
Comparison of the Stand Structure Diversity of Open Pinus brutia Ten. Forests in Areas of Different Productivity in Central Cyprus



Abstract
:1. Introduction
2. Materials and Methods
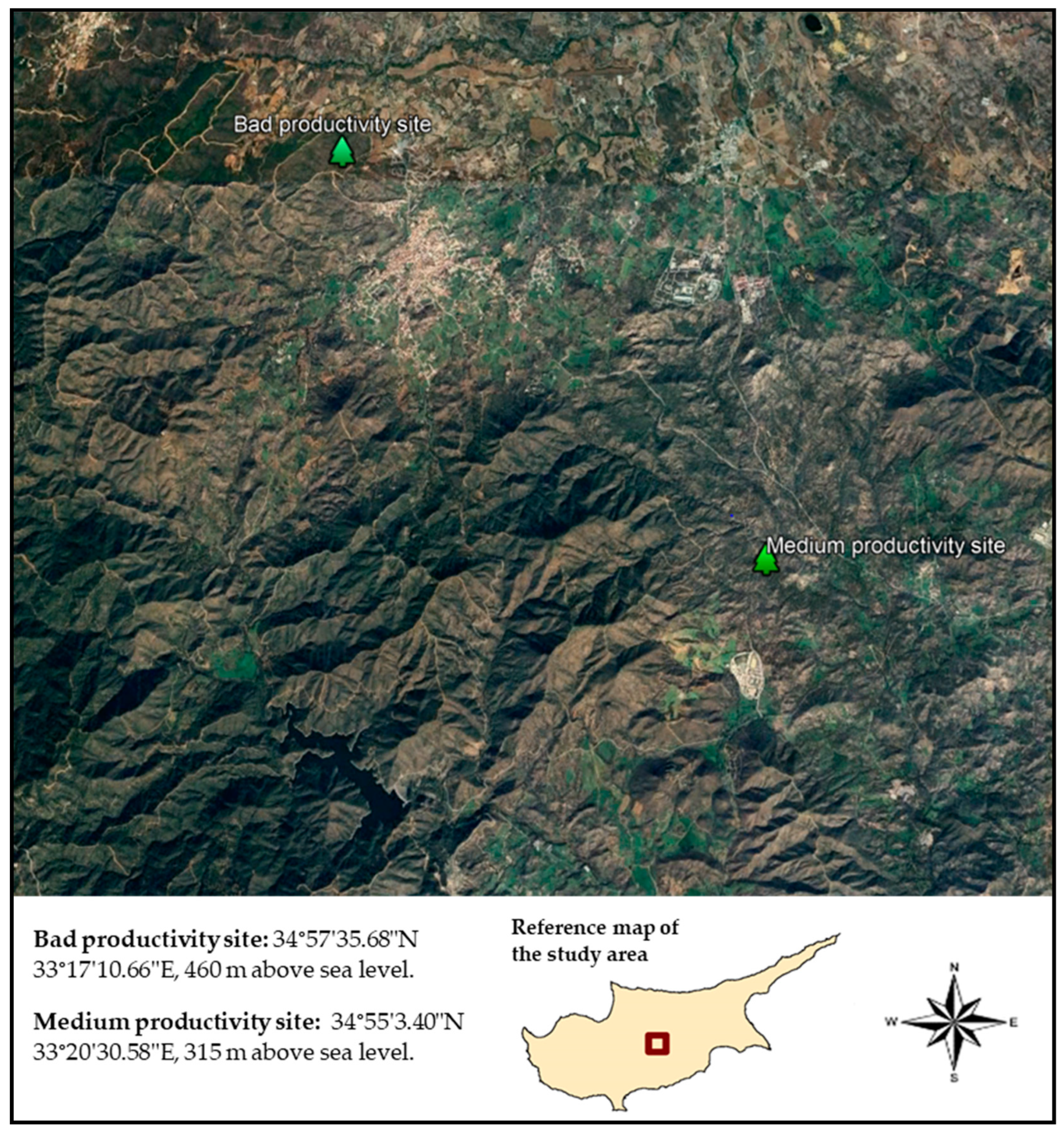
2.1. Study Area
2.2. Research Method
2.3. Statistical Analysis
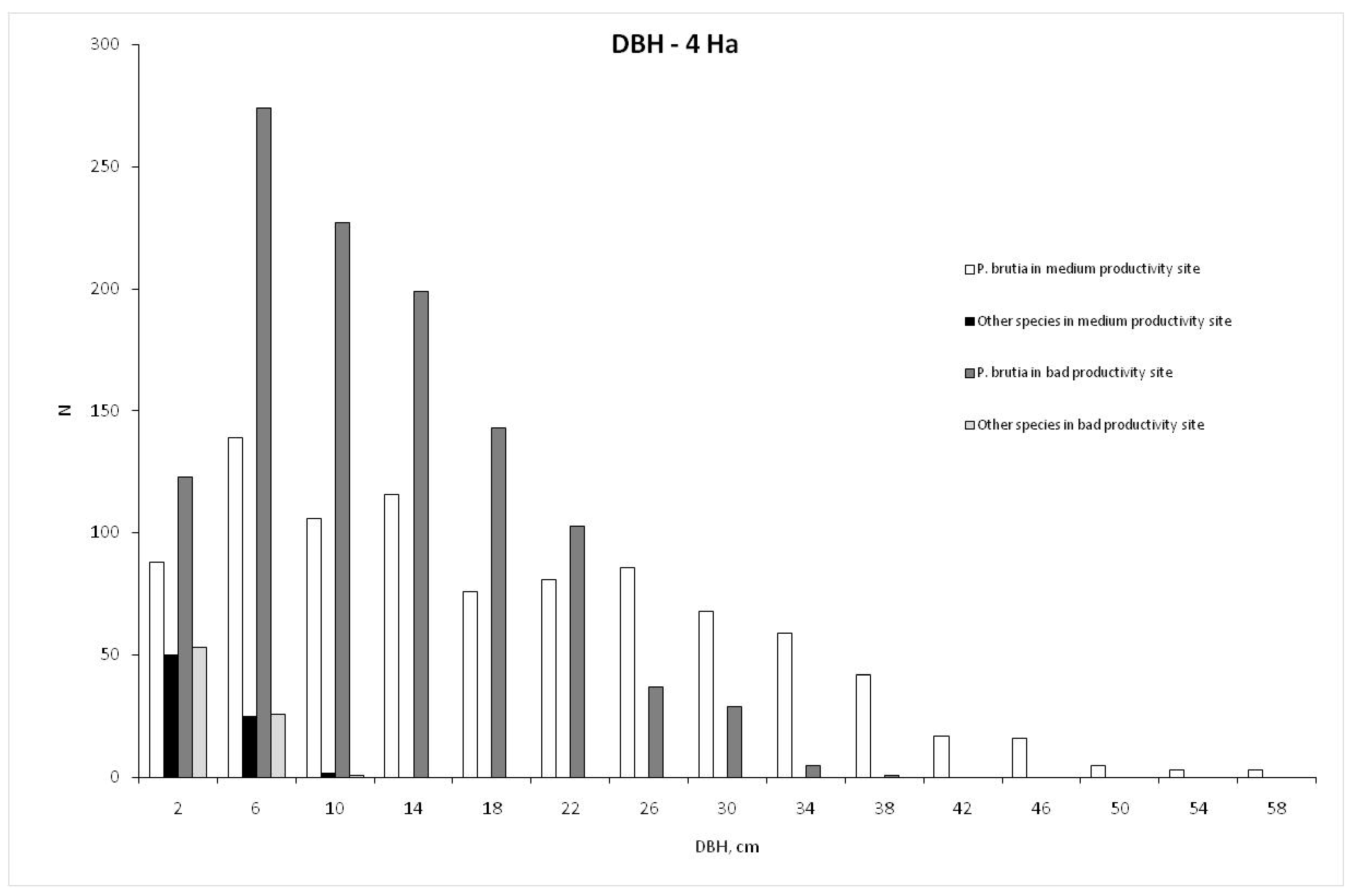
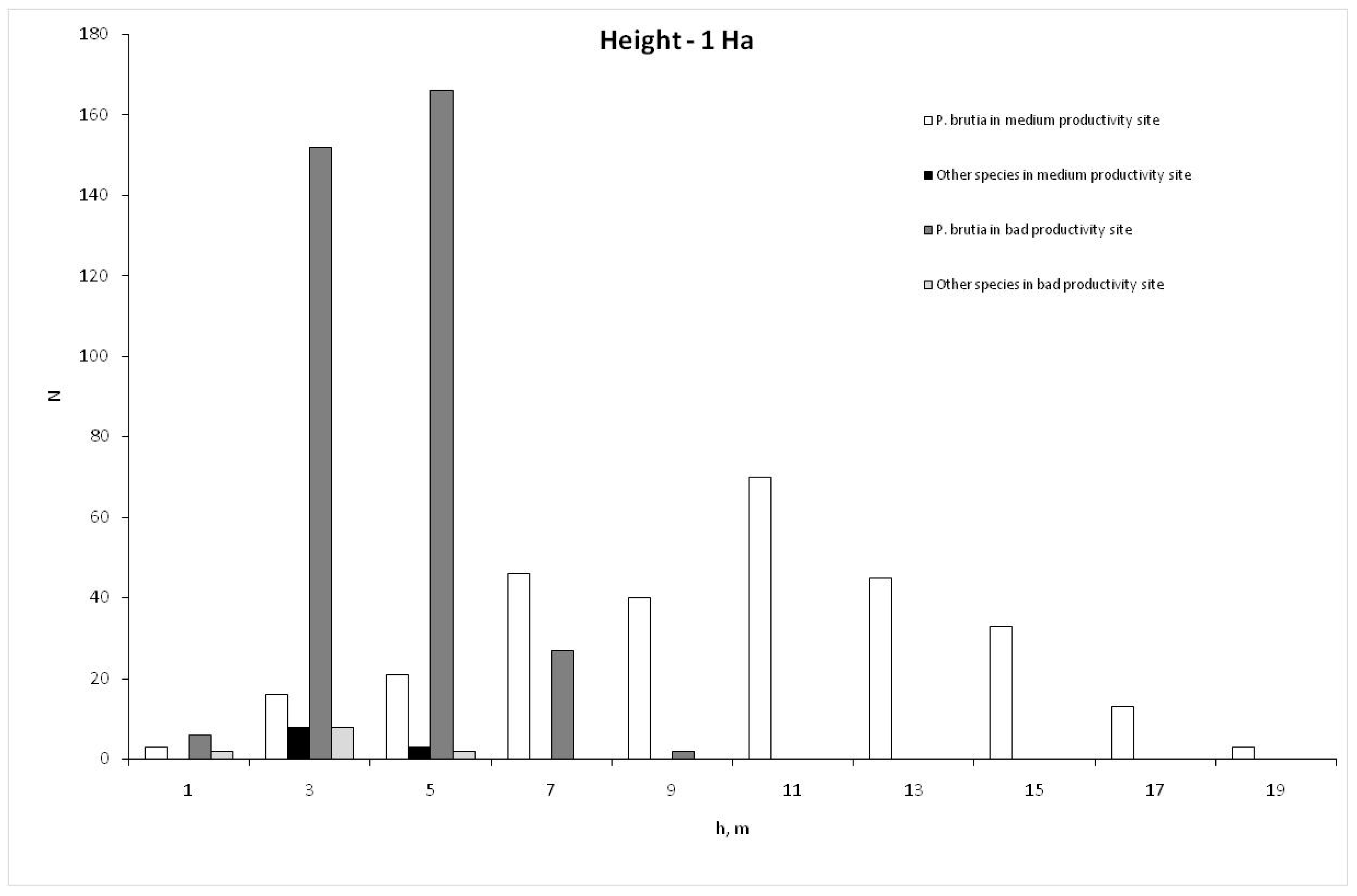
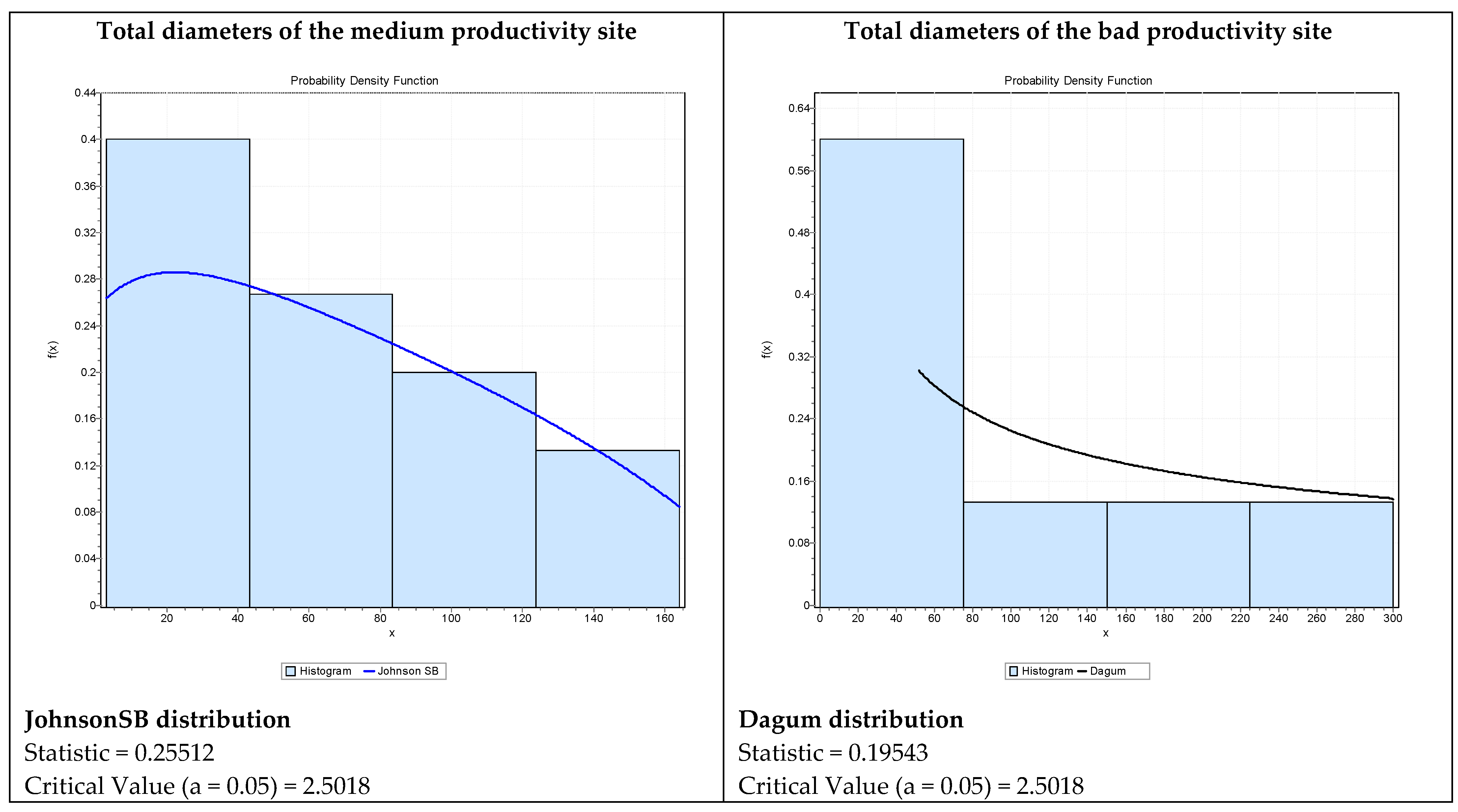
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milios, E. Facilitation process and sprouting ability as silvicultural tools in the frame of climate change. In Forest Ecosystems and Climate Changes, Proceedings of the Plenary Lectures of the International Scientific Conference: Forest Ecosystems and Climate Changes, Belgrade, Serbia, 9–10 March 2010; Institute of Forestry: Belgrade, Serbia, 