Concentration of Nutrients in Individual Organs of European Beech (Fagus sylvatica L.) Seedlings and Root System Development as a Result of Different Fertilization


Abstract
1. Introduction
2. Material and Methods
2.1. Experiment Design
2.2. Laboratory Analysis
2.3. Statistical Analysis
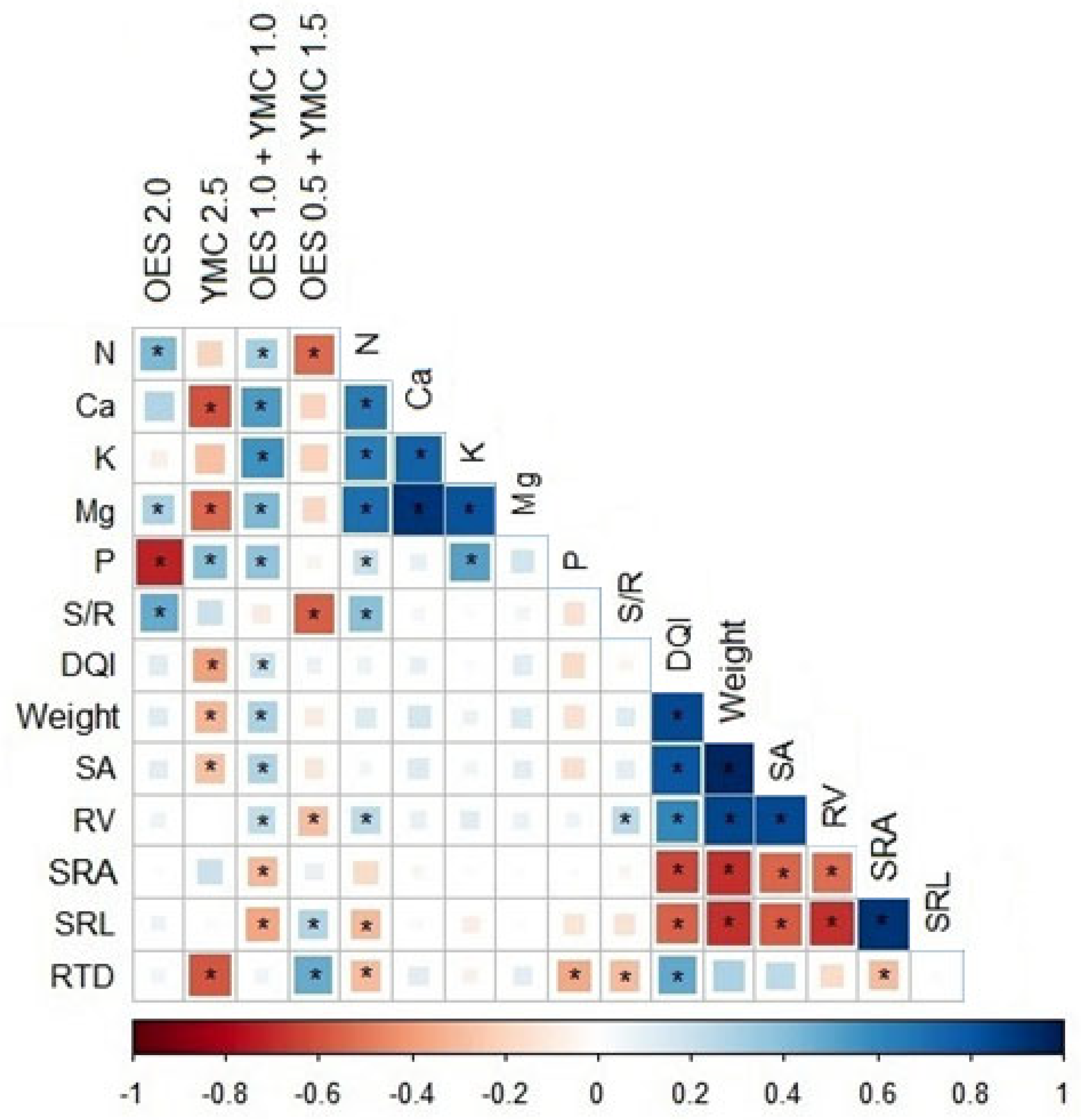
3. Results
3.1. Seedling Analysis
3.2. The Relationship Between Root Traits, Biometric and Fertilizer
4. Discussion
4.1. Chemical Properties of Seedlings
4.2. Biometric Analysis of Seedlings
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Trnka, M.; Olesen, J.E.; Kersebaum, A.C.; Skjelvåg, A.O.; Eitzinger, J.; Seguin, B.; Dubrovský, M. Agrolimatic conditions in Europe under climate change. Glob. Chang. Biol. 2011, 17, 2298–2318. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Roy, D.B. Altered geographic and temporal variability in phenology in response to climate change. Glob. Ecol. Biogeogr. 2006, 15, 498–504. [Google Scholar] [CrossRef]
- Jurásek, A.; Bartoš, J.; Nárovcová, J. Intensively fertilised seedlings of the beech (Fagus sylvatica L.) for artificial regeneration of the spruce stands in the process of conversion. J. For. Sci. 2008, 54, 452–458. [Google Scholar] [CrossRef]
- Jonard, M.; Fürst, A.; Verstraeten, A.; Thimonier, A.; Timmermann, V.; Potočić, N.; Rautio, P. Tree mineral nutrition is deteriorating in Europe. Glob. Chang. Biol. 2015, 21, 418–430. [Google Scholar] [CrossRef]
- Lukac, M.; Calfapietra, C.; Lagomarsino, A.; Loreto, F. Global climate change and tree nutrition: Effects of elevated CO2 and temperature. Tree Physiol. 2010, 30, 1209–1220. [Google Scholar] [CrossRef]
- Sierota, Z. Czy wprowadzanie w odnowieniach sadzonek z zakrytym systemem korzeniowym wszędzie znajduje uzasadnienie—Punkt widzenia fitopatologa (Is the introduction of covered root seedlings in every renewal reasonable—Phytopathological point of view). Sylwan 2019, 163, 989–996. [Google Scholar] [CrossRef]
- Hirano, Y.; Mizoguchi, T.; Brunner, I. Root parameters of forest trees as sensitive indicators of acidifying pollutants: A review of research of Japanese forest trees. J. For. Res. 2007, 12, 134–142. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. For. Ecol. Manag. 2011, 262, 2008–2023. [Google Scholar] [CrossRef]
- Staszel, K.; Błońska, E.; Lasota, J. Fine root morphology and soil properties under influence of different tree stands along an altitudinal climosequence in the Carpathian mountains. For. Ecosyst. 2022, 9, 100066. [Google Scholar] [CrossRef]
