

Effects of Elevated CO2 Concentration and Temperature on the Growth and Photosynthetic Characteristics of Populus simonii × P. nigra ‘1307’ Leaves


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Determination of the Tree Growth Status
2.4. Data Analysis
3. Results
3.1. Effect of CO2 Concentration and Temperature on Populus simonii × P. nigra ‘1307’ Leaves Biomass
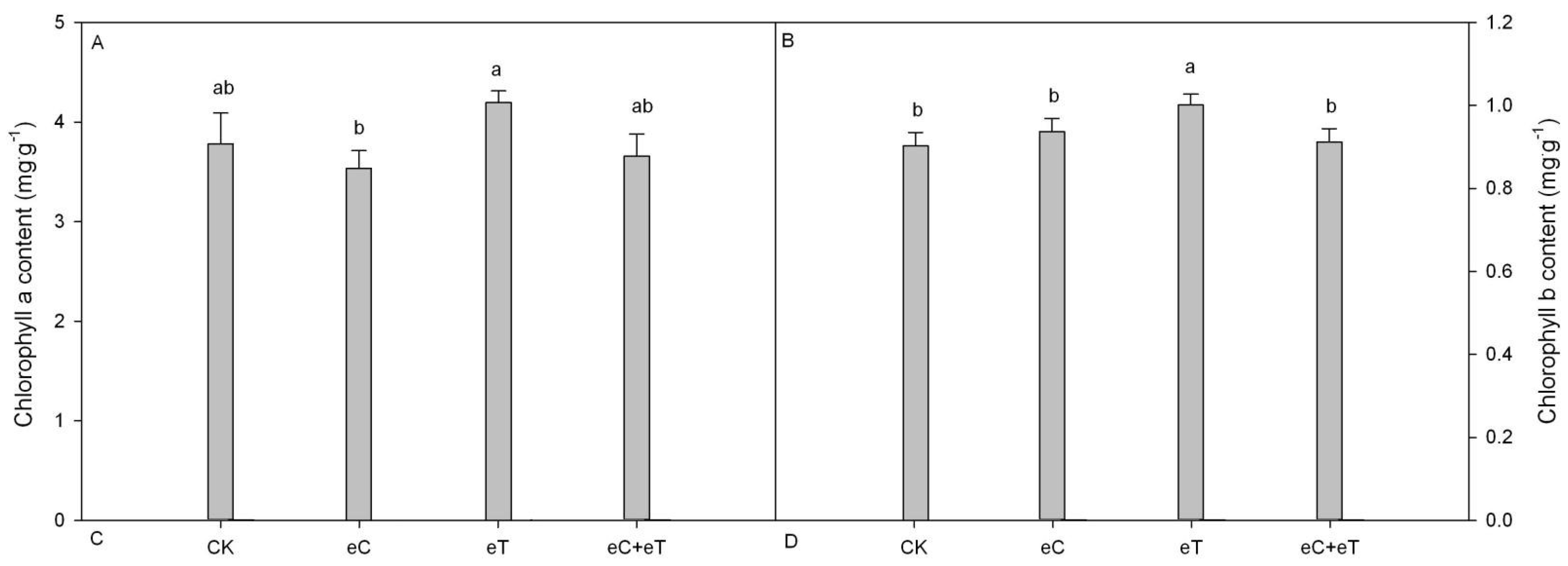
3.2. Effects of CO2 Concentration and Temperature on Chlorophyll Parameters on Populus simonii × P. nigra ‘1307’ Leaves
3.3. CO2 Concentration and Temperature Effects on Photosynthetic Parameters and Dry Matter Allocation in Populus simonii × P. nigra ‘1307’ Functional Leaves
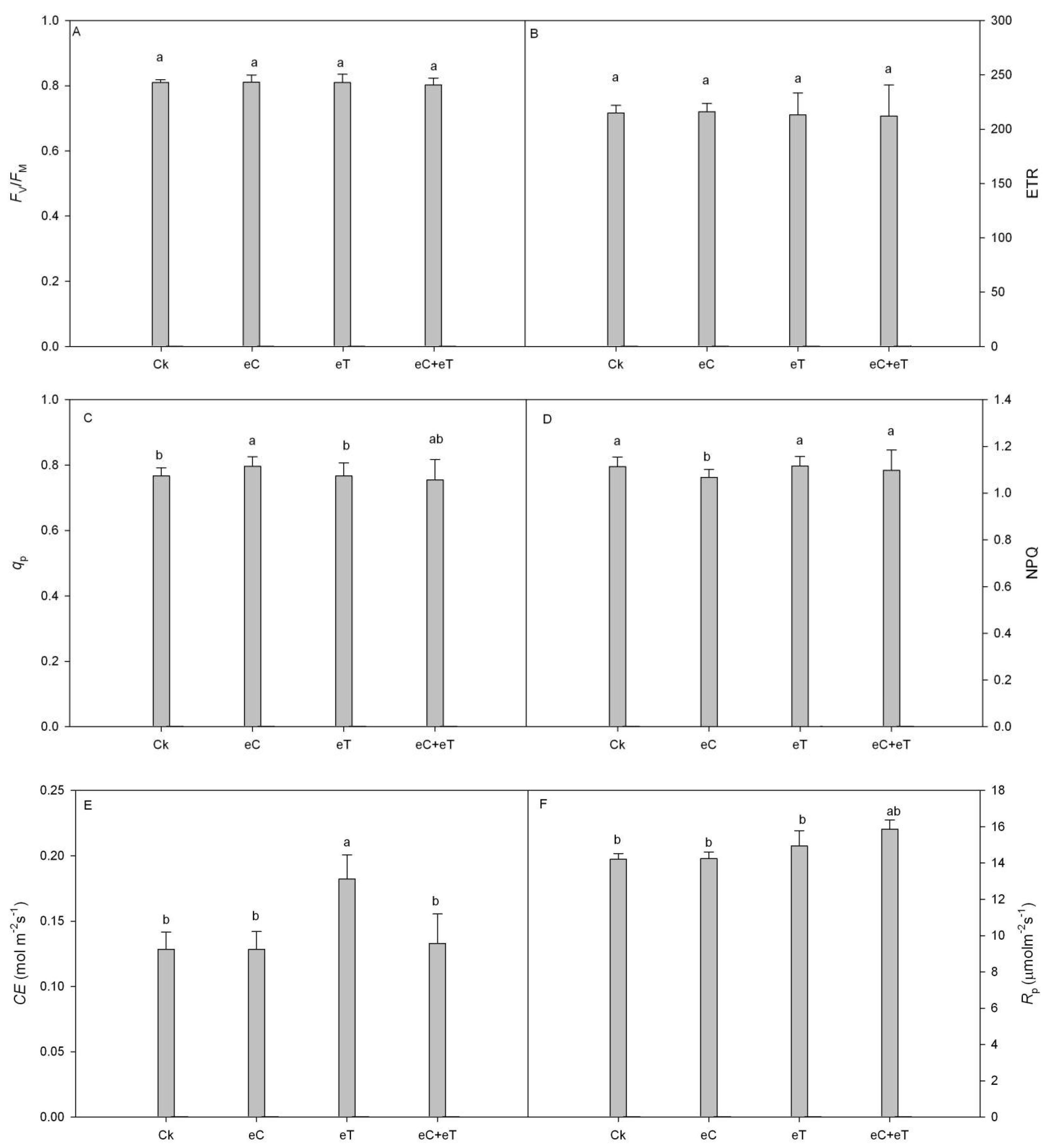
3.4. Effects of CO2 Concentration and Temperature on Chlorophyll Fluorescence Parameters of Functional Leaves
3.5. Effects of High CO2 Concentration and Elevated Temperature on the Activity of PSII Reaction Center in Populus simonii × P. nigra ‘1307’ Leaves
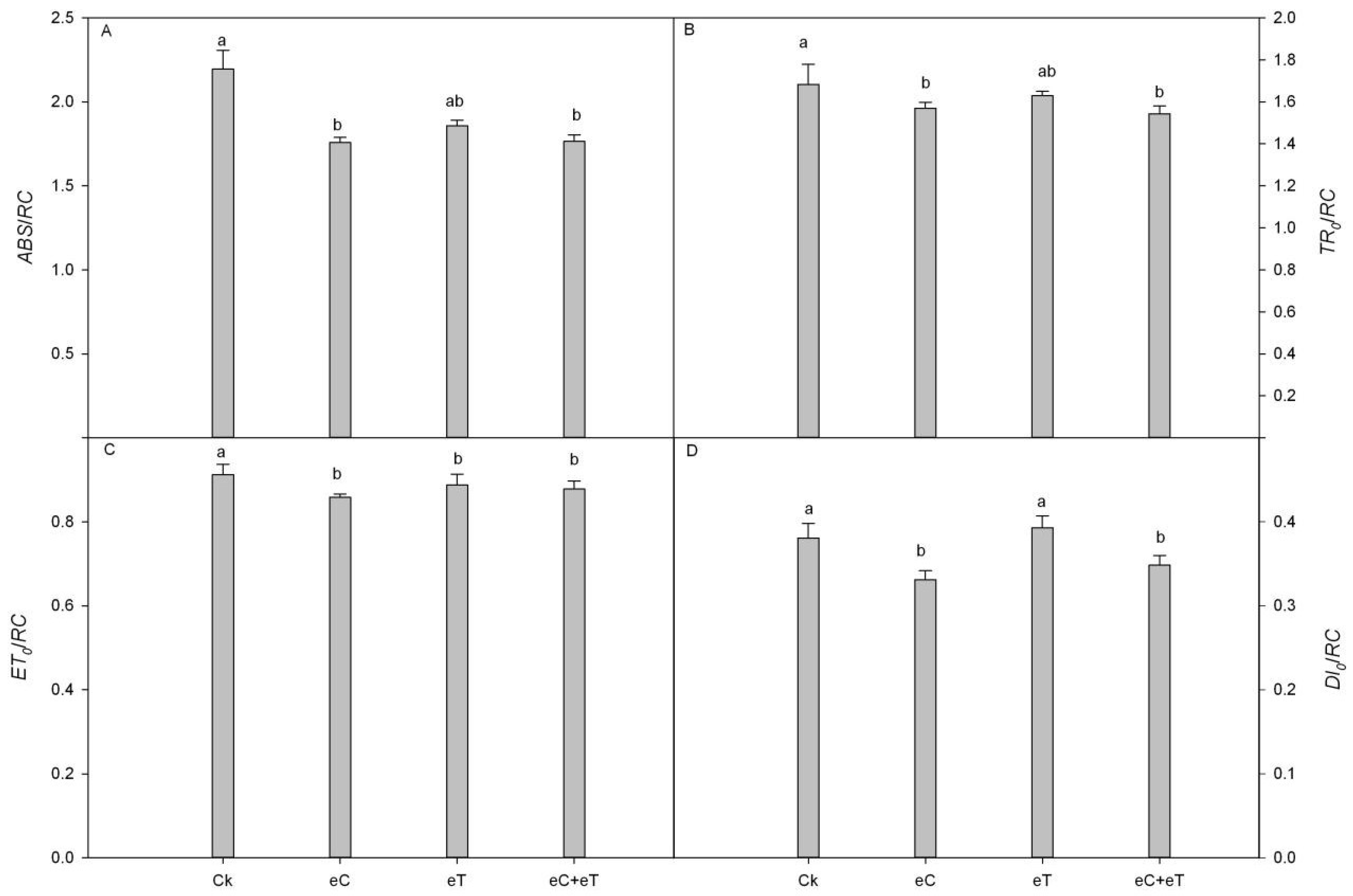
3.6. Effects of Elevated CO2 Concentration and Temperature on Donor and Receptor Sides Parameters of PSII in Populus simonii × P. nigra ‘1307’
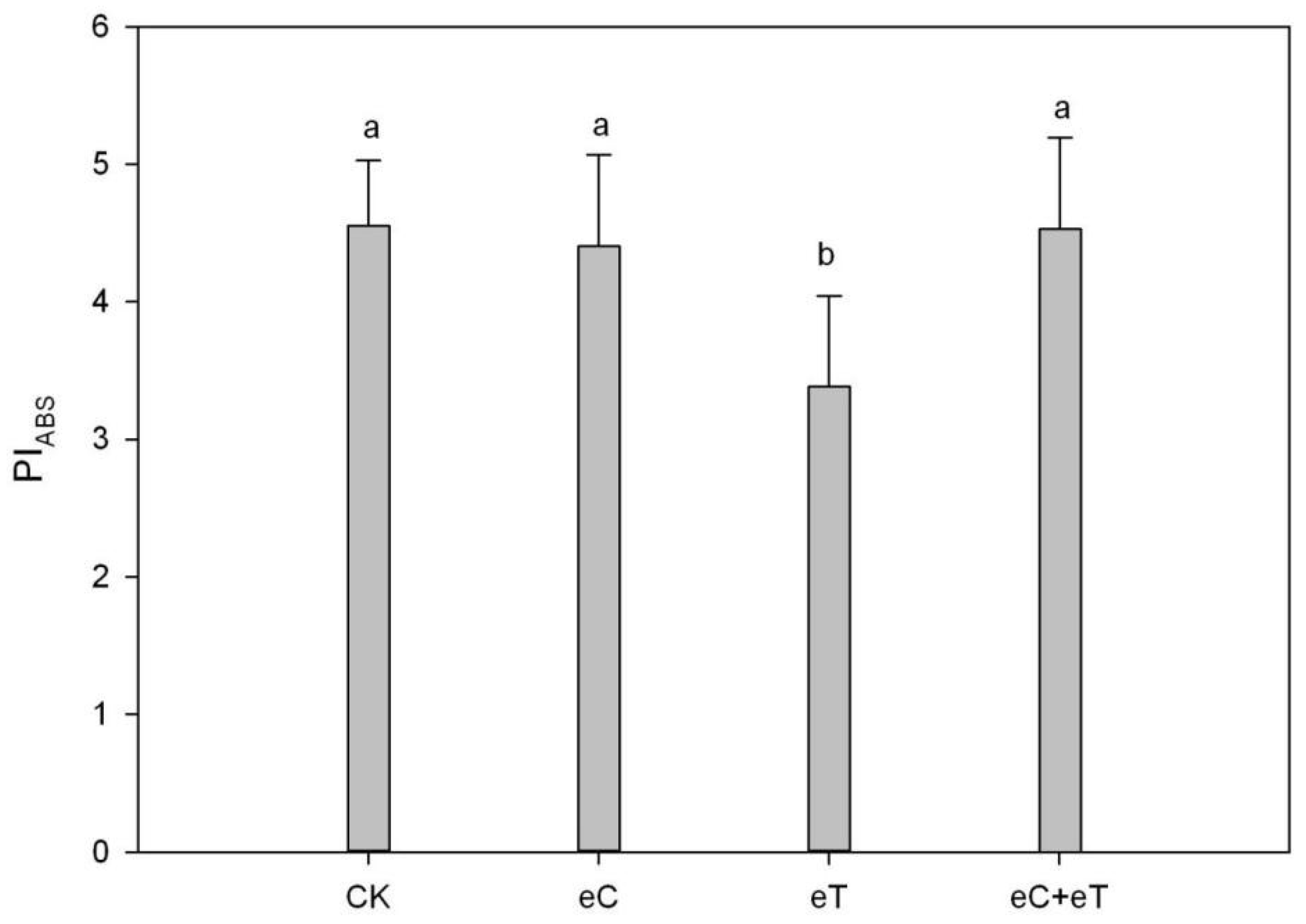
3.7. Effect of Elevated CO2 Concentration and Temperature on the PSII Performance Indicators in Populus simonii × P. nigra ‘1307’
4. Discussion
4.1. Effects of Increased Atmospheric CO2 and Elevated Temperatures on Leaf Growth and Chlorophyll Content of Populus simonii × P. nigra ‘1307’ Leaves
4.2. Effects of Increased Atmospheric CO2 and Elevated Temperatures on Photosynthetic Physiology of Populus simonii × P. nigra ‘1307’ Leaves
4.3. Enhanced Energy Conversion by Isncreased CO2 Concentration in the PSII Reaction Center
4.4. Enhanced Performance of PSII Donor and Receptor by Increased CO2 Concentration and Temperature
