Abstract
Water use efficiency (WUE) is key to linking the water, carbon, and nutrient cycles in terrestrial ecosystems. However, the coupling between WUE and leaf nutrients is still poorly understood in subtropical forests. Here, the stable carbon isotope technique was employed to estimate the leaf-scale WUE of five common tree species (Castanopsis eyrei, Symplocos laurina, Machilus grijsii, Ternstroemia gymnanthera, and Rhododendron ovatum) in different habitat types (i.e., hillside, near the top of the peak, and peak) in a subtropical broad-leaf evergreen forest on the western slope of Wuyi Mountain, southern China. In addition, leaf carbon (C), nitrogen (N), and phosphorus (P) contents were also measured to assess plant nutrient utilization and its relationship with WUE. From the hillside to the peak, soil water content showed a decreasing trend, whereas the soil total C, N, and P contents showed an increasing trend. Regardless of species, the leaf δ13Cp value and WUE showed an increasing trend from the hillside to the peak, mainly due to an increase in soil water deficit and light. The leaf N and P contents showed an increasing trend from hillside to peak due to an increase in soil nutrients, while the leaf C: N ratio, C:P ratio, and N:P ratio showed a decreasing trend. The regression analysis showed that leaf-scale WUE was positively correlated with the leaf N and P contents but negatively correlated with the leaf N:P ratio, especially for the three species (C. eyrei, S. laurina, and T. gymnanthera). These results indicated that the differences in soil water availability, light, and soil development resulting from different habitats have a significant impact on leaf-scale WUE and nutrient status on Wuyi Mountain. Therefore, there may be a close relationship between WUE and leaf nutrients, which would help us to better understand the water-, carbon-, and nutrient-coupled relationships for the evergreen broad-leaved tree species in this region.
1. Introduction
In recent years, global climate warming and corresponding environmental changes have significantly affected the plant community structure and function in terrestrial ecosystems [1]. Firstly, to cope with climate and environmental changes, plants will develop a series of physiological and ecological regulatory mechanisms, the critical aspect of which is to modulate plant water use efficiency (WUE) [2,3]. WUE is defined as the function between carbon assimilation rate and stomatal conductance [4]. Hence, WUE not only reflects the balance of water supply and consumption for plant growth [5] but also can indicate the plant’s adaptation to environmental changes [2,6]. Secondly, as the essential nutrients for plant growth, carbon (C), nitrogen (N), and phosphorus (P) can substantially influence the physiological and ecological function of the plants [7,8,9]. As the most crucial structural element in plant tissues, C accounts for about 50% of plant dry matter and is a substrate and energy source for various physiological and biochemical processes in plants [10,11]. N and P are the two most important limiting elements for plant growth, and the C, N, and P contents, as well as their stoichiometric ratios (i.e., C:N ratio, C:P ratio, and N:P ratio), can effectively reflect the uptake assimilation capacity and nutrient limitation of plants [8,12,13]. There have been many studies on leaf-scale WUE [2,3,5,6,14,15,16,17,18] and C, N, and P nutrients [7,8,10,19,20]. However, the intrinsic correlation between the leaf-level WUE and leaf nutrients is not well studied [9,21], which limits our understanding of the coupled relationships between carbon–water cycling and nutrient cycling in terrestrial ecosystems.
In C3 plants, leaf-scale ‘intrinsic’ WUE can be identified by the stable carbon isotope ratio in a leaf (δ13Cp), due to the fact that it is primarily related to the ratio of the intercellular (Ci) to the atmospheric (Ca) partial pressure of CO2 (Ci/Ca; [4,22,23]). As a sensitive indicator in response to climate and environmental changes, leaf-scale WUE is generally influenced by external environmental conditions (such as water availability and light) and internal leaf nutrients. For example, Liu et al. (2021) found that the leaf δ13Cp and WUE values of desert plants in northern China decreased with increasing annual precipitation, which was mainly because the increase in rainfall enhanced plant water availability and reduced light radiation [15]. Moreover, a soil water deficit can lead to stomatal closure and reduced stomatal conductance, resulting in a lower ratio of Ci/Ca and a higher leaf δ13Cp and WUE [4,22,23,24]. Moreover, light differences can affect the leaf δ13Cp and WUE by influencing chlorophyll content, leaf phototropism, carboxylase activity, and other biochemical processes that are related to photosynthesis in plants [11]. In addition to environmental factors, the plant’s nutrient status also can influence leaf-scale WUE by modulating plant photosynthesis. For instance, the leaf P content tends to be positively correlated with the photosynthetic rate [25]. This is because, under a shortage of P, the influence of chloroplasts on the photosynthetic rate is greater than that of stomatal conductance [26]. Likewise, the increase in leaf N content could enhance photosynthesis mainly by promoting the activity of enzymes [27]. At the leaf level, WUE was positively correlated with leaf P content, but it was not significantly associated with leaf N content in a P-limited subtropical forest [9,28]. However, the WUE of broad-leaf trees was positively correlated both with the leaf N and P contents in a temperate forest [11]. In rubber forests, WUE was positively correlated with the leaf N and P content but negatively correlated with the leaf N:P ratio [29].