2010; pp. 137–142. [Google Scholar]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley & Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.; Mark, S. The Practice of Silviculture, 9th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F. Conserving Forest Biodiversity: A Comprehensive Multiscaled Approach; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Dafis, S. Forest Ecology; Giahoudis-Giapoulis: Thessaloniki, Greece, 1986. [Google Scholar]
- Barnes, B.V.; Zak, D.R.; Denton, S.R.; Spurr, S.H. Forest Ecology, 4th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1998. [Google Scholar]
- O’Hara, K. Multiaged Silviculture: Managing for Complex Forest Stand Structures; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Dafis, S. Applied Silviculture; Giahoudis-Giapoulis: Thessaloniki, Greece, 1992. [Google Scholar]
- Richardson, D.M.; Rundel, P.W. Ecology and Biogeography of Pinus: An Introduction. In Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998; pp. 3–40. [Google Scholar]
- Demiguel, S. Growth and Yield Modelling for Optimal Multi-Objective Forest Management of Eastern Mediterranean Pinus brutia. Diss. For. 2014, 170, 59. [Google Scholar] [CrossRef]
- Boydak, M. Silvicultural Characteristics and Natural Regeneration of Pinus brutia Ten.—A Review. Plant Ecol. 2004, 171, 153–163. [Google Scholar] [CrossRef]
- Boydak, M.; Dirik, H.; Çalıkoğlu, M. Forest Stand Dynamic, Regeneration and Fire in Pinus brutia Ecosystems. In Biology and Silviculture of Turkish Red Pine (Pinus brutia Ten.); Istanbul Universitesi Orman Fakultesi: Ankara, Turkey, 2006. [Google Scholar]
- Quézel, P. Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Neeman, L., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 1–12. [Google Scholar]
- Gezer, A. The sylviculture of Pinus brutia in Turkey. In Le Pin d’Alep et le Pin Brutia Dans la Sylviculture Méditerranéenne; CIHEAM: Paris, France, 1986; pp. 55–66. [Google Scholar]
- Fischer, R.; Lorenz, M.; Köhl, M.; Becher, G.; Granke, O.; Christou, A. The Condition of Forests in Europe: 2008 Executive Report; United Nations Economic Commission for Europe, Convention on Long-Range Transboundary Air Pollution: Hamburg, Germany, 2008; p. 23. [Google Scholar]
- Paionidis, L. Forestry in Cyprus (115 Years of Contribution to the Country 1879–1994); Ministry of Agriculture, Natural Resources and Environment, Department of Forestry of Cyprus: Aglantzia, Cyprus, 1994.
