1. Introduction
Invasive alien species comprise one of the consequential drivers of anthropogenic global change [
1], and the damage that newly added invasive species cause to resident native species may differ between island habitats and continental habitats [
2]. The isolated, insular features that define the evolution of island flora may lead to a lack of resistance to invasive herbivores. Additionally, specialist herbivores that invade an island may proliferate in the absence of natural enemies. For example, the cycad species from Micronesia is described as
Cycas micronesica K.D. Hill [
3], and there are no known native herbivores that consume leaves. The native
Anatrachyntis sp. Meyrick larvae consume microstrobilus tissues after the adults provide entomophilous pollination services [
4,
5]. The rapid removal of the strobilus tissue is beneficial for hastening the timing of subsequent reproductive events [
6]. The native flying fox,
Pteropus mariannus Desmarest, consumes the seed integuments [
7], which may improve seed germination and may provide a zoochory service. The native stem borer,
Acalolepta marianarum Aurivillius, exploits
C. micronesica stem cortex tissue as larval food [
5,
8]. This stem borer exhibits typical stem and bark borer behaviors by avoiding ovipositioning on healthy host trees and selectively targeting unhealthy individuals. In the absence of novel non-native stressors, these three native herbivores do not pose threats to the host tree with which they coevolved.
Non-native species that invade these Micronesian islands with a specialized affinity for consuming cycad leaf tissue may be at an advantage in the insular habitats because no pre-existing natural enemies exist and the host tree possesses no evolved resistance. The beginning of the 21st century was not a good time in this regard for
C. micronesica, as the armored scale
Aulacaspis yasumatsui Takagi, the specialist butterfly
Luthrodes pandava Horsfield, and the microlepidoptera leaf miner
Erechthias Meyrick sp. were first identified on Guam between 2003 and 2005 [
5,
8,
9]. These three herbivores exhibited rapid movement throughout the island because the host tree was the most abundant tree in Guam’s forests at the time [
10]. The newly developed residence of
A. yasumatsui in Micronesia led to secondary invasions of Rota in 2007 and the urban areas of Palau in 2008 [
11]. The islands of Yap contain
C. micronesica populations that remain free of the threats of these invasive insect herbivores.
This case study has been extensively studied. For example, interactions among the invasive insect herbivores [
5,
12,
13] and long-term changes to Guam’s
C. micronesica populations [
14,
15,
16] have been reported as consequences of the
A. yasumatsui invasion. The number of herbivores and omnivores that feed on
C. micronesica is considerable, but
A. yasumatsui is the only one that is known to be lethal when acting alone [
5]. One factor that has not been adequately reported involves how the consequential co-occurring herbivores in a subpopulation influence the plant’s initial mortality response to the
A. yasumatsui invasion. Therefore, the aims of this study were to determine how the
C. micronesica population within each of six disparate subpopulations with differing abiotic conditions and numbers of competing consumers responded immediately following the onset of
A. yasumatsui infestations. Would the addition of other consumers exert any consequential influence on the features of early host tree mortality caused by
A. yasumatsui? The answer to this question may aid conservationists by improving predictions of the timing and extent of host plant mortality as the
A. yasumatsui invasive range continues to expand to other islands.
2. Materials and Methods
Six sites on the islands of Guam and Rota were selected for the establishment of plots designed to determine changes to C. micronesica plant density and demography over a period of three years following the initial outbreak of A. yasumatsui within each site. The sites were selected to ensure disparity in abiotic factors and the number of co-occurring herbivore threats to the host tree. In Guam, site selection was further defined by geographic distance and high plant density. In Rota, site selection was further defined by the inclusion of the three disjunct areas of occupancy.
2.1. Site Descriptions
The initial
A. yasumatsui infestations ranged from May 2005 in the northern Guam site until April 2010 in the western Rota site (
Table 1). The invasive Rota population required three years to enter a western Rota habitat because the small forest fragment was located on an isolated peninsula that was separated from the remaining forested habitats by an urban area. These sites exhibited great disparity in aspect and ranged in elevation from 6 to 334 masl. Although α-diversity was similar in all of the habitats, β-diversity was dissimilar. The dominant species were not the same within the footprint of the designated plots (
Table 1).
The six sites were highly contrasting in number and identity of co-occurring consequential consumer species (
Table 2). In addition to the two invasive lepidopteran leaf herbivores and the native stem borer, some
C. micronesica habitats in Guam were impacted by non-native ungulates. These included the omnivore feral pig (
Sus scrofa L.) and the herbivore naturalized deer (
Rusa marianna Desmarest). The level of damage to plant health that each of these consumers causes has been discussed elsewhere [
5]. The disparity in ungulate damage among the Guam sites was a result of differential access by local hunters. Site 1 in Guam was located within a military installation where local hunting is highly regulated and restricted [
17], and this was the site with abundant pig and deer damage. In contrast, local hunters are allowed unrestricted access to Sites 2 and 3, and these sites never revealed ungulate damage to
C. micronesica trees. One of the six sites exhibited signs of direct plant herbivory from all five consumers, and one of the sites was devoid of all five consumers (
Table 2). The other four sites contained one to three of the co-occurring consumers.
Sites 1 (centered at 13.5541 N, 144.9376 E) and 2 (13.3376 N, 144.6683 E) were located in calcareous soils with abundant karst outcrops (Clayey-skeletal, gibbsitic, nonacid, isohyperthermic Lithic Ustorthents). Site 3 (13.2651 N, 144.7119 E) was supported with a volcanic clay soil that was poorly drained and subject to erosion (Clayey, montmorillonitic, isohyperthermic, shallow Udic Haplustolls). The littoral soils in Site 4 (14.1825 N, 145.2094 E) were sands (Loamy-skeletal, carbonatic, isohyperthermic Lithic Haplustolls). The alkaline soils in Site 5 (14.1376 N, 145.2247 E) contained fewer rock outcrops than sites 1 and 2 (Clayey-skeletal, kaolinitic, isohyperthermic Lithic Haplustolls). The soils in Site 6 (14.1272 N, 145.1262 E) exhibited chemical and physical properties that were similar to those in Sites 1, 2, and 5, although the soil series differed (Loamy, oxidic, nonacid, isohyperthermic Lithic Ustorthents). The latitudinal range of this study was approximately 100 km, and the longitudinal range was 60 km. The two closest sites were separated by 6 km. Although the sites varied in edaphic and topographic characteristics, there were no climate differences within the footprint of this study. The most influential weather factor that may have caused heterogeneous plant damage among the sites was the occurrence of tropical cyclones. There were no consequential tropical cyclones that influenced the plant population between 2005, when data collection was initiated at Site 1, and 2013, when data collection was terminated at Site 6.