The subtropical evergreen broad-leaved forest is one of the most important vegetation types in the Wuyi Mountains of southern China [30]. Changes in intrinsic WUE have been reported for different life types (i.e., trees, shrubs, and herbs) along the altitudinal gradient of Wuyi Mountain [21]. However, to date, a systematic study on the leaf-scale WUE changes and their coupling relationship with leaf nutrients in this region under different habitat conditions is still unclear. In this study, we selected five common trees (i.e., Castanopsis eyrei, Symplocos laurina, Machilus grijsii, Ternstroemia gymnanthera, and Rhododendron ovatum) as the target plants under different habitats (i.e., hillside, near the top of the peak, and peak) in a subtropical broad-leaf evergreen forest on the western slope of Wuyi Mountain, southern China. By measuring the leaf δ13Cp and the C, N, and P contents for these five tree species, our main aims are (1) to determine the variation patterns of leaf-scale WUE from hillside to peak; (2) to analyze the differences in leaf C, N, P contents and their stoichiometric ratios under different habitat conditions; and (3) to explore the relationship between intrinsic WUE and leaf nutrients for the five tree species. These results would help us to better understand the coupled relationships between WUE and nutrients for the evergreen broad-leaved tree species in this region.
2. Materials and Methods
2.1. Study Area, Experimental Design, and Sample Collection
We conducted the study in a subtropical evergreen broad-leaved forest (27°40′50″ N, 117°09′11″ E), which is located within Jiangxi Matou Mountain National Nature Reserve in Zixi County, Jiangxi Province, southern China (Figure 1). The study area lies on the western slope of Wuyi Mountain, where it preserves a larger area of native evergreen broad-leaved forests with a high forest coverage of 96.3%, making it an important gene pool of biological resources in the world. It has a typical subtropical humid monsoon climate with an annual average temperature of 17 °C, an annual average precipitation of 1930 mm, and a relative humidity of 83%. In this forest, the dominant species are mainly composed of evergreen and deciduous trees such as C. eyrei, Lithocarpus litseifolius, Schima superba, S. laurina, M. grijsii, T. gymnanthera, Alniphyllum fortune., etc. The soil types belong to red soil and red–yellow soil according to the Genetic Soil Classification of China [31].
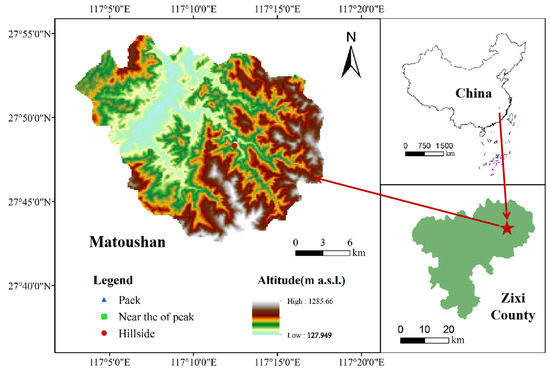
Figure 1.
Location of the study area and sampling points.
In October 2020, three sample plots with different habitat types (i.e., hillside in the northeast slope (H–NE), near the top of the peak in the southwest slope (NP–SW), and peak in the southwest slope (P–SW)) were set up in this subtropical evergreen broad-leaved forest (Figure 1 and Table 1). Five common dominant tree species (i.e., C. eyrei, S. laurina, M. grijsii, T. gymnanthera, and R. ovatum) were selected as target plants in each sample plot. In each plot, four quadrats with a size of 20 m × 20 m were randomly set up to investigate the five target trees. For each target species, five plants with similar heights and diameters at breast height (DBH) in each quadrat were selected to collect 10–15 mature leaves. The collected leaf samples were washed with pure water to remove surface impurities and dust, brought back to the laboratory, and oven-dried at 80 °C for 48 h. Each dried sample was then crushed using an onyx grinder, sieved through a 100-mesh sieve, and used as a sample to be tested. Meanwhile, soil samples from 0 to 30 cm were collected with a soil core (5 cm in diameter) after removing the forest floor in each sample plot according to a 10 m × 10 m grid to determine the initial soil moisture and nutrient contents.

Table 1.
Basic situation of plant growth environments of three sample plots in a subtropical broad-leaf evergreen forest on the western slope of Wuyi Mountain in southern China.
2.2. Sample Chemical Analysis
The soil water content was measured using the oven-drying method. Air-dried soil samples were milled to pass through a 0.15 mm sieve to analyze the soil nutrient contents. In this experiment, N and P contents were measured colorimetrically on an Autoanalyzer (AA3, Seal Analytical, Norderstedt, Germany) after acid digestion with 98% H2SO4 and 30% H2O2. Organic C content was measured with an elemental analyzer (Flash 2000 HT, Thermo Fisher Scientific, Bremen, Germany) [31]. In addition, 0.04~0.05 mg of dried leaf sample was weighed and placed in pre-burned tin capsules (3.5 × 9 mm) for stable carbon isotope measurement. The leaf δ13C was determined using an isotope ratio mass spectrometer (IRMS) (Delta V Advantage, Thermo Fisher Scientific, Bremen, Germany) coupled with an elemental analyzer (Flash EA 2000 HT, Thermo Fisher Scientific, Bremen, Germany) at the IRMS laboratory of Jiangxi Agricultural University. The cellulose standard (IAEA-CH3; δ13C = −24.724‰), L-glutamic acid (USGS40; δ13C = −26.39‰), and urea (CO(NH2)2; δ13C = −41.30‰) were used to calibrate the δ13C measurements, respectively. The standard deviation for the repeated analysis of an internal standard was <0.03‰.
2.3. Calculation of Water Use Efficiency (WUE)
The leaf-scale WUE is defined as the ratio of the rate of carbon assimilation (A, photosynthesis) and stomatal conductance (gs) and can be estimated from the stable carbon isotope discrimination value in leaves (∆13Cp) using the following equation [4,18,22,23]:
where ∆13Cp is the stable carbon isotope discrimination value (Equation (2)), Ci is the intercellular CO2 concentration, Ca is the atmospheric CO2 concentration obtained from Equation (3), a (4.4‰) is the fractionation during the diffusion of CO2, b (27‰) is the biochemical fractionation, and 1.6 is the ratio of the diffusivities of water vapor and carbon dioxide [22,23].