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P. Preserving the nutritional quality of crop plants under a changing climate: Importance and strategies. Plant Soil 2019, 443, 1–26. [Google Scholar] [CrossRef]
- Matías, L.; Castro, J.; Zamora, R. Soil-nutrient availability under a global-change scenario in a Mediterranean mountain ecosystem. Glob. Chang. Biol. 2011, 17, 1646–1657. [Google Scholar] [CrossRef]
- Arndal, M.F.; Merrild, M.P.; Michelsen, A.; Schmidt, I.K.; Mikkelsen, T.N.; Beier, C. Net root growth and nutrient acquisition in response to predicted climate change in two contrasting heathland species. Plant Soil 2013, 369, 615–629. [Google Scholar] [CrossRef]
- Pang, W.; Hou, D.; Wang, H. Preparation of Microcapsules of Slow-Release NPK Compound Fertilizer and the Release Characteristics. J. Braz. Chem. Soc. 2018, 29, 2397–2404. [Google Scholar] [CrossRef]
- Hasan, M.M.; Teixeira da Silva, J.A.; Li, X. Regulation of phosphorus uptake and utilization: Transitioning from current knowledge to practical strategies. Cell. Mol. Biol. Lett. 2016, 21, 7. [Google Scholar] [CrossRef] [PubMed]
- Talkner, U.; Meiwes, K.J.; Potočić, N.; Seletković, I.; Cools, N.; de Vos, B.; Rautio, P. Phosphorus nutrition of beech (Fagus sylvatica L.) is decreasing in Europe. Ann. For. Sci. 2015, 72, 919–928. [Google Scholar] [CrossRef]
- Nilsson, L.O.; Wallander, H. Production of external mycelium by ectomycorrhizal fungi in a Norway spruce forest was reduced in response to nitrogen fertilization. New Phytol. 2003, 158, 409–416. [Google Scholar] [CrossRef]
- Mašková, T.; Herben, T. Root: Shoot ratio in developing seedlings: How seedlings change their allocation in response to seed mass and ambient nutrient supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Nursery fertilization affects seedling traits but not field performance in Quercus suber L. J. Arid Environ. 2010, 74, 491–497. [Google Scholar] [CrossRef]
- Bosiacki, M.; Golcz-Polaszewska, M.; Kozik, E. Slow-release fertilizers in the production of horticultural plants. Part I. Effect of Osmocote Exact Standard fertilizer on the growth and condition of nourishing of selected taxons of ornamental trees and shrubs. J. Res. Appl. Agric. Eng. 2009, 54, 29–35. [Google Scholar]
- Wu, L.; Liu, M. Preparation and properties of chitosan-coated NPK compound fertilizer with controlled-release and water-retention. Carbohydr. Polym. 2008, 72, 240–247. [Google Scholar] [CrossRef]
