
Lobaria pulmonaria (L.) Hoffm.: The Multifaceted Suitability of the Lung Lichen to Monitor Forest Ecosystems

 , ,
, ,  , ,
, ,  ,
,  , , and
, , and 
Abstract
1. Introduction
- 1.
- It has a wide distribution, being present in boreal, temperate, mountainous, and oceanic regions of the northern hemisphere and in tropical forests of Eastern and Southern Africa [8].
- 2.
- Its long life cycle (the time to reach sexual fertility has been estimated up to 35 years) makes it an excellent indicator of undisturbed forest ecosystems, as well as forest areas with high ecological continuity (e.g., [9]).
- 3.
- It is a tripartite lichen, with the green alga Symbiochloris reticulata (Tschermak-Woess) Skaloud, Friedl, A.Beck and Dal Grande as its main photobiont and N-fixing cyanobacteria of the genus Nostoc, which are confined in structures called cephalodia inside the thallus [10]. This allows L. pulmonaria to actively contribute to the N balance of forest ecosystems [11,12].
- 4.
- 5.
- Compared to the usual small size of lichens, this species has a large thallus, often exceeding 20–30 cm in diameter, rather showy and easy to distinguish even by non-specialist personnel. For this reason, it can be used in large-scale monitoring networks involving numerous forest operators [16].
2. Materials and Methods
3. Results
3.1. Contextual Topics
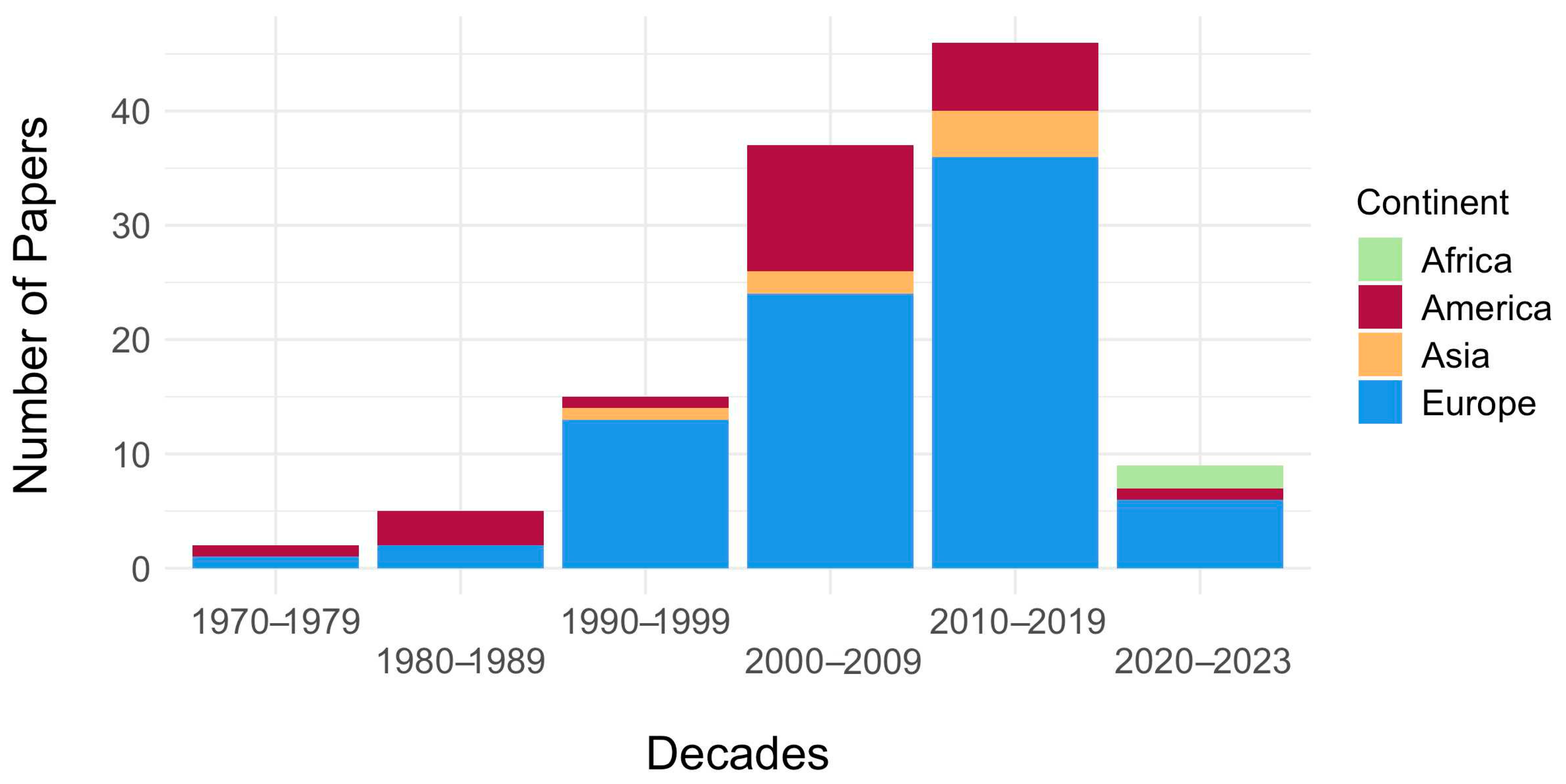
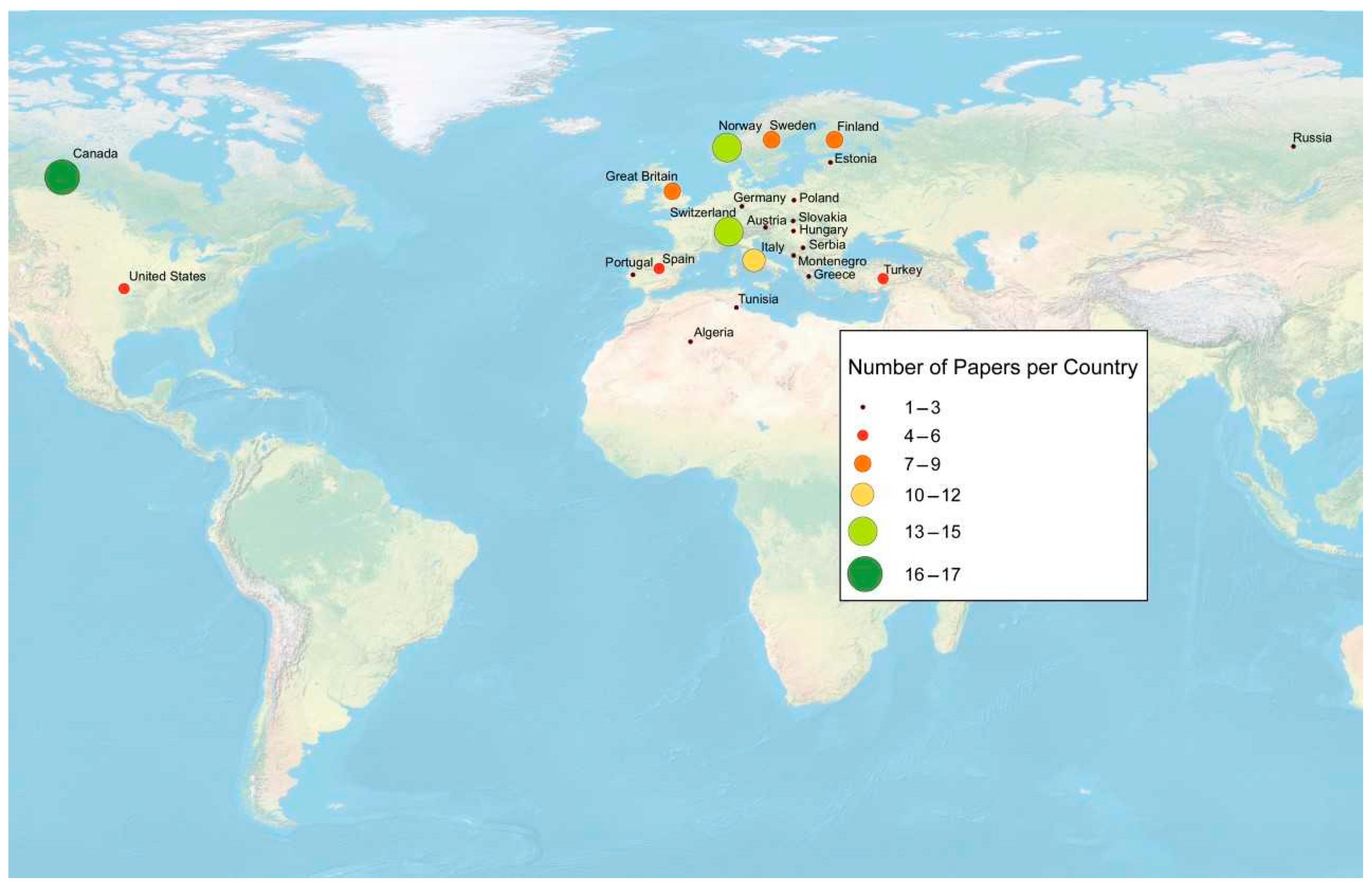
3.2. Temporal and Spatial Distribution
4. Discussion
4.1. Ecosystem Functioning and Ecosystem Services
4.1.1. Carbon and Nitrogen Cycling
4.1.2. Growth, Dispersal, and Grazing
4.2. Lobaria pulmonaria in Air Pollution Monitoring
4.2.1. Monitoring Lobaria pulmonaria
4.2.2. Monitoring with Lobaria pulmonaria
4.2.3. Monitoring the Effects of Acid Rain
4.2.4. Monitoring around Pollution Sources
4.2.5. Manipulative Experiments in Relation to Specific Pollutants
4.2.6. Monitoring Radionuclides