WUE = A/gs = (Ca − Ci)/1.6 = Ca (b − ∆13Cp)/[1.6 (b − a)]
The stable carbon isotope discrimination value (∆13Cp; Equation (2)) [4,22,23], atmospheric CO2 concentration (Ca; Equation (3)) [5,17], and stable carbon isotopic ratio (δ13Catm; Equation (4)) [5,17] were calculated from the following equations, respectively:
∆13Cp = a + (b − a) Ci/Ca = (δ13Catm − δ13Cp)/[(1 + δ13Cp)/1000]
Ca = 277.78 + 1.350exp (0.01572 (t − 1740))
δ13Catm = −6.429 − 0.006exp (0.0217 (t − 1740))
In the formula, δ13Cp is the stable carbon isotopic ratio of the leaves, and t is the sampling year. In this study, t = 2020, and substituting this into Equations (3) and (4), the Ca in 2020 was calculated to be 389.66 μmol mol−1, and δ13Catm was −6.93‰.
2.4. Statistical Analyses
The data were statistically analyzed using R-studio 4.1 software. A one-way analysis of variance (ANOVA) was employed to compare the differences in the leaf δ13Cp values, WUE, and nutrient contents of the same tree species under different habitat conditions. Additionally, the differences in soil water content, soil organic C content, soil total N content, and soil total P content were examined across different sampling plots. The significance level was set at 0.05. Linear fits were performed through a regression analysis of leaf-scale WUE to leaf N and P concentrations and the N:P ratio for each species, with 95% confidence intervals.
3. Results
3.1. Environmental Conditions under Different Habitat Types
There were significant differences in the environmental conditions under the three different habitat types (Table 1). Specifically, there was a smaller slope gradient and a lower altitude on the hillside on the northeast slope (H–NE), while there was the largest slope gradient and the highest altitude at the peak on the southwest slope (P–SW). For the soil properties, the soil water content decreased with an increasing altitudinal gradient from the hillside to the peak. However, the soil total C content, soil total N content, and soil total P content increased with an increasing altitudinal gradient from the hillside to the peak.
3.2. Leaf δ13Cp and WUE under Different Habitat Conditions
There were significant differences in the leaf δ13Cp and WUE for the five tree species under the different habitat conditions (Figure 2). From the hillside to the peak, a positive trend was observed in the leaf δ13Cp values of the five tree species. Consequently, the WUE values exhibited a corresponding increasing trend. Regardless of species, the relatively more negative leaf δ13Cp values are mainly found at the hillside, but the relatively positive leaf δ13Cp values are generated at the peak (Figure 2a). Thus, there was lower WUE at the hillside but higher WUE at the peak (Figure 2b).
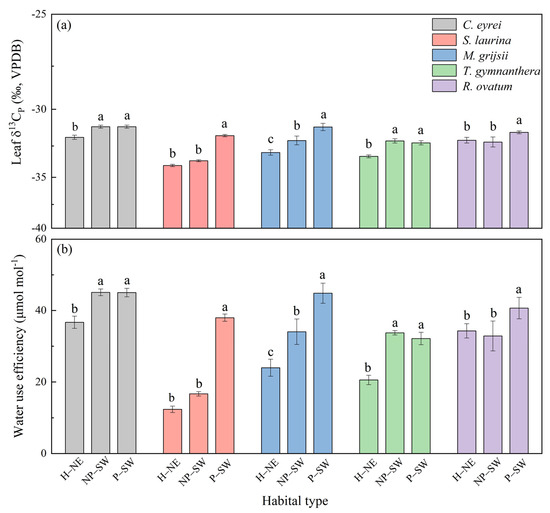
Figure 2.
Leaf δ13Cp (a) and WUE (b) of five tree species under different habitat conditions in a subtropical broadleaf evergreen forest on the western slope of Wuyi Mountain in southern China. Notes: H–NE: hillside on the northeast slope; NP–SW: near the top of the peak on the southwest slope; P–SW: the peak on the southwest slope. The different lowercase letters (a, b, c) mean that there is a significant difference among the treatments (H–NE, NP–SW, and P–SW) at p < 0.05.
3.3. Leaf Nutrients and Their Stoichiometric Ratios under Different Habitat Conditions
There were also significant differences in the leaf N and P contents and stoichiometric ratios under the different habitat conditions, but no significant differences in the leaf C contents (Figure 3 and Figure 4). From the hillside to the peak, the leaf N and P contents of the five tree species showed an increasing trend (Figure 3), but the leaf C:N ratio, C:P ratio, and N:P ratio exhibited a decreasing trend (Figure 4). Specifically, the leaf N content ranged from 6.95 mg g−1 to 16.29 mg g−1, the leaf P content ranged from 0.69 mg g−1 to 1.51 mg g−1, the leaf C:N ratio ranged from 26.07 to 72.37, the leaf C:P ratio ranged from 290.56 to 744.28, and the leaf N:P ratio ranged from 8.77 to 14.98. Overall, the leaf N and P contents at the peak were higher than those at the hillside (Figure 3b,c), while the leaf C:N, C:P, and N:P ratios at the peak were lower than those at the hillside (Figure 4).
