
The Distributional Range Changes of European Heterobasidion Under Future Climate Change

Abstract
1. Introduction
2. Materials and Methods
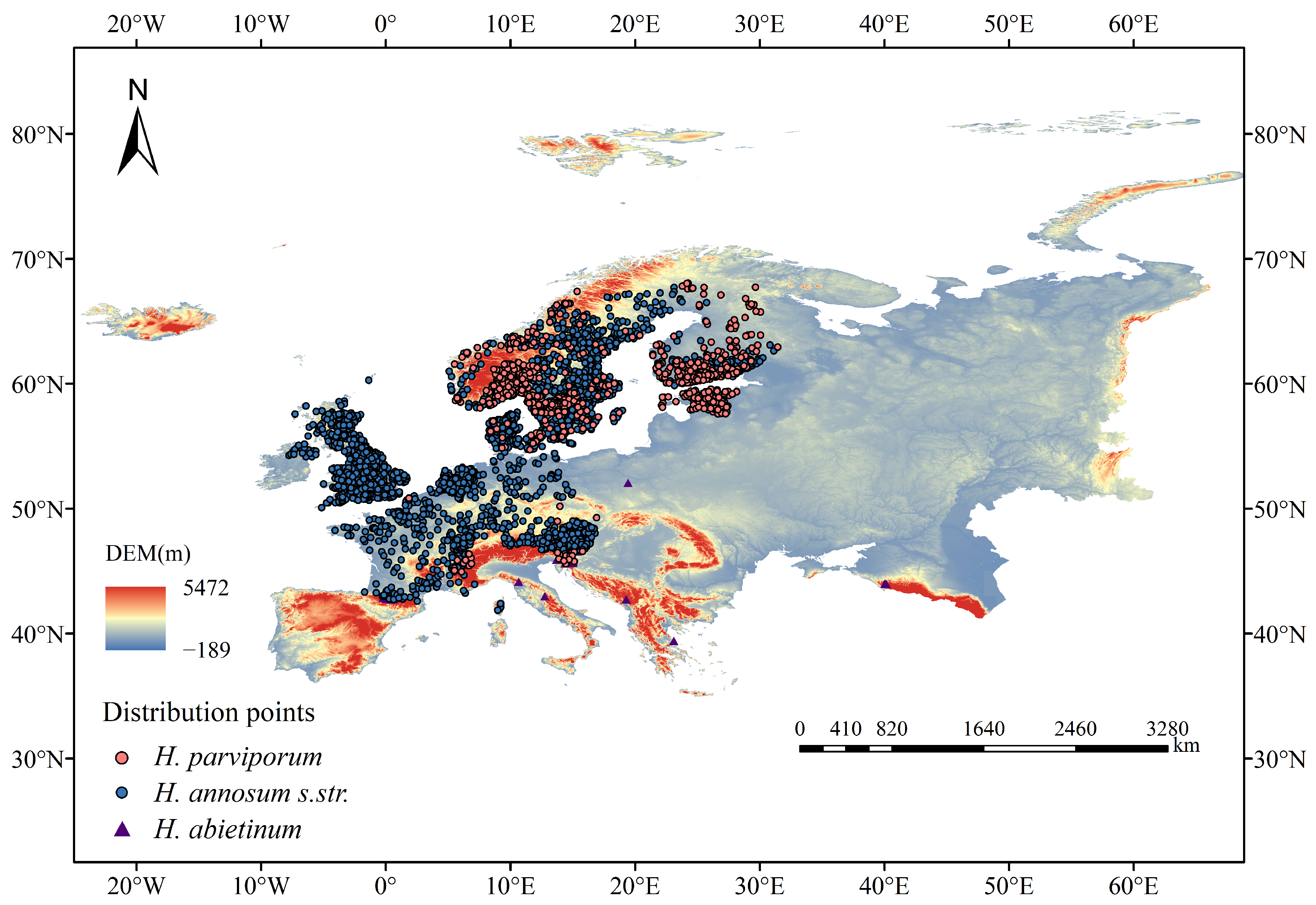
2.1. Collecting and Processing Obtained Species Occurrence Data
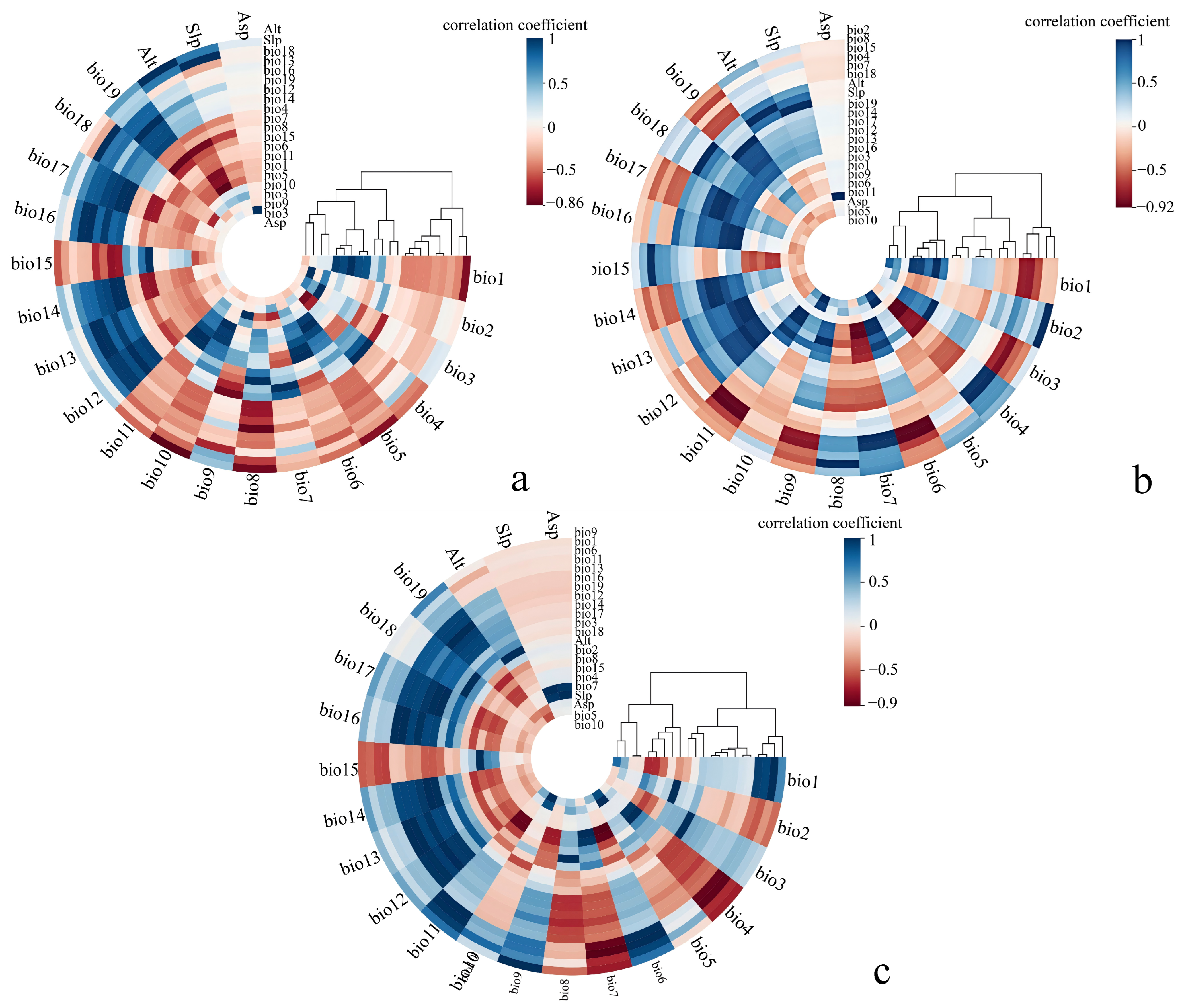
2.2. Collecting and Processing of Environmental Variables
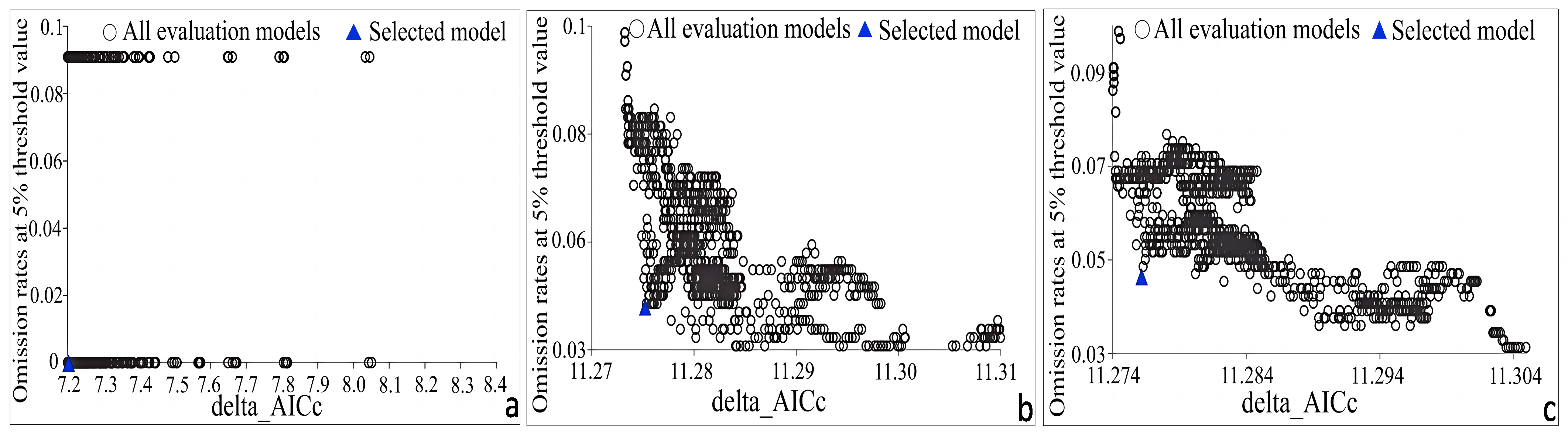
2.3. Model Evaluation
2.4. Changes of Suitable Habitat Area and Centroids
3. Results
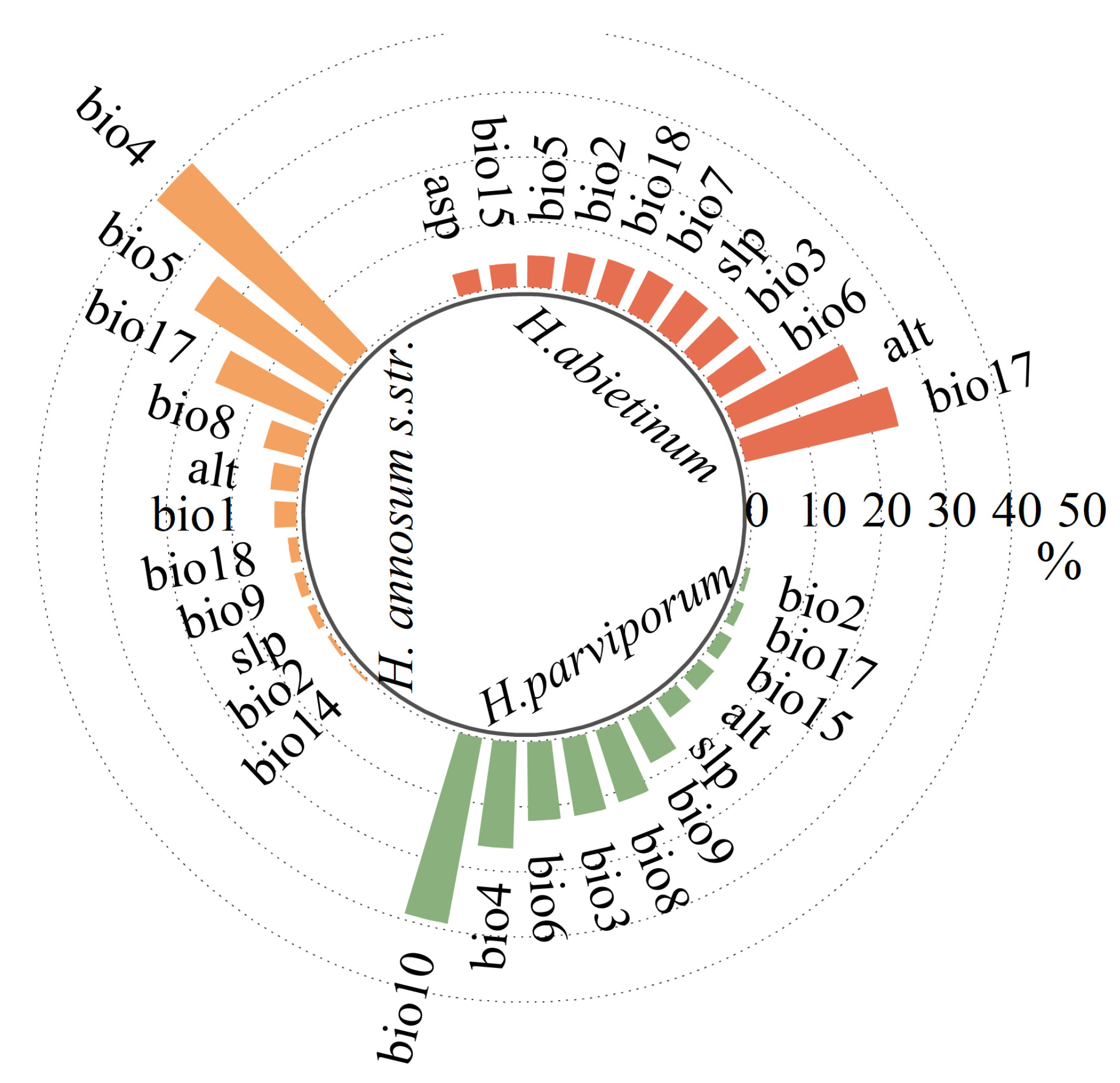
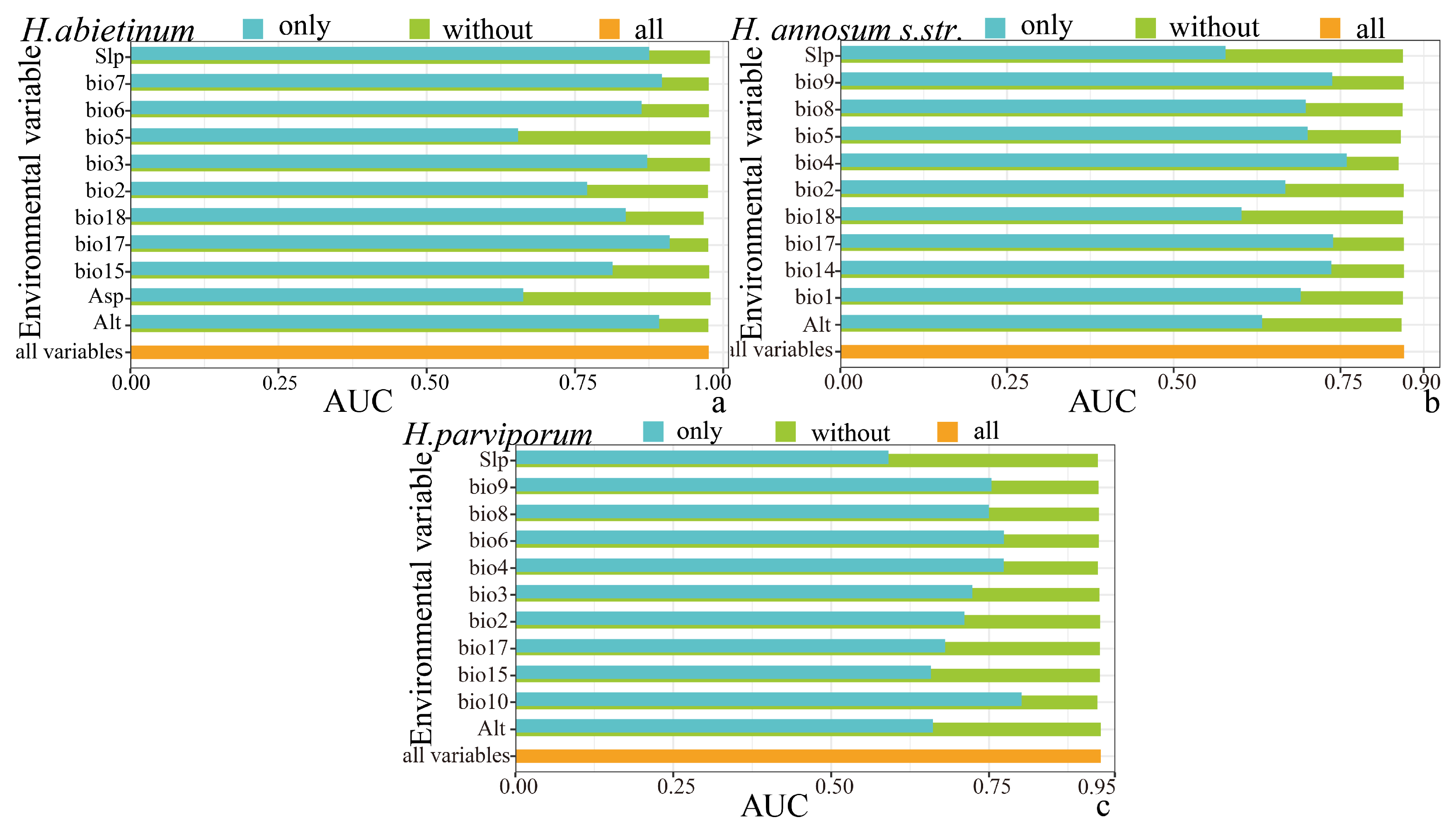
3.1. Model Variables and Performance Evaluation
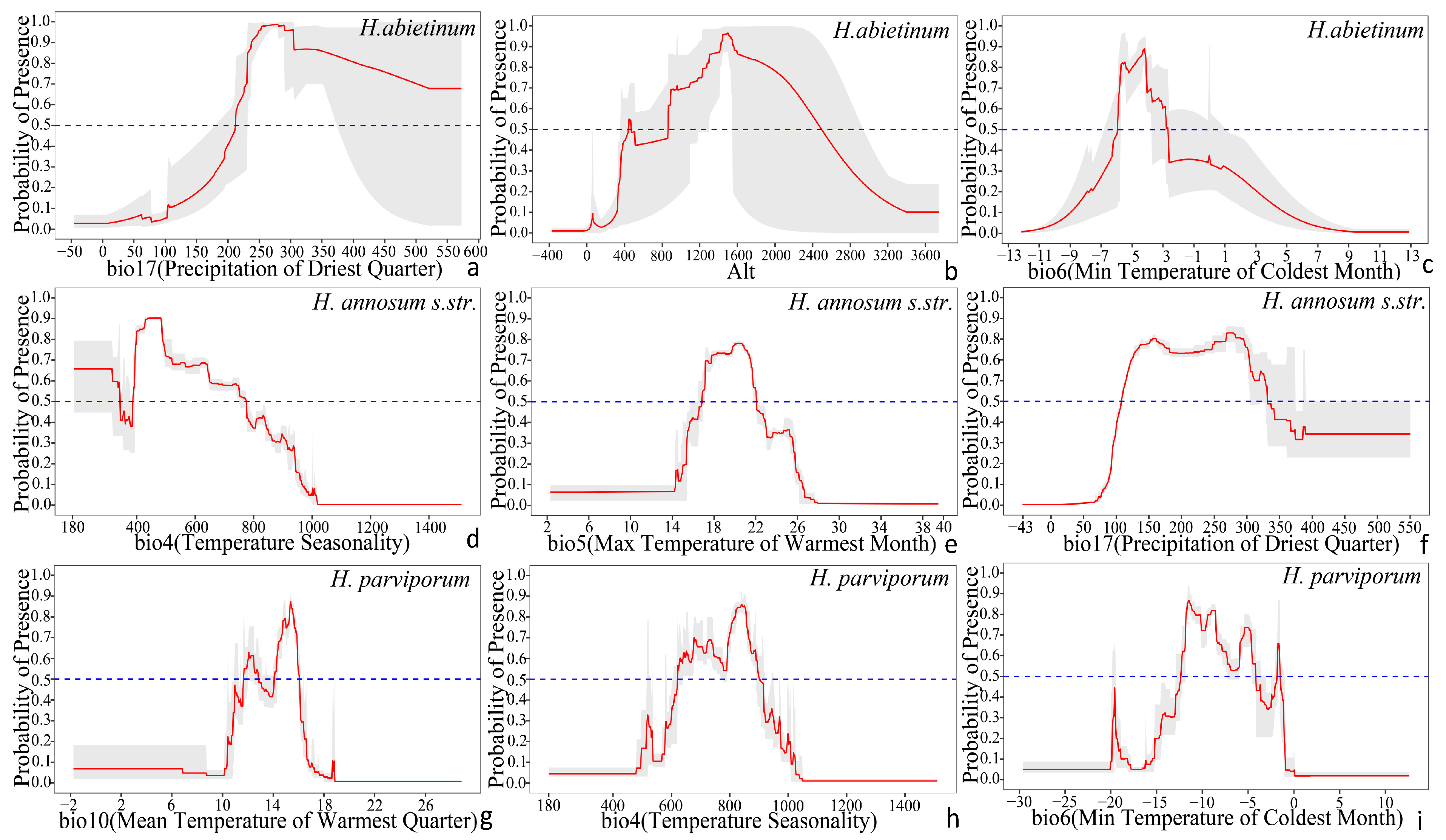
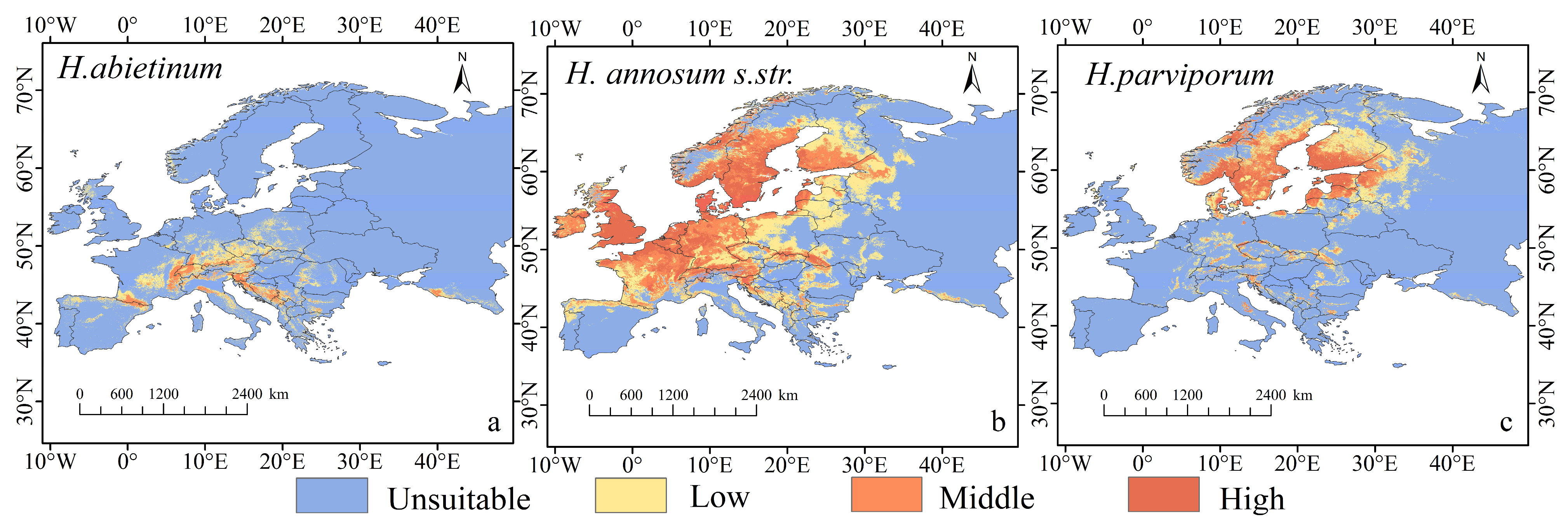
3.2. Current Distribution and Dominant Environmental Variables
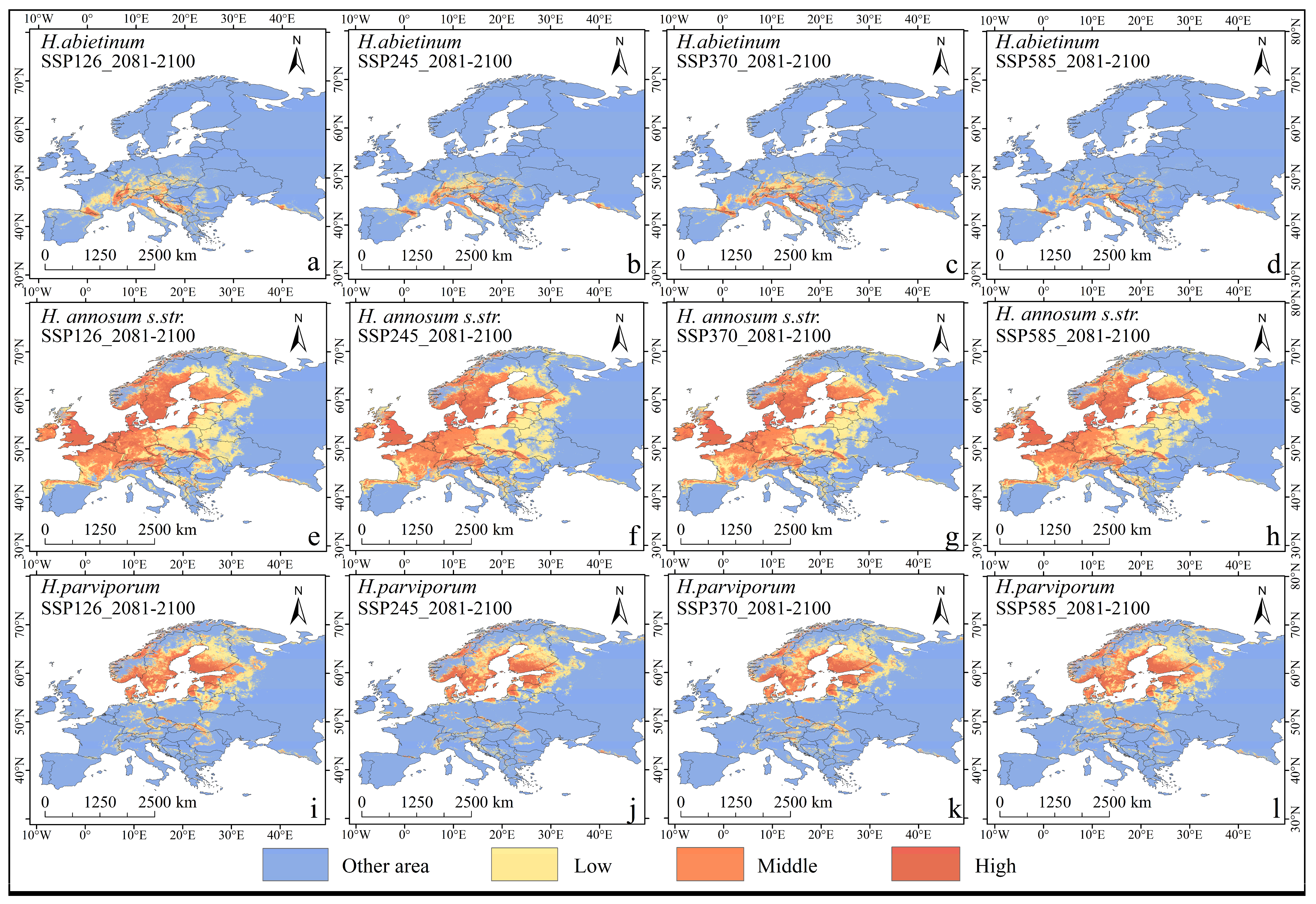
3.3. Distribution Characteristics Under Future Climate Models
3.4. Changes in Spatial Pattern and Centroid
4. Discussion
4.1. The Impact of Environmental Factors on Distribution
4.2. Changes in Spatial Pattern and Centroid
4.3. Study Limitations
4.4. Future Perspectives
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Yuan, Y.; Huang, S.; Ma, Y.; Hong, Z.; Wang, Y.; Wu, X.; Li, Z.; Ye, J.; Zhang, L. Geographical distribution and predict potential distribution of Angelica L. genus. Environ. Sci. Pollut. Res. 2023, 30, 46562–46573. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Sun, Y.; Zhang, X. Temperature and precipitation projection at 1.5 and 2 °C increase in global mean temperature. Chin. Sci. Bull. 2017, 62, 3098–3111. [Google Scholar] [CrossRef]
- Linnakoski, R.; Kasanen, R.; Dounavi, A.; Forbes, K.M. Editorial: Forest Health Under Climate Change: Effects on Tree Resilience, and Pest and Pathogen Dynamics. Front. Plant Sci. 2019, 10, 1157. [Google Scholar] [CrossRef]
- Ostry, M.E.; Laflamme, G. Fungi and diseases—Natural components of healthy forests. Botany 2009, 87, 22–25. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Brockerhoff, E.G.; Wingfield, B.D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef]
- Ayres, M.P.; Lombardero, M.J. Assessing the consequences of global change for forest disturbance from herbivores and pathogens. Sci. Total Environ. 2000, 262, 263–286. [Google Scholar] [CrossRef]
- Maijala, P.; Harrington, T.C.; Raudaskoski, M. A peroxidase gene family and gene trees in Heterobasidion and related genera. Mycologia 2003, 95, 209–221. [Google Scholar] [CrossRef]