4.2.7. Major and Trace Elements and PAHs in Remote Areas
4.3. Forest Management
4.3.1. Monitoring Lobaria pulmonaria
4.3.2. Influence of Forest Management
4.3.3. Genetic Diversity
4.3.4. Forest Edges
4.3.5. Tree-Level Factors
4.3.6. Forest Type
4.3.7. Dispersal Capacity
4.3.8. Monitoring with Lobaria pulmonaria
5. Conclusions
- In terms of ecosystem functioning and services, L. pulmonaria contributes to C cycle and moreover N2 fixation (thanks to the presence of cephalodia). Lobaria pulmonaria is important for the forest’s food network, especially for snails and slugs, and it supports the diversity of some organisms, e.g., offering suitable microhabitats for several small animals. However, the interaction between L. pulmonaria and lichen-eating animals still requires investigations.
- Lobaria pulmonaria is considered to be extremely sensitive to pollution. Its application in air pollution monitoring refers to past acid rain and SO2 depositions, heavy metals, radionuclides, and PAHs in forest habitats. Hence, air pollution (especially in the past) and habitat fragmentation (nowadays) appear as the main anthropogenic factors influencing Lobarion vegetation. On the other hand, its use to monitor air pollution is in fact limited by its sensitivity and rarity. Actually, there is particular interest in understanding whether current air quality still limits the possibility of recolonisation in forest sites where L. pulmonaria disappeared, as well as the interaction between pollution, microclimatic parameters, and forest structure and dynamics in a conservation perspective. In this sense, recent research also highlighted the possibility of translocation practices for conservation purposes in relation to potential anthropogenic impact on L. pulmonaria in threatened habitats.
