
The Diversity and Community Pattern of Liverworts on Sygera Mountain, Tibet


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Identification
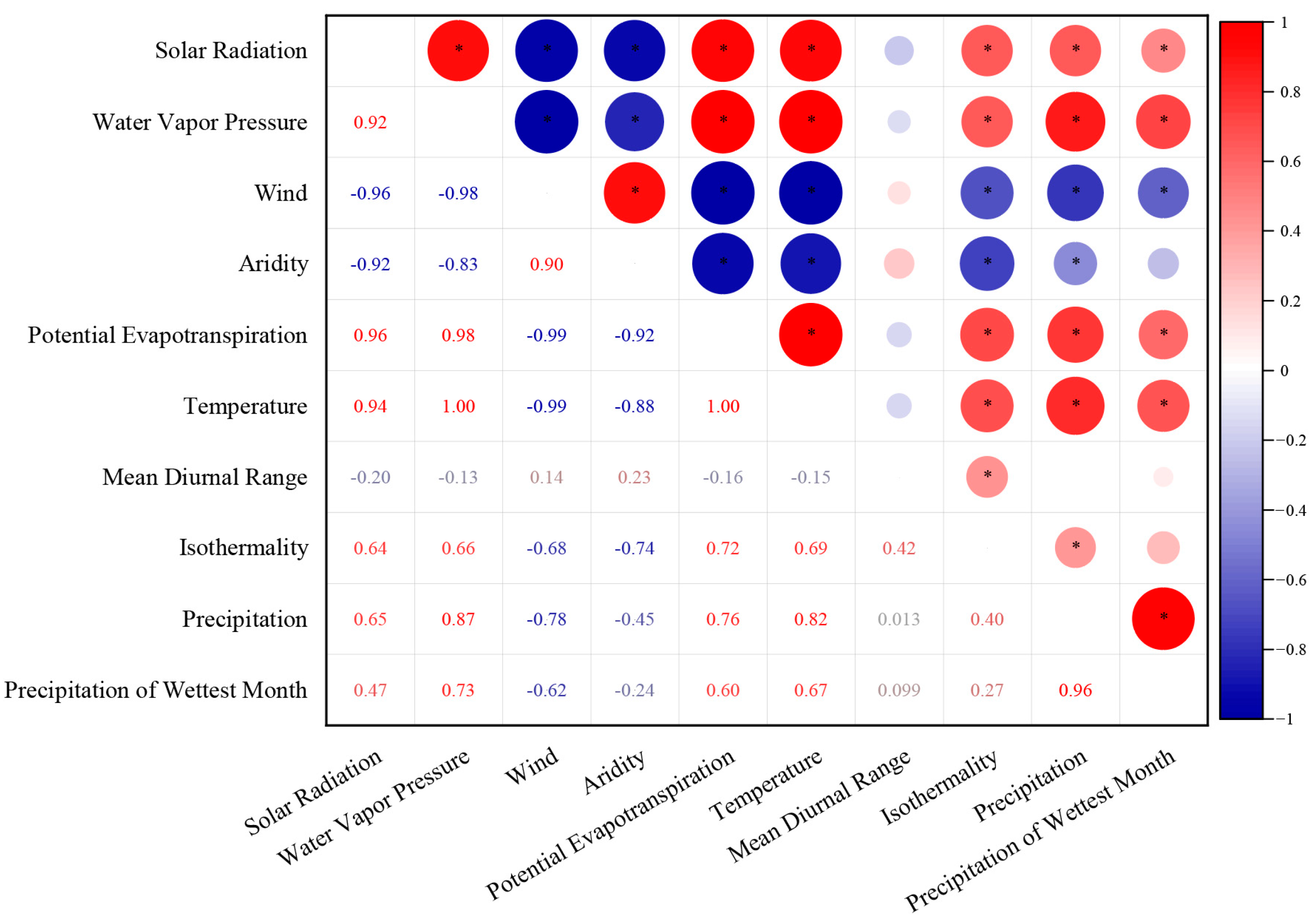
2.3. Environmental Variables
2.4. Data Analyses
3. Results
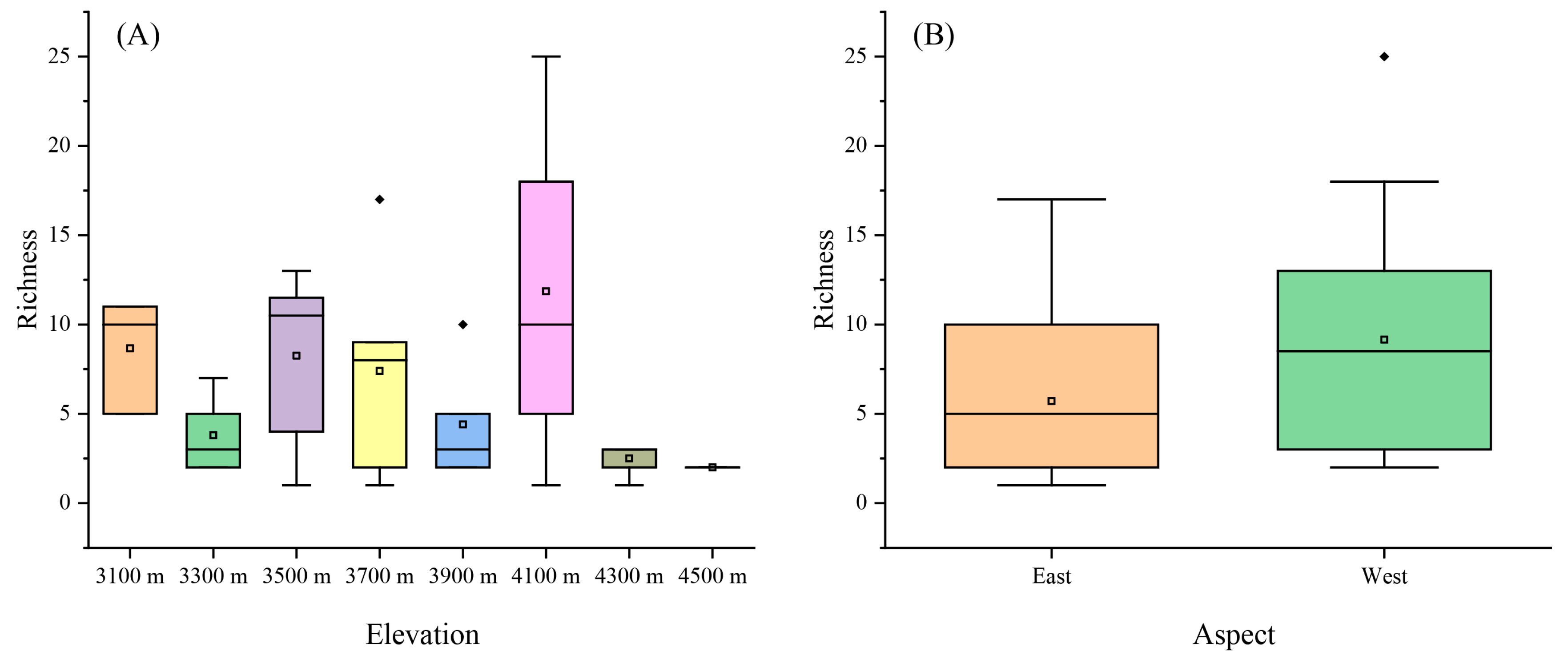
3.1. Variation in Liverwort Diversity with Elevation and Aspect
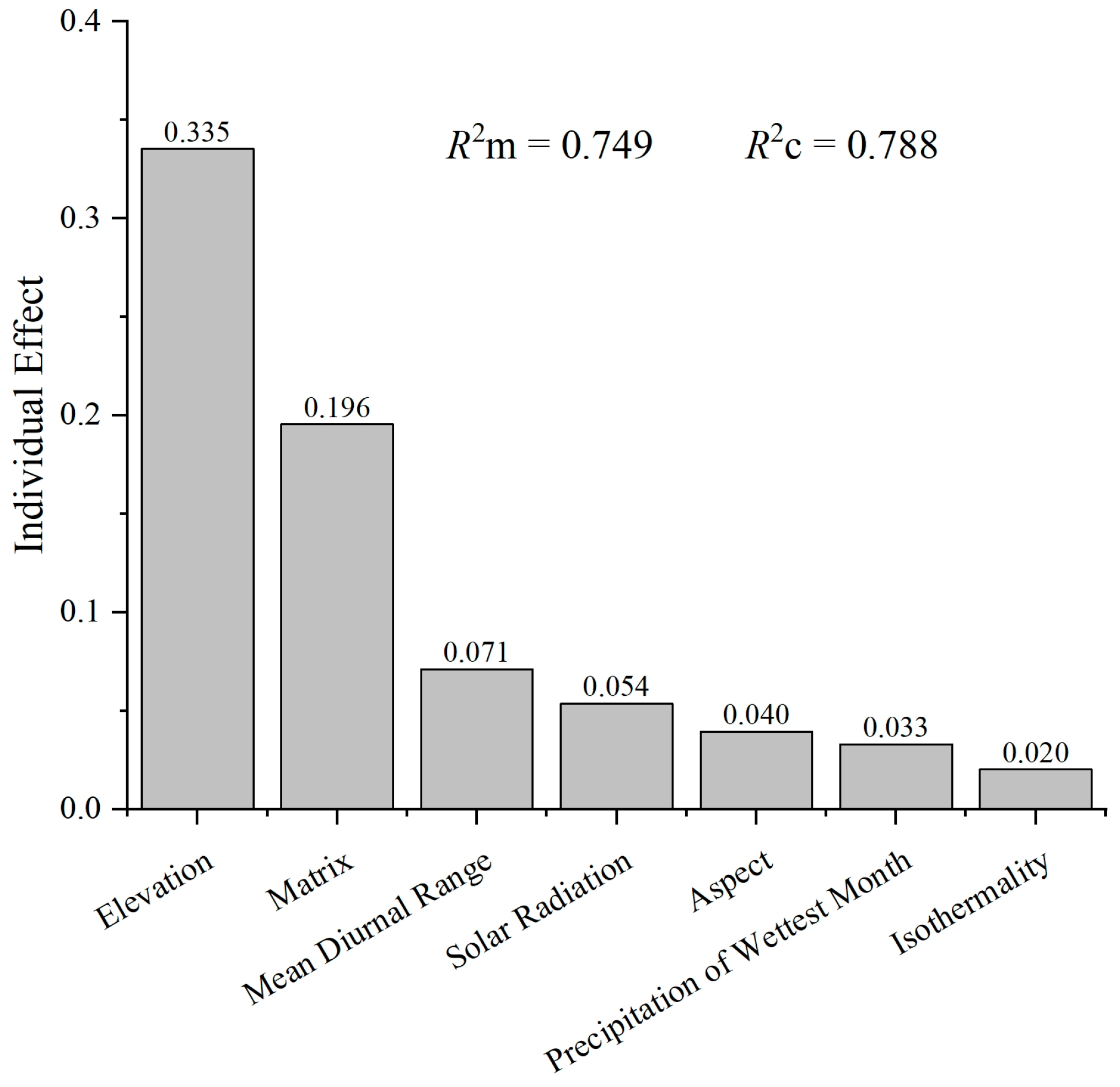
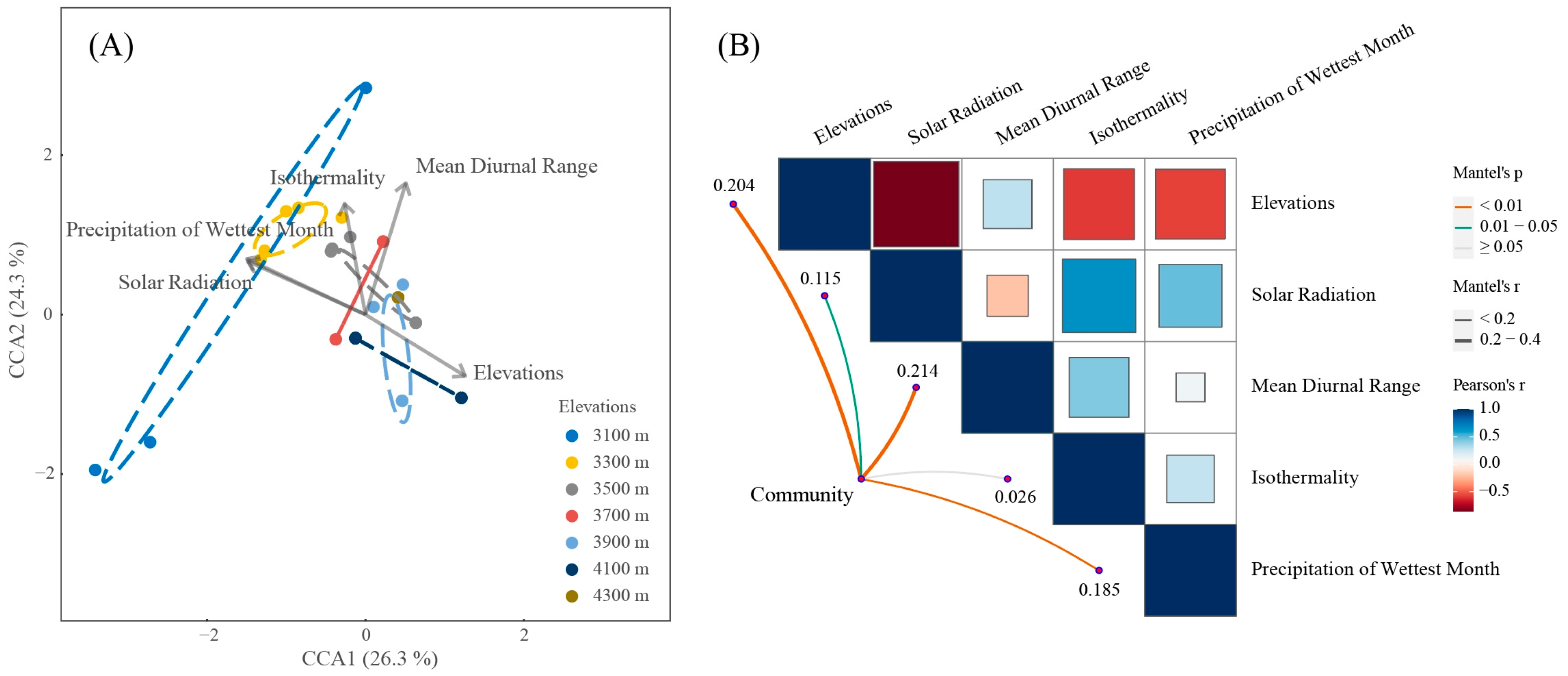
3.2. Relationship between Liverwort Communities and Environmental Factors
4. Discussion
4.1. The Influence of Elevation on Liverwort Diversity and Community Composition
4.2. The Impact of Climatic Factors on the Community Structure of Liverworts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Humboldt, A.V.; Bonpland, A. Essai sur la Géographie des Plantes Accompagné d’un Tableau Physique des Régions Équinoxiales; Levrault, Schoell et Cie: Paris, France, 1805. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; Volume 2. [Google Scholar]
- Ovaskainen, O.; Abrego, N. Joint Species Distribution Modelling: With Applications in R; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- McCain, C.M.; Grytnes, J. Elevational gradients in species richness. eLS 2010, 10, a0022548. [Google Scholar]
- Zhang, W.; Huang, D.; Wang, R.; Liu, J.; Du, N. Altitudinal patterns of species diversity and phylogenetic diversity across temperate mountain forests of northern China. PLoS ONE 2016, 11, e159995. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Anderson, M.J.; Arita, H.T.; Chao, A.; Colwell, R.K.; Connolly, S.R.; Currie, D.J.; Dunn, R.R.; Graves, G.R.; Green, J.L. Patterns and causes of species richness: A general simulation model for macroecology. Ecol. Lett. 2009, 12, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Kluge, J.; Worm, S.; Lange, S.; Long, D.; Böhner, J.; Yangzom, R.; Miehe, G. Elevational seed plants richness patterns in Bhutan, Eastern Himalaya. J. Biogeogr. 2017, 44, 1711–1722. [Google Scholar] [CrossRef]