4.5. Role of the Increased CO2 Concentration on the Enhanced PSII Comprehensive Performance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects; Contribution of Working GroupII to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- De Kauwe, M.D.; Medlyn, B.E.; Zaehle, S.; Anthony, P.W.; Michael, C.D.; Thomas, H.; Atul, K.J.; Luo, Y.Q.; Parton, W.J.; Prentice, I.C.; et al. Forest water use and water use efficiency at elevated CO2: A model-data intercomparison at two contrasting temperate forest FACE sites. Glob. Change Biol. 2013, 19, 1759–1779. [Google Scholar] [CrossRef] [PubMed]
- Saurer, B.; Matthis, C.; Paolo, C.; McCarthy, C.C.; Heather, R.; Norby, R.J.; Francesca, C.M. Elevated CO2 increases tree-level intrinsic water use efficiency: Insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites. New Phytol. 2013, 197, 544–554. [Google Scholar]
- Keenan, T.; Holliger, D.; Bohrer, G.; Danilo, D.J.; William, M.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Peppe, D.J.; Royer, D.L.; Cariglino, B. Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications. New Phytol. 2011, 190, 724–739. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Xu, Y.B.; Song, S.Y.; Shen, Y.F.; Li, S.Q. Effect of doubled atmospheric CO2 and nitrogen application on photosynthetic rate and chlorophyll fluorescence character of winter wheat. Acta Bot. Boreal. Occid. Sin. 2011, 31, 144–151. [Google Scholar]
- Zhou, N.; Jing, L.Q.; Zhu, J.G.; Wang, Y.X.; Wang, Y.L.; Yang, L.X. Dynamic responses of leaf photosynthesis and chlorophyll fluorescence to elevated atmospheric CO2 concentration for two hybrid rice cultivars. J. Nucl. Agric. Sci. 2019, 33, 772–780. [Google Scholar]
