

Optimal Soil Calcium for the Growth of Mulberry Seedlings Is Altered by Nitrogen Addition

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Experimental Design
2.2. Plant and Soil Analysis
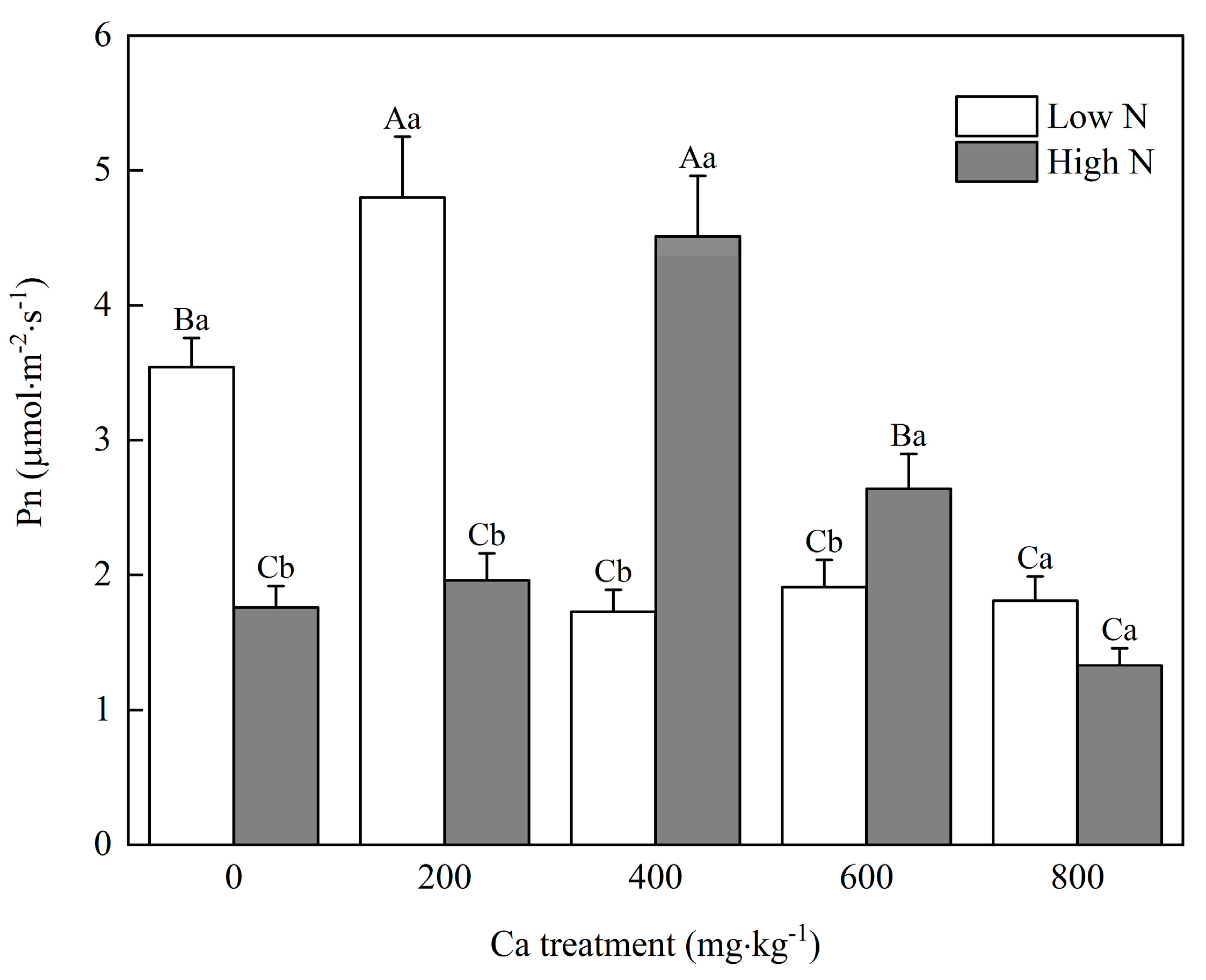
2.2.1. Determination of Net Photosynthetic Rate
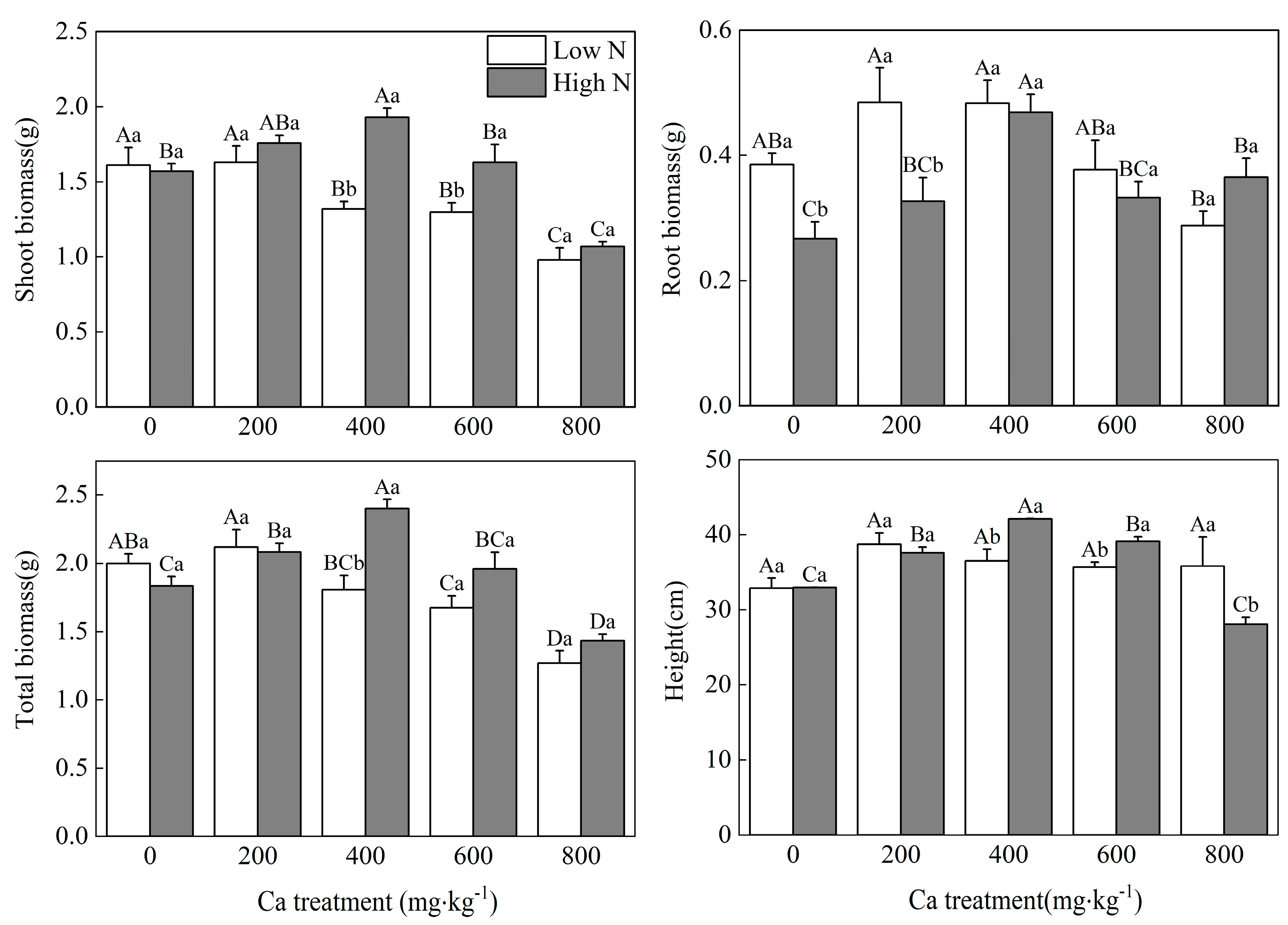
2.2.2. Growth Characteristics
2.2.3. Determination of Soil Ca
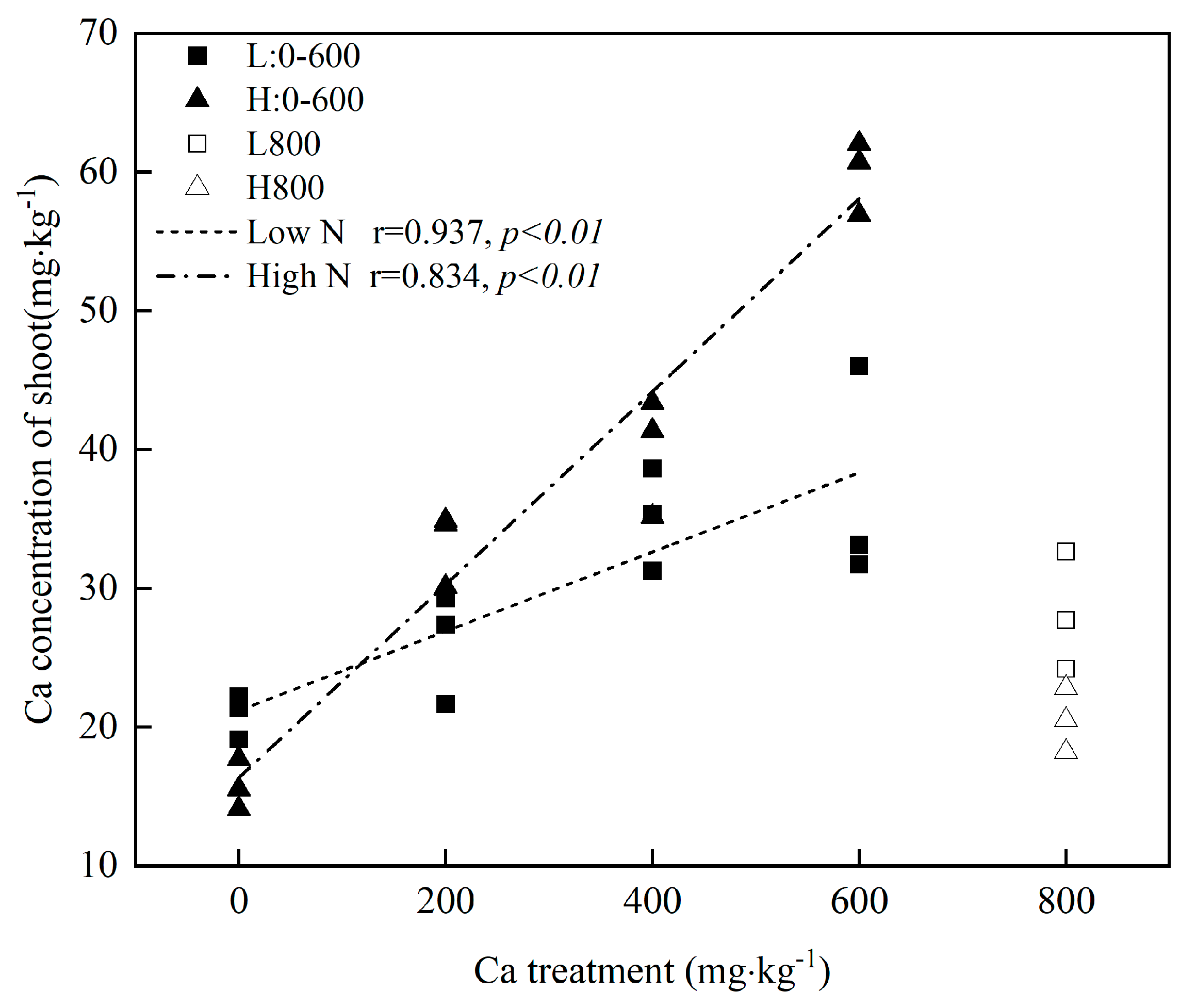
2.2.4. Shoot Ca Concentration
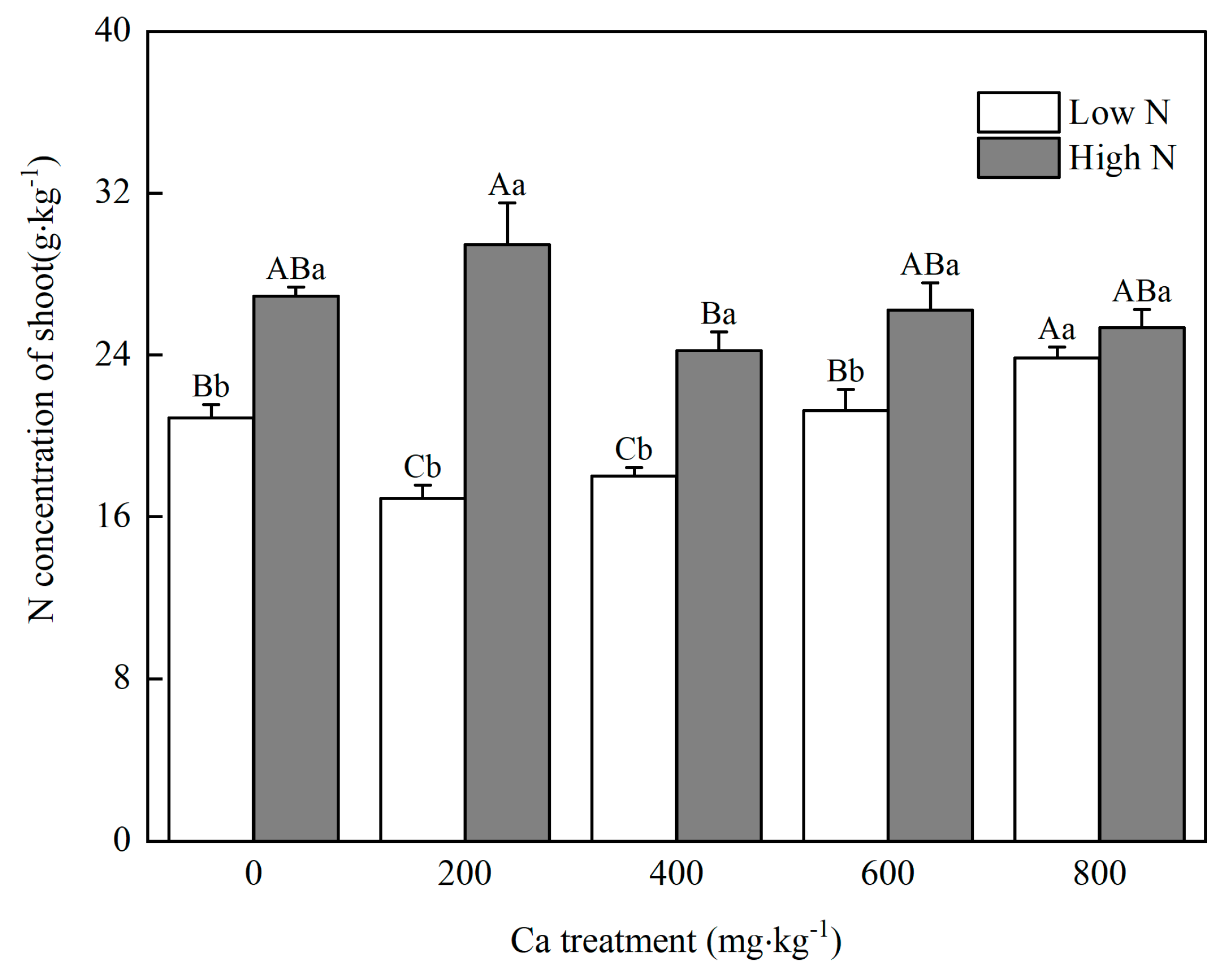
2.2.5. Shoot N Concentration
2.2.6. Leaf δ13C and iWUE
2.3. Statistical Analysis
3. Results
3.1. Available Concentrations of Ca in Soil
3.2. Mulberry Growth
3.3. Uptake of N and Ca in Mulberry
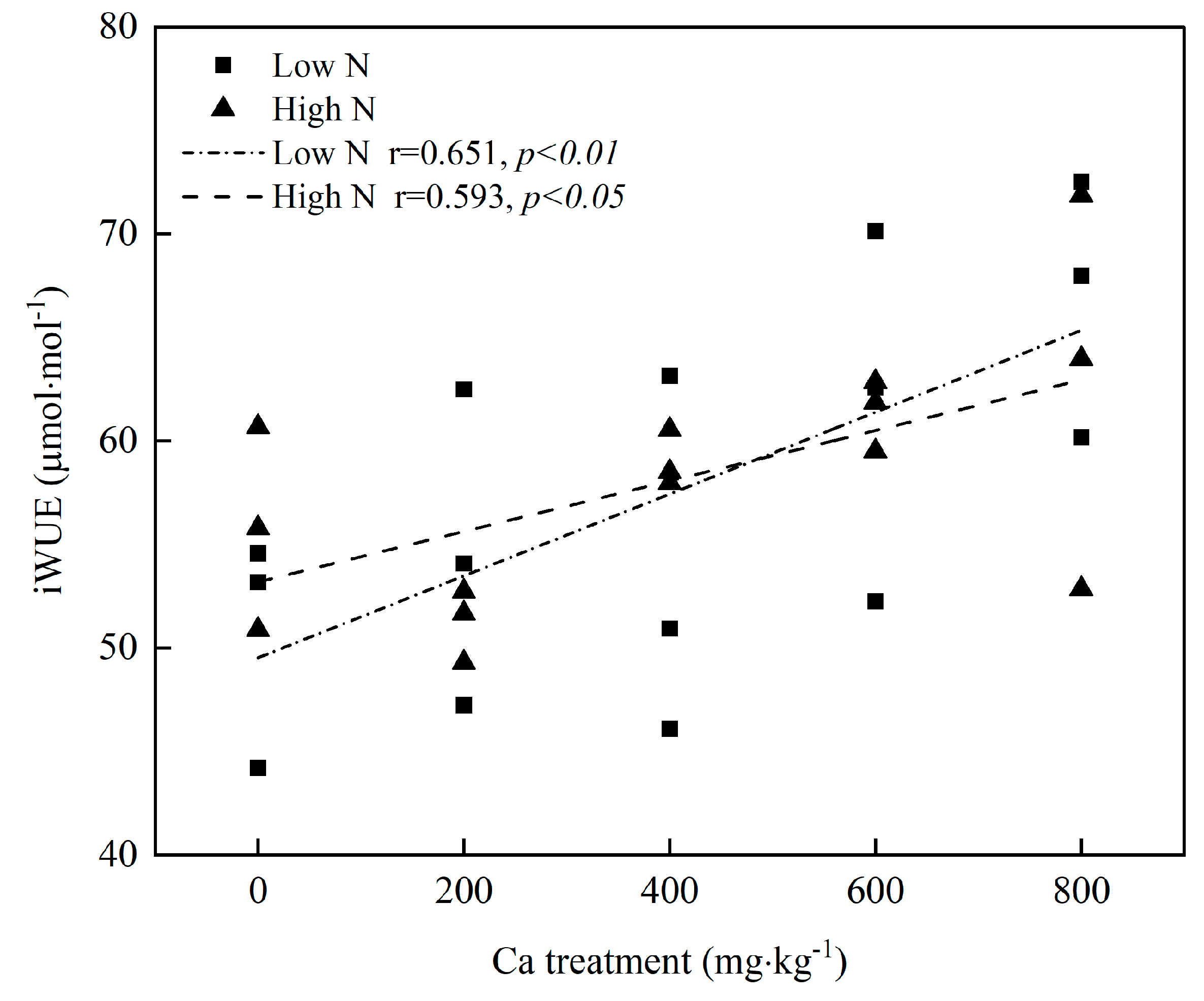
3.4. Relationship between iWUE and Ca
4. Discussion
4.1. N Influences the Optimal Soil Ca Level for Mulberry Seedlings
4.2. Soil Ca Influences Plant Ca Uptake
4.3. Ca Increases Plant iWUE and Drought Resistance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perakis, S.S.; Maguire, D.A.; Bullen, T.D.; Cromack, K.; Waring, R.H.; Boyle, J.R. Coupled Nitrogen and Calcium Cycles in Forests of the Oregon Coast Range. Ecosystems 2006, 9, 63–74. [Google Scholar] [CrossRef]
- Broadley, M.R.; Bowen, H.C.; Cotterill, H.L.; Hammond, J.P.; Meacham, M.C.; Mead, A.; White, P.J. Variation in the shoot calcium content of angiosperms. J. Exp. Bot. 2003, 54, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; Wirén, N.V. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, K.D. The calcium conundrum. Both versatile nutrient and specific signal. Plant Physiol. 2004, 136, 2438–2442. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q. Exogenous calcium alters activities of antioxidant enzymes in Trifolium repens L. leaves under PEG-induced water deficit. J. Plant Nutr. 2010, 33, 1874–1885. [Google Scholar] [CrossRef]
