
Elevational Patterns of Tree Species Richness and Forest Biomass on Two Subtropical Mountains in China

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
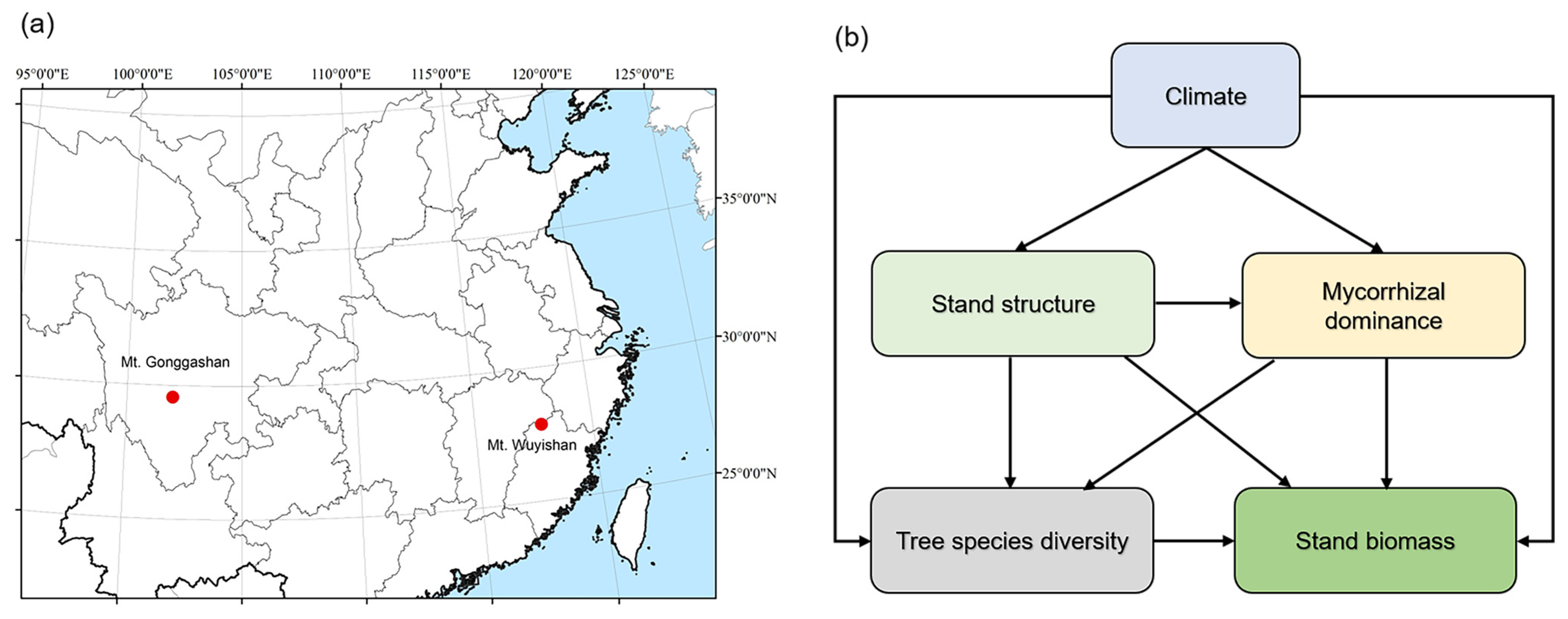
2.1. Study Sites and Field Sampling
2.2. Species Richness, Biomass, and Stand Structural Variables
2.3. Mycorrhizal Information
2.4. Climate Data
2.5. Statistical Analyses
3. Results
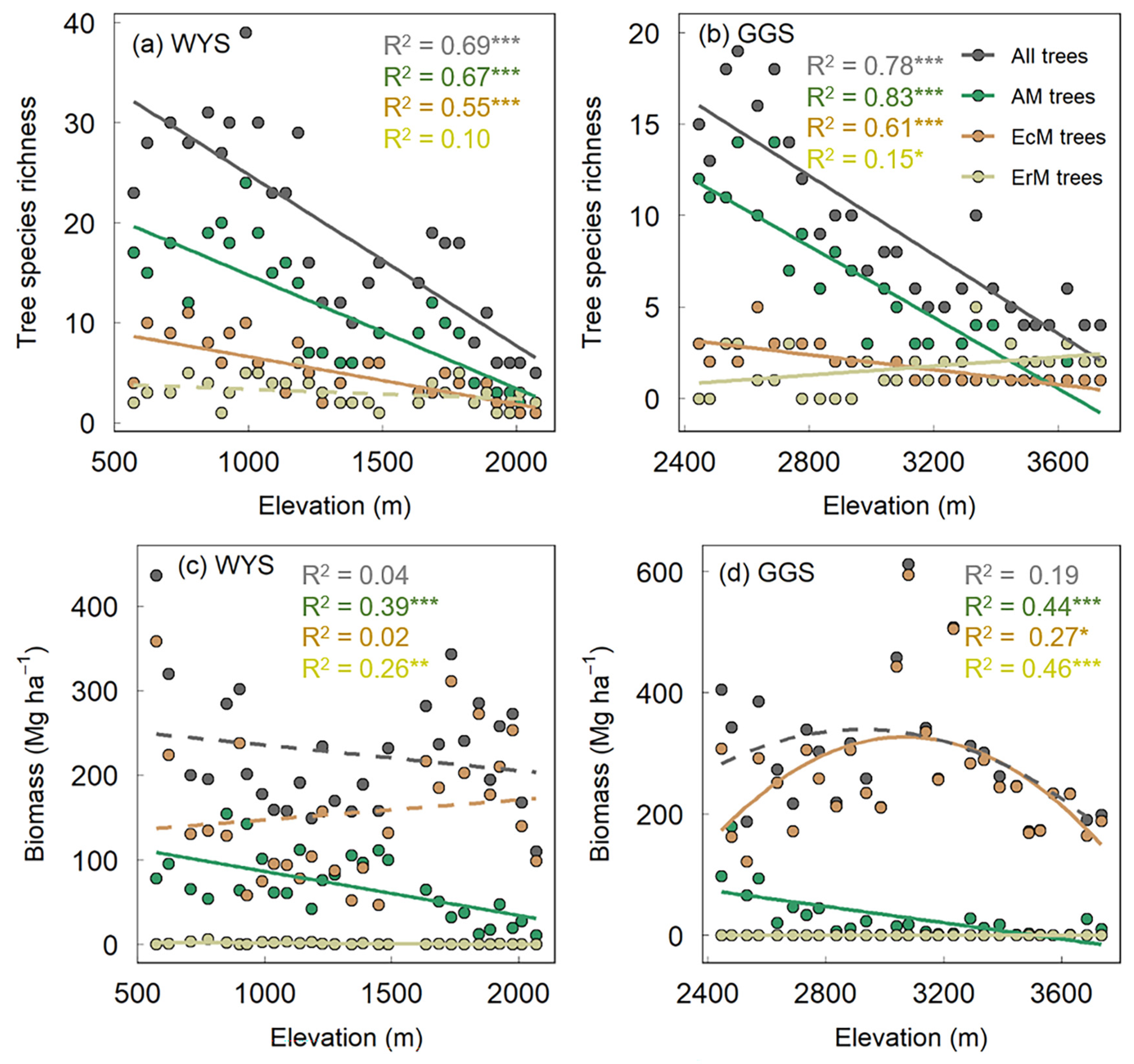
3.1. Elevational Patterns of Tree Species Richness and Biomass
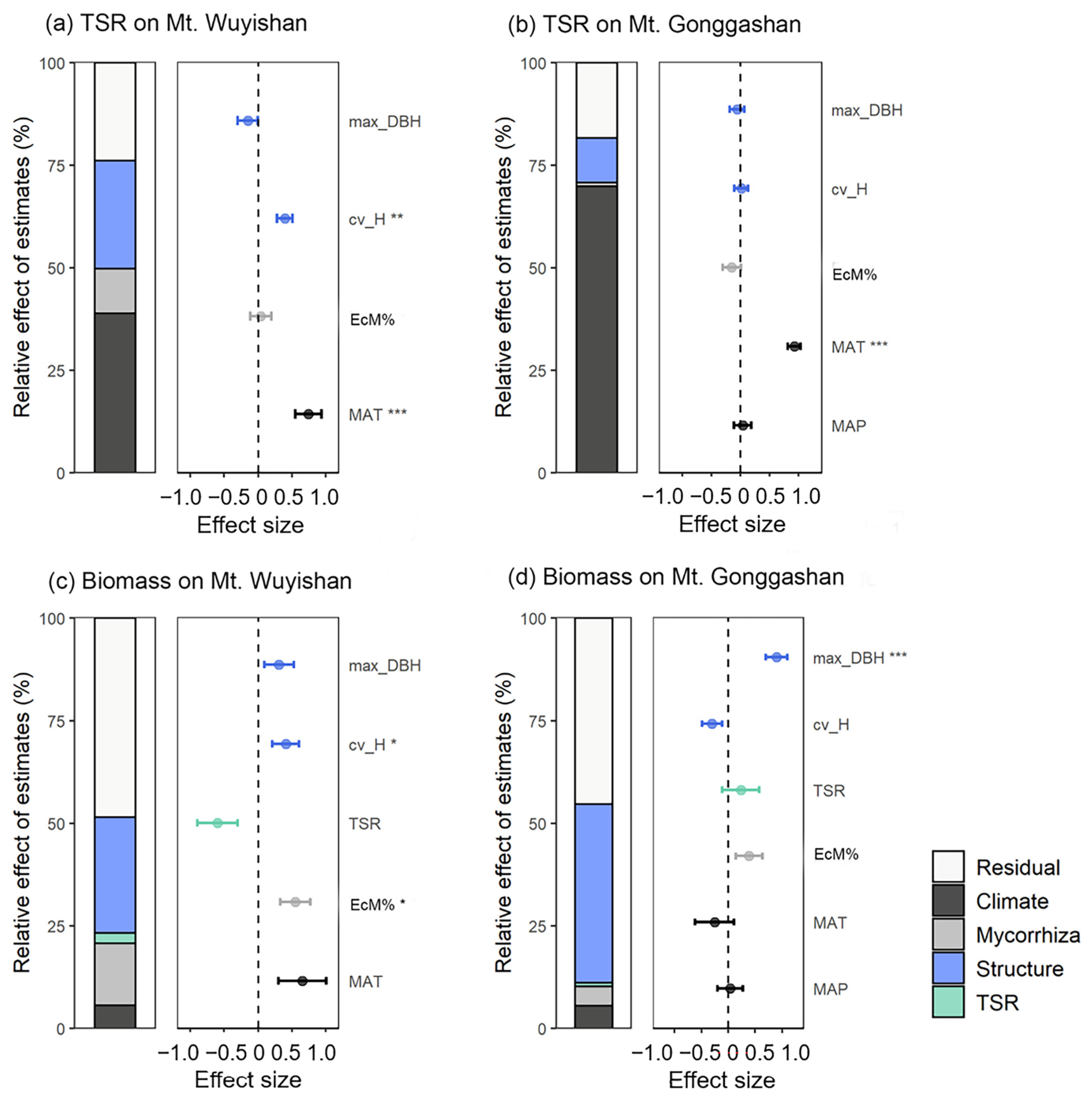
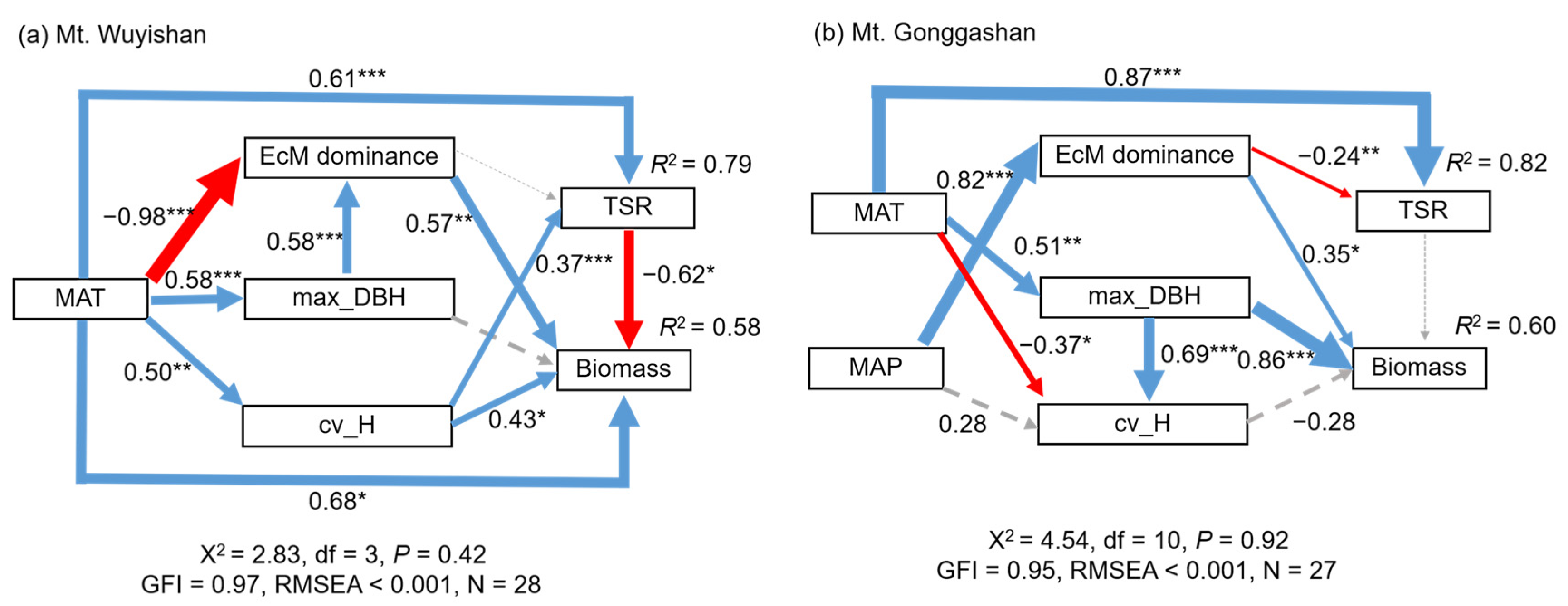
3.2. Possible Drivers of Tree Species Richness and Biomass
4. Discussion
4.1. Temperature Controls Tree Species Richness
4.2. Forest Biomass Was Greatly Driven by Stand Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Davies, S.J.; Abiem, I.; Abu Salim, K.; Aguilar, S.; Allen, D.; Alonso, A.; Anderson-Teixeira, K.; Andrade, A.; Arellano, G.; Ashton, P.S.; et al. Forestgeo: Understanding Forest Diversity and Dynamics through a Global Observatory Network. Biol. Conserv. 2021, 253, 108907. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive Biodiversity-Productivity Relationship Predominant in Global Forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Li, Y.; Härdtle, W.; von Oheimb, G.; Yang, X.; et al. Impacts of Species Richness on Productivity in a Large-Scale Subtropical Forest Experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Schmid, B.; Loreau, M.; Forrester, D.I.; Fei, S.; Zhu, J.; Tang, Z.; Zhu, J.; Hong, P.; Ji, C.; et al. Multispecies Forest Plantations Outyield Monocultures across a Broad Range of Conditions. Science 2022, 376, 865–868. [Google Scholar] [CrossRef]