- Asiegbu, F.O.; Adomas, A.; Stenlid, J. Conifer root and butt rot caused by Heterobasidion annosum (Fr.) Bref. s.l. Mol. Plant Pathol. 2010, 6, 395–409. [Google Scholar] [CrossRef]
- Paul, N.C.; Deng, J.X.; Shin, K.S.; Yu, S.H. Molecular and Morphological Characterization of Endophytic Heterobasidion araucariae from Roots of Capsicum annuum L. in Korea. Mycobiology 2012, 40, 85–90. [Google Scholar] [CrossRef]
- Hellsten, S.; Helmisaari, H.S.; Melin, Y.; Skovsgaard, J.P.; Kaakinen, S.; Kukkola, M.; Saarsalmi, A.; Petersson, H.; Akselsson, C. Nutrient concentrations in stumps and coarse roots of Norway spruce, Scots pine and silver birch in Sweden, Finland and Denmark. (Special Issue: Environmental effects of tree-stump harvesting). For. Ecol. Manag. 2013, 290, 40–48. [Google Scholar] [CrossRef]
- Dai, Y.C.; Korhonen, K. Heterobasidion australe, a new polypore derived from the Heterobasidion insulare complex. Mycoscience 2009, 50, 353–356. [Google Scholar] [CrossRef]
- Tokuda, S.; Hattori, T.; Dai, Y.C.; Ota, Y.; Buchanan, P.K. Three species of Heterobasidion (Basidiomycota, Hericiales), H. parviporum, H. orientale sp. nov. and H. ecrustosum sp. nov. from East Asia. Mycoscience 2009, 50, 190–192. [Google Scholar] [CrossRef]
- Kashif, M.; Hyder, R.; Perez, D.D.V.; Hantula, J.; Vainio, E. Heterobasidion wood decay fungi host diverse and globally distributed viruses related to Helicobasidium mompa partitivirus V70. Virus Res. 2015, 195, 119–123. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Zaļuma, A.; Kenigsvalde, K.; Kļaviņa, D.; Brauners, I.; Piri, T. Susceptibility of Small-Diameter Norway Spruce Understory Stumps to Heterobasidion Spore Infection. Forests 2019, 10, 521. [Google Scholar] [CrossRef]
- Hyder, R.; Piri, T.; Hantula, J.; Nuorteva, H.; Vainio, E.J. Distribution of Viruses Inhabiting Heterobasidion annosum in a Pine-Dominated Forest Plot in Southern Finland. Microb. Ecol. Int. J. 2018, 75, 631. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hyder, R.; Aday, G.; Hansen, E.; Piri, T.; Doğmuş-Lehtijärvi, T.; Lehtijärvi, A.; Korhonen, K.; Hantula, J. Population structure of a novel putative mycovirus infecting the conifer root-rot fungus Heterobasidion annosum sensu lato. Virology 2012, 422, 366–376. [Google Scholar] [CrossRef]