- Su, X.Q.; Wang, M.Y.; Shu, S.; Sun, J.; Guo, S.R. Effects of exogenous Spd on the fast chlorophyll fluorescence induction dynamics in tomato seedlings under high temperature stress. Acta Hortic. Sin. 2013, 40, 2409–2418. [Google Scholar]
- Kanchan, J.; Virender, S.B. Interactive effect of temperature and water stress on physiological and biochemical processes in soybean. Physiol. Mol. Biol. Plants 2019, 25, 667–681. [Google Scholar]
- Wheeler, J.K.; Huggett, B.A.; Tofte, A.N. Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism. Plant Cell Environ. 2013, 36, 1938–1949. [Google Scholar] [CrossRef]
- Vincent, G. Leaf photosynthetic capacity and nitrogen content adjustment to canopy openness in tropical forest tree seedlings. J. Trop. Ecol. 2001, 17, 495–509. [Google Scholar] [CrossRef]
- Johnson, R.C.; Li, Y.Y. Water relations, forage production, and photosynthesis in tall fescue divergently selected for carbon isotope discrimination. Crop Sci. 1999, 39, 5692–5698. [Google Scholar] [CrossRef]
- Palmroth, S.; Bach, H.; Nordin, A.; Palmqvist, K. Nitrogen-addition effects on leaf traits and photosynthetic carbon gain of boreal forest understory shrubs. Oecologia 2014, 175, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Trifilò, P.; Nardini, A.; Lo Gullo, M.A.; Barbera, P.M.; Tadeja, S.; Fabio, R. Diurnal changes in embolism rate in nine dry forest trees: Relationships with species-specific xylem vulnerability, hydraulic strategy and wood traits. Tree Physiol. 2015, 35, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R.S.; Smith, E.S. Functional responses of plants to elevated atmospheric CO2: Do photosynthetic and productivity data from FACE experiments support early predictions? New Phytol. 2010, 162, 253–280. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Oleksyn, J.; Reich, P.B. Seedlings of five boreal tree species differ in acclimation of net photosynthesis to elevated CO2 and temperature. Tree Physiol. 1998, 18, 715–726. [Google Scholar] [CrossRef]