- Qiao, D.; Liu, H.; Yu, L. Preparation and characterization of slow-release fertilizer encapsulated by starch-based superabsorbent polymer. Carbohydr. Polym. 2016, 147, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Zhu, Z.; Jiang, Y. Long-term impact of fertilization on soil pH and fertility in an apple production system. J. Soil Sci. Plant Nutr. 2018, 18, 282–293. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.W.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Qin, J.; Jiang, X.; Chen, S.; Gao, F.; Shen, D. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Banach, J.; Kempf, M.; Skrzyszewska, K.; Olejnik, K. The effect of starter fertilization on the growth of seedlings of European beech Fagus sylvatica L. Sylwan 2021, 165, 565–576. [Google Scholar] [CrossRef]
- Szołtyk, G.; Zajączkowski, P. Nawożenie doglebowe. In Szkółkarstwo Leśne od, A. do Z: Praca Zbiorowa; Wesoły, W., Hauke, M., Eds.; Centrum Informacyjne Lasów Państwowych: Warszawa, Poland, 2009; pp. 233–241. ISBN 978-83-89744-81-4. [Google Scholar]
- Banach, J.; Kormanek, M.; Małek, S.; Durło, G.; Skrzyszewska, K. Effect of the changing seedlings density of Quercus robur L. grown in nursery containers on their morphological traits and planting suitability. Sylwan 2023, 167, 1–12. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Lasn, R. The role of soil conditions in fine root ecomorphology in Norway spruce (Picea abies (L.) Karst.). Plant Soil 1999, 208, 283–292. [Google Scholar] [CrossRef]
- Staszel, K.; Lasota, J.; Błońska, E. Effect of drought on root exudates from Quercus petraea and enzymatic activity of soil. Sci. Rep. 2022, 12, 7635. [Google Scholar] [CrossRef]
- Wesoły, W.; Hauke, M.; Sienkiewicz, A. Nawożenie Dolistne oraz Stosowanie Nawozów Wieloskładnikowych o długim Okresie Działania w Szkółkach Kontenerowych i Otwartych (Foliar Fertilization and the Use of Compound Fertilizers with a Long Period of Action in Container and Open Nurseries); Wesoły, W., Ed.; Szkółkarstwo leśne od, A. do Z. Warszawa; Centrum Informacyjne Lasów Państwowych: Warszawa, Poland, 2009; pp. 241–254. (In Polish)
- Mellert, K.H.; Göttlein, A. Comparison of new foliar nutrient thresholds derived from VAN DEN BURG’s literature compilation with established central European references. Eur. J. For. Res. 2012, 131, 1461–1472. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 December 2023).