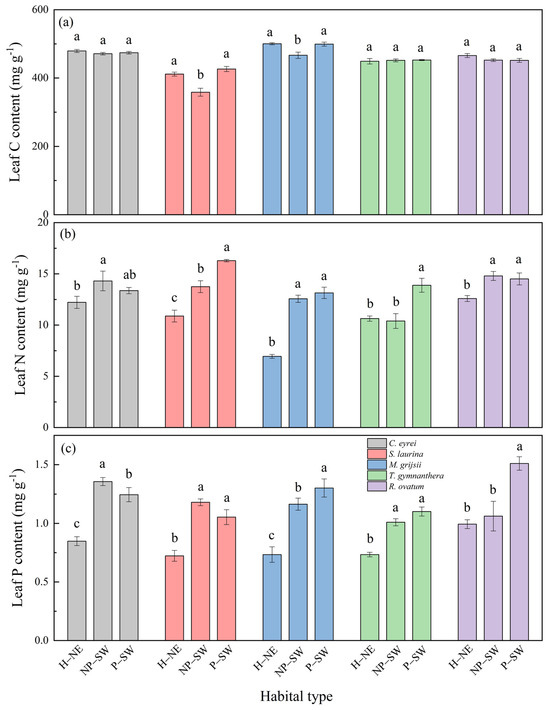
Figure 3.
Leaf C content (a), N content (b), and P content (c) of five tree species under different habitat conditions in a subtropical broad-leaf evergreen forest on the western slope of Wuyi Mountain in southern China. Notes: H–NE: hillside on the northeast slope; NP–SW: near the top of the peak on the southwest slope; P–SW: the peak on the southwest slope. The different lowercase letters (a, b, c) mean that there is a significant difference among the treatments (H–NE, NP–SW, and P–SW) at p < 0.05.
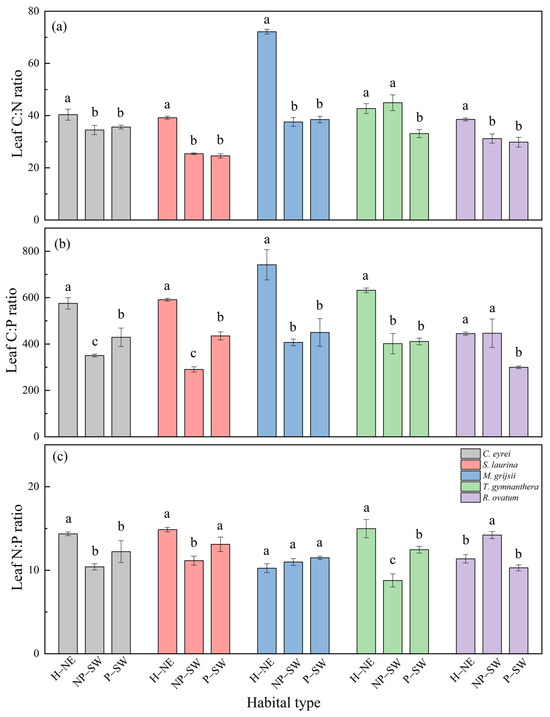
Figure 4.
Leaf C:N ratio (a), C:P ratio (b), and N:P ratio (c) of five tree species under different habitat conditions in a subtropical broadleaf evergreen forest on the western slope of Wuyi Mountain in southern China. Notes: H–NE: hillside on the northeast slope; NP–SW: near the top of the peak on the southwest slope; P–SW: the peak on the southwest slope. The different lowercase letters (a, b, c) mean that there is a significant difference among the treatments (H–NE, NP–SW, and P–SW) at p < 0.05.
3.4. Relationships between Leaf Nutrients and WUE
The leaf nutrients (i.e., N, P, and N:P ratio) both had a significant relationship with WUE for three of the tree species, namely C. eyrei, S. laurina, and T. gymnanthera, while only either N or P had a significant relationship with WUE for the remaining two tree species, M. grijsii and R. ovatum (Figure 5). Specifically, the leaf N content was significantly positively correlated with WUE for C. eyrei (r = 0.494; Slope = 1.13; p < 0.05; Figure 5a), S. laurina (r = 0.442; Slope = 1.47; p < 0.05; Figure 5d), M. grijsii (r = 0.691; Slope = 2.33; p < 0.01; Figure 5g), and T. gymnanthera (r = 0.728; Slope = 2.66; p < 0.01; Figure 5j), but not for R. ovatum (p > 0.05; Figure 5m). The leaf P content was also significantly positively correlated with WUE for C. eyrei (r = 0.759; Slope = 13.82; p < 0.01; Figure 5b), S. laurina (r = 0.899; Slope = 32.77; p < 0.01; Figure 5e), T. gymnanthera (r = 0.558; Slope = 18.89; p < 0.05; Figure 5k), and R. ovatum (r = 0.474; Slope = 7.98; p < 0.05; Figure 5n), but not for M. grijsii (p > 0.05; Figure 5h). However, the leaf N:P ratio was significantly negatively correlated with WUE for C. eyrei (r = −0.483; Slope = −0.83; p < 0.05; Figure 5c), S. laurina (r = −0.646; Slope = −1.85; p < 0.01; Figure 5f), and T. gymnanthera (r = −0.492; Slope = −4.09; p < 0.05; Figure 5l), but not for M. grijsii (p > 0.05; Figure 5i) and R. ovatum (p > 0.05; Figure 5o).
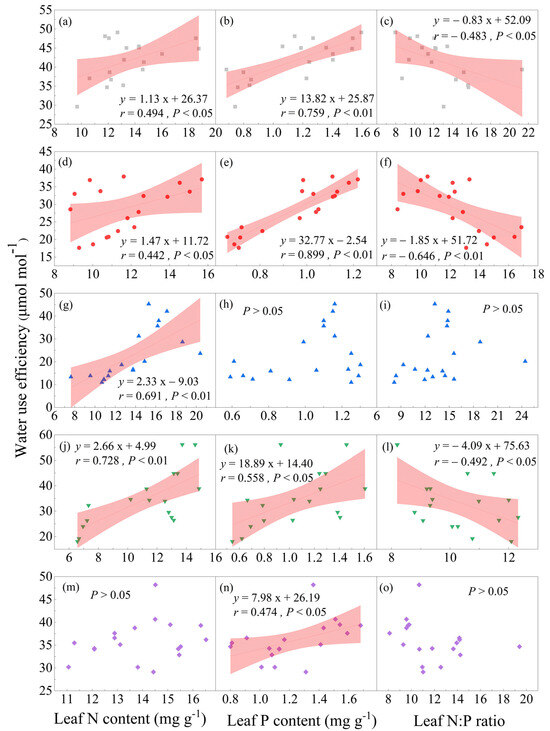
Figure 5.