- Long, S.P. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations: Has its importance been underestimated? Plant Cell Environ. 1991, 14, 729–739. [Google Scholar] [CrossRef]
- Mott, K.A. Environment. Opinion: Stomatal responses to light and CO2 depend on the mesophyll. Plant Cell Environ. 2009, 32, 1479–1486. [Google Scholar] [CrossRef]
- Chavan, S.G.; Duursma, R.A.; Michael, T.; Ghannoum, O. Elevated CO2 alleviates the negative impact of heat stress on wheat physiology but not on grain yield. J. Exp. Bot. 2019, 70, 6447–6459. [Google Scholar] [CrossRef]
- Sun, P.; Mantri, N.; Lou, H.Q.; Hu, Y.; Sun, D.; Zhu, Y.Q. Effects of elevated CO2 and temperature on yield and fruit quality of strawberry (Fragaria ananassa Duch.) at two levels of nitrogen application. PLoS ONE 2012, 7, e41000. [Google Scholar] [CrossRef]
- Taylor, G.E.; Gunderson, G.A. Physiological site of ethylene effects on carbon dioxide assimilation in Glycine max L. Merr. Plant Physiol. 1988, 86, 85–92. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, S.; Sun, L.; Song, F.B.; Liu, F.L.; Li, X.N. Cold tolerance of photosynthetic electron transport system is enhanced in wheat plants grown under elevated CO2. Front. Plant Sci. 2018, 9, 933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Xu, N.; Li, X.; Jin, W.W.; Tian, Q.; Sun, G.Y.; Gu, S.Y. Overexpression of 2-Cys Prx increased salt tolerance of photosystem Ⅱ(PSⅡ) in tobacco. Inter. J. Agric. Biol. 2017, 19, 735–745. [Google Scholar] [CrossRef]
- Li, F.S.; Kang, S.Z.; Zhang, F.C. Effects of CO2 enrichment, nitrogen and water on photosynthesi, evapotranspiration and water use efficiency of spring wheat. Chin. J. Appl. Ecol. 2003, 14, 387–393. [Google Scholar]