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13. 2017. Available online: https://docs.tibco.com/products/spotfire-statistica/archive (accessed on 12 December 2023).
- Mellert, K.H.; Prietzel, J.; Straussberger, R.; Rehfuess, K.E.; Kahle, H.P.; Perez, P.; Spiecker, H. Relationships between long-term trends of air temperature, precipitation, nitrogen nutrition and growth of coniferous stands in Central Europe and Finland. Eur. J. For. Res. 2008, 127, 507–524. [Google Scholar] [CrossRef]
- Sardans, J.; Alonso, R.; Janssens, I.A.; Carnicer, J.; Vereseglou, S.; Rillig, M.C.; Fernandez-Martinez, M.; Sanders, T.G.M.; Penuelas, J. Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across European Pinus sylvestris forests: Relationships with climate, N deposition and tree growth. Funct. Ecol. 2016, 30, 676–689. [Google Scholar] [CrossRef]
- Talkner, U.; Riek, W.; Dammann, I.; Kohler, M.; Göttlein, A.; Mellert, K.H.; Meiwes, K.J. Nutritional status of major forest tree species in Germany. In Status and Dynamics of Forests in Germany: Results of the National Forest Monitoring; Wellbrock, N., Bolte, A., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 261–293. [Google Scholar]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 2008; Volume 2, pp. 11–99. [Google Scholar]
- Xiulan, L.; Yingchun, L.; Hai, N.; Lixia, X. The effect of low phosphorus stress on main physiological traits of different maize genotypes. Zuo Wu Xue Bao 2005, 31, 667–669. [Google Scholar]
- Razaq, M.; Zhang, P.; Shen, H.L. Salahuddin Influence of nitrogen and phosphorus on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [PubMed]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.Q.; Guo, D.L.; Xu, X.L. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Eissenstat, D.M. Costs and benefits of constructing roots of small diameter. J. Plant Nutr. 1992, 15, 763–782. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Uscola, M.; Jacobs, D.F. The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New For. 2015, 46, 813–839. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; Ivetić, V. Root system development and field establishment: Effect of seedling quality. New Forests 2022, 53, 1021–1067. [Google Scholar] [CrossRef]
- Luoranen, J.; Rikala, R. Nutrient loading of Norway spruce seedlings hastens bud burst and enhances root growth after out planting. Silva Fenn. 2011, 45, 319–329. [Google Scholar] [CrossRef]
- Bernier, P.Y.; Lamhamedi, M.S.; Simpson, D.G. Shoot:root ratio is of limited use in evaluating the quality of container conifer stock. Tree Plant. Notes 1995, 46, 102–106. [Google Scholar]
- Tsakaldimi, M.; Ganatsas, P.; Jacobs, D.F. Prediction of planted seedling survival of five Mediterranean species based on initial seedling morphology. New For. 2012, 44, 327–339. [Google Scholar] [CrossRef]
- Ivetić, V.; Grossnickle, S.; Škorić, M. Forecasting the field performance of Austrian pine seedlings using morphological attributes. iForest 2016, 10, 99–107. [Google Scholar] [CrossRef]