The relationship between WUE and the leaf N content, leaf P content, and leaf N:P of the five tree species in a subtropical broad-leaf evergreen forest on the western slope of Wuyi Mountain in southern China. Notes: the gray square represents C. eyrei (a–c), the red circle represents S. laurina (d–f), the blue triangle up represents M. grijsii (g–i), the green triangle down represents T. gymnanthera (j–l), and the pink diamond represents R. ovatum (m–o). The linear fits were used to test the coupling between WUE and leaf nutrients with the significance level set at 0.05. The shadow indicates 95% confidence intervals.
4. Discussion
4.1. Effect of Habitat Types on Leaf-Scale WUE
At the local scale, water availability and light conditions are considered the two most important environmental factors to influence the leaf-scale WUE, mainly by controlling the stomatal conductance or photosynthetic capacity [4,11,17]. In this subtropical forest, the δ13Cp values of five tree species showed an increasing trend with decreasing soil moisture from the hillside to the peak (Table 1 and Figure 2), suggesting that soil water content has a more significant influence on leaf-scale WUE. Similarly, Huang et al. (2019) found that the leaf δ13Cp values of trees gradually increased from the depression to the hilltop due to decreased soil moisture in a karst seasonal rainforest [17]. Zhang et al. (2017) also found that whether the instantaneous WUE derived from the gas exchange or the intrinsic WUE calculated from the carbon isotope model for the Platycladus orientalis seedling, both reached their maximum value at the lowest soil water content [16]. Thus, these two results are consistent with our study, implying that leaf-scale WUE is highly sensitive to soil water availability. The regulation mechanisms of soil water availability for the leaf-scale WUE may be the following: In habitats with limited soil moisture, plant defense cells may reduce intracellular solutes through metabolic processes as a response to water scarcity in the root systems. This can lead to an increase in water potential and subsequent water loss within cells [4]. In this case, part of the stomata will be closed due to dehydration, and this physiological strategy can not only reduce plant transpiration but also decrease the ratio of Ci/Ca, thereby resulting in a more positive value of δ13C and higher WUE [22,23]. In contrast, stomatal conductance functions at its full capacity, resulting in a more negative value of δ13C and lower WUE in a moist habitat [6].
In addition to water availability, light is another important environmental factor that can affect leaf-scale WUE [11,17]. This is mainly because photosynthetic processes such as chlorophyll distribution, leaf phototropism, and carboxylase activity are all closely related to light [32]. Considering that the insolation index at the peak may be higher than that of the hillside, more light will lead to an increase in CO2 assimilated by photosynthesis, which may decrease stomatal conductance and result in a higher leaf δ13Cp value and WUE for the high-altitude species than for the low-altitude species (Figure 2). Huang et al. (2019) also found that the leaf δ13Cp value and WUE of Sterculia monosperma gradually increased from the depression to the peak with an increase in the insolation index [17]. Similarly, the δ13Cp and WUE values were higher in leaves at the top of the canopy than in leaves at the lower canopy due to differences in light intensity in the forest canopy structure [11,32]. Therefore, this evidence suggested that habitat-induced differences in soil water availability and light intensity can significantly affect leaf-scale WUE values.
4.2. Effect of Habitat Types on Leaf Nutrients and Their Stoichiometric Ratios
Leaf C, N, and P contents could reflect the nutrient uptake status of the plants, which is mainly influenced by soil nutrients, temperature, precipitation, mycorrhizal, and succession time [12,19,20,33]. In this study, we found that the leaf N and P contents showed an increasing trend with an altitudinal gradient from the hillside to the peak for the five species, despite no significant difference in the leaf C content (Figure 3b,c). In general, as the most crucial element in the dry matter of the plant, C is primarily derived from CO2 assimilation during photosynthesis; thus, it is not a limiting element for plant growth [7,20]. On the one hand, plants’ N sources are influenced by the weathering of parent rocks and the decomposition of organic matter in the soil. On the other hand, plants can also obtain N sources from the atmosphere through biological and high-energy N-fixation [34]. In this region, the succession time of the subtropical broad-leaved evergreen forests dominated by Castanopsis eyrei at the peak was often longer than that at the hillside, which led to the soil organic matter and soil total N contents being greater at the peak than at the hillside (Table 1). Thus, we found that the leaf N content was increased with increasing altitude from the hillside to the peak (Figure 3b). However, a plant’s P content is mainly derived from a parent rock’s weathering; hence, the soil total P content is lower if the weathering time of the parent rock is longer [13]. In this case, the plant will suffer from a stronger P limitation, particularly in subtropical forests [35]. We found that the leaf P content showed an increasing trend from the hillside to the peak (Figure 3c). This is mainly because the chemical weathering of the parent rock decreases with increasing altitude on Wuyi Mountain [21]. This leads to increased soil P content in the high-altitude habitats, resulting in a higher leaf P content at the peak.