- Mao, Z.; van der Plas, F.; Corrales, A.; Anderson-Teixeira, K.J.; Bourg, N.A.; Chu, C.; Hao, Z.; Jin, G.; Lian, J.; Lin, F.; et al. Scale-Dependent Diversity–Biomass Relationships Can Be Driven by Tree Mycorrhizal Association and Soil Fertility. Ecol. Monogr. 2023, 93, e1568. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.; Coomes, D. Individual Size Inequality Links Forest Diversity and Above-ground Biomass. J. Ecol. 2015, 103, 1245–1252. [Google Scholar] [CrossRef]
- Ammer, C. Diversity and Forest Productivity in a Changing Climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef]
- Ullah, F.; Gilani, H.; Sanaei, A.; Hussain, K.; Ali, A. Stand Structure Determines Aboveground Biomass across Temperate Forest Types and Species Mixture Along a Local-Scale Elevational Gradient. Forest Ecol. Manag. 2021, 486, 118984. [Google Scholar] [CrossRef]
- Yan, G.; Bongers, F.J.; Trogisch, S.; Li, Y.; Chen, G.; Yan, H.; Deng, X.; Ma, K.; Liu, X. Climate and Mycorrhizae Mediate the Relationship of Tree Species Diversity and Carbon Stocks in Subtropical Forests. J. Ecol. 2022, 110, 2462–2474. [Google Scholar] [CrossRef]
- Wang, Z.; Brown, J.H.; Tang, Z.; Fang, J. Temperature Dependence, Spatial Scale, and Tree Species Diversity in Eastern Asia and North America. Proc. Natl. Acad. Sci. USA 2009, 106, 13388–13392. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The Effect of Biodiversity on Tree Productivity: From Temperate to Boreal Forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Fang, J.; Shen, Z.; Tang, Z.; Wang, X.; Wang, Z.; Feng, J.; Liu, Y.; Qiao, X.; Wu, X.; Zheng, C. Forest Community Survey and the Structural Characteristics of Forests in China. Ecography 2012, 35, 1059–1071. [Google Scholar] [CrossRef]
- Chu, C.; Lutz, J.A.; Král, K.; Vrška, T.; Yin, X.; Myers, J.A.; Abiem, I.; Alonso, A.; Bourg, N.; Burslem, D.F.R.P.; et al. Direct and Indirect Effects of Climate on Richness Drive the Latitudinal Diversity Gradient in Forest Trees. Ecol. Lett. 2018, 22, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Raich, J.W.; Russell, A.E.; Kitayama, K.; Parton, W.J.; Vitousek, P.M. Temperature Influences Carbon Accumulation in Moist Tropical Forests. Ecology 2006, 87, 76–87. [Google Scholar] [CrossRef]
- Keith, H.; Mackey, B.G.; Lindenmayer, D.B. Re-evaluation of Forest Biomass Carbon Stocks and Lessons from the World’s Most Carbon-dense Forests. Proc. Natl. Acad. Sci. USA 2009, 106, 11635–11640. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.C.; Swenson, N.G.; Enquist, B.J.; White, E.P.; Phillips, O.L.; Jørgensen, P.M.; Weiser, M.D.; Monteagudo Mendoza, A.; Núñez Vargas, P. Variation in Above-ground Forest Biomass across Broad Climatic Gradients. Global Ecol. Biogeogr. 2011, 20, 744–754. [Google Scholar] [CrossRef]
- Chen, G.; Cai, Q.; Ma, S.; Feng, Y.; Fang, W.; Ji, C.; Zhu, J.; Wang, Z.; Wang, S.; Tang, Z.; et al. Climate and Forest Attributes Influence Above-ground Biomass of Deciduous Broadleaf Forests in China. J. Ecol. 2022, 111, 495–508. [Google Scholar] [CrossRef]