- Varga, B.; Vida, G.; Varga-László, E.; Hoffmann, B.; Veisz, O. Combined Effect of Drought Stress and Elevated Atmospheric CO2 Concentration on the Yield Parameters and Water Use Properties of Winter Wheat (Triticum aestivum L.) Genotypes. J. Agron. Crop Sci. 2017, 203, 192–205. [Google Scholar] [CrossRef]
- Xu, F.; Jiang, M.; Meng, F. Short-term effect of elevated CO2 concentration (0.5%) on mitochondria in diploid and tetraploid black locust (Robinia pseudoacacia L.). Ecol. Evol. 2017, 7, 4651–4660. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Jian, Y.; Jia, B.; Zhou, G. Elevated-CO2 response of stomatal and its dependence on environmental factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef]
- Wang, A.; Lam, S.K.; Hao, X.Y.; Li, F.Y.; Zong, Y.; Wang, H.; Li, P. Elevated CO2 reduces the adverse effects of drought stress on a high-yielding soybean (Glycine max (L.) Merr.) cultivar by increasing water use efficiency. Plant Physiol. Biochem. 2018, 132, 660–665. [Google Scholar] [CrossRef]
- Zhang, H.H.; Xu, N.; Teng, Z.Y.; Wang, J.R.; Ma, S.L.; Wu, X.Y.; Li, X.; Sun, G.Y. 2-Cys Prx plays a critical role in scavenging H2O2 and protecting photosynthetic function in leaves of tobacco seedlings under drought stress. J. Plant Inter. 2019, 14, 119–128. [Google Scholar] [CrossRef]
- Woodward, F.I.; Kelly, C.K. The influence of CO2 concentration on stomatal density. New Phytol. 2006, 131, 311–327. [Google Scholar] [CrossRef]
- Pazzaglia, P.T.; Weiner, J.; Liu, F. Effects of CO2 elevation and irrigation regimes on leaf gas exchange plant water relations, and water use efficiency of two tomato cultivars. Agric. Water Manag. 2019, 169, 26–33. [Google Scholar] [CrossRef]