The leaf C:N and C:P ratios reflect the plant’s ability to assimilate C as it absorbs N and P; hence, they are often used to indicate a plant’s nutrient use efficiency [7,11,20]. In this subtropical forest, we found that the leaf C:N and C:P ratios decreased with an increase in altitudinal gradient, suggesting that the hillside has a higher nutrient utilization efficiency than the peak due to the relatively lower leaf N and P contents in the low-altitude habitat (Figure 4a,b). In addition, the leaf N:P ratio can be used to assess the nutrient limitation status of plants to some extent [13]. At the community level, Townsend et al. (2007) suggested that N limitation likely occurs at an N:P ratio < 14, with P limitation probable at an N:P ratio > 16, and maybe both N- and P limitations when 14 < N:P < 16 [36]. In this region, we found that the leaf N:P ratio of the five tree species was chiefly less than 14 at the peak (Figure 4c), suggesting that plant growth was generally limited by N in high-altitude habitats. Interestingly, the leaf N:P ratio of most of the tree species on the hillside ranged between 14 and 16 (Figure 4c), suggesting that the growth of plants in this habitat is likely co-limited both by N and P. However, the subtropical forest was often considered a P-limited ecosystem [35,36]. This is not quite consistent with our results, and the main reason for this may be related to the degree of soil development. As mentioned earlier, on Wuyi Mountain, there is a weaker weathering intensity of the parent rock on the high-altitude mountain than that of the low-altitude mountain [21]. Therefore, at the mountain peak, the soil has a higher P content, which results in plants experiencing N limitation (Table 1). On the contrary, there is a more serious loss in P in mineral soils on the hillside due to the stronger soil development degree; hence, plant growth in this habitat is often subjected to the co-limitation of N and P. These results further implied that the difference in soil development caused by habitats will significantly affect the leaf nutrients and their C N P stoichiometric ratios in the subtropical forest ecosystem.
4.3. The Coupling Relationships between WUE and Leaf Nutrients for the Five Tree Species
Theoretically, there is a close coupling relationship between intrinsic WUE and leaf traits in terrestrial ecosystems [14]. Leaf nutrients (e.g., N and P) influence intrinsic WUE mainly by modulating photosynthetic and transpiration rates [9,11,21,29]. N is an essential component of chlorophyll, proteins, and enzymes in plants, which can significantly impact plant photosynthesis [37]. It has been shown that an increased N content can promote chlorophyll and protein synthesis and increase leaf area, thereby increasing plant photosynthesis [14,27]. Moreover, increased N availability can also enhance the nutrient content of non-light organs, which will further increase cellular osmotic pressure and reduce water loss in leaves, thereby increasing WUE [38]. Similarly, the effects of P on leaf-scale WUE are mainly due to its influence on the Rubisco carboxylase, chlorophyll content, and corresponding photosynthetic rate [25]. Generally, P in mineral soils can be easily transported to the surface of the fine roots to increase the P concentration in the leaves under transpiration stress, thus causing a positive correlation between leaf P content and transpiration rate [27]. A recent study has proved that an increase in foliar P concentration could indirectly enhance the leaf-scale WUE in a subtropical forest [9]. The results of the above studies both support a positive correlation between WUE and both leaf N and P content, which is consistent with our research (Figure 5). In addition, the leaf N:P ratio was also closely correlated with intrinsic WUE [8,28]. It was found that the photosynthesis rate could reach its optimum value when the leaf N:P ratio was 12. Thus, the leaf N:P ratio was positively correlated with the photosynthesis rate at an N:P ratio < 12; however, the leaf N:P ratio was negatively correlated with the photosynthetic rate at an N:P ratio > 12 [39]. In our study, for most of the species, the leaf N:P ratio was greater than 12; hence, intrinsic WUE was negatively correlated with the leaf N:P ratio (Figure 5). However, it is worth noting that the WUE of three of the tree species (i.e., C. eyrei, S. laurina, and T. gymnanthera) showed significant relationships both with the leaf N content, leaf P content, and leaf N:P ratio, while the WUE of the other two tree species (i.e., M. grijsii and R. ovatum) showed a significant positive correlation with only either the leaf N content or the leaf P content (Figure 5). We found that the increased rates of P for the three tree species were faster than those of N (i.e., the slope values of P were higher than those of N) with increased WUE from the hillside to the peak, meaning that P may be more important for modulating WUE for these tree species in the subtropic forest, which has been well-demonstrated in a recent study [9]. Thus, WUE was negatively correlated with the leaf N:P ratio for the three tree species (Figure 5). On the contrary, the increased rates of N for the other two tree species (i.e., M. grijsii and R. ovatum) were faster or the same as that of P, which would result in no significant correlations between WUE and the leaf N:P ratio for these two tree species. Thus, we suppose that this asynchrony among plants may be caused by differences in the rates of N or P uptake of species, which still requires further research in the future. Overall, our results suggested that N and P and their stoichiometry balance could co-modulate the water utilization for the five evergreen broad-leaved tree species in this region.
5. Conclusions
In this study, we investigated the leaf-scale WUE and leaf nutrients of five tree species (C. eyrei, S. laurina, M. grijsii, T. gymnanthera, and R. ovatum) in three different habitat types (i.e., hillside, near the top of the peak, and peak) on the western slope of the Wuyi Mountain in southern China. From the hillside to the peak, the leaf-scale WUE of the tree species increased gradually with the decrease in soil moisture and the increase in light. The leaf N and P contents showed an increasing trend due to an increase in soil nutrients, whereas the leaf C:N, C:P, and N:P ratios showed a decreasing trend. More importantly, we found that leaf-scale WUE was positively correlated with the leaf N and P contents but was negatively correlated with the leaf N:P ratio, especially for three of the five species (i.e., C. eyrei, S. laurina, and T. gymnanthera). These results indicated that changes in leaf-scale WUE and nutrients of evergreen broad-leaved tree species under different habitat types are mainly driven by differences in soil water availability, light, and soil development sequence, despite there being interspecific differences. It was also emphasized that leaf nutrients can be used as potential proxies to identify the variations in leaf-scale WUE for specific species. In summary, our findings are valuable for understanding the water-, carbon-, and nutrient-coupled relationships for the evergreen broad-leaved tree species, which are important in the face of global changes.