- He, L.; Lu, X.; Tian, J.; Yang, Y.; Li, B.; Li, J.; Guo, S. Proteomic analysis of the effects of exogenous calcium on hypoxic-responsive proteins in cucumber roots. Proteome Sci. 2012, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Sanavy, S.A.M.M.; Gholamhoseini, M.; Joghan, A.K.; Majdi, M.; Kashkooli, A.B. The role of calcium in improving photosynthesis and related physiological and biochemical attributes of spring wheat subjected to simulated acid rain. Physiol. Mol. Biol. Plants 2013, 19, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zhang, B. Effect of exogenous calcium on growth, nutrients uptake and plasma membrane H+-ATPase and Ca2+-ATPase activities in soybean (Glycine max) seedlings under simulated acid rain stress. Ecotoxicol. Environ. Saf. 2018, 165, 261–269. [Google Scholar] [CrossRef]
- Gradowski, T.; Thomas, S.C. Responses of Acer saccharum canopy trees and saplings to P, K and lime additions under high N deposition. Tree Physiol. 2008, 28, 173–185. [Google Scholar] [CrossRef]
- Li, W.; Xu, F.; Chen, S.; Zhang, Z.; Zhao, Y.; Jin, Y.; Li, M.; Zhu, Y.; Liu, Y.; Yang, Y.; et al. A comparative study on Ca content and distribution in two Gesneriaceae species reveals distinctive mechanisms to cope with high rhizospheric soluble calcium. Front. Plant Sci. 2014, 5, 647. [Google Scholar] [CrossRef]
- Kosiba, A.M.; Schaberg, P.G.; Rayback, S.A.; Hawley, G.J. The surprising recovery of red spruce growth shows links to decreased acid deposition and elevated temperature. Sci. Total Environ. 2018, 637–638, 1480–1491. [Google Scholar] [CrossRef]