- Woodward, S.; Stenlid, J.; Karjalainen, R.; Hüttermann, A. Heterobasidion annosum: Biology, Ecology, Impact and Control; CAB International: Wallingford, UK, 2000; Volume 148, pp. 127–128. [Google Scholar]
- Zu, K.; Wang, Z.; Zhu, X.; Lenoir, J.; Shrestha, N.; Lyu, T.; Luo, A.; Li, Y.; Ji, C.; Peng, S.; et al. Upward shift and elevational range contractions of subtropical mountain plants in response to climate change. Sci. Total Environ. 2021, 783, 146896. [Google Scholar] [CrossRef]
- Atwater, D.Z.; Barney, J.N. Climatic niche shifts in 815 introduced plant species affect their predicted distributions. Glob. Ecol. Biogeogr. 2021, 30, 1671–1684. [Google Scholar] [CrossRef]
- Qiao, H.; Lin, C.; Ji, L.; Jiang, Z. mMWeb—An Online Platform for Employing Multiple Ecological Niche Modeling Algorithms. PLoS ONE 2012, 7, e43327. [Google Scholar] [CrossRef]
- Barbosa, F.G.; Schneck, F. Characteristics of the top-cited papers in species distribution predictive models. Ecol. Model. 2015, 313, 77–83. [Google Scholar] [CrossRef]
- Vaz, U.L.; Cunha, H.F.; Nabout, J.C. Trends and biases in global scientific literature about ecological niche models. Braz. J. Biol. 2015, 75, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, R.; Perez, V.; Gégout, J.-C. Disregarding the edaphic dimension in species distribution models leads to the omission of crucial spatial information under climate change: The case of Quercus pubescens in France. Glob. Chang. Biol. 2012, 18, 2648–2660. [Google Scholar] [CrossRef]
- Wang, Y.H.; Jiang, W.M.; Comes, H.P.; Hu, F.S.; Qiu, Y.X.; Fu, C.X. Molecular phylogeography and ecological niche modelling of a widespread herbaceous climber, Tetrastigma hemsleyanum (Vitaceae): Insights into Plio-Pleistocene range dynamics of evergreen forest in subtropical China. New Phytol. 2015, 206, 852–867. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2010, 14, 763–773. [Google Scholar] [CrossRef]
- van Proosdij, A.S.; Sosef, M.S.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 39, 366–376. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 6, 1–5. [Google Scholar]
- Zheng, C.; Wen, Z.M.; Liu, Y.Y.; Guo, Q.; Jiang, Y.M.; Ren, H.Y.; Fan, Y.M.; Yang, Y.T. Integrating Habitat Suitability and the Near-Nature Restoration Priorities into Revegetation Plans Based on Potential Vegetation Distribution. Forests 2021, 12, 218. [Google Scholar] [CrossRef]