- Li, P.M.; Gao, H.Y.; Strasser, R.J. Application of the fast chlorophyll fluorescence in duction dynamics analysis in photosynthesis study. J. Plant Physiol. Mol. Biol. 2005, 31, 559–566. [Google Scholar]
- Fan, P.P.; Feng, F.; Liu, C.; Sun, W.J.; Yu, L.F.; Ke, H.N.; Chen, S.T.; Hu, Z.H. Effect of different levels of elevated CO2 concentration on leaf chlorophyll fluorescence characteristics of Japonica rice. Chin. J. Appl. Ecol. 2019, 30, 3735–3744. [Google Scholar]
- Jin, L.Q.; Che, X.K.; Zhang, Z.S.; Gao, H.Y. The relationship between the changes in Wk and different damage degree of PSII donor side and acceptor side under high temperature with high light in cucumber. Plant Physiol. J. 2015, 51, 969–976. [Google Scholar]
- Du, G.D.; Lü, D.G.; Zhao, L.; Wang, S.S.; Cai, Q. Effects of high temperature on leaf photosynthetic characteristics and photosystem II photo-chemical activity of kernel-used apricot. Chin. J. Appl. Ecol. 2011, 22, 701–706. [Google Scholar]
- Chen, F.; Tian, J.C.; Meng, Q.W.; Zhao, S.J. Effects of short-term high temperature stress on flag leaf photosystemII functions of high-yielding wheat at late grain-filling stage. Chin. J. Appl. Ecol. 2006, 17, 1854–1858. [Google Scholar]
- Van Heerden, P.D.R.; Strasser, R.J.; Krüger, G.H.J. Reduction of dark chilling stress in N2-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics. Physiol. Plant 2004, 121, 239–249. [Google Scholar]
- Cosentino, S.L.; Sanzone, E.; Testa, G.; Testa, G.; Patanè, C.; Anastasi, U.; Scordia, D. Does post-anthesis heat stress affect plant phenology, physiology, grain yield and protein content of durum wheat in a semi-arid mediterranean environment? J. Agron. Crop Sci. 2019, 205, 309–323. [Google Scholar] [CrossRef]
- Zhang, L.T.; Liu, J.G. Effects of heat stress on photosynthetic electron transport in a marine cyanobacterium Arthrospira sp. J. Appl. Phycol. 2016, 28, 757–763. [Google Scholar]