- Liu, L.; Xu, Y.; Tang, Y.; Du, W.; Shao, C.; Wu, J.; Zhao, L.; Zhang, L.; Liu, J.; Xu, X. The importance of including soil properties when disentangling the drivers of species richness: The case of the alpine genus Saxifraga L. in China. Front. Ecol. Evol. 2020, 8, 244. [Google Scholar] [CrossRef]
- Descombes, P.; Leprieur, F.; Albouy, C.; Heine, C.; Pellissier, L. Spatial imprints of plate tectonics on extant richness of terrestrial vertebrates. J. Biogeogr. 2017, 44, 1185–1197. [Google Scholar] [CrossRef]
- Körner, C. Significance of temperature in plant life. In Plant Growth and Climate Change; Morison, J.I.L., Morecroft, M.D., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2006; pp. 48–69. [Google Scholar]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Kewlani, P.; Negi, V.S.; Bhatt, I.D.; Rawal, R.S.; Nandi, S.K. Soil nutrients concentration along altitudinal gradients in Indian Western Himalaya. Scand. J. For. Res. 2021, 36, 98–104. [Google Scholar] [CrossRef]
- Antonelli, A.; Kissling, W.D.; Flantua, S.G.; Bermúdez, M.A.; Mulch, A.; Muellner-Riehl, A.N.; Kreft, H.; Linder, H.P.; Badgley, C.; Fjeldså, J. Geological and climatic influences on mountain biodiversity. Nat. Geosci. 2018, 11, 718–725. [Google Scholar] [CrossRef]
- Perrigo, A.; Hoorn, C.; Antonelli, A. Why mountains matter for biodiversity. J. Biogeogr. 2020, 47, 315–325. [Google Scholar] [CrossRef]
- Gu, J.; Song, X.; Liao, Y.; Ye, Y.; Wang, R.; Ma, H.; Shao, X. Tree Species Drive the Diversity of Epiphytic Bryophytes in the Alpine Forest Ecosystem: A Case Study in Tibet. Forests 2022, 13, 2154. [Google Scholar] [CrossRef]
