272. https://doi.org/10.3390/f15020272
Wang T, Li W, Cui H, Song Y, Liu C, Yan Q, Wu Y, Jia Y, Fang L, Qi L. Predicting the Potential Habitat Distribution of Relict Plant Davidia involucrata in China Based on the MaxEnt Model. Forests. 2024; 15(2):272. https://doi.org/10.3390/f15020272
Chicago/Turabian StyleWang, Tianxiang, Wenting Li, Hongxia Cui, Yunrui Song, Changyong Liu, Qing Yan, Yaoxing Wu, Yihang Jia, Lizheng Fang, and Lianghua Qi. 2024. "Predicting the Potential Habitat Distribution of Relict Plant Davidia involucrata in China Based on the MaxEnt Model" Forests 15, no. 2: 272. https://doi.org/10.3390/f15020272
APA StyleWang, T., Li, W., Cui, H., Song, Y., Liu, C., Yan, Q., Wu, Y., Jia, Y., Fang, L., & Qi, L. (2024). Predicting the Potential Habitat Distribution of Relict Plant Davidia involucrata in China Based on the MaxEnt Model. Forests, 15(2), 272. https://doi.org/10.3390/f15020272