- Thirgood, J.V. Cyprus: A Chronicle of Its Forests, Land and People; University of British Columbia Press: Vancouver, BC, Canada, 1987; ISBN 978-0-7748-0268-0. [Google Scholar]
- Forests Department of Cyprus. Mapping of Woody Forest Vegetation in Cyprus; Forests Department of Cyprus: Nicosia, Cyprus, 2020.
- Meikle, D.R. Flora of Cyprus; Bentham—Moxon Trust Royal Botanic Gardens Kew: Richmond, UK, 1977; Volume I, p. 832. [Google Scholar]
- Greuter, W.; Burdet, H.M.; Long, G. Med-Checklist; Conservatoire et Jardin botaniques de la Ville de Genève: Geneva, Switzerland, 1984; Volume 4, Available online: http://ww2.bgbm.org/mcl/ (accessed on 8 August 2023).
- Pantelas, V. The forest of brutia pine in Cyprus. Options Mediterr. Ser. Stud. 1986, 1, 43–46. [Google Scholar]
- Tsintidis, T.; Hadjikyriakos, G.N.; Christodoulou, C.S. Trees and Shrubs in Cyprus; Cyprus Forest Association: Nicosia, Cyprus, 2002; p. 442. [Google Scholar]
- FAO. Global Forest Resource Assessment 2010; Report Cyprus; Food and Agriculture Organization of the United Nation: Rome, Italy, 2010. [Google Scholar]
- Valladares, F.; Pearcy, R.W. Drought Can Be More Critical in the Shade than in the Sun: A Field Study of Carbon Gain and Photo-Inhibition in a Californian Shrub during a Dry El Niño Year. Plant Cell Environ. 2002, 25, 749–759. [Google Scholar] [CrossRef]
- Petrou, P.; Milios, E. Investigation of the Factors Affecting Artificial Seed Sowing Success and Seedling Survival in Pinus brutia Natural Stands in Middle Elevations of Central Cyprus. Forests 2020, 11, 1349. [Google Scholar] [CrossRef]
- Geological Survey Department of Cyprus. Geological Map of Cyprus, Revised ed.; Press and Information Office: Nicosia, Cyprus, 1995.
- Petrou, P. Regeneration, Growth and Structure of Calabrian Pine (Pinus brutia Ten.) Stands in the Middle Elevations of Central Cyprus. Ph.D. Thesis, Democritus University of Thrace, Orestiada, Greece, 2015. [Google Scholar]