- We discussed the impact of forest management on L. pulmonaria, a sensitive species to the variation of microclimatic conditions. Forest management practices, such as logging and fragmentation, can negatively affect L. pulmonaria populations, leading to a loss of biomass, changes in population composition, and reduced vitality. Retaining unlogged forest aggregates is essential to preserve vital populations of L. pulmonaria. Genetic diversity in L. pulmonaria can be influenced by forest disturbances, potentially leading to clonal vegetative reproduction. Genetic dissimilarity is influenced also by tree species and trunk diameter, and forest management can impact population structure also at a genetic level. We also discussed the impact of forest edges on L. pulmonaria, with varying effects depending on edge type and local conditions. The species tends to have a higher growth rate near ‘soft’ forest edges but may not show the same enhancement near ‘hard’ edges. Factors at tree and stand levels, such as tree age, canopy openness, and habitat continuity, play a crucial role in L. pulmonaria colonisation and survival. The species is associated with old-growth forests with long ecological continuity, and the ecological requirements vary during its life cycle. Forest type, light availability, and climatic conditions also influence the presence and growth of L. pulmonaria. The species is more likely to colonise older trees with large circumferences, but it can adapt to different conditions, especially with optimal climatic conditions. Finally, L. pulmonaria is used as an indicator species for assessing forest characteristics and the presence of endangered species. Its abundance is often correlated with forest continuity and the presence of specific (and often desirable) ecological conditions. Hence, the complex relationship between forest management, microclimatic conditions, genetic diversity, and the ecological role of L. pulmonaria in forest ecosystems further highlight the need for sustainable and modern forest management practices.