- Lee, C.M.; Lee, D.-S.; Kwon, T.-S.; Athar, M.; Park, Y.-S. Predicting the Global Distribution of Solenopsis geminata (Hymenoptera: Formicidae) under Climate Change Using the MaxEnt Model. Insects 2021, 12, 229. [Google Scholar] [CrossRef]
- Fordham, D.A.; Wigley, T.M.L.; Brook, B.W. Multi-model climate projections for biodiversity risk assessments. Ecol. Appl. 2011, 21, 3317–3331. [Google Scholar] [CrossRef]
- Lovato, T.; Peano, D.; Butenschön, M.; Materia, S.; Iovino, D.; Scoccimarro, E.; Fogli, P.G.; Cherchi, A.; Bellucci, A.; Gualdi, S.; et al. CMIP6 Simulations with the CMCC Earth System Model (CMCC-ESM2). J. Adv. Model. Earth Syst. 2022, 14, e2021MS002814. [Google Scholar] [CrossRef]
- Wyser, K.; van Noije, T.; Yang, S.T.; von Hardenberg, J.; O’Donnell, D.; Döscher, R. On the increased climate sensitivity in the EC-Earth model from CMIP5 to CMIP6. Geosci. Model Dev. 2020, 13, 3465–3474. [Google Scholar] [CrossRef]
- Volodin, E.M. Possible Climate Change in Russia in the 21st Century Based on the INM-CM5-0 Climate Model. Russ. Meteorol. Hydrol. 2022, 47, 327–333. [Google Scholar] [CrossRef]
- Müller, W.A.; Jungclaus, J.H.; Mauritsen, T.; Baehr, J.; Bittner, M.; Budich, R.; Bunzel, F.; Esch, M.; Ghosh, R.; Haak, H.; et al. A Higher-resolution Version of the Max Planck Institute Earth System Model (MPI-ESM1.2-HR). J. Adv. Model. Earth Syst. 2018, 10, 1383–1413. [Google Scholar] [CrossRef]
- Tatebe, H.; Ogura, T.; Nitta, T.; Komuro, Y.; Ogochi, K.; Takemura, T.; Sudo, K.; Sekiguchi, M.; Abe, M.; Saito, F.; et al. Description and basic evaluation of simulated mean state, internal variability, and climate sensitivity in MIROC6. Geosci. Model Dev. 2019, 12, 2727–2765. [Google Scholar] [CrossRef]
- Yukimoto, S.; Kawai, H.; Koshiro, T.; Oshima, N.; Yoshida, K.; Urakawa, S.; Tsujino, H.; Deushi, M.; Tanaka, T.; Hosaka, M.; et al. The Meteorological Research Institute Earth System Model Version 2.0, MRI-ESM2.0: Description and Basic Evaluation of the Physical Component. J. Meteorol. Soc. Jpn. 2019, 97, 931–965. [Google Scholar] [CrossRef]
- Kumar, S.; Graham, J.; West, A.M.; Evangelista, P.H. Using district-level occurrences in Max Ent for predicting the invasion potential of an exotic insect pest in India. Comput. Electron. Agric. 2014, 103, 55–62. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef]