2.2. Data Collection
Within each of the sites, four 20 m × 20 m plots were established when A yasumatsui had entered the habitat but before there were any signs of infestation within each plot’s boundaries. From experience, this meant A. yasumatsui would enter the plots within weeks of the initial plant survey. The distance among the four plots within each site was not fixed, and the maximum distance ranged from 830 m in the western Rota site to 470 m in the northeast Guam site. The stem count and height of every C. micronesica stem were recorded initially, and then these same measurements were repeated over a three-year period. The sequential population counts were conducted more often on Guam, but the Rota counts were never separated by more than 12 months. These methods enabled an annual assessment of plant mortality using the date of initial A. yasumatsui herbivory as the starting date for each site.
Plants with stems less than 20 cm in height were designated as newly emerged seedlings if the leaf number was four or less and the stem height was difficult to measure, and designated as saplings if the leaf number was greater than four and the stem height could be easily measured. Plants with stems between 20 and 100 cm in height were designated as juveniles, as reproductive structures are rarely observed on in situ individuals less than 100 cm in height. The adult plants were separated into 100–200 cm, 200–300 cm, and 300+ cm demographic categories. As a result, there were six plant size categories. There were 548 individuals in the initial census of Site 1, 513 individuals in Site 2, and 563 individuals in Site 3. Rota habitats contained more initial plants within the areas of occupancy than did Guam habitats. There were 1992 individuals in the initial census of Site 4, 1772 individuals in Site 5, and 1816 individuals in Site 6.
2.3. Data Analysis
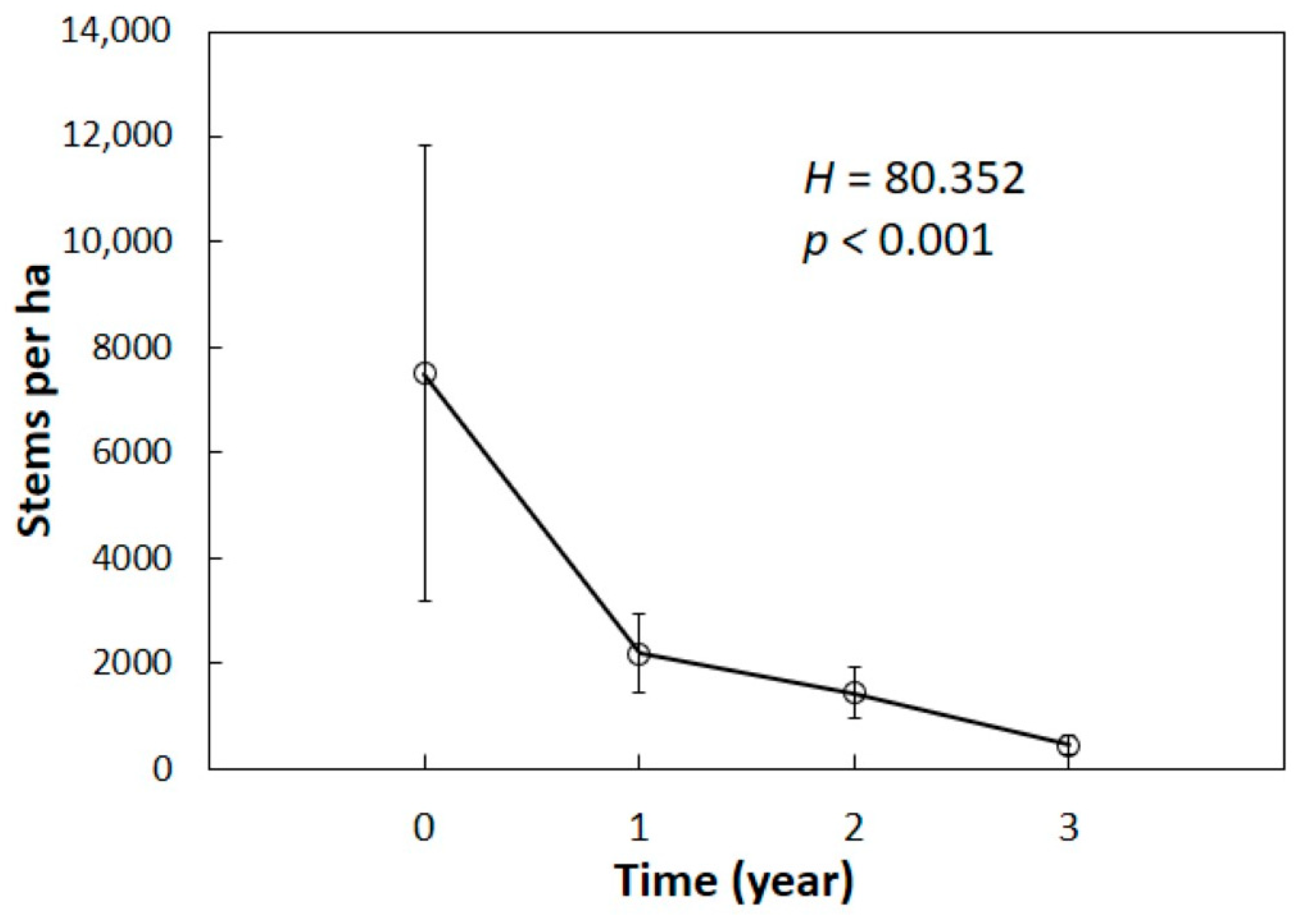
Our primary objective was to determine how annual changes in alive
C. micronesica plants caused by
A. yasumatsui herbivory differed among the sites in order to tease apart the influence of co-occurring consumers. Long-term survival of
C. micronesica has been studied in many habitats on Guam [
14]. In order to determine if our six sites behaved similarly to the sites that were previously reported, the survival of the entire population within each plot was tabulated over the four measurement periods to compare overall survival among the years. The data did not conform to parametric prerequisites, so the non-parametric Kruskal–Wallis