- Blum, J.D.; Klaue, A.; Nezat, C.A.; Driscoll, C.T.; Johnson, C.E.; Siccama, T.G.; Eagar, C.; Fahey, T.J.; Fahey, G.E. Mycorrhizal weathering of apatite as an important calcium source in base-poor forest ecosystems. Nature 2002, 417, 729–731. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Bernhardt, E.S. Chapter 6-The Biosphere: Biogeochemical Cycling on Land. Biogeochemistry 2013, 173–231. [Google Scholar] [CrossRef]
- Hynicka, J.D.; Pett-Ridge, J.C.; Perakis, S.S. Nitrogen enrichment regulates calcium sources in forests. Glob. Chang. Biol. 2016, 22, 4067–4079. [Google Scholar] [CrossRef]
- Santos MAd Valadares, R.V.; Neves, J.C.L.; Silvac IRd Tótolad, M.R.; Costa, M.D. Ammonium nitrogen increases Ca uptake from non-exchangeable reservoirs by eucalypt plants. Forest Ecol. Manag. 2020, 465, 118062. [Google Scholar] [CrossRef]
- Khalaj, K.; Ahmadi, N.; Souri, M.K. Improvement of Postharvest Quality of Asian Pear Fruits by Foliar Application of Boron and Calcium. Horticulturae 2017, 3, 15. [Google Scholar] [CrossRef]
- Lee, J.A. The calcicole-calcifuge problem revisited. Adv. Bot. Res. 1998, 29, 1–30. [Google Scholar] [CrossRef]
- Liu, T.; Wu, F.; Wang, W.; Chen, J.; Li, Z.; Dong, X. Effects of calcium on seed germination, seedling growth and photosynthesis of six forest tree species under simulated acid rain. Tree Physiol. 2011, 31, 402–413. [Google Scholar] [CrossRef]
- Martín-Diana, A.B.; Rico, D.; Frías, J.M.; Barat, J.M.; Henehan, G.T.M.; Barry-Ryan, C. Calcium for extending the shelf life of fresh whole and minimally processed fruits and vegetables: A review. Trends Food Sci. Technol. 2007, 18, 210–218. [Google Scholar] [CrossRef]
- Ciccarese, A.; Stellacci, A.M.; Gentilesco, G.; Rubino, P. Effectiveness of pre- and post-veraison calcium applications to control decay and maintain table grape fruit quality during storage. Postharvest Biol. Technol. 2013, 75, 135–141. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium:Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef]