- Wang, E.; Lu, Z.R.; Rohani, E.R.; Ou, J.M.; Tong, X.H.; Han, R.C. Current and future distribution of Forsythia suspensa in China under climate change adopting the MaxEnt model. Front. Plant Sci. 2024, 15, 1394799. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P.; Mcpherson, J. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2015, 5, 1198–1205. [Google Scholar] [CrossRef]
- Yan, X.; Wang, S.; Duan, Y.; Han, J.; Huang, D.; Zhou, J. Current and future distribution of the deciduous shrub Hydrangea macrophylla in China estimated by MaxEnt. Ecol. Evol. 2021, 11, 16099–16112. [Google Scholar] [CrossRef]
- Martinez, F.; Blancard, D.; Lecomte, P.; Levis, C.; Dubos, B.; Fermaud, M. Phenotypic Differences Between vacuma and transposa subpopulations of Botrytis cinerea. Eur. J. Plant Pathol. 2003, 109, 479–488. [Google Scholar] [CrossRef]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. Des. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef]
- Lonsdale, D. Effects of climate change on fungal disease of trees. Fungi Environ. Chang. 1996, 6, 1253022. [Google Scholar]
- Miles, J.W.; Grof, B. Recent advances in studies of anthracnose of Stylosanthes. III. Stylosanthes breeding approaches South America. Trop. Grassl. 1997, 31, 430–434. [Google Scholar]
- Milici, V.R.; Dalui, D.; Mickley, J.G.; Bagchi, R. Responses of plant–pathogen interactions to precipitation: Implications for tropical tree richness in a changing world. J. Ecol. 2020, 108, 1800–1809. [Google Scholar] [CrossRef]
- Munier, A.; Hermanutz, L.; Jacobs, J.D.; Lewis, K. The interacting effects of temperature, ground disturbance, and herbivory on seedling establishment: Implications for treeline advance with climate warming. Plant Ecol. 2010, 210, 19–30. [Google Scholar] [CrossRef]
- Ungerer, M.J.; Lombardero, A.M.J. Climate and the northern distribution limits of Dendroctonus frontalis Zimmermann (Coleoptera: Scolytidae). J. Biogeogr. 1999, 26, 1133–1145. [Google Scholar] [CrossRef]
- Burgess, S.S.O. Measuring transpiration responses to summer precipitation in a Mediterranean climate: A simple screening tool for identifying plant water-use strategies. Physiol. Plant 2006, 127, 404–441. [Google Scholar] [CrossRef]