- Sagheb-Talebi, K.; Schütz, J.-P. The Structure of Natural Oriental Beech (Fagus orientalis) Forests in the Caspian Region of Iran and Potential for the Application of the Group Selection System. Forestry 2002, 75, 465–472. [Google Scholar] [CrossRef]
- Husch, B.; Miller, C.I.; Beers, T.W. Forest Mensuration; John Wiley & Sons, Inc.: New York, NY, USA, 1982. [Google Scholar]
- Kitikidou, K.; Petrou, P.; Milios, E. Dominant height growth and site index curves for Calabrian pine (Pinus brutia Ten.) in central Cyprus. Renew. Sustain. Energy Rev. 2012, 16, 1323–1329. [Google Scholar] [CrossRef]
- Delta-T Devices Ltd. HemiView User Manual; Delta-T Devices Ltd.: Cambridge, UK, 1999. [Google Scholar]
- Hale, S.E. Light regime beneath Sitka spruce plantations in northern Britain: Preliminary results. For. Ecol. Manag. 2001, 151, 61–66. [Google Scholar] [CrossRef]
- Montgomery, R.A. Effects of understory foliage on patterns of light attenuation near the forest floor. Biotropica 2004, 36, 33. [Google Scholar] [CrossRef]
- Rényi, A. On Measures of Information and Entropy. In Proceedings of the 4th Berkeley Symposium on Mathematics, Statistics and Probability, Berkeley, CA, USA, 20 June–30 July 1960; University of California Press: Berkeley, CA, USA, 1961; pp. 547–561. [Google Scholar]
- Jost, L. Entropy and Diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Kitikidou, K.; Milios, E.; Stampoulidis, A.; Pipinis, E.; Radoglou, K. Within-Forest Stand (or Formation, or Plot) and between-Forest Stand (or Formation, or Plot) Biodiversity Indices. MethodsX 2022, 9, 101919. [Google Scholar] [CrossRef] [PubMed]
- Schittkowski, K. EASY-FIT: A Software System for Data Fitting in Dynamical Systems. Struct. Multidiscip. Optim. 2002, 23, 153–169. [Google Scholar] [CrossRef]
- Anderson, T.W.; Darling, D.A. A Test of Goodness of Fit. J. Am. Stat. Assoc. 1954, 49, 765–769. [Google Scholar] [CrossRef]
- IBM Corp. Released IBM SPSS Statistics for Windows, Version 21.0; IBM Corp: Armonk, NY, USA, 2012. [Google Scholar]