- Juice, S.M.; Fahey, T.J.; Siccama, T.G.; Driscoll, C.T.; Denny, E.G.; Eagar, C. Response of sugar maple to calcium addition to northern hardwoood forest. Ecology 2006, 87, 1267–1280. [Google Scholar] [CrossRef]
- Minick, K.J.; Fisk, M.C.; Groffman, P.M. Calcium and phosphorus interact to reduce mid-growing season net nitrogen mineralization potential in organic horizons in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 271–279. [Google Scholar] [CrossRef]
- Indrasumunar, A.; Menzies, N.W.; Dart, P.J. Calcium affects the competitiveness of acid-sensitive and acid-tolerant strains of Bradyrhizobium japonicum in nodulating and fixing nitrogen with two soybean cultivars in acid soil. Soil Biol. Biochem. 2012, 46, 115–122. [Google Scholar] [CrossRef]
- Kang, J.; Zhao, W.; Zheng, Y.; Zhang, D.; Zhou, H.; Sun, P. Calcium chloride improves photosynthesis and water status in the C4 succulent xerophyte Haloxylon ammodendron under water deficit. Plant Growth Regul. 2017, 82, 467–478. [Google Scholar] [CrossRef]
- He, N.; Zhang, C.; Qi, X.; Zhao, S.; Tao, Y.; Yang, G.; Lee, T.H.; Wang, X.; Cai, Q.; Li, D.; et al. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013, 4, 2445. [Google Scholar] [CrossRef]
- Li, M.; Hassan, F.U.; Tang, Z.; Peng, L.; Liang, X.; Li, L.; Peng, K.; Xie, F.; Yang, C. Mulberry Leaf Flavonoids Improve Milk Production, Antioxidant, and Metabolic Status of Water Buffaloes. Front. Vet. Sci. 2020, 7, 599. [Google Scholar] [CrossRef]