Author Contributions
K.-X.H.: investigation, writing—original draft; Z.-J.X., J.-C.W., H.W., H.-Q.Z., Z.-B.X., W.Z., J.-F.C. and L.-W.H.: investigation and data curation; J.-S.R., S.-S.X. and Y.Z.: methodology and reviewing; F.-X.S.: conceptualization, funding, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 42067049 and 32160357), the Double Thousand Plan of Jiangxi Province (jxsq2023102213), the Project for Forestry Science and Technology Extension Demonstration of Central Government-funded (JXTG(2023)15), the Jiangxi Provincial Natural Science Foundation (20212BAB203027), the Jiangxi Provincial Key Research and Development Program (20202BBGL73097), the Research Project of the Jiangxi Provincial Department of Forestry (CXZX(2022)38), and the Open Research Fund of Jiangxi Province Institute of Water Sciences (Grant Nos. 2022SKTR03 and 2022SKTR05).
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, M.T.; Piao, S.L.; Sun, Y.; Ciais, P.; Cheng, L.; Mao, J.F.; Poulter, B.; Shi, X.Y.; Zeng, Z.Z.; Wang, Y.P. Change in terrestrial ecosystem water-use efficiency over the last three decades. Glob. Chang. Biol. 2015, 21, 2366–2378. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, A.W.; Bitter, N.Q.; Sandquist, D.R.; Ehleringer, J.R. Multidecadal records of intrinsic water-use efficiency in the desert shrub Encelia farinose reveal strong responses to climate change. Proc. Natl. Acad. Sci. USA 2020, 117, 18161–18168. [Google Scholar] [CrossRef]
- Wang, X.; Chen, G.; Wu, M.Q.; Li, X.Z.; Wu, Q.; Wang, P.; Zeng, H.; Yang, R.; Tang, X.L. Differences in the patterns and mechanisms of leaf and ecosystem-scale water use efficiencies on the Qinghai-Tibet Plateau. Catena 2023, 222, 106874. [Google Scholar] [CrossRef]
- Ma, W.T.; Tcherkez, G.; Wang, X.M.; Schäufele, R.; Schnyder, H.; Yang, Y.; Gong, X.Y. Accounting for mesophyll conductance substantially improves 13C-based estimates of intrinsic water-use efficiency. New. Phytol. 2021, 229, 1326–1338. [Google Scholar] [CrossRef]
- Feng, X.H. Long-term ci/ca response of trees in western North America to atmospheric CO2 concentration derived from carbon isotope chronologies. Oecologia 1998, 117, 19–25. [Google Scholar] [CrossRef]
- Ale, R.; Zhang, L.; Li, X.; Raskoti, B.B.; Pugnaire, F.I.; Luo, T.X. Leaf δ13C as an indicator of water availability along elevation gradients in the dry Himalayas. Ecol. Indic. 2018, 94, 266–273. [Google Scholar] [CrossRef]
- ÅGREN, G.I. The C: N: P stoichiometry of autotrophs–theory and observations. Ecol. Lett. 2004, 7, 185–191. [Google Scholar] [CrossRef]
- Yan, Z.B.; Kim, N.; Han, W.X.; Guo, Y.L.; Han, T.S.; Du, E.Z.; Fang, J.Y. Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant Soil. 2015, 388, 147–155. [Google Scholar] [CrossRef]
- Hang, Z.Q.; Ran, S.S.; Fu, Y.R.; Wan, X.H.; Song, X.; Chen, Y.X.; Yu, Z.P. Functionally dissimilar neighbours increase tree water use efficiency through enhancement of leaf phosphorus concentration. J. Ecol. 2022, 110, 2179–2189. [Google Scholar] [CrossRef]
- Hessen, D.O.; Ågrer, G.I.; Anerson, T.R.; Elser, J.J.; Ruiter, P.C. Carbon sequestration in ecosystems: The role of stoichiometry. Ecology 2004, 85, 1179–1192. [Google Scholar] [CrossRef]
- Tian, J.Y.; Yuan, F.H.; Guan, D.X.; Wu, J.B.; Wang, A.Z. Water use efficiency and leaf nutrient characteristics of five major tree species in broadleaved Korean pine forest in Changbai Mountains, China. Chin. J. Appl. Ecol. 2022, 33, 304–310, (In Chinese with English Abstract). [Google Scholar]
- Zheng, Z.M.; Lu, J.; Su, Y.Q.; Yang, Q.S.; Lin, Y.H.; Liu, H.M.; Yang, J.; Huang, H.; Wang, X.H. Differential effects of N and P additions on foliar stoichiometry between species and community levels in a subtropical forest in eastern China. Ecol. Indic. 2020, 117, 106537. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef]
- Rumman, R.; Atkin, O.K.; Bloomfield, K.J.; Eamus, D. Variation in bulk-leaf 13C discrimination, leaf traits and water-use efficiency-trait relationships along a continental-scale climate gradient in Australia. Glob. Chang. Biol. 2018, 24, 1186–1200. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Su, Y.G.; Li, Y.; Huang, G. Shrub colonization regulates δ13Cp enrichment between soil and vegetation in deserts by affecting edaphic variables. Catena 2021, 203, 105365. [Google Scholar] [CrossRef]
- Zhang, Y.E.; Yu, X.X.; Chen, L.H.; Jia, G.D.; Zhao, N.; Li, H.Z.; Chang, X.M. Foliar water use efficiency of Platycladus orientalis sapling under different soil water contents. Chin. J. Appl. Ecol. 2017, 28, 2149–2154, (In Chinese with English Abstract). [Google Scholar]