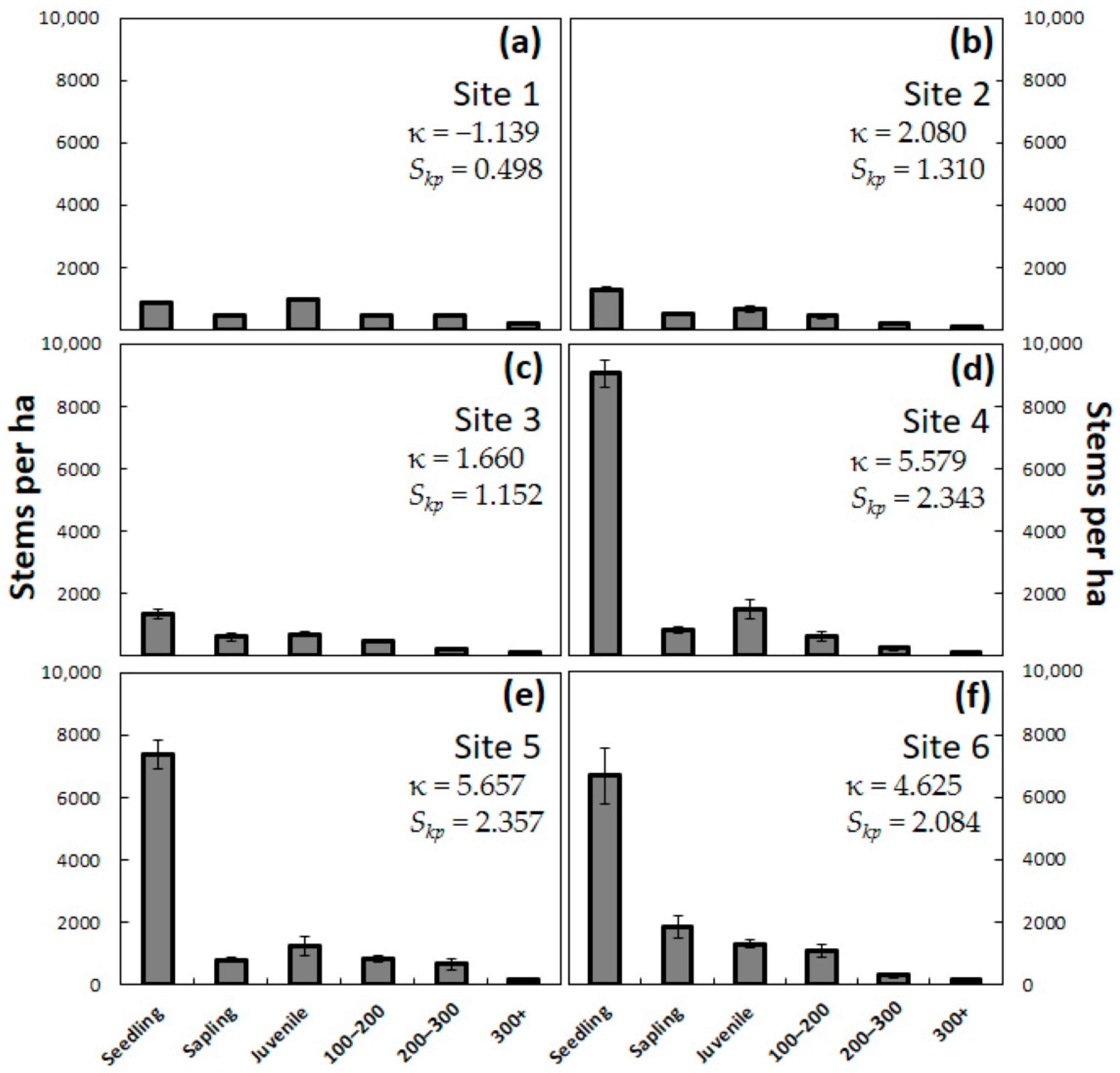
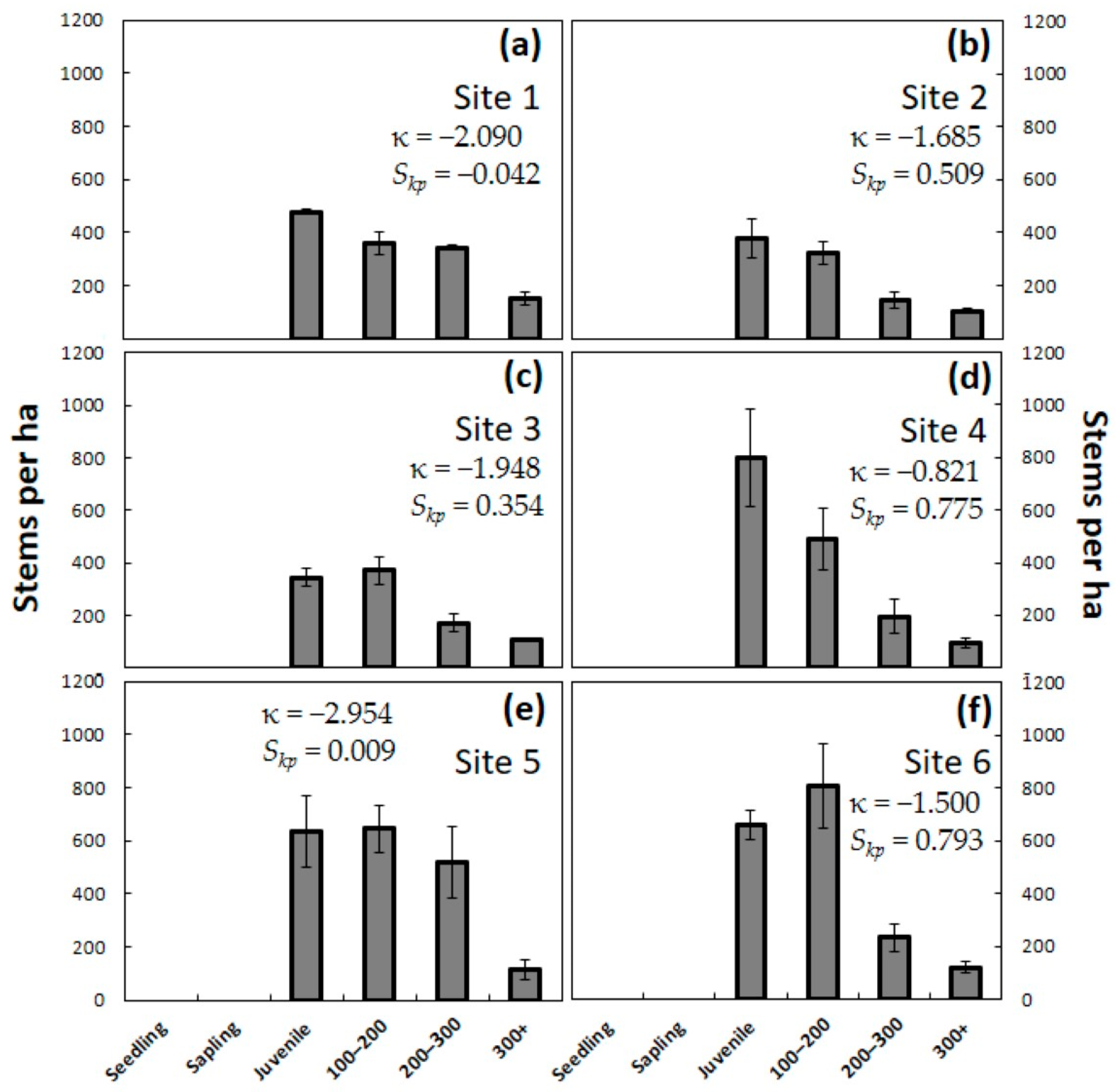
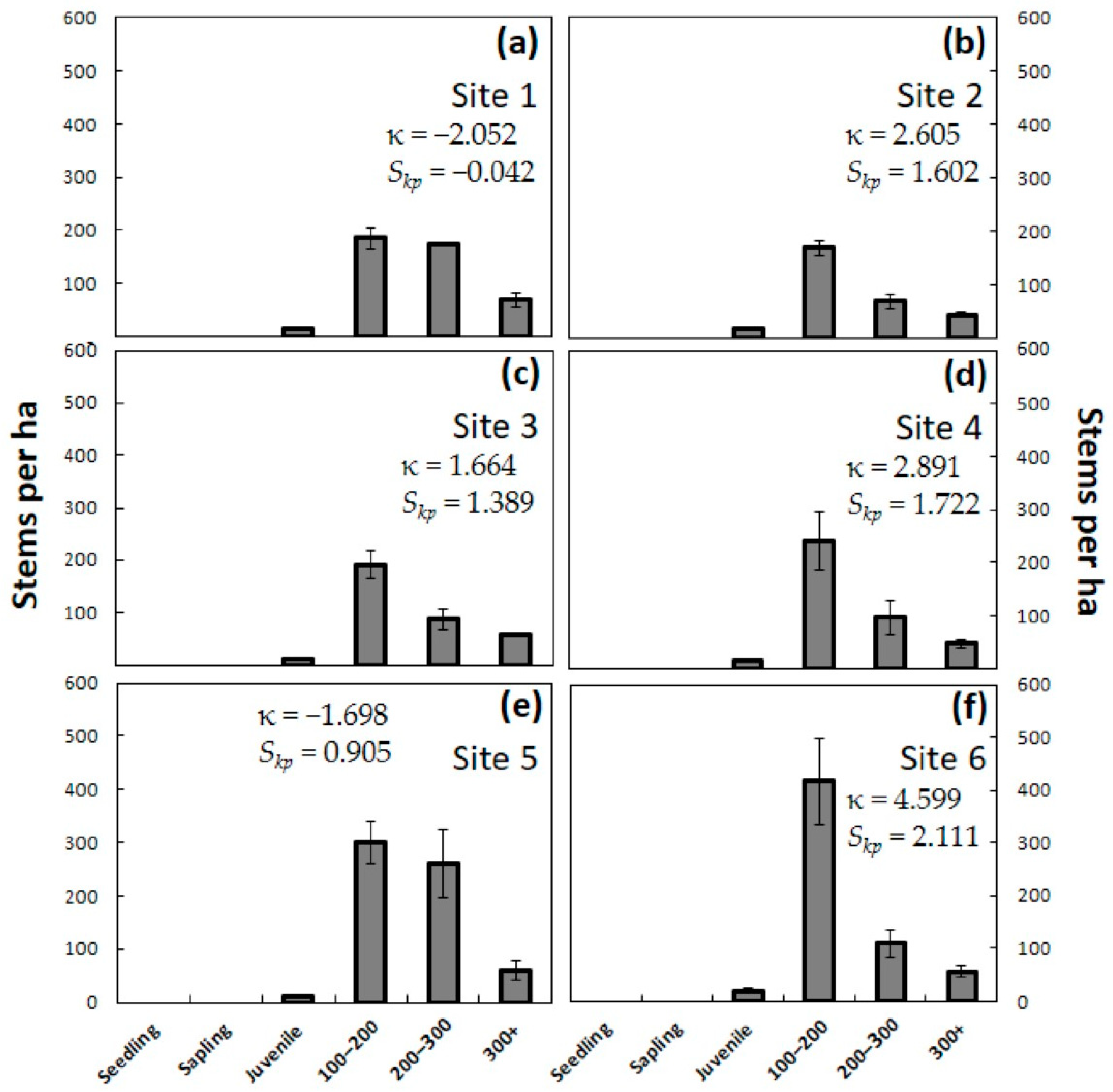
H test was used to compare the four observation years. The means were separated using a post-hoc Dunn-Bonferroni test. Second, the data from each of the years were tabulated separately within size categories to visualize the changes in population demography. Frequency distributions of plant size within each site and year were created, then kurtosis (κ) and skewness (
Skp) were calculated [
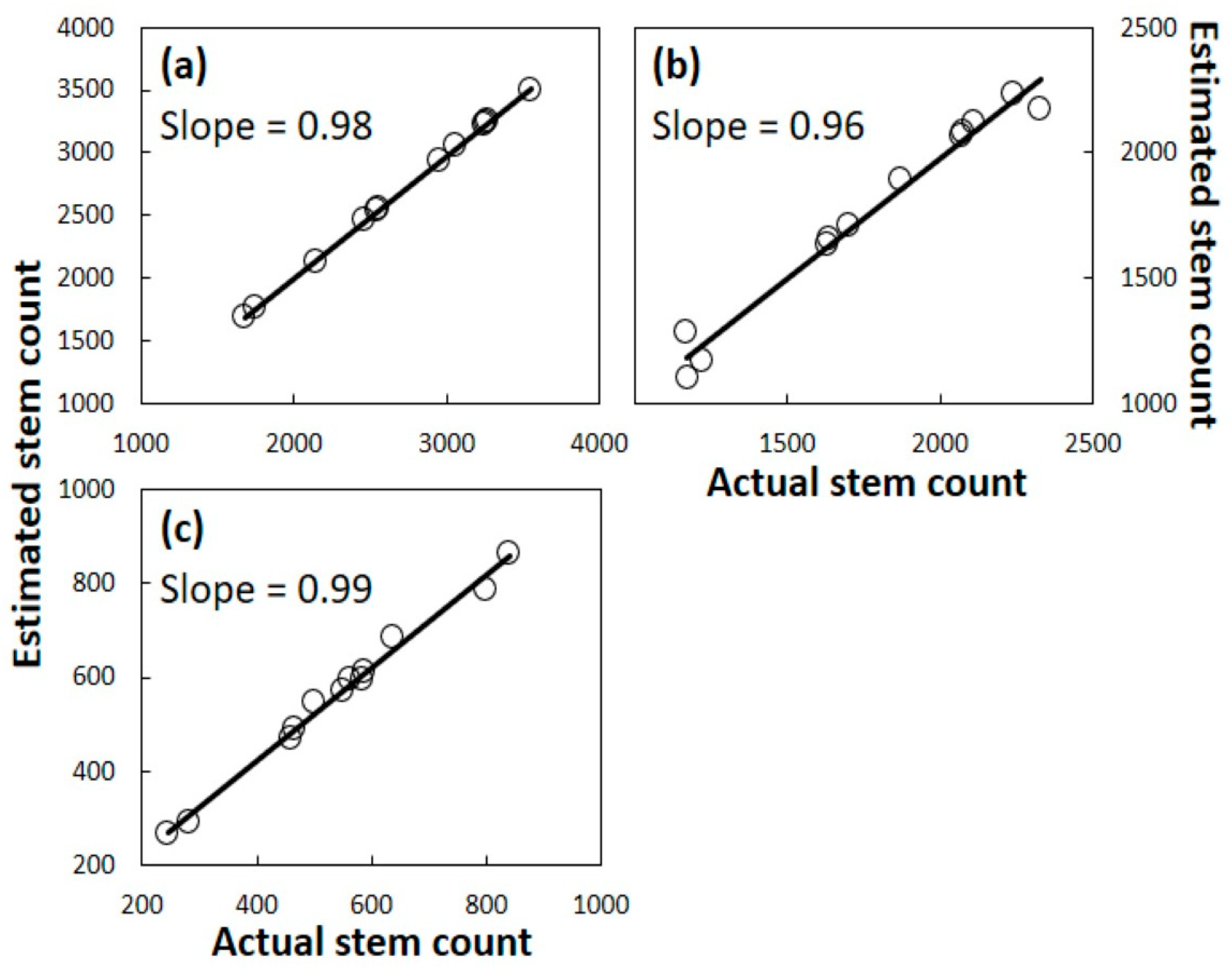
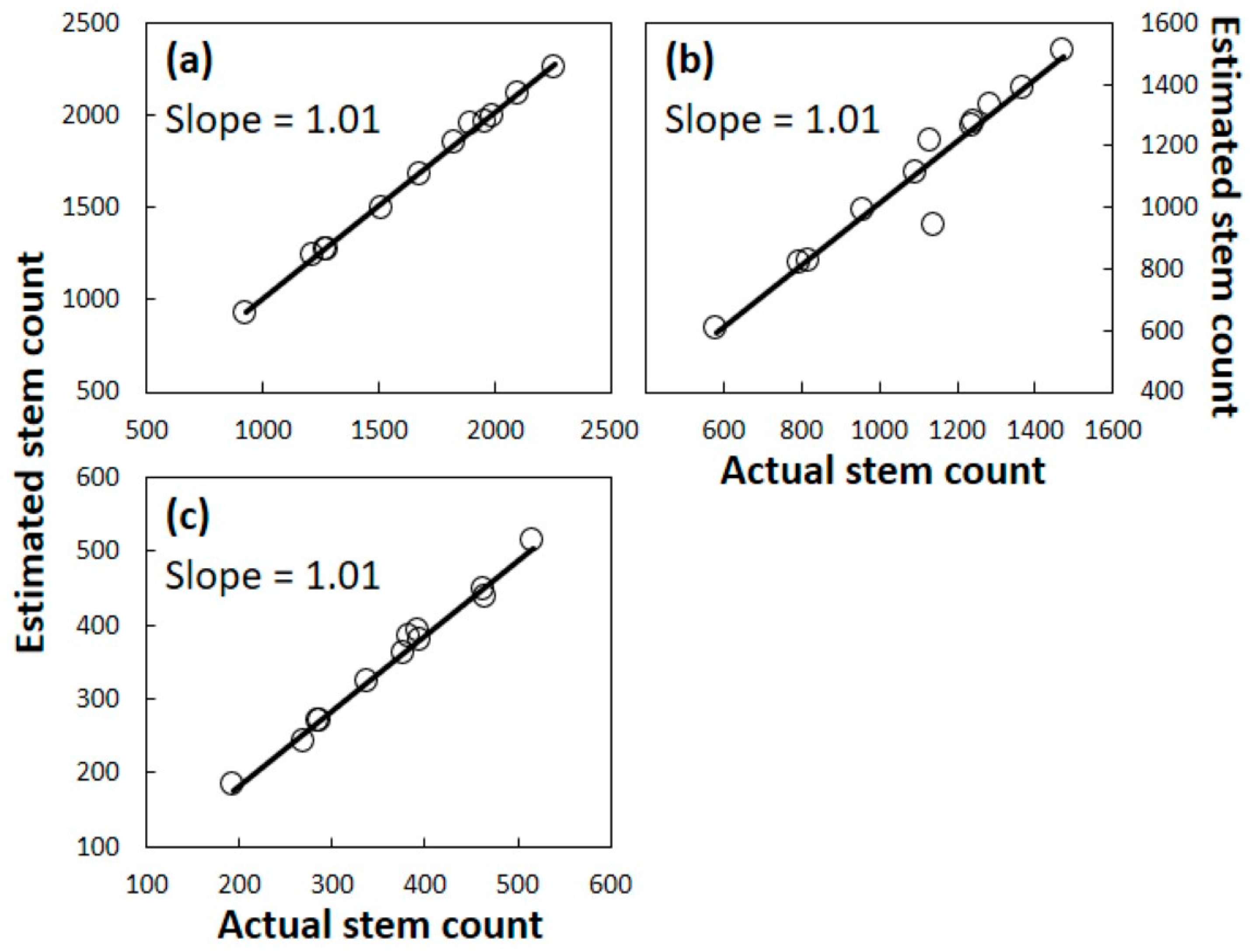
18]. Third, models were created based on coefficients that described the proportion of surviving plants within each size category at the end of each year. Models were developed from the Guam data, and these models were used with pre-invasion Rota data to generate predicted survival after one, two, or three years. Models were developed from the Rota data, and these models were used with pre-invasion Guam data to generate predicted survival after one, two, or three years. Fourth, the results from each of the individual plots were subjected to linear regression to determine significance and fit for the actual versus predicted population of surviving plants.
4. Discussion
Cycas micronesica experienced no major threats prior to 2003, when it was the most abundant tree species on Guam [
10] and was thriving as a foundation species throughout its native range. Today, this unique tree is listed as Endangered under the IUCN Red List [
19] and Threatened under the United States Endangered Species Act (ESA) [
20]. The IUCN listing was initiated in 2006, revealing the speed with which invasive specialist herbivore species can alter insular ecosystems if an abundant native plant serves as a host for the herbivore. Although long-term trends in mortality of the tree have been reported [
14,
21], this is the first study to show that the presence of other consequential consumers [
5] found on Guam and Rota did not influence the speed and demographic traits of the initial years of mortality after herbivory by
A. yasumatsui was initiated within a new subpopulation. One of our sites contained five consequential co-occurring herbivores and omnivores, and one of our sites contained no other observable consumers, yet the influence of plant size on the mortality response was homogeneous among all 24 of our plots. Indeed, the Guam model coefficients were able to predict the Rota outcomes with accuracy, and the Rota model coefficients were able to predict the Guam outcomes with accuracy.