- Cui, H.; Lu, T.; Wang, M.; Zou, X.; Zhang, Y.; Yang, X.; Dong, Y.; Zhou, H. Flavonoids from Morus alba L. leaves: Optimization of extraction by response surface methodology and comprehensive evaluation of their antioxidant, antimicrobial, and inhibition of α-Amylase activities through analytical hierarchy process. Molecules 2019, 24, 2398. [Google Scholar] [CrossRef]
- Lu, X.; Vitousekc, P.M.; Mao, Q.; Gilliamd, F.S.; Luo, Y.; Zhou, G. Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest. Proc. Natl. Acad. Sci. USA 2018, 115, 5187–5192. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Hall, A.E.; Farquhar, G.D. Stable Isotopes and Plant Carbon-Water Relations; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Lawrence, G.B.; David, M.B.; Shortle, W.C. A new mechanism for calcium loss in forest-floor soils. Nature 1995, 378, 162–165. [Google Scholar] [CrossRef]
- Likens, G.E.; Driscoll, C.T.; Buso, D.C.; Siccama, T.G.; Johnson, C.E.; Lovett, G.M.; Fahey, T.J.; Reiners, W.A.; Ryan, D.F.; Martin, C.W.; et al. The biogeochemistry of calcium at Hubbard Brook. Biogeochemistry 1998, 41, 89–173. [Google Scholar] [CrossRef]
- Page, B.D.; Bullen, T.D.; Mitchell, M.J. Influences of Calcium Availability and Tree Species on Ca Isotope Fractionation in Soil and Vegetation. Biogeochemistry 2008, 88, 1–13. [Google Scholar] [CrossRef]
- Kobe, R.K.; Likens, G.E.; Eagar, C. Tree seedling growth and mortality responses to manipulations of calcium and aluminum in a northern hardwood forest. Can. J. Forest Res. 2002, 32, 954–966. [Google Scholar] [CrossRef]
- Gabara, B.; Sklodowska, M.; Wyrwicka, A.; Glińska, S.; Gapińska, M. Changes in the ultrastructure of chloroplasts and mitochondria and antioxidant enzyme activity in Lycopersicon esculentum Mill. leaves sprayed with acid rain. Plant Sci. 2003, 164, 507–516. [Google Scholar] [CrossRef]