- The conservation of the species per se, and as an umbrella species for epiphytic communities of conservation concern. Future efforts should focus on regular, standardised, and harmonised monitoring of the conservation status of L. pulmonaria at continental and global scales to ensure its adequate protection in the context of climate change scenarios;
- as part of monitoring the effects of forest management plans in order to provide biological data to support policymakers’ decisions and to guide the use of forest resources to be both economically and environmentally sustainable.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Nascimbene, J.; Benesperi, R.; Giordani, P.; Grube, M.; Marini, L.; Vallese, C.; Mayrhofer, H. Could Hair-Lichens of High-Elevation Forests Help Detect the Impact of Global Change in the Alps? Diversity 2019, 11, 45. [Google Scholar] [CrossRef]
- Jovan, S.E.; McCune, B. Air-quality bioindication in the greater Central Valley of California, with epiphytic macrolichen communities. Ecol. Appl. 2005, 15, 1712–1726. [Google Scholar] [CrossRef]
- Jovan, S.; Riddell, J.; Padgett, P.E.; Nash, T.H. Eutrophic lichens respond to multiple forms of N: Implications for critical levels and critical loads research. Ecol. Appl. 2012, 22, 1910–1922. [Google Scholar] [CrossRef] [PubMed]
- Geiser, L.H.; Neitlich, P.N. Air pollution and climate gradients in western Oregon and Washington indicated by epiphytic macrolichens. Environ. Pollut. 2007, 145, 203–218. [Google Scholar] [CrossRef]
- Will-Wolf, S.; Geiser, L.H.; Neitlich, P.; Reis, A.H. Forest lichen communities and environment—How consistent are relationships across scales? J. Veg. Sci. 2006, 17, 171–184. [Google Scholar] [CrossRef]
- Mayer, A.L.; Vihermaa, L.; Nieminen, N.; Luomi, A.; Posch, M. Epiphytic macrolichen community correlates with modeled air pollutants and forest conditions. Ecol. Indic. 2009, 9, 992–1000. [Google Scholar] [CrossRef]
- Grimm, M.; Grube, M.; Schiefelbein, U.; Zühlke, D.; Bernhardt, J.; Riedel, K. The Lichens’ Microbiota, Still a Mystery? Front. Microbiol. 2021, 12, 623839. [Google Scholar] [CrossRef]
- Jüriado, I.; Liira, J. Distribution and habitat ecology of the threatened forest lichen Lobaria pulmonaria in Estonia. Folia Crypt. Eston. 2009, 46, 55–65. [Google Scholar]
- Jordan, W.P. The internal cephalodia of the genus Lobaria. Bryol. 1970, 73, 669–681. [Google Scholar] [CrossRef]
- Millbank, J.W.; Kershaw, K.A. Nitrogen metabolism in lichens. New Phytol. 1970, 69, 595–597. [Google Scholar] [CrossRef]
- Eymann, C.; Lassek, C.; Wegner, U.; Bernhardt, J.; Fritsch, O.A.; Fuchs, S.; Otto, A.; Albrecht, D.; Schiefelbein, U.; Cernava, T.; et al. Symbiotic interplay of fungi, algae, and bacteria within the lung lichen Lobaria pulmonaria L. Hoffm. as assessed by State-of-the-Art metaproteomics. J. Proteome Res. 2017, 16, 2160–2173. [Google Scholar] [CrossRef] [PubMed]
- Rose, F. Phytogeographical and ecological aspects of Lobarion communities in Europe. Bot. J. Linnean. Soc. 1988, 96, 69–79. [Google Scholar] [CrossRef]
- Gauslaa, Y. Lobarion, an epiphytic community of ancient forests, threatened by acid rain. Lichenologist 1995, 27, 59–76. [Google Scholar] [CrossRef]
- Richardson, D.H.S.; Cameron, R.P. Cyanolichens: Their response to pollution and possible management strategies for their conservation in northeastern North America. Northeast. Nat. 2004, 11, 1–22. [Google Scholar] [CrossRef]