- Semenov, M.A.; Halford, N.G. Identifying target traits and molecular mechanisms for wheat breeding under a changing climate. J. Exp. Bot. 2009, 60, 2791–2804. [Google Scholar]
- Prasad, P.V.V.; Djanaguiraman, M. Response of floret fertility and individual grain weight of wheat to high temperature stress: Sensitive stages and thresholds for temperature and duration. Funct. Plant Biol. 2014, 41, 1261–1269. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CK | eC | eT | eC + eT | |
|---|---|---|---|---|
| Leaf dry weight (g) | 11.48 ± 1.26 a | 11.96 ± 1.34 a | 11.12 ± 2.32 a | 13.50 ± 2.33 a |
| Stem dry weight (g) | 45.23 ± 4.02 a | 33.56 ± 2.34 b | 34.03 ± 2.34 a | 33.12 ± 3.78 a |
| Root dry weight (g) | 29.76 ± 5.23 c | 53.18 ± 9.56 a | 34.25 ± 8.15 bc | 47.22 ± 8.23 a |
| Plant Height (cm) | 171.32 ± 3.13 a | 163.44 ± 5.28 ab | 158.89 ± 5.97 bc | 153962 ± 3.13 c |
| Ground diameter (cm) | 0.84 ± 0.02 a | 0.85 ± 0.32 a | 0.85 ± 0.31 a | 0.84 ± 0.13 a |
| RWC (%) | 95.28 ± 2.54 c | 97.27 ± 5.23 ab | 98.45 ± 4.21 a | 96.48 ± 0.06 a |
| LMA (gm−2) | 75.34 ± 6.23 c | 113.23 ± 6.51 a | 108.25 ± 5.12 a | 92.33 ± 4.45 b |
| CK | eC | eC + eT | eT | |
|---|---|---|---|---|
| Pmax (μmol m−2s−1) | 15.48 ± 1.06 b | 17.22 ± 1.23 b | 19.22 ± 1.82 a | 15.01 ± 0.86 b |
| Pn (μmol m−2s−1) | 18.23 ± 0.46 b | 21.13 ± 0.34 a | 22.45 ± 1.26 a | 16.23 ± 0.83 b |
| Gs (mol m−2s−1) | 1.44 ± 0.36 a | 0.84 ± 0.24 c | 1.01 ± 0.32 bc | 1.32 ± 0.54 ab |
| Tr (mmol m−2s−1) | 14.45 ± 0.88 b | 12.24 ± 1.34 a | 13.82 ± 0.12 a | 14.13 ± 0.24 b |
| Ci (μmol mol−1) | 385.28 ± 15.36 a | 552.13 ± 23.52 a | 549.65 ± 18.52 b | 398.45 ± 12.21 a |
| WUE (μmol mmol−1) | 0.97 ± 0.03 b | 1.37 ± 0.23 a | 1.67 ± 0.23 a | 0.97 ± 0.03 b |