- Baribault, T.W.; Kobe, R.K.; Rothstein, D.E. Soil calcium, nitrogen, and water are correlated with aboveground net primary production in northern hardwood forests. For. Ecol. Manag. 2010, 260, 723–733. [Google Scholar] [CrossRef]
- Michalet, R.; Gandoy, C.; Joud, D.; Pages, J.P.; Choler, P. Plant Community Composition and Biomass on Calcareous and Siliceous Substrates in the Northern French Alps: Comparative Effects of Soil Chemistry and Water Status. Arct. Antarct. Alp. Res. 2002, 34, 102–113. [Google Scholar] [CrossRef]
- Reich, P.B.; Schoettle, A.W. Role of phosphorus and nitrogen in photosynthetic and whole plant carbon gain and nutrient use efficiency in eastern white pine. Oecologia 1988, 77, 25–33. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef]
- Reyer, C.; Lasch-Born, P.; Suckow, F.; Gutsch, M.; Murawski, A.; Pilz, T. Projections of regional changes in forest net primary productivity for different tree species in Europe driven by climate change and carbon dioxide. Ann. Forest Sci. 2014, 71, 211–225. [Google Scholar] [CrossRef]
- Nakji, T.; Fukami, M.; Dokiya, Y.; Izuta, T. Effects of high nitrogen load on growth, photosynthesis and nutrient status of Cryptomeria japonica and Pinus densiflora seedlings. Trees 2001, 15, 453–461. [Google Scholar] [CrossRef]
- Wu, C.; Wang, Z.; Fan, Z.; Sun, H. Effects of different concentrations and form ratios of nitrogenon chlorophyll biosynthesis, photosynthesis, and biomass partitioning in Fraxinus Mandshurica seedlings. Acta Phytoecologica Sinica 2003, 27, 771–779. [Google Scholar] [CrossRef]
- White, P.J. The pathways of calcium movement to the xylem. J. Exp. Bot. 2001, 52, 891–899. [Google Scholar] [CrossRef]
- Blatt, M.R. Ca2+ signalling and control of guard-cell Volume in stomatal movements. Curr. Opin. Plant Biol. 2000, 3, 196–204. [Google Scholar] [CrossRef]