- Huang, F.Z.; Li, D.X.; Wang, B.; Xiang, W.S.; Guo, Y.L.; Wen, S.J.; Chen, T.; Li, X.K. Foliar stable carbon isotope composition and water use efficiency of plant in the Karst seasonal rain forest. Chin. J. Appl. Ecol. 2019, 30, 1833–1839, (In Chinese with English Abstract). [Google Scholar]
- Gong, X.Y.; Ma, W.T.; Yu, Y.Z.; Fang, K.Y.; Yang, Y.S.; Tcherkez, G.; Adams, M.A. Overestimated gains in water-use efficiency by global forests. Glob. Chang. Biol. 2022, 28, 4923–4934. [Google Scholar] [CrossRef]
- Lu, J.N.; Zhao, X.Y.; Wang, S.K.; Feng, S.; Ning, Z.Y.; Wang, R.X.; Chen, X.P.; Zhao, H.S.; Chen, M. Untangling the influence of abiotic and biotic factors on leaf C, N, and P stoichiometry along a desert-grassland transition zone in northern China. Sci. Total. Environ. 2023, 884, 163902. [Google Scholar] [CrossRef]
- Li, L.; Liu, L.; Yu, Z.; Peñuelas, J.; Sardans, J.; Chen, Q.F.; Xu, J.B.; Zhou, G.Y. Carbon, nitrogen and phosphorus stoichiometry in natural and plantation forests in China. Forests 2022, 13, 755. [Google Scholar] [CrossRef]
- Kong, L.L.; Lin, J.; Huang, Z.Q.; Yu, Z.P.; Xu, Z.K.; Liang, Y.F. Variations of water use efficiency and its relationship with leaf nutrients of different altitudes of Wuyi Mountains, China. Chin. J. Appl. Ecol. 2017, 28, 2102–2110, (In Chinese with English Abstract). [Google Scholar]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Funct. Plant. Biol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Adiredjo, A.L.; Navaud, O.; Muños, S.; Langlade, N.B.; Lamaze, T.; Grieu, P. Genetic control of water use efficiency and leaf carbon isotope discrimination in sunflower (Helianthus annuus L.) subjected to two drought scenarios. PLoS ONE 2014, 9, e101218. [Google Scholar] [CrossRef]
- Talbi Zribi, O.; Abdelly, C.; Debez, A. Interactive effects of salinity and phosphorus availability on growth, water relations, nutritional status and photosynthetic activity of barley (Hordeum vulgare L.). Plant Biology 2011, 13, 872–880. [Google Scholar] [CrossRef]
- Jacob, J.; Lawlor, D.W. Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower, maize and wheat plants. J. Exp. Bot. 1991, 42, 1003–1011. [Google Scholar] [CrossRef]
- Chen, Y.H.; Han, W.X.; Tang, L.Y.; Tang, Z.Y.; Fang, J.Y. Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography 2013, 36, 178–184. [Google Scholar] [CrossRef]
- Huang, Z.Q.; Liu, B.; Davis, M.; Sardans, J.; Peñuelas, J.; Billings, S. Long-term nitrogen deposition linked to reduced water use efficiency in forests with low phosphorus availability. New. Phytol. 2016, 210, 431–442. [Google Scholar] [CrossRef]
- Zeng, H.H.; Wu, J.E.; Liu, W.J. Water use efficiency and leaf nutrient contents of plants in jungle rubber agroforestry system. Subtrop. Plant Sci. 2019, 48, 125–133, (In Chinese with English Abstract). [Google Scholar]
- Wu, Z.Y. Vegetation of China; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]
- Chen, H.M.; Shi, F.X.; Xu, J.W.; Liu, X.P.; Mao, R. Tree mycorrhizal type controls over soil water-extractable organic matter quantity and biodegradation in a subtropical forest of southern China. Forest. Ecol. Manag. 2023, 535, 120900. [Google Scholar] [CrossRef]
- Hanba, Y.T.; Mori, S.; Lei, T.T.; Koike, T.; Wada, E. Variations in leaf δ13C along a vertical profile of irradiance in a temperate Japanese forest. Oecologia 1997, 110, 253–261. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J. The increasing importance of distinguishing among plant nitrogen sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef]
- Cui, E.Q.; Lu, R.L.; Xu, X.N.; Sun, H.F.; Qiao, Y.; Ping, J.Y.; Qiu, S.Y.; Lin, Y.H.; Bao, J.H.; Yong, Y.T.; et al. Soil phosphorus drives plant trait variations in a mature subtropical forest. Glob. Chang. Biol. 2022, 28, 3310–3320. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.P.; Bustamante, M.M. Controls over foliar N:P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.D.; Greer, D.H.; Laing, W.A.; McManus, M.T. Changes in photosynthetic efficiency and carotenoid composition in leaves of white clover at different developmental stages. Plant Physiol. Bioch. 2003, 41, 887–893. [Google Scholar] [CrossRef]
- Shangguan, Z.P.; Shao, M.A.; Dyckmans, J. Nitrogen nutrition and water stress effects on leaf photosynthetic gas exchange and water use efficiency in winter wheat. Environ. Exp. Bot. 2000, 44, 141–149. [Google Scholar] [CrossRef]
- Garrish, V.; Cernusak, L.A.; Winter, K.; Turner, B.L. Nitrogen to phosphorus ratio of plant biomass versus soil solution in a tropical pioneer tree, Ficus insipida. J. Exp. Bot. 2010, 61, 3735–3748. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).