- Abbasi, U.A.; Mattsson, E.; Nissanka, S.P.; Ali, A. Species α-diversity Promotes but β-diversity Restricts Aboveground Biomass in Tropical Forests, Depending on Stand Structure and Environmental Factors. J. For. Res. 2022, 1–13. [Google Scholar] [CrossRef]
- Ali, A. Forest Stand Structure and Functioning: Current Knowledge and Future Challenges. Ecol. Indic. 2019, 98, 665–677. [Google Scholar] [CrossRef]
- Aponte, C.; Kasel, S.; Nitschke, C.R.; Tanase, M.A.; Vickers, H.; Parker, L.; Fedrigo, M.; Kohout, M.; Ruiz-Benito, P.; Zavala, M.A.; et al. Structural Diversity Underpins Carbon Storage in Australian Temperate Forests. Global Ecol. Biogeogr. 2020, 29, 789–802. [Google Scholar] [CrossRef]
- Wen, Z.; Jiang, Z.; Zheng, H.; Ouyang, Z. Tropical Forest Strata Shifts in Plant Structural Diversity-Aboveground Carbon Relationships along Altitudinal Gradients. Sci. Total Environ. 2022, 838, 155907. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Wang, Q.; Han, S.; Guo, Z.; Yu, J.; Wang, W.; Fan, C.; Cao, W.; Wang, L.; Xing, Y.; et al. Beneficial Effects of Warming on Temperate Tree Carbon Storage Depend on Precipitation and Mycorrhizal Types. Sci. Total Environ. 2022, 819, 153086. [Google Scholar] [CrossRef]
- Deng, M.; Hu, S.; Guo, L.; Jiang, L.; Huang, Y.; Schmid, B.; Liu, C.; Chang, P.; Li, S.; Liu, X.; et al. Tree Mycorrhizal Association Types Control Biodiversity-Productivity Relationship in a Subtropical Forest. Sci. Adv. 2023, 9, eadd4468. [Google Scholar] [CrossRef]