- Donovan, L.A.; Ehleringer, J.R. Ecophysiological differences among juvenile and reproductive plants of several woody species. Oecologia 1991, 86, 594–597. [Google Scholar] [CrossRef]
- Beer, C. Impact of soil freezing and thawing dynamics on soil organic carbon stocks in permafrost regions. IOP Conf. Ser. Earth Environ. Sci. 2009, 6, 052013. [Google Scholar] [CrossRef]
- Pei, Z.M.; Ghassemian, M.; Kwak, C.M.; McCourt, P.; Schroeder, J.I. Role of Farnesyltransferase in ABA Regulation of Guard Cell Anion Channels and Plant Water Loss. Science 1998, 282, 287–290. [Google Scholar] [CrossRef]
- Pei, Z.M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Reddy, A.S.N.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with Stresses: Roles of Calcium-and Calcium/Calmodulin-Regulated Gene Expression. Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic C concentration (g·kg−1) | 18.02 ± 0.53 |
| Total N concentration (g·kg−1) | 1.47 ± 0.09 |
| Total P concentration (g·kg−1) | 1.18 ± 0.10 |
| Available N concentration (mg·kg−1) | 174.2 ± 7.32 |
| Available P concentration (mg·kg−1) | 13.07 ± 0.13 |
| Water-extractable Ca concentration (mg·kg−1) | 113.62 ± 5.22 |
| Exchangeable Ca concentration (mg·kg−1) | 4567.32 ± 54.70 |
| Maximum water-holding capacity (%) | 30.5 ± 0.33 |
| Variables | Ca | N | Interaction | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Water-extractable Ca2+ | 84.302 | <0.01 | 3.078 | 0.095 | 0.197 | 0.937 |
| Exchangeable Ca2+ | 9.328 | <0.01 | 11.704 | <0.01 | 1.596 | 0.214 |
| Pn | 30.053 | <0.01 | 7.390 | <0.05 | 70.486 | <0.01 |
| Shoot biomass | 29.129 | <0.01 | 21.770 | <0.01 | 9.199 | <0.01 |
| Root biomass | 32.139 | <0.01 | 27.142 | <0.01 | 7.306 | <0.01 |