The pre-invasion C. micronesica populations contained seedlings and saplings as the most abundant demographic category, and this entire category was eliminated immediately during the first year. By the end of year one, the combination of data from all 24 plots in this study indicated a new subpopulation responding to A. yasumatsui herbivory, which can be predicted by subjecting the pre-invasions data to the equation: Stems per ha = (seedlings × 0.000) + (saplings × 0.000) + (juveniles × 0.962) + (adults × 1.000). The second year of scale damage eliminated all individuals less than 50 cm in height and began to cull some of the adult trees. By the end of year two, the combination of data from all 24 plots in this study indicated a new subpopulation responding to A. yasumatsui herbivory, which can be predicted by the equation: Stems per ha = (seedlings × 0.000) + (saplings × 0.000) + (juveniles × 0.517) + (adults × 0.760). By the end of the third year, there were no living individuals less than 90 cm in height, and more than half of the plants greater than 90 cm had been killed. By the end of year three, the combination of data from all 24 plots in this study indicated a new subpopulation responding to A. yasumatsui herbivory, which can be predicted by the equation: Stems per ha = (seedlings × 0.000) + (saplings × 0.000) + (juveniles × 0.016) + (adults × 0.378).
Our previous long-term studies on Guam indicated that no
C. micronesica trees were less than 100 cm in height after 5 years of herbivory in one location [
21] and after 9 years in all 12 locations [
14]. Moreover, by 2020, the smallest individuals measured on Guam were 175 cm in height, indicating the elimination of every small individual by this hemipteran pest deleted at least 70 years of population-level recruitment [
22]. Even if conservation efforts are successful in beginning to mitigate the biotic threats and the tree species begins to recover in the future, this extensive recruitment gap will leave a permanent mark on the demographic traits of the tree populations into the future. This permanent mark will be at least 2 m in vertical stem length and will persist for centuries into the future. Heeding the early advice to construct a coalition of biological control organisms [
23,
24] would not have allowed these permanent changes to demography to occur.
One profound confirmation for the assertion that A. yasumatsui is the most influential threat to C. micronesica is illuminated by looking more closely at the comparison of Site 1 and Site 6. Site 1 contained five co-occurring herbivores or omnivores, exhibited the least healthy pre-invasion population, and became infested in May 2005. In contrast, Site 6 was herbivore-free when A. yasumatsui initially infested the forest fragment in April 2010. While both sites experienced the elimination of seedlings, saplings, and most juveniles, 34% of the adults in Site 1 but only 31% of the adults in Site 6 were persisting after three years of damage by the armored scale herbivore. Clearly, the absence of co-occurring consumers in Site 6 did not offer any benefits to the host tree population when it was forced to respond to the first three years of the A. yasumatsui invasion.
4.1. Practical Applications
The utility of this new knowledge may be profound for predicting how a newly invaded C. micronesica subpopulation will respond for three years. Indeed, the predicted number of plant deaths based on pre-invasion subpopulation data in each of the 24 plots was remarkably consistent with the actual plant deaths over the timeline of this study. Therefore, conservationists may obtain pre-invasion density and demography data that can be used to accurately predict the dynamics of population changes during the first three years of A. yasumatsui herbivory. By the end of three years of A. yasumatsui herbivory, a once-thriving C. micronesica population is predicted to consist of about 2% of the initial juvenile population and about 38% of the initial adult population.
Over the course of the first two years, new seedlings are predicted to germinate from the pre-existing seed bank and from the dispersal of seeds from megastrobili that were close to maturity when A. yasumatsui herbivory was initiated. These seedlings are predicted to be killed shortly after germination, with no recruitment to the sapling stage. No newly germinated seedlings will be found after year two. Megastrobili that develop after the A. yasumatsui invasion are predicted to contain undeveloped ovules at the end of three years, but no developing seeds will reach maturity in the C. micronesica habitats at this stage.
4.2. The Other Consumers
Site 1 contained the least number and proportion of seedlings and saplings among the three Guam sites, even though the observed number of fecund female trees and seeds in the standing trees was no less at this site than at the other sites. Signs of soil disturbance due to pig rooting were widespread. These observations indicated that the relatively low seedling and sapling counts were likely due to the rooting and seed herbivory of this feral omnivore.
The pre-invasion data indicated that Site 6 exhibited the greatest evidence of recruitment from seedling to sapling stage among the Rota sites. This was the only site that did not contain L. pandava herbivory. These observations indicated that the decreased recruitment of seedlings to saplings in Rota Sites 4 and 5 was likely due to the long-term pressures of butterfly herbivory.
When a foundation tree species is selectively removed from an island’s forests, the conservation community is expected to understand the possible influences on other native organisms that rely on the tree species in order to consider the potential for coextinction. The microlepidoptera mutualist pollinators are the most important native animals to discuss in this regard. Our observations indicate that the Mariana, Western Caroline, and Palau islands contain three different taxa of pollinators. Moreover, we do not know of an alternate larval food that augments C. micronesica microstrobilus tissue herbivory. Therefore, the pollinator taxon from the Mariana Islands of Guam and Rota appears to be a two-island endemic species, and these native animals have been and remain threatened by the loss of C. micronesica trees.
The native flying fox,
P. mariannus, is a frugivore and may rely on
C. micronesica seed availability during seasons when other preferred fruits are difficult to find [
7]. The invasion of
A. yasumatsui has threatened this source of food for the flying fox communities in Guam and Rota [
25].
The specialist butterfly,
L. pandava, appears to be in direct competition with
A. yasumatsui. In Guam, ephemeral irruptions of
A. yasumatsui were accompanied by decreases in
L. pandava herbivory, and periods of decreased
A. yasumatsui infestations were accompanied by increases in
L. pandava herbivory [
12]. The incidence of
A. yasumatsui on Guam has declined in recent years for unknown reasons [
26], and this development may generate an increase in butterfly herbivory in the near future.
The native stem borer
A. marianarum poses a secondary threat to
C. micronesica because stem borer damage increases after any abiotic or biotic stress that reduces the health of the host tree, and therefore irruptions of the non-native
A. yasumatsui are followed by irruptions of the native
A. marianarum [
12]. Subsequently, the incidence of windsnap during tropical cyclones is increased by the
A. marianarum stem herbivory [
26,
27], illuminating how the invasion of
A. yasumatusi has led to cascading negative interactive outcomes. This case study reveals a conservation conundrum because the native beetle is no less deserving of conservation than the host tree,
C. micronesica.
Within the context of population dynamics and recruitment behaviors of
Anatrachyntis sp.,
P. mariannus, and
A. marianarum, we view
C. micronesica as a keystone species [
28]. The immense population loss and the less frequent production of strobili among the living trees have undoubtedly decimated the services that this native tree species provides for these native animals. In addition to fostering the recovery of regeneration and recruitment of
C. micronesica, conservationists may also need to study and foster re-entanglement of the food webs that have been disrupted. We believe the
Anatrachyntis populations may be at greater risk than any other involved taxon, including
C. micronesica, because the Guam and Rota pollinator species likely occur nowhere else worldwide.
4.3. Biological Control in the Study Sites
Ongoing work within the native range of the armored scale in Thailand has revealed that
C. micronesica trees are not threatened by chronic infestations of
A. yasumatsui, and the ex situ plant populations are healthy and thriving as a result of widespread native biological control organisms that maintain the
A. yasumatsui density below damaging levels [
28]. These observations support the contention that the establishment of a biological control program on Guam and Rota would adequately mitigate the threats to
C. micronesica that are imposed by
A. yasumatsui [
26,
27].