- Guimarães, Z.T.M.; Da Silva, D.C.; Ferreira, M.J. Seedling quality and short-term field performance of three Amazonian forest species as affected by site conditions. iForest 2024, 17, 80–89. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| pH | Macronutrients | |||||
|---|---|---|---|---|---|---|
| C | N | Ca | K | Mg | P | |
| % | ||||||
| 4.87 ± 0.08 | 46.3 ± 1.2 | 0.79 ± 0.0 | 0.8604 ± 0.08 | 0.10 ± 0.01 | 0.37 ± 0.07 | 0.03 ± 0.00 |
| Variant Designation | Fertilizer Dose in kg per 1 m3 of Substrate | Macroelement | ||||
|---|---|---|---|---|---|---|
| Yara Mila Complex (YMC) | Osmocote Exact Standard (OES) | N | P | K | Mg | |
| OES 2.0 | – | 2.0 | 1 | 4 | 4 | 4 |
| YMC 2.5 | 2.5 | – | 2 | 1 | 1 | 1 |
| OES 1.0 + YMC 1.0 | 1.0 | 1.0 | 3 | 3 | 3 | 3 |
| OES 0.5 + YMC 1.5 | 1.5 | 0.5 | 4 | 2 | 2 | 2 |
| Nutrient | Wesoły et al., 2009 [29] | Mellert and Goettlein 2012 [30] | |
|---|---|---|---|
| Optimal Values | Normal Range | Surplus Luxury | |
| N | 1.30–2.20 | 1.87–2.32 | 2.32–2.75 |
| P | 0.15–0.30 | 0.12–0.19 | 0.19–0.20 |
| K | 0.50–2.00 | 0.61–0.97 | 0.97–1.30 |
| Ca | 0.20–0.80 | 0.67–1.40 | >1.40 |
| Mg | 0.10–3.0 | 0.11–0.23 | >0.23 |
| Root Parameters | OES 2.0 | YMC 2.5 | OES 1.0 + YMC 1.0 | OES 0.5 + YMC 1.5 |
|---|---|---|---|---|
| S/R | 1.67 ± 0.13 a | 1.60 ± 0.14 a,b | 1.52 ± 0.10 b,c | 1.40 ± 0.11 c |
| DQI | 0.60 ± 0.03 a | 0.46 ± 0.02 b | 0.63 ± 0.04 a | 0.59 ± 0.03 a |
| Weight [mg] | 421.09 ± 100.31 a,b | 346.96 ± 100.91 b | 497.73 ± 167.64 a | 386.75 ± 115.11 a,b |
| SA [cm2] | 79.00 ± 13.06 a,b | 68.28 ± 18.31 b | 89.87 ± 26.57 a | 73.56 ± 17.99 a,b |
| RV [cm3] | 0.26 ± 0.06 a,b | 0.26 ± 0.07 a,b | 0.31 ± 0.11 a | 0.22 ± 0.06 b |
| SRL [m kg−1] | 224.18 ± 37.93 a,b | 219.80 ± 23.84 a,b | 200.92 ± 23.96 b | 236.77 ± 29.09 a |
| SRA [m2 kg−1] | 19.78 ± 2.29 a | 20.36 ± 1.54 a | 18.91 ± 1.28 a | 20.00 ± 1.55 a |
| RTD [kg m−3] | 175.25 ± 17.10 a | 151.43 ± 13.48 b | 175.53 ± 21.67 a | 191.15 ± 20.37 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasik, M.; Staszel-Szlachta, K.; Małek, S.; Banach, J. Concentration of Nutrients in Individual Organs of European Beech (Fagus sylvatica L.) Seedlings and Root System Development as a Result of Different Fertilization. Forests 2024, 15, 2016. https://doi.org/10.3390/f15112016
Jasik M, Staszel-Szlachta K, Małek S, Banach J. Concentration of Nutrients in Individual Organs of European Beech (Fagus sylvatica L.) Seedlings and Root System Development as a Result of Different Fertilization. Forests. 2024; 15(11):2016. https://doi.org/10.3390/f15112016
Chicago/Turabian StyleJasik, Michał, Karolina Staszel-Szlachta, Stanisław Małek, and Jacek Banach. 2024. "Concentration of Nutrients in Individual Organs of European Beech (Fagus sylvatica L.) Seedlings and Root System Development as a Result of Different Fertilization" Forests 15, no. 11: 2016. https://doi.org/10.3390/f15112016
APA StyleJasik, M., Staszel-Szlachta, K., Małek, S., & Banach, J. (2024). Concentration of Nutrients in Individual Organs of European Beech (Fagus sylvatica L.) Seedlings and Root System Development as a Result of Different Fertilization. Forests, 15(11), 2016. https://doi.org/10.3390/f15112016