- Scheidegger, C.; Frey, B.; Walser, J.C. Reintroduction and augmentation of populations of the endangered Lobaria pulmonaria: Methods and concepts. In Proceedings of the Darwin International Workshop Lobarion Lichens as Indicators of Primaeval Forests of Eastern Carpathians, Kostrino, Ukraine, 25–30 May 1998; Kondratyuk, S., Coppins, B., Eds.; pp. 33–52. [Google Scholar]
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Nordén, B.; Paltto, H.; Götmark, F.; Wallin, K. Indicators of biodiversity, what do they indicate?—Lessons for conservation of cryptogams in oak-rich forest. Biol. Conserv. 2007, 135, 369–379. [Google Scholar] [CrossRef]
- Nascimbene, J.; Brunialti, G.; Ravera, S.; Frati, L.; Caniglia, G. Testing Lobaria pulmonaria (L.) Hoffm as an indicator of lichen conservation importance of Italian forests. Ecol. Indic. 2010, 10, 353–360. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Arup, U.; Baranowski, R.; Ekman, S. Tree-dependent lichens and beetles as indicators in conservation forests. Conserv. Biol. 1995, 9, 1208–1215. [Google Scholar] [CrossRef]
- Scheidegger, C.; Werth, S. Conservation strategies for lichens: Insights from population biology. Fungal Biol. Rev. 2009, 23, 55–66. [Google Scholar] [CrossRef]
- Nascimbene, J.; Thor, G.; Nimis, P.L. Effects of forest management on epiphytic lichens in temperate deciduous forests of Europe—A review. For. Ecol. Manag. 2013, 298, 27–38. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Ravera, S. Structural variables drive the distribution of the sensitive lichen Lobaria pulmonaria in Mediterranean old-growth forests. Ecol. Indic. 2015, 53, 37–42. [Google Scholar] [CrossRef]
- Paoli, L.; Benesperi, R.; Fačkovcová, Z.; Nascimbene, J.; Ravera, S.; Marchetti, M.; Anselmi, B.; Landi, M.; Landi, S.; Bianchi, E.; et al. Impact of forest management on threatened epiphytic macrolichens: Evidence from a Mediterranean mixed oak forest (Italy). IForest—Biogeosci. For. 2019, 12, 383. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Nimis, P.L. Influence of forest management on epiphytic lichens in a temperate beech forest of northern Italy. For. Ecol. Manag. 2007, 247, 43–47. [Google Scholar] [CrossRef]
- Martellos, S.; Pittao, E.; Cesaroni, D.; Mereu, A.; Petruzzella, D.; Pinzari, M.; Sbordoni, V.; Tallone, G.; Attorre, F. Volunteers recruitment, retention, and performance during the CSMON-LIFE (Citizen Science MONitoring) Project and 3 years of follow-up. Sustainability 2021, 13, 11110. [Google Scholar] [CrossRef]
- Botting, R.S.; Campbell, J.; Fredeen, A.L. Contrasting arboreal and terrestrial macrolichen and bryophyte communities in old-growth sub-boreal spruce forests of central British Columbia. Bryologist 2008, 111, 607–619. [Google Scholar] [CrossRef]
- Campbell, J.; Fredeen, A.L. Lobaria pulmonaria abundance as an indicator of macrolichen diversity in interior cedar-hemlock forests of east-central British Columbia. Canad. J. Bot. 2004, 82, 970–982. [Google Scholar] [CrossRef]
- Campbell, J.; Fredeen, A.L. Contrasting the abundance, nitrogen, and carbon of epiphytic macrolichen species between host trees and soil types in a sub-boreal forest. Canad. J. Bot. 2007, 85, 31–42. [Google Scholar] [CrossRef]