- Ali, A.; Yan, E.R.; Chen, H.Y.; Chang, S.X.; Zhao, Y.T.; Yang, X.D.; Xu, M.S. Stand Structural Diversity rather than Species Diversity Enhances Aboveground Carbon Storage in Secondary Subtropical Forests in Eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar] [CrossRef]
- Wang, S.; Jiménez-Alfaro, B.; Pan, S.; Yu, J.; Sanaei, A.; Sayer, E.J.; Ye, J.; Hao, Z.; Fang, S.; Lin, F.; et al. Differential Responses of Forest Strata Species Richness to Paleoclimate and Forest Structure. Forest Ecol. Manag. 2021, 499, 119605. [Google Scholar] [CrossRef]
- Cai, Q.; Ji, C.; Zhou, X.; Bruelheide, H.; Fang, W.; Zheng, T.; Zhu, J.; Shi, L.; Li, H.; Zhu, J.; et al. Changes in Carbon Storages of Fagus Forest Ecosystems along an Elevational Gradient on Mt. Fanjingshan in Southwest China. J. Plant Ecol. 2020, 13, 139–149. [Google Scholar] [CrossRef]
- Mensah, S.; Salako, V.K.; Seifert, T.; Ostertag, R. Structural Complexity and Large-Sized Trees Explain Shifting Species Richness and Carbon Relationship across Vegetation Types. Funct. Ecol. 2020, 34, 1731–1745. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zobel, M. How Mycorrhizal Associations Drive Plant Population and Community Biology. Science 2020, 367, eaba1223. [Google Scholar] [CrossRef]
- Carteron, A.; Vellend, M.; Laliberté, E. Mycorrhizal Dominance Reduces Local Tree Species Diversity across US Forests. Nat. Ecol. Evol. 2022, 6, 370–374. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and Common Traits in Mycorrhizal Symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The Mycorrhizal-Associated Nutrient Economy: A New Framework for Predicting Carbon–Nutrient Couplings in Temperate Forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef]