- Porter, J.H.; Parry, M.L.; Carter, T.R. The potential effects of climatic change on agricultural insect pests. Agric. For. Meteorol. 1991, 57, 221–240. [Google Scholar] [CrossRef]
- Asante, S.K.; Danthanarayana, W.; Heatwole, H. Bionomics and population growth statistics of apterous virginoparae of woolly apple aphid, Eriosoma lanigerum, at constant temperatures. Entomol. Exp. Appl. 1991, 60, 261–270. [Google Scholar] [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nat. Rev. Microbiol. 2023, 21, 640–656. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; p. 2391. [Google Scholar]
- Bordi, I.; Sutera, F. The analysis of the Standardized Precipitation Index in the Mediterranean area: Regional patterns. Ann. Geophys. 2001, 44, 964–978. [Google Scholar] [CrossRef]
- Muller, M.; Olsson, P.O.; Eklundh, L.; Jamali, S.; Ardo, J. Features predisposing forest to bark beetle outbreaks and their dynamics during drought. For. Ecol. Manag. 2022, 523, 120480. [Google Scholar] [CrossRef]
- Benestad, R.E. Climate change scenarios for Northern Europe from multi-model IPCC AR4 climate simulations. Geophys. Res. Lett. 2005, 32, 261. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.T.W.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef]
- Rubio-Cuadrado, Á.; Camarero, J.J.; Gómez, C.; Cañellas, I.; Aulló-Maestro, I.; Gil, L.; Montes, F. Scots pine plantations growth adaptation to climate warming in locations at the southernmost distribution limit of the species. Dendrochronologia 2020, 63, 125745. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Climate warming will reduce growth and survival of Scots pine except in the far north. Ecol. Lett. 2008, 11, 588–597. [Google Scholar] [CrossRef]
- Wang, C.J.; Wan, J.Z. Envelope, Functional trait perspective on suitable habitat distribution of invasive plant species at a global scale. Perspect. Ecol. Conserv. 2021, 19, 475–486. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Species | Pathogenicity | Distribution |
|---|---|---|---|
| Pine-Parasitic Group | Heterobasidion annosum s.str. | Highly pathogenic | Europe |
| Heterobasidion irregulare | Highly pathogenic | North America | |
| Spruce-Parasitic Group | Heterobasidion abietinum | Highly pathogenic | Europe |
| Heterobasidion occidentale | Highly pathogenic | North America | |
| Heterobasidion parviporum | Highly pathogenic | Europe | |