- Bourma, K.; Milios, E.; Radoglou, K.; Kitikidou, K. Development of a Graded Biodiversity Assessment (GBA) Index for the Assessment of the Biodiversity of Managed Natural Forests. Ecologies 2023, 4, 614–626. [Google Scholar] [CrossRef]
- Stampoulidis, A.; Pipinis, E.; Petrou, P.; Kitikidou, K. Regeneration Analysis of the Juniperus Excelsa Mixed Stands in Prespa National Park of Greece as a Base for the Assessment of the Appropriate Silvicultural Treatment for the Conservation of the Species. South-East Eur. For. 2023, 14, 47–52. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
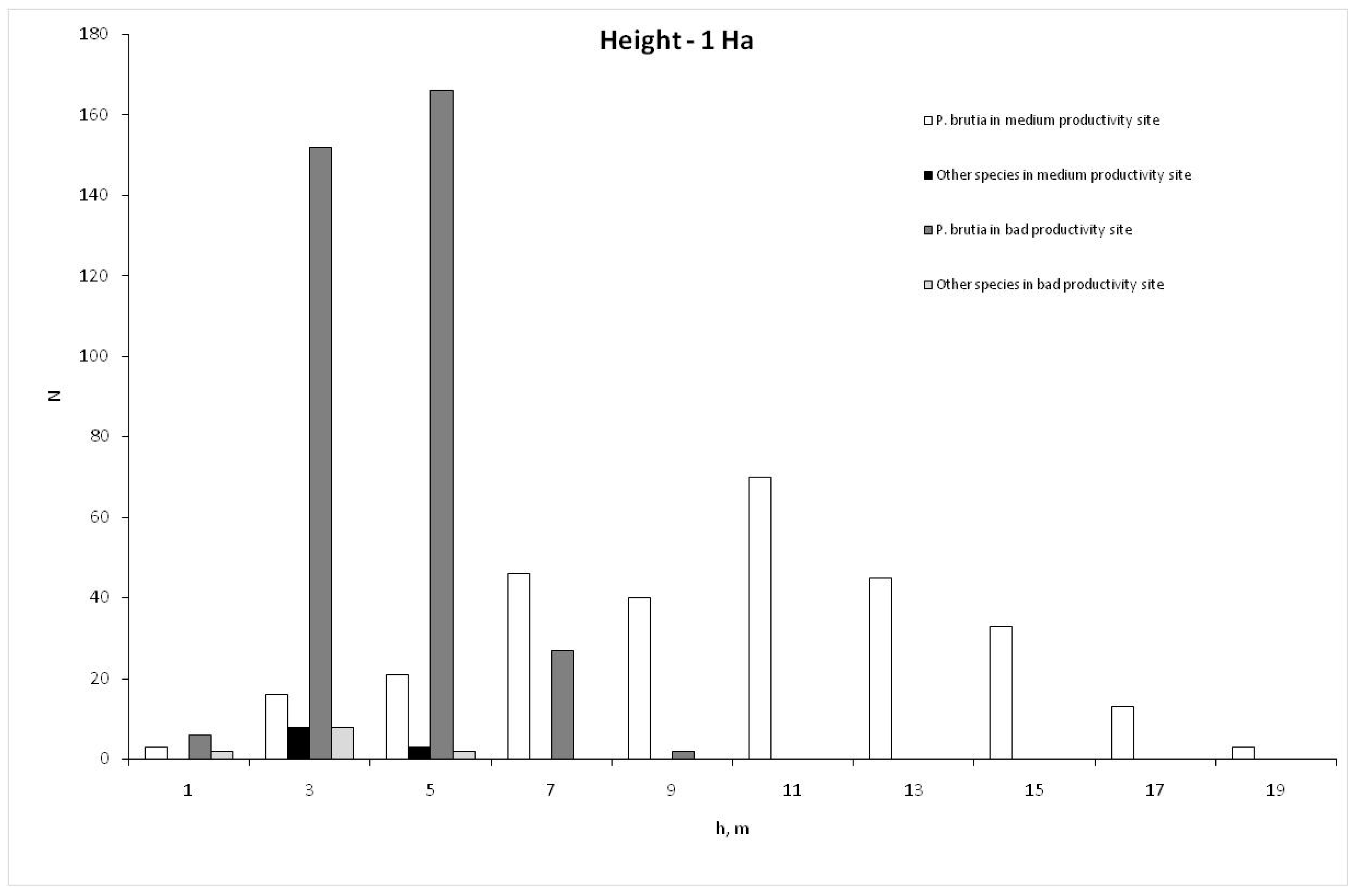{kind=link}
{kind=link}
{kind=link}
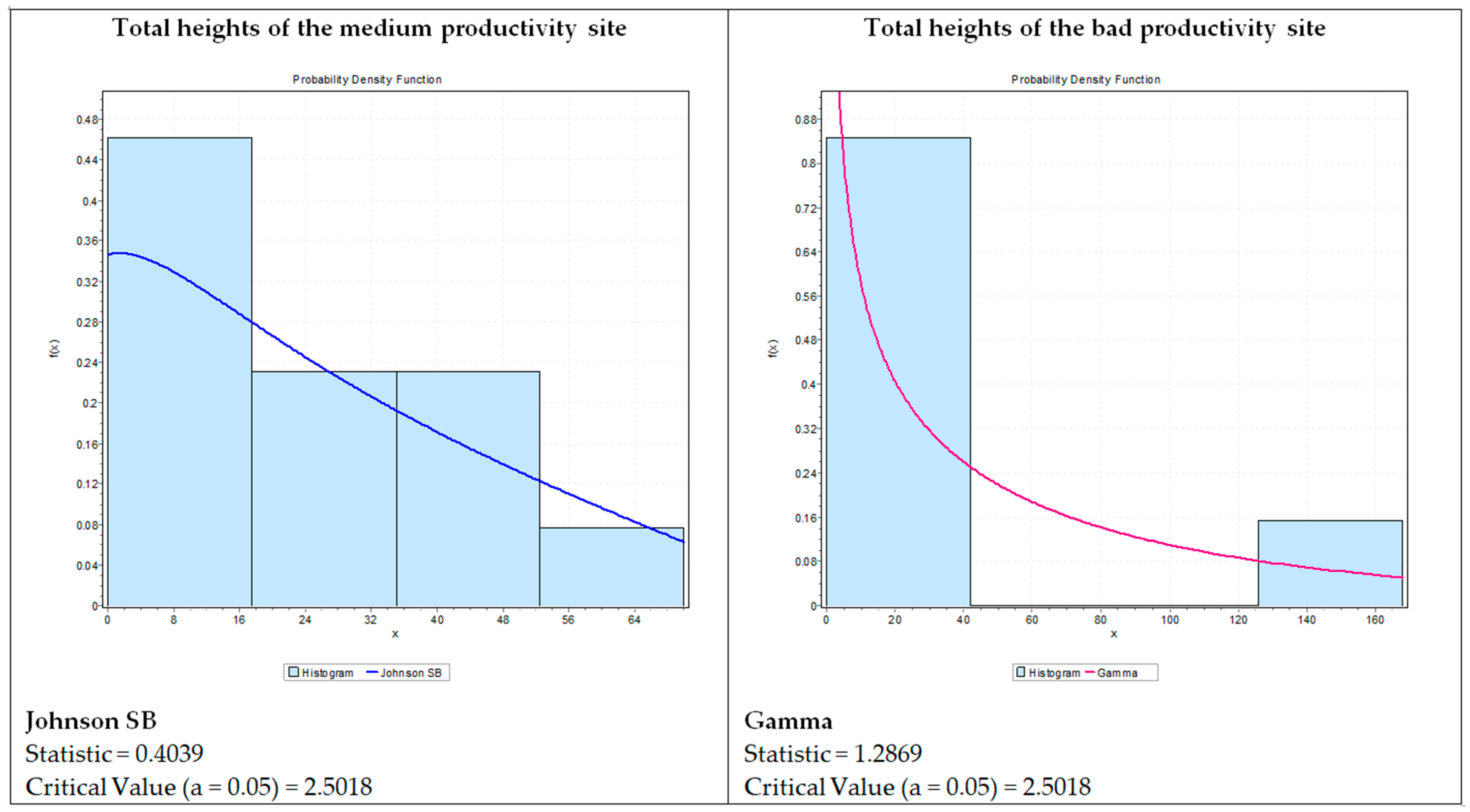{kind=link}
| DBH Range (cm) | Age Range (Years) | |
|---|---|---|
| Medium Productivity Site | Bad Productivity Site | |
| 0–8 | 40–65 | 43–68 |
| 8–16 | 56–65 | 44–90 |
| 16–24 | 62–75 | 75–96 |
| 24–32 | 77–90 | 78–94 |
| 32–40 | 81–102 | 85–98 |
| 40–48 | 108–125 | – |
| 48–56 | 107–115 | – |
| 56–64 (58.3 *) | 100–119 | – |
| Medium Productivity Site | ||||||||||
| Species | DBH (cm) | Height (m) | Basal Area (m2/ha) | Density (n/ha) | ||||||
| Mean | S.D. | Min. | Max. | Mean | S.D. | Min. | Max. | Total | Total | |
| P. brutia | 18.49 | 12.614 | 1.0 | 58.3 | 9.85 | 3.928 | 1.3 | 18.0 | 8.740 | 227 |
| Other species | 3.70 | 12.602 | 1.0 | 9.6 | 3.26 | 3.903 | 2.0 | 5.5 | 0.027 | 19 |
| Bad Productivity Site | ||||||||||
| Species | DBH (cm) | Height (m) | Basal Area (m2/ha) | Density (n/ha) | ||||||
| Mean | S.D. * | Min. | Max. | Mean | S.D. * | Min. | Max. | Total | Total | |
| P. brutia | 12.10 | 7.382 | 0.6 | 38.2 | 4.05 | 1.374 | 1.7 | 8.2 | 4.407 | 285 |
| Other species | 3.49 | 7.366 | 1.0 | 9.5 | 2.28 | 1.421 | 1.60 | 3.0 | 0.024 | 20 |
| Order q | DBH | Height | ||||||
|---|---|---|---|---|---|---|---|---|
| Proportions | z- Value | p- Value | Bad Site’s Diversity, Compared with the Medium Site’s Diversity | Proportions | z- Value | p- Value | Bad Site’s Diversity, Compared with the Medium Site’s Diversity | |