- Körner, C. Mountain Biodiversity, Its Causes and Function. AMBIO J. Hum. Environ. 2004, 33, 11–17. [Google Scholar] [CrossRef]
- Tang, Z.; Fang, J.; Chi, X.; Feng, J.; Liu, Y.; Shen, Z.; Wang, X.; Wang, Z.; Wu, X.; Zheng, C.; et al. Patterns of Plant Beta-Diversity Along Elevational and Latitudinal Gradients in Mountain Forests of China. Ecography 2012, 35, 1083–1091. [Google Scholar] [CrossRef]
- Zhou, G.; Yin, G.; Tang, X. Carbon Stocks of Forest Ecosystems in China: Biomass Equation; Science Press: Beijing, China, 2018. (In Chinese) [Google Scholar]
- Soudzilovskaia, N.A.; van Bodegom, P.M.; Terrer, C.; Zelfde, M.V.T.; McCallum, I.; Luke McCormack, M.; Fisher, J.B.; Brundrett, M.C.; de Sá, N.C.; Tedersoo, L. Global Mycorrhizal Plant Distribution Linked to Terrestrial Carbon Stocks. Nat. Commun. 2019, 10, 5077. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Classification and Elevational Gradient Analysis of Diversity and Carbon Density of Three Mountains’ Plant Communities in Southern China. Master Thesis, Peking University, Beijing, China, 2020. (In Chinese). [Google Scholar]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing Hierarchical and Variation Partitioning in Multiple Regression and Canonical Analyses Using the Rdacca.Hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 15 April 2023).