| Heterobasidion subparviporum | Weakly pathogenic | Asia | |
| Saprotrophic Conifer Group | Heterobasidion amyloideum | Saprotrophic | Asia |
| Heterobasidion araucariae | Saprotrophic | Oceania | |
| Heterobasidion armandii | Saprotrophic | Asia | |
| Heterobasidion australe | Saprotrophic | Asia | |
| Heterobasidion insulare | Saprotrophic | Asia | |
| Heterobasidion linzhiense | Saprotrophic | Asia | |
| Heterobasidion orientale | Saprotrophic | Asia | |
| Heterobasidion subinsulare | Saprotrophic | Asia | |
| Heterobasidion tibeticum | Saprotrophic | Asia |
| Environmental Variable | Variable Description | Unit |
|---|---|---|
| Bio1 | Annual Mean Temperature | °C |
| Bio2 | Mean Diurnal Range (Mean of monthly (max temp-min temp)) | °C |
| Bio3 | Isothermality | - |
| Bio4 | Temperature Seasonality | - |
| Bio5 | Max Temperature of Warmest Month | °C |
| Bio6 | Min Temperature of Coldest Month | °C |
| Bio7 | Temperature Annual Range | °C |
| Bio8 | Mean Temperature of Wettest Quarter | °C |
| Bio9 | Mean Temperature of Driest Quarter | °C |
| Bio10 | Mean Temperature of Warmest Quarter | °C |
| Bio11 | Mean Temperature of Coldest Quarter | °C |
| Bio12 | Annual Precipitation | mm |
| Bio13 | Precipitation of Wettest Month | mm |
| Bio14 | Precipitation of Driest Month | mm |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) | mm |
| Bio16 | Precipitation of Wettest Quarter | mm |
| Bio17 | Precipitation of Driest Quarter | mm |
| Bio18 | Precipitation of Warmest Quarter | mm |
| Bio19 | Precipitation of Coldest Quarter | mm |
| Slp | Slope | - |
| Asp | Aspect | - |
| Alt | Altitude | m |
| Climate Scenario | Definition | Characteristics |
|---|---|---|
| SSP126 | Low emissions sustainable development pathway | Emphasizes the use of renewable energy, reducing greenhouse gas emissions, and implementing sustainable economic policies to achieve low emission targets. |
| SSP245 | Moderate emissions pathway | Combines economic growth with greenhouse gas reduction, adopting moderate policies to address climate change, resulting in medium-level emissions. |
| SSP370 | High emissions pathway | Continues current economic and energy policies, leading to a continuous rise in greenhouse gas emissions, with a lack of effective climate policies. |