- Chen, Z.; Hsieh, C.; Jiang, F.; Hsieh, T.; Sun, I. Relations of soil properties to topography and vegetation in a subtropical rain forest in southern Taiwan. Plant Ecol. 1997, 132, 229–241. [Google Scholar] [CrossRef]
- Shaw, J. Biogeographic patterns and cryptic speciation in bryophytes. J. Biogeogr. 2001, 28, 253–261. [Google Scholar] [CrossRef]
- Ah-Peng, C.; Wilding, N.; Kluge, J.; Descamps-Julien, B.; Bardat, J.; Chuah-Petiot, M.; Strasberg, D.; Hedderson, T.A. Bryophyte diversity and range size distribution along two altitudinal gradients: Continent vs. island. Acta Oecol. 2012, 42, 58–65. [Google Scholar] [CrossRef]
- Grau, O.; Grytnes, J.A.; Birks, H. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J. Biogeogr. 2007, 34, 1907–1915. [Google Scholar] [CrossRef]
- Costa, D.P.; Nadal, F.; Da Rocha, T.C. The first botanical explorations of bryophyte diversity in the Brazilian Amazon mountains: High species diversity, low endemism, and low similarity. Biodivers. Conserv. 2020, 29, 2663–2688. [Google Scholar] [CrossRef]
- Tuba, Z.; Slack, N.G.; Stark, L.R. Bryophyte Ecology and Climate Change; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Virtanen, R. Diaspore and shoot size as drivers of local, regional and global bryophyte distributions. Glob. Ecol. Biogeogr. 2014, 23, 610–619. [Google Scholar] [CrossRef]
- Graham, L.; Lewis, L.A.; Taylor, W.; Wellman, C.; Cook, M. Early terrestrialization: Transition from algal to bryophyte grade. In Photosynthesis in Bryophytes and Early Land Plants; Hanson, D.T., Rice, S.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 9–28. [Google Scholar]