| Total biomass | 23.323 | <0.01 | 8.722 | <0.01 | 5.335 | <0.01 |
| Height | 9.341 | <0.01 | 0.015 | 0.903 | 5.392 | <0.01 |
| N concentration in shoot | 3.496 | <0.05 | 96.979 | <0.01 | 7.966 | <0.01 |
| Ca concentration in shoot | 53.774 | <0.01 | 9.575 | <0.01 | 14.350 | <0.01 |
| iWUE | 3.050 | <0.05 | 0.013 | 0.909 | 0.768 | 0.558 |
| Ca Treatment (mg⋅kg−1) | Water-Extractable Ca (mg⋅kg−1) | Exchangeable Ca (mg⋅kg−1) | ||
|---|---|---|---|---|
| Low N | High N | Low N | High N | |
| 0 | 61.4 ± 2.5 Eb | 91.9 ± 2.3 Ea | 4264.8 ± 47.5 Ba | 4188.4 ± 50.7 Ba |
| 200 | 225.1 ± 6.5 Da | 257.7 ± 4.9 Da | 4279.4 ± 57.8 Ba | 4246.4 ± 24.1 ABa |
| 400 | 379.2 ± 10.5 Cb | 436.5 ± 11.6 Ca | 4326.5 ± 93.1 Ba | 4287.8 ± 94.5 ABa |
| 600 | 595.6 ± 8.8 Ba | 620.8 ± 16.1 Ba | 4533.9 ± 64.3 Aa | 4259.9 ± 43.8 ABb |
| 800 | 715.3 ± 13.1 Aa | 804.0 ± 14.5 Aa | 4595.2 ± 38.8 Aa | 4445.2 ± 77.5 Aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Li, Y.; Li, H.; Zhang, S.; Zhou, Y. Optimal Soil Calcium for the Growth of Mulberry Seedlings Is Altered by Nitrogen Addition. Forests 2023, 14, 399. https://doi.org/10.3390/f14020399
Zhang T, Li Y, Li H, Zhang S, Zhou Y. Optimal Soil Calcium for the Growth of Mulberry Seedlings Is Altered by Nitrogen Addition. Forests. 2023; 14(2):399. https://doi.org/10.3390/f14020399
Chicago/Turabian StyleZhang, Tengzi, Yanan Li, Hui Li, Songzhu Zhang, and Yongbin Zhou. 2023. "Optimal Soil Calcium for the Growth of Mulberry Seedlings Is Altered by Nitrogen Addition" Forests 14, no. 2: 399. https://doi.org/10.3390/f14020399
APA StyleZhang, T., Li, Y., Li, H., Zhang, S., & Zhou, Y. (2023). Optimal Soil Calcium for the Growth of Mulberry Seedlings Is Altered by Nitrogen Addition. Forests, 14(2), 399. https://doi.org/10.3390/f14020399