- Allen, A.P.; Brown, J.H.; Gillooly, J.F. Global Biodiversity, Biochemical Kinetics, and the Energetic-Equivalence Rule. Science 2002, 297, 1545–1548. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a Metabolic Theory of Ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Mao, Z.; Corrales, A.; Zhu, K.; Yuan, Z.; Lin, F.; Ye, J.; Hao, Z.; Wang, X. Tree Mycorrhizal Associations Mediate Soil Fertility Effects on Forest Community Structure in a Temperate Forest. New Phytol. 2019, 223, 475–486. [Google Scholar] [CrossRef]
- Chen, L.; Swenson, N.G.; Ji, N.; Mi, X.; Ren, H.; Guo, L.; Ma, K. Differential Soil Fungus Accumulation and Density Dependence of Trees in a Subtropical Forest. Science 2019, 366, 124–128. [Google Scholar] [CrossRef]
- Bennett, J.A.; Maherali, H.; Reinhart, K.O.; Lekberg, Y.; Hart, M.M.; Klironomos, J. Plant-Soil Feedbacks and Mycorrhizal Type Influence Temperate Forest Population Dynamics. Science 2017, 355, 181–184. [Google Scholar] [CrossRef]
- Teste, F.P.; Kardol, P.; Turner, B.L.; Wardle, D.A.; Zemunik, G.; Renton, M.; Laliberté, E. Plant-Soil Feedback and the Maintenance of Diversity in Mediterranean-Climate Shrublands. Science 2017, 355, 173–176. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Paoli, G.; McGuire, K.; Amaral, I.; Barroso, J.; Bastian, M.; Blanc, L.; Bongers, F.; Boundja, P.; Clark, C.; et al. Large Trees Drive Forest Aboveground Biomass Variation in Moist Lowland Forests across the Tropics. Global Ecol. Biogeogr. 2013, 22, 1261–1271. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon Cycling and Storage in World Forests: Biome Patterns Related to Forest Age. Global Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Corrales, A.; Mangan, S.A.; Turner, B.L.; Dalling, J.W.; Chave, J. An Ectomycorrhizal Nitrogen Economy Facilitates Monodominance in a Neotropical Forest. Ecol. Lett. 2016, 19, 383–392. [Google Scholar] [CrossRef]
- Harley, J.L.; Harley, E.L. A Check-list of Mycorrhiza in the British Flora. New Phytol. 1987, 105, 1–102. [Google Scholar] [CrossRef]
- Haug, I.; Weber, R.; Oberwinkler, F.; Tschen, J. The Mycorrhizal Status of Taiwanese Trees and the Description of Some Ectomycorrhizal Types. Trees 1994, 8, 237–253. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Abbreviation | Unit | Mt. Wuyishan | Mt. Gonggashan | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Min | Max | Mean | SD | Min | Max | |||
| Response variables | ||||||||||
| Tree species richness | TSR | species/600 m2 | 19 | 9.5 | 5 | 39 | 9.1 | 4.9 | 4 | 19 |
| Stand biomass | Biomass | Mg ha−1 | 225.3 | 71.6 | 110.0 | 436.7 | 295.0 | 106.4 | 171.8 | 612.5 |
| Possible predictive variables | ||||||||||
| Elevation | Elevation | m | 1340 | 458 | 573 | 2070 | 3085 | 397 | 2445 | 3736 |