| SSP585 | Very high emissions pathway | Faces significant environmental challenges with insufficient policy responses, resulting in very high greenhouse gas emissions and severe impacts of climate change. |
| Species | Climate Scenario | Area of Suitable Areas at Different Levels (×104 km2) and Variation Relative to Current Period (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Unsuitable | Low | Middle | High | ||||||
| ×104 km2 | % | ×104 km2 | % | ×104 km2 | % | ×104 km2 | % | ||
| H. abietinum | Current | 1187.93 | - | 70.09 | - | 16.34 | - | 7.38 | - |
| SSP126 | 1177.74 | −0.9 | 73.61 | 5 | 17.88 | 9 | 6.75 | −9 | |
| SSP245 | 1189.67 | 0.1 | 58.58 | −16 | 20.02 | 23 | 7.46 | 1 | |
| SSP370 | 1180.94 | −0.6 | 59.82 | −15 | 22.79 | 39 | 12.18 | 65 | |
| SSP585 | 1211.67 | 2 | 53.50 | −24 | 14.81 | −9 | 5.76 | −22 | |
| H. annosum s.str. | Current | 816.15 | - | 199.60 | - | 144.75 | - | 115.24 | - |
| SSP126 | 770.05 | −6 | 232.78 | 16 | 155.23 | 7 | 117.69 | 2 | |
| SSP245 | 782.85 | −4 | 224.07 | 5 | 161.31 | 11 | 107.51 | −7 | |
| SSP370 | 812.69 | −0.4 | 208.92 | 5 | 147.96 | 2 | 106.17 | −8 | |
| SSP585 | 816.48 | 0.04 | 196.98 | −1 | 150.70 | 4 | 111.57 | −3 | |
| H. parviporum | Current | 1048.12 | - | 117.60 | - | 63.87 | - | 46.15 | - |
| SSP126 | 1015.47 | −3 | 142.58 | 21 | 72.69 | 14 | 44.99 | −3 | |
| SSP245 | 1036.38 | −1 | 132.70 | 13 | 65.01 | 2 | 41.65 | −10 | |
| SSP370 | 1050.81 | 0.3 | 120.19 | 2 | 64.31 | 1 | 40.42 | −12 | |
| SSP585 | 1061.43 | 1 | 108.76 | −8 | 66.01 | 3 | 39.55 | −14 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, S.; Zhang, X.; Jian, S. The Distributional Range Changes of European Heterobasidion Under Future Climate Change. Forests 2024, 15, 1863. https://doi.org/10.3390/f15111863
Shen S, Zhang X, Jian S. The Distributional Range Changes of European Heterobasidion Under Future Climate Change. Forests. 2024; 15(11):1863. https://doi.org/10.3390/f15111863
Chicago/Turabian StyleShen, Shen, Xueli Zhang, and Shengqi Jian. 2024. "The Distributional Range Changes of European Heterobasidion Under Future Climate Change" Forests 15, no. 11: 1863. https://doi.org/10.3390/f15111863
APA StyleShen, S., Zhang, X., & Jian, S. (2024). The Distributional Range Changes of European Heterobasidion Under Future Climate Change. Forests, 15(11), 1863. https://doi.org/10.3390/f15111863