| 0 | 0.600 | 9.334 | 0.000 | Significantly Decreasing | 0.667 | 8.567 | 0.000 | Significantly Decreasing |
| 0.400 | 0.333 | |||||||
| 1 | 0.430 | 7.785 | 0.000 | Significantly Decreasing | 0.519 | 9.061 | 0.000 | Significantly Decreasing |
| 0.272 | 0.186 | |||||||
| 2 | 0.381 | 7.018 | 0.000 | Significantly Decreasing | 0.462 | 8.412 | 0.000 | Significantly Decreasing |
| 0.243 | 0.163 | |||||||
| 3 | 0.354 | 6.561 | 0.000 | Significantly Decreasing | 0.428 | 7.786 | 0.000 | Significantly Decreasing |
| 0.227 | 0.156 | |||||||
| 4 | 0.354 | 6.561 | 0.000 | Significantly Decreasing | 0.428 | 7.786 | 0.000 | Significantly Decreasing |
| 0.227 | 0.156 | |||||||
| ∞ | 0.240 | 4.500 | 0.000 | Significantly Decreasing | 0.287 | 4.481 | 0.000 | Significantly Decreasing |
| 0.163 | 0.145 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrou, P.; Stampoulidis, A.; Kitikidou, K.; Milios, E. Comparison of the Stand Structure Diversity of Open Pinus brutia Ten. Forests in Areas of Different Productivity in Central Cyprus. Forests 2023, 14, 2200. https://doi.org/10.3390/f14112200
Petrou P, Stampoulidis A, Kitikidou K, Milios E. Comparison of the Stand Structure Diversity of Open Pinus brutia Ten. Forests in Areas of Different Productivity in Central Cyprus. Forests. 2023; 14(11):2200. https://doi.org/10.3390/f14112200
Chicago/Turabian StylePetrou, Petros, Athanasios Stampoulidis, Kyriaki Kitikidou, and Elias Milios. 2023. "Comparison of the Stand Structure Diversity of Open Pinus brutia Ten. Forests in Areas of Different Productivity in Central Cyprus" Forests 14, no. 11: 2200. https://doi.org/10.3390/f14112200
APA StylePetrou, P., Stampoulidis, A., Kitikidou, K., & Milios, E. (2023). Comparison of the Stand Structure Diversity of Open Pinus brutia Ten. Forests in Areas of Different Productivity in Central Cyprus. Forests, 14(11), 2200. https://doi.org/10.3390/f14112200