- Laenen, B.; Shaw, B.; Schneider, H.; Goffinet, B.; Paradis, E.; Désamoré, A.; Heinrichs, J.; Villarreal, J.C.; Gradstein, S.R.; McDaniel, S.F. Extant diversity of bryophytes emerged from successive post-Mesozoic diversification bursts. Nat. Commun. 2014, 5, 5134. [Google Scholar] [CrossRef]
- Ligrone, R.; Duckett, J.G.; Renzaglia, K.S. Conducting tissues and phyletic relationships of bryophytes. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 795–813. [Google Scholar] [CrossRef]
- Mishler, B.D.; Churchill, S.P. Transition to a land flora: Phylogenetic relationships of the green algae and bryophytes. Cladistics 1985, 1, 305–328. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, S.; Bell, N.; Hedenäs, L. The evolutionary diversity of mosses–taxonomic heterogeneity and its ecological drivers. Crit. Rev. Plant Sci. 2018, 37, 128–174. [Google Scholar] [CrossRef]
- Valarezo, E.; Meneses, M.A.; Jaramillo-Fierro, X.; Radice, M.; Benítez, Á. Volatile Compounds and Oils from Mosses and Liverworts. In Bioactive Compounds in Bryophytes and Pteridophytes; Springer: Cham, Switzerland, 2022; pp. 1–53. [Google Scholar]
- Ligrone, R.; Duckett, J.G.; Renzaglia, K.S. Major transitions in the evolution of early land plants: A bryological perspective. Ann. Bot.-Lond. 2012, 109, 851–871. [Google Scholar] [CrossRef] [PubMed]
- Marks, R.A.; Burton, J.F.; McLetchie, D.N. Sex differences and plasticity in dehydration tolerance: Insight from a tropical liverwort. Ann. Bot.-Lond. 2016, 118, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, C.P.; Franks, P.J.; Rees, M.; Bidartondo, M.I.; Leake, J.R.; Beerling, D.J. Mutualistic mycorrhiza-like symbiosis in the most ancient group of land plants. Nat. Commun. 2010, 1, 103. [Google Scholar] [CrossRef] [PubMed]
- Hallingbäck, T.; Hodgetts, N.G. Mosses, Liverworts, and Hornworts: Status Survey and Conservation Action Plan for Bryophytes; IUCN in Collaboration with the Swedish Threatened Species Unit: Gland, Switzerland, 2000. [Google Scholar]