- Campbell, J.; Fredeen, A.L.; Prescott, C.E. Decomposition and nutrient release from four epiphytic lichen litters in sub-boreal spruce forests. Can. J. For. Res. 2010, 40, 1473–1484. [Google Scholar] [CrossRef]
- Kobylinski, A.; Fredeen, A.L. Vertical distribution and nitrogen content of epiphytic macrolichen functional groups in sub-boreal forests of central British Columbia. For. Ecol. Manag. 2014, 329, 118–128. [Google Scholar] [CrossRef]
- Kobylinski, A.; Fredeen, A.L. Importance of arboreal cyanolichen abundance to nitrogen cycling in sub-boreal spruce and fir forests of Central British Columbia, Canada. Forests 2015, 6, 2588–2607. [Google Scholar] [CrossRef]
- Blacklock, N.E.; Goldsmith, F.B.; Patriquin, D.G. Nitrogen fixation (acetylene reduction) by lungwort lichen (Lobaria pulmonaria) on red maple (Acer rubrum). Proc. Nova Scotian Inst. Sci. 1980, 30, 89–107. [Google Scholar]
- Gauslaa, Y.; Goward, T.; Pypker, T. Canopy settings shape elemental composition of the epiphytic lichen Lobaria pulmonaria in unmanaged conifer forests. Ecol. Indic. 2020, 113, 106294. [Google Scholar] [CrossRef]
- Millbank, J.W. The assessment of nitrogen fixation and throughput by lichens. New Phytol. 1982, 92, 229–234. [Google Scholar] [CrossRef]
- Pentecost, A. Estimates of abundance and biomass of cephalodia and their relationship to nitrogen deposition in some British populations of Lobaria pulmonaria (L.) Hoffm. Lichenologist 2021, 53, 335–339. [Google Scholar] [CrossRef]
- Denison, R.; Caldwell, B.; Bormann, B.; Lindell, E.; Swanberg, C.; Anderson, S. Contents of carbon based defence compounds in the old forest lichen Lobaria pulmonaria vary along environmental gradients. Water Air Soil Pollut. 1977, 8, 21–34. [Google Scholar] [CrossRef]
- Sigal, L.L.; Johnston, W. Effects of acidic rain and ozone on nitrogen fixation and photosynthesis in the lichen Lobaria pulmonaria (L.) Hoffm. Environ. Exp. Bot. 1986, 26, 59–64. [Google Scholar] [CrossRef]
- Gilbert, O.L. Field evidence for an acid rain effect on lichens. Environ. Pollut. Ser. A Ecol. Biol. 1986, 40, 227–231. [Google Scholar] [CrossRef]
- Rose, F. Lichenological indicators of age and environmental continuity in woodlands. In Lichenology: Progress and Problems; Brown, D.H., Hawksworth, D.L., Bailey, R.H., Eds.; Academic Press: London, UK; New York, NY, USA, 1976; pp. 279–307. [Google Scholar]
- Paoli, L.; Fačkovcová, Z.; Lackovičová, A.; Guttová, A. Air pollution in Slovakia (Central Europe): A story told by lichens (1960–2020). Biologia 2021, 76, 3235–3255. [Google Scholar] [CrossRef]
- Gauslaa, Y. Lobaria pulmonaria, an indicator of species-rich forests of long ecological continuity. Blyttia 1994, 52, 119–128. [Google Scholar]
- Kalwij, J.M.; Wagner, H.H.; Scheidegger, C. Effects of stand-level disturbances on the spatial distribution of a lichen indicator. Ecol. Appl. 2005, 15, 2015–2024. [Google Scholar] [CrossRef]
- Carlsson, R.; Nilsson, K. Status of the red-listed lichen Lobaria pulmonaria on the Åland Islands, SW Finland. Ann. Bot. Fenn. 2009, 46, 549–554. [Google Scholar] [CrossRef]
- Dymytrova, L.; Brändli, U.-B.; Ginzler, C.; Scheidegger, C. Forest history and epiphytic lichens: Testing indicators for assessing forest autochthony in Switzerland. Ecol. Indic. 2018, 84, 847–857. [Google Scholar] [CrossRef]