| PNUE (μmol mol−1s−1) | 113.27 ± 8.23 b | 108.24 ± 6.24 b | 132.56 ± 22.35 a | 103.56 ± 2.22 b |
| Wk | Vj | Mo | ΨPo | Ψo | ΨEo | ΨDo | |
|---|---|---|---|---|---|---|---|
| CK | 0.48 ± 0.03 a | 0..44 ± 0.03 a | 0.68 ± 0.02 ab | 0.86 ± 0.04 ab | 0.67 ± 0.02 a | 0.56 ± 0.04 a | 0.19 ± 0.04 b |
| eC | 0.42 ± 0.04 b | 0.47 ± 0.03 ab | 0.63 ± 0.03 ab | 0.87 ± 0.05 a | 0.64 ± 0.01 ab | 0.54 ± 0.03 a | 0.19 ± 0.02 b |
| eT | 0.46 ± 0.02 ab | 0.49 ± 0.04 a | 0.73 ± 0.03 a | 0.85 ± 0.03 b | 0.62 ± 0.02 b | 0.52 ± 0.02 b | 0.20 ± 0.02 a |
| eC + eT | 0.42 ± 0.05 b | 0.16 ± 0.02 a | 0.61 ± 0.05 b | 0.86 ± 0.06 ab | 0.65 ± 0.03 a | 0.55 ± 0.03 a | 0.20 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, N.; Ding, J.; Zhang, T.; Dong, J.; Wang, Y.; Yang, X. Effects of Elevated CO2 Concentration and Temperature on the Growth and Photosynthetic Characteristics of Populus simonii × P. nigra ‘1307’ Leaves. Forests 2023, 14, 2156. https://doi.org/10.3390/f14112156
Xu N, Ding J, Zhang T, Dong J, Wang Y, Yang X. Effects of Elevated CO2 Concentration and Temperature on the Growth and Photosynthetic Characteristics of Populus simonii × P. nigra ‘1307’ Leaves. Forests. 2023; 14(11):2156. https://doi.org/10.3390/f14112156
Chicago/Turabian StyleXu, Nan, Junnan Ding, Tianyi Zhang, Juexian Dong, Yuan Wang, and Xuechen Yang. 2023. "Effects of Elevated CO2 Concentration and Temperature on the Growth and Photosynthetic Characteristics of Populus simonii × P. nigra ‘1307’ Leaves" Forests 14, no. 11: 2156. https://doi.org/10.3390/f14112156
APA StyleXu, N., Ding, J., Zhang, T., Dong, J., Wang, Y., & Yang, X. (2023). Effects of Elevated CO2 Concentration and Temperature on the Growth and Photosynthetic Characteristics of Populus simonii × P. nigra ‘1307’ Leaves. Forests, 14(11), 2156. https://doi.org/10.3390/f14112156