The armored scale predator
Rhyzobius lophanthae Blaisdell was intentionally released in Guam in 2005 and in Rota in 2007 [
5]. The predator entered each of our six research sites shortly after
A. yasumatsui entered each subpopulation, either naturally or by anthropogenic distribution and release. Therefore, the initial plant mortality in each plot occurred in the presence of this scale predator. Ongoing plant mortality has not ceased, and the predator is clearly not sufficient to fully mitigate the lethal threat caused by the armored scale. The limitations of
R. lophanthae have been discussed elsewhere [
5,
11,
28].
Attempts to introduce the parasitoids
Aphytis lingnanensis Compere and
Coccobius fulvus Compere & Annecke to Guam were unsuccessful [
5]. However, the fortuitous presence of the parasitoid
Arrhenophagus chionaspidis Aurivillius was first noticed on Guam in 2013 [
5]. The Guam portion of the present study was terminated before this date, so there was no parasitoid biocontrol of
A. yasumatsui within any of our six study sites.
4.4. Species Conservation
When a cornucopia of anthropogenic, abiotic, and biotic threats coalesce to endanger a tree species, developing a conservation action plan to slow down plant mortality may require a triage approach where the single greatest threat is identified and then countered with the greatest level of conservation efforts. For this approach to be successful, knowledge about each of the threats needs to be generated through observation and experimentation, and then those facts need to be communicated to the empowered decision-makers who control funding and programming of conservation efforts. In order to enable the most effective conservation actions, this communication best occurs through peer-reviewed journal publications that are filtered through the expert vetting process. Therefore, developing a conservation action plan to mitigate the threats and a recovery plan to rebuild the plant populations depends on bringing knowledgeable research biologists into the agenda, as these are the contributors who possess the wherewithal to compile and interpret the copious information from myriad directions.
What is the primary threat to
C. micronesica? All evidence to date indicates that land conversion is not a consequential threat. The loss of
C. micronesica trees due to construction activities has not occurred to a great extent on Rota, Yap, or Palau and is restricted to military lands on Guam. The forests that have experienced the loss of
C. micronesica trees within federal construction sites contain numerous trees in the habitats adjacent to the construction sites, and in situ conservation of these trees constitutes the most effective mitigation endeavor to counteract the consequence of land conversion [
26]. The other native and non-native consumers that have been discussed are also not primary threats to the tree species; they are secondary threats. The consequential historical
C. micronesica plant mortality since 2003 has been a result of
A. yasumatsui herbivory. The threat is manifested as direct threats caused by carbohydrate depletion that leads to individual plant mortality following
A. yasumatsui damage [
29], but there are also cascading threats that only emerge because of antecedent
A. yasumatsui herbivory. These include lethal damage during and after tropical cyclones [
26,
27,
30] and herbivory from other animals that exploit the increased vulnerability of the unhealthy trees [
5,
12].
The results herein provide more evidence to support the contention that A. yasumatsui alone persists as the greatest single threat to the host tree. A predictive equation based on Guam data calculated with accuracy the mortality from the three Rota subpopulations. Similarly, the predictive equations based on Rota data calculated with accuracy the mortality from the three Guam subpopulations. These outcomes occurred even though the number of co-occurring consumers and abiotic conditions varied greatly among the sites.
This new information is valuable for two reasons. First, the development of an adequate biological control program that manages
A. yasumatsui below lethal densities would enable the recovery of the
C. micronesica population [
26]. Second, the focus of emergency conservation funding and planning to ensure the host tree species can persist into the future can justifiably ignore the other co-occurring threats, such as salvage from construction sites, to fully focus all resources on biological control of the armored scale. These secondary threats would inadvertently decline without any management if the primary threat were adequately addressed. Therefore, contemporary conservation projects designed to translocate trees from construction sites unjustifiably consume available financial resources and do not contribute to the actionable conservation of
C. micronesica. These large-scale tree transplanting and propagation projects have consumed millions of dollars in attempts to rescue trees from federal construction sites on the island of Guam. Our results indicate that these expensive projects are ill-informed and will not generate any benefit to species conservation because the primary threat is not being addressed in any capacity.
Establishing a coalition of specialist biological control organisms to maintain the
A. yasumatsui herbivory below lethal levels was the greatest conservation need in 2003 on Guam and in 2007 on Rota. This conservation action was the most important need when the species was Red-listed in 2006 [
19] and ESA-listed in 2015 [
20]. In order to address the persistence of
C. micronesica into the future, this endeavor, which was recommended by the scientific community in 2005 [
23,
24] remains the greatest conservation need today. Invasive species impose an enormous cost on society [
31], and group apathy toward the control of invasive species cannot be good for society. This is essentially what describes the past two decades in Guam. These types of difficult conservation needs define many biodiversity threats that are global in nature and are sometimes described as wicked problems [
32,
33,
34,
35]. While the management of biological control programs has been implemented for more than a century [
36], many case studies are characterized by difficult hurdles [
37]. Turning the corner toward success within these case studies cannot occur without collaborations among all stakeholders, including the scientists who possess the knowledge to inform each decision within information-deficit conditions [
36,
38]. Being successful in solving the wicked problems of conservation requires a change in the manner in which decision-makers interact with the problems. The Guam and Rota invasive species control needs demand a multi-year, multi-discipline approach [
28].
4.5. Island Comparisons
There are three island groups that comprise the indigenous range of
C. micronesica. The Mariana Island of Guam stands alone in containing vast areas of occupancy that stretch for tens of kilometers and traverse numerous soil and habitat types. The Mariana island of Rota and the Caroline island of Yap are similar in that three disjunct areas of occupancy make up three subpopulations. The plant density within these subpopulations is immense in Yap and was immense in Rota prior to the 2007
A. yasumatsui invasion. The independent state of Palau contains numerous island subpopulations, but all are limited in plant number and geographic range. Land conversion within any of the conscribed subpopulations in Rota, Yap, and Palau would be devastating to the global genetic diversity of the tree species. But land conversion in Guam is inconsequential to species survival because so many trees persist in the same habitats adjacent to the construction sites that are causing the land conversion. In situ conservation of these populations adjacent to the construction sites would benefit species survival without the expense and added health threat that is required to excavate, propagate, and translocate from the construction sites [
39]. The greatest conservation needs for Guam and Rota are to obey the 2005 recommendations by scientists [
23,
24] and ensure biocontrol becomes a reality. The greatest conservation needs for Yap and the isolated islands of Palau are to implement appropriate hygiene protocols for human travelers to ensure
A. yasumatsui is not vectored into the uninfested islands.
The data herein are the first
C. micronesica density data to be reported from the island of Rota. There were three known areas of occupancy at the time of the
A. yasumatsui invasion. The plant density within these Rota habitats was extensive, with more than 1000 adults per ha. These same densities were reported from the Yap subpopulations [
14]. If the conservation deciders allow local extirpation of any of the six Rota or Yap subpopulations, the result will be extensive genetic erosion that cannot be recovered.