- Dymytrova, L.; Brändli, U.-B.; Stofer, S.; Scheidegger, C. Recognizing autochthonous mountain forests in Switzerland by means of epiphytic lichens. Schweiz. Z. Forstwes. 2019, 170, 258–265. [Google Scholar] [CrossRef]
- Kuusinen, M. Cyanobacterial macrolichens on Populus tremula as indicators of forest continuity in Finland. Biol. Conserv. 1996, 75, 43–49. [Google Scholar] [CrossRef]
- Ojala, E.; Mönkkönen, M.; Inkeröinen, J. Epiphytic bryophytes on European aspen Populus tremula in old-growth forests in northeastern Finland and in adjacent sites in Russia. Can. J. Bot. 2000, 78, 529–536. [Google Scholar] [CrossRef]
- Sundberg, B.; Palmqvist, K.; Esseen, P.-A.; Renhorn, K.-E. Growth and vitality of epiphytic lichens: II. Modelling of carbon gain using field and laboratory data. Oecologia 1997, 109, 10–18. [Google Scholar] [CrossRef]
- Paoli, L.; Guttová, A.; Sorbo, S.; Lackovičová, A.; Ravera, S.; Landi, S.; Landi, M.; Basile, A.; Sanità di Toppi, L.; Vannini, A.; et al. Does air pollution influence the success of species translocation? Trace elements, ultrastructure and photosynthetic performances in transplants of a threatened forest macrolichen. Ecol. Indic. 2020, 117, 106666. [Google Scholar] [CrossRef]
- Riga-Karandinos, A.N.; Karandinos, M.G. Assessment of air pollution from a lignite power plant in the plain of Megalopolis (Greece) using as biomonitors three species of lichens; impacts on some biochemical parameters of lichens. Sci. Total Environ. 1998, 215, 167–183. [Google Scholar] [CrossRef]
- Çiçek, A.; Koparal, A.S.; Aslan, A.; Yazici, K. Accumulation of heavy metals from motor vehicles in transplanted lichens in an urban area. Commun. Soil Sci. Plant Anal. 2008, 39, 168–176. [Google Scholar] [CrossRef]
- Yemets, O.A.; Solhaug, K.A.; Gauslaa, Y. Spatial dispersal of airborne pollutants and their effects on growth and viability of lichen transplants along a rural highway in Norway. Lichenologist 2014, 46, 809–823. [Google Scholar] [CrossRef]
- Kouadria, N.; Alioua Berrebbah, A.; Belhoucine, F.; Bouredja, N.; Aitkaci, M. Cellular impact of metal trace elements on the lichen Lobaria pulmonaria (L.) Hoffm. (1796), a bioindicator of atmospheric pollution and identification of its antioxidant response. Appl. Ecol. Environ. Res. 2021, 19, 4721–4738. [Google Scholar] [CrossRef]
- Blasco, M.; Domeño, C.; Lópeza, P.; Nerín, C. Behaviour of different lichen species as biomonitors of air pollution by PAHs in natural ecosystems. J. Environ. Monit. 2011, 13, 2588–2596. [Google Scholar] [CrossRef]
- Bolshunova, T.; Rikhvanov, L.; Mezhibor, A.; Zhornyak, L.; Baranovskaya, N.; Eremina, E. Biogeochemical characteristics of epiphytic lichen Lobaria pulmonaria of the Barguzin nature reserve (The republic of Buryatia, Russia). J. Environ. Eng. Landsc. 2018, 26, 120–127. [Google Scholar] [CrossRef]
- Chahloul, N.; Khadhri, A.; Vannini, A.; Mendili, M.; Raies, A.; Loppi, S. Bioaccumulation of potentially toxic elements in some lichen species from two remote sites of Tunisia. Biologia 2022, 77, 2469–2473. [Google Scholar] [CrossRef]
- Muir, P.S.; Shirazi, A.M. Effects of formaldehyde-enriched mists on Pseudotsuga menziesii (Mirbel) Franco and Lobaria pulmonaria (L.) Hoffm. Environ. Pollut. 1996, 94, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Branquinho, C.; Brown, D.H.; Máguas, C.; Catarino, F. Lead (Pb) uptake and its effects on membrane integrity and chlorophyll fluorescence in different lichen species. Environ. Exp. Bot. 1997, 37, 95–105. [Google Scholar] [CrossRef]