| Climate | ||||||||||
| Mean annual temperature | MAT | °C | 13.2 | 2.0 | 10.1 | 16.5 | 3.4 | 2.7 | −0.9 | 7.7 |
| Mean annual precipitation | MAP | mm | 2170 | 241 | 1894 | 2642 | 2111 | 83 | 1946 | 2202 |
| Stand structure | ||||||||||
| Stem density | Density | individuals ha−1 | 3120 | 1027 | 1633 | 6300 | 1556 | 953 | 350 | 4200 |
| Maximum DBH | max_DBH | cm | 43.5 | 10.2 | 29.6 | 75.4 | 82.8 | 23.4 | 42.8 | 126.1 |
| DBH variance | cv_DBH | unitless | 0.69 | 0.13 | 0.43 | 0.95 | 1.08 | 0.25 | 0.64 | 1.62 |
| Tree height variance | cv_H | unitless | 0.43 | 0.09 | 0.19 | 0.58 | 0.83 | 0.18 | 0.54 | 1.32 |
| Mycorrhizal dominance | ||||||||||
| Ectomycorrhizal dominance | EcM% | % | 58.9 | 22.1 | 25.6 | 95.1 | 80.4 | 15.7 | 39.7 | 98.3 |
| Arbuscular mycorrhizal dominance | AM% | % | 32.8 | 19.9 | 3.8 | 68.5 | 11.5 | 15.2 | 0.1 | 60.3 |
| Ericoid mycorrhizal dominance | ErM% | % | 8.3 | 7.4 | 0.2 | 24.5 | 8.1 | 11.3 | 0.0 | 39.3 |
| Variables | Standardized Total Effects | Standardized Direct Effects | Standardized Indirect Effects | |||
|---|---|---|---|---|---|---|
| TSR | Biomass | TSR | Biomass | TSR | Biomass | |
| Mt. Wuyishan | ||||||
| MAT | 0.83 | 0.20 | 0.61 | 0.68 | 0.22 | −0.48 |
| max_DBH | −0.03 | 0.67 | 0 | 0.32 | −0.03 | 0.35 |
| cv_H | 0.37 | 0.20 | 0.37 | 0.43 | 0 | −0.23 |
| EcM% | −0.06 | 0.60 | −0.06 | 0.57 | 0 | 0.04 |
| TSR | 0 | −0.62 | / | −0.62 | 0 | 0 |
| Mt. Gonggashan | ||||||
| MAT | 0.87 | 0.37 | 0.87 | 0 | 0 | 0.37 |
| MAP | −0.20 | 0.23 | 0 | 0 | −0.20 | 0.23 |
| max_DBH | 0 | 0.67 | 0 | 0.86 | 0 | −0.19 |
| cv_H | 0 | −0.28 | 0 | −0.28 | 0 | 0 |
| EcM% | −0.24 | 0.37 | −0.24 | 0.35 | 0 | 0.02 |
| TSR | 0 | −0.09 | 0 | −0.09 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Q.; Ma, S.; Sun, L.; Chen, G.; Xiao, J.; Fang, W.; Ji, C.; Tang, Z.; Fang, J. Elevational Patterns of Tree Species Richness and Forest Biomass on Two Subtropical Mountains in China. Forests 2023, 14, 1337. https://doi.org/10.3390/f14071337
Cai Q, Ma S, Sun L, Chen G, Xiao J, Fang W, Ji C, Tang Z, Fang J. Elevational Patterns of Tree Species Richness and Forest Biomass on Two Subtropical Mountains in China. Forests. 2023; 14(7):1337. https://doi.org/10.3390/f14071337
Chicago/Turabian StyleCai, Qiong, Suhui Ma, Lijuan Sun, Guoping Chen, Jian Xiao, Wenjing Fang, Chengjun Ji, Zhiyao Tang, and Jingyun Fang. 2023. "Elevational Patterns of Tree Species Richness and Forest Biomass on Two Subtropical Mountains in China" Forests 14, no. 7: 1337. https://doi.org/10.3390/f14071337
APA StyleCai, Q., Ma, S., Sun, L., Chen, G., Xiao, J., Fang, W., Ji, C., Tang, Z., & Fang, J. (2023). Elevational Patterns of Tree Species Richness and Forest Biomass on Two Subtropical Mountains in China. Forests, 14(7), 1337. https://doi.org/10.3390/f14071337