The unusually high absolute and proportional numbers of pre-invasion juvenile plants in northeast Guam (Site 1) were also reported from a northwest Guam location [
21]. These two northern coastal habitats were positioned in the terrain that received massive environmental destruction from the bombing raids imposed by the United States military when Guam was recaptured following the Japanese occupation of WWII. This bombardment began during the first week of May 1944 and reached its greatest intensity in July 1944 [
40]. Therefore, aircraft and battleship firepower destroyed Guam’s terrain, demolished civilian dwellings, and killed civilian residents for more than two months in order to lessen military casualties when United States troops began to land on 21 July 1944. This environmental destruction from months of conventional wartime firepower occurred because the United States enabled the Japanese occupation by abandoning the legal responsibility to defend Guam in 1941 [
41]. In this study, Site 1 exhibited a number of juvenile plants, which was much greater in proportion to the adult tree categories than at any other site. These extreme numbers of juvenile plants may illuminate a lingering demographic consequence of the destruction of the forest resources 60 years prior and the permanent mark on
C. micronesica subpopulations. Sites 2 and 3 in Guam were located in isolated forests in remote areas that were not likely targeted by the bombing of 1944. Similarly, the islands of Rota, Yap, and Palau did not receive military bombardment during WWII.
4.6. Lessons Learned
Our results directly inform the conservation decisions that are needed in Guam and Rota, with several applications previously discussed. But the cumulative actionable research in this case study highlights lessons that may have regional or global application.
Models applicable throughout the genus range. The genus
Cycas is the most speciose and widespread genus of cycad [
42], with numerous insular subpopulations throughout this range. This entire range remains at risk from
A. yasumatsui. This armored scale has invaded the Ryukyu Archipelago in recent months [
43], and these islands comprise the endemic range of
Cycas revoluta Thunb. Therefore, the Yap and Palau subpopulations of
C. micronesica and the uninvaded subpopulations of
C. revoluta in the Ryukyu islands exhibit the greatest risk of future
A. yasumatsui invasion. Improving our ability to predict future outcomes in conservation science is an emerging challenge for the global conservation community [
44]. We believe our predictive equations from Guam and Rota will accurately define the plant response in all of these uninvaded subpopulations if an invasion occurs. Local scientists will be required to collect plant density and demography data prior to any plant mortality in order to capitalize on this predictive power.
Benchmarking. Our data illuminate the importance of securing population-level descriptors prior to the onset of damage by an acute threat in order to provide future conservationists with a benchmark or baseline foundation. When apathy and procrastination allow nascent threats to impose large-scale population changes before the conservation community decides to fund and conduct actionable research, subsequent species recovery goals are devoid of the requisite benchmark data that informs those goals. In the case of a
Cycas subpopulation threatened by
A. yasumatsui, a delay of only a few months will cause the collection of accurate benchmark data to be impossible. For example, four of our sites exhibited leptokurtic kurtosis before
A. yasumatsui damage, then exhibited platykurtic kurtosis only 12 months after the damage ensued. Our results show that a delay of only 3 years will result in the failure to document 90% of the pre-existing population. How can one restore a plant population if one does not know the restoration goal as defined by pre-threat descriptors? This issue is at the core of the unsuccessful funded projects in Guam. The funding agency has employed a revolving door policy for the biologists empowered with managing the sequential projects, as none of them possess direct cycad biology education and all are replaced before they have enough time to develop appropriate knowledge. Newly hired practitioners who possess no verifiable cycad biology knowledge are allowed to spend millions of dollars. Parachute scientists [
45] from United States universities are flown into Guam to guide decisions about tropical island invasive species despite never living and working within the native range of
C. micronesica. The incomprehension leads to an inability to draw on the awareness of the physiognomy and gestalt appearance of the forests in which a pre-scale cycad plant thrived. These contemporary contributors, who are collectively spending more public funds on cycad conservation than any other humans worldwide, have no first-hand knowledge about how the habitats looked prior to the onset of the threats. This is a large part of why the funded activities have been unsuccessful.
Balancing current knowledge with uncertainties. Almost 40% of known vascular plants are currently threatened with extinction [
46]. One of the challenges in conservation decision-making is how to manage the interactions between what is known from the literature and what is not known [
47]. This
C. micronesica case study has emerged as an unsuccessful example in this regard, as members of the International Union for Conservation of Nature’s Species Survival Council with international cycad conservation knowledge have been actively marginalized by empowered administrators, and actionable knowledge from the peer-reviewed literature has been actively ignored by funded practitioners [
26]. The consequence has been ongoing plant mortality despite the millions of dollars invested in ill-conceived conservation actions, resulting in a federal lawsuit in the District Court of Guam designed to stop the ongoing ecocide [
48]. The funded conservation actions to conserve
Lodoicea maldivica Gmelin in the Seychelles provide a successful example that is in stark contrast to the approach that has been implemented in Guam. The empowered conservation deciders in this developing nation initiated conservation actions by first funding scientists to determine the bottlenecks and second by developing action plans based on the new information that was published in peer-reviewed journals [
49,
50]. These unsuccessful examples in Guam and successful examples in the Seychelles highlight the importance of establishing a management team that values independent review and fosters the synthesis of ongoing monitoring and research to achieve tangible conservation outcomes. The development of effective species conservation plans will require embracing the best available information and combining it with collaborative development among all stakeholders [
51].
Long-term monitoring. Finally, our study provides an explicit example of the value of long-term monitoring of permanent plots. Relying on permanent plots to monitor long-term changes in tropical forests is a key theme in global change forest science [
52]. Permanent plots have been exploited to learn many
C. micronesica lessons from Guam, Rota, and Yap [
14,
21]. However, the value of using repeated measurements can be diminished if the observations are not sustained for an appropriate length of time. Indeed, rates of mortality at 5 years after perennial plant translocation were not similar to rates of mortality in the same sites at 15 years after translocation [
53]. Monitoring to document the success of plant translocations should extend beyond documenting translocated plant survival and should continue until evidence of regeneration and recruitment is verified at the recipient site [
54,
55]. One of the reasons for information deficits in the translocation ecology literature is that unsuccessful outcomes are purposefully not reported [
53]. Short-term observations of only 1 year have been used in Guam to declare the success of translocation projects [
48]. These methods are indefensible, and taxpayers who fund conservation actions in every country deserve to know the success rate of plant translocations at 5, 10, and 15 years. Based on our decades of Guam-based research, we believe the use of an appropriate duration of monitoring will reveal that these recent translocation projects that were deemed successful have already reached 100% mortality.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}