- He, X.; He, K.S.; Hyvönen, J. Will bryophytes survive in a warming world? Perspect. Plant Ecol. Evol. Syst. 2016, 19, 49–60. [Google Scholar] [CrossRef]
- Dos Santos, N.D.; Da Costa, D.P. Altitudinal zonation of liverworts in the Atlantic Forest, Southeastern Brazil. Bryologist 2010, 113, 631–645. [Google Scholar] [CrossRef]
- Corlett, R. The Ecology of Tropical East Asia; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Shen, Z.; Lu, J.; Hua, M.; Tang, X.; Qu, X.; Xue, J.; Fang, J. Population structure and spatial pattern analysis of Quercus aquifolioides on Sejila Mountain, Tibet, China. J. For. Res. 2018, 29, 405–414. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Trabucco, A.; Zomer, R.J. Global Aridity Index and Potential Evapo-Transpiration (ET0) Climate Database v2; CGIAR Consortium for Spatial Information (CGIAR-CSI): Washington, DC, USA, 2018. [Google Scholar]
- Schielzeth, H.; Nakagawa, S. Nested by design: Model fitting and interpretation in a mixed model era. Methods Ecol. Evol. 2013, 4, 14–24. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Dray, S.; Dufour, A. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2015, 82, 1–26. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, S.; Zhang, X.; Mao, L. glmm.hp: An R package for computing individual effect of predictors in generalized linear mixed models. J. Plant Ecol. 2022, 15, 1302–1307. [Google Scholar] [CrossRef]
- Lai, J.; Zhu, W.; Cui, D.; Mao, L. Extension of the glmm.hp package to Zero-Inflated Generalized Linear Mixed Models and multiple regression. J. Plant Ecol. 2023, rtad038. [Google Scholar] [CrossRef]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots. 2020. Available online: https://rpkgs.datanovia.com/ggpubr/ (accessed on 20 October 2023).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.; Wagner, H. vegan: Community Ecology Package, R package version 2.5-3; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Grytnes, J.A. Ecological interpretations of the mid-domain effect. Ecol. Lett. 2003, 6, 883–888. [Google Scholar] [CrossRef]
- Kessler, M.; Hofmann, S.; Krömer, T.; Cicuzza, D.; Kluge, J. The impact of sterile populations on the perception of elevational richness patterns in ferns. Ecography 2011, 34, 123–131. [Google Scholar] [CrossRef]
- Frahm, J.; Gradstein, S.R. An altitudinal zonation of tropical rain forests using byrophytes. J. Biogeogr. 1991, 18, 669–678. [Google Scholar] [CrossRef]
- Gradstein, S.R.; Churchill, S.P.; Salazar-Allen, N. Guide to the Bryophytes of Tropical America; Memoirs-New York Botanical Garden: Bronx, NY, USA, 2001. [Google Scholar]
- Wolf, J. Diversity patterns and biomass of epiphytic bryophytes and lichens along an altitudinal gradient in the Northern Andes. Ann. Mo. Bot. Gard. 1993, 80, 928–960. [Google Scholar] [CrossRef]
- Glime, J.M. Temperature: Cold. In Bryophyte Ecology; Michigan Technological University: Houghton, MI, USA, 2013; Volume 1, Chapter 10-2. [Google Scholar]
- Górski, P.; Gądek, B.; Gąbka, M. Snow as a parameter of bryophyte niche partitioning in snow-beds of the Tatra Mountains (Western Carpathians). Ecol. Indic. 2020, 113, 106258. [Google Scholar] [CrossRef]
- Coelho, M.C.; Gabriel, R.; Hespanhol, H.; Borges, P.A.; Ah-Peng, C. Bryophyte diversity along an elevational gradient on pico island (Azores, portugal). Diversity 2021, 13, 162. [Google Scholar] [CrossRef]
- Aranda, S.C.; Gabriel, R.; Borges, P.A.; Santos, A.M.; de Azevedo, E.B.; Patiño, J.; Hortal, J.; Lobo, J.M. Geographical, temporal and environmental determinants of bryophyte species richness in the Macaronesian Islands. PLoS ONE 2014, 9, e101786. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Niu, S.; Li, P.; Jia, H.; Wang, H.; Ye, Y.; Yuan, Z. Stand structure and substrate diversity as two major drivers for bryophyte distribution in a temperate montane ecosystem. Front. Plant Sci. 2017, 8, 874. [Google Scholar] [CrossRef] [PubMed]
- Ah-Peng, C.; Flores, O.; Wilding, N.; Bardat, J.; Marline, L.; Hedderson, T.A.; Strasberg, D. Functional diversity of subalpine bryophyte communities in an oceanic island (La Réunion). Arct. Antarct. Alp. Res. 2014, 46, 841–851. [Google Scholar] [CrossRef]
- Patiño, J.; Bisang, I.; Hedenäs, L.; Dirkse, G.; Bjarnason, Á.H.; Ah Peng, C.; Vanderpoorten, A. Baker’s law and the island syndromes in bryophytes. J. Ecol. 2013, 101, 1245–1255. [Google Scholar] [CrossRef]
- Horwath, A.B.; Royles, J.; Tito, R.; Gudiño, J.A.; Salazar Allen, N.; Farfan-Rios, W.; Rapp, J.M.; Silman, M.R.; Malhi, Y.; Swamy, V. Bryophyte stable isotope composition, diversity and biomass define tropical montane cloud forest extent. Proc. R. Soc. B 2019, 286, 20182284. [Google Scholar] [CrossRef]
- Peters, K.; Poeschl, Y.; Blatt-Janmaat, K.L.; Uthe, H. Ecometabolomics Studies of Bryophytes. In Bioactive Compounds in Bryophytes and Pteridophytes; Springer: Cham, Switzerland, 2023; pp. 637–679. [Google Scholar]
- Yadav, S.; Srivastava, A.; Biswas, S.; Basu, S.; Singh, S.K.; Mishra, Y. Seasonal changes in the antioxidative defence system of a liverwort Dumortiera hirsuta. J. Plant Growth Regul. 2022, 41, 1265–1275. [Google Scholar] [CrossRef]
- Fenton, N.J.; Frego, K.A.; Sims, M.R. Changes in forest floor bryophyte (moss and liverwort) communities 4 years after forest harvest. Can. J. Bot.-Rev. Can. Bot. 2003, 81, 714–731. [Google Scholar] [CrossRef]
- Robinson, S.A.; Waterman, M.J. Sunsafe bryophytes: Photoprotection from excess and damaging solar radiation. In Photosynthesis in Bryophytes and Early Land Plants; Springer: Dordrecht, The Netherlands, 2013; pp. 113–130. [Google Scholar]
- Newsham, K.K.; Geissler, P.A.; Nicolson, M.J.; Peat, H.J.; Lewis-Smith, R.I. Sequential reduction of UV-B radiation in the field alters the pigmentation of an Antarctic leafy liverwort. Environ. Exp. Bot. 2005, 54, 22–32. [Google Scholar] [CrossRef]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dyn. 2015, 6, 245–265. [Google Scholar] [CrossRef]
- Bates, J.W. Effects of intermittent desiccation on nutrient economy and growth of two ecologically contrasted mosses. Ann. Bot.-Lond. 1997, 79, 299–309. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Gu, J.; Ye, Y.; Li, W.; Liao, Y.; Wang, R.; Ma, H.; Shao, X. The Diversity and Community Pattern of Liverworts on Sygera Mountain, Tibet. Forests 2024, 15, 48. https://doi.org/10.3390/f15010048
Song X, Gu J, Ye Y, Li W, Liao Y, Wang R, Ma H, Shao X. The Diversity and Community Pattern of Liverworts on Sygera Mountain, Tibet. Forests. 2024; 15(1):48. https://doi.org/10.3390/f15010048
Chicago/Turabian StyleSong, Xiaotong, Jiqi Gu, Yanhui Ye, Wei Li, Yujia Liao, Ruihong Wang, Heping Ma, and Xiaoming Shao. 2024. "The Diversity and Community Pattern of Liverworts on Sygera Mountain, Tibet" Forests 15, no. 1: 48. https://doi.org/10.3390/f15010048
APA StyleSong, X., Gu, J., Ye, Y., Li, W., Liao, Y., Wang, R., Ma, H., & Shao, X. (2024). The Diversity and Community Pattern of Liverworts on Sygera Mountain, Tibet. Forests, 15(1), 48. https://doi.org/10.3390/f15010048