- Hauck, M.; Spribille, T. The Mn/Ca and Mn/Mg ratios in bark as possible causes for the occurrence of Lobarion lichens on conifers in the dripzone of Populus in western North America. Lichenologist 2002, 34, 527–532. [Google Scholar] [CrossRef]
- Paul, A.; Hauck, M. Effects of manganese on chlorophyll fluorescence in epiphytic cyano- and chlorolichens. Flora Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 451–460. [Google Scholar] [CrossRef]
- Hauck, M.; Paul, A.; Spribille, T. Uptake and toxicity of manganese in epiphytic cyanolichens. Environ. Exp. Bot. 2006, 56, 216–224. [Google Scholar] [CrossRef]
- Yemets, O.; Gauslaa, Y.; Solhaug, K.A. Monitoring with lichens—Conductivity methods assess salt and heavy metal damage more efficiently than chlorophyll fluorescence. Ecol. Indic. 2015, 55, 59–64. [Google Scholar] [CrossRef]
- Jüriado, I.; Liira, J. Threatened forest lichen Lobaria pulmonaria—Its past, present and future in Estonia|[Harilik kopsusamblik (Lobaria pulmonaria)—Ohustatud metsasamblik eestis minevikus, tänapäeval ja tulevikus]. For. Stud. 2010, 53, 15–24. [Google Scholar] [CrossRef]
- Nascimbene, J.; Caniglia, G.; Nicli, M.; Dalle Vedove, M. Populations of Lobaria pulmonaria (L.) Hoffm. in the Cansiglio Regional Forest (Veneto, Pre-Alps, north-east Italy): Distribution, diversity and conservation issues. Plant Biosyst. 2006, 140, 34–42. [Google Scholar] [CrossRef]
- Gu, W.-D.; Kuusinen, M.; Konttinen, T.; Hanski, I. Spatial pattern in the occurrence of the lichen Lobaria pulmonaria in managed and virgin boreal forests. Ecography 2001, 24, 139–150. [Google Scholar] [CrossRef]
- Snäll, T.; Pennanen, J.; Kivistö, L.; Hanski, I. Modelling epiphyte metapopulation dynamics in adynamic forest landscape. Oikos 2005, 109, 209–222. [Google Scholar] [CrossRef]
- Bianchi, E.; Benesperi, R.; Brunialti, G.; Di Nuzzo, L.; Fačkovcová, Z.; Frati, L.; Giordani, P.; Nascimbene, J.; Ravera, S.; Vallese, C.; et al. Vitality and growth of the threatened lichen Lobaria pulmonaria (L.) Hoffm. in response to logging and implications for its conservation in mediterranean oak forests. Forests 2020, 11, 995. [Google Scholar] [CrossRef]
- Di Nuzzo, L.; Giordani, P.; Benesperi, R.; Brunialti, G.; Fačkovcová, Z.; Frati, L.; Nascimbene, J.; Ravera, S.; Vallese, C.; Paoli, L.; et al. Microclimatic Alteration after Logging Affects the Growth of the Endangered Lichen Lobaria pulmonaria. Plants 2022, 11, 295. [Google Scholar] [CrossRef] [PubMed]
- Fačkovcová, Z.; Guttová, A.; Benesperi, R.; Loppi, S.; Bellini, E.; Sanità di Toppi, L.; Paoli, L. Retaining unlogged patches in mediterranean oak forests may preserve threatened forest macrolichens. Iforest 2019, 12, 187–192. [Google Scholar] [CrossRef]
- Hazell, P.; Gustafsson, L. Retention of trees at final harvest—Evaluation of a conservation technique using epiphytic bryophyte and lichen transplants. Biol. Conserv. 1999, 90, 133–142. [Google Scholar] [CrossRef]
- Gustafsson, L.; Fedrowitz, K.; Hazell, P. Survival and vitality of a macrolichen 14 years after transplantation on aspen trees retained at clearcutting. For. Ecol. Manag. 2013, 291, 436–441. [Google Scholar] [CrossRef]
- Larsson, P.; Solhaug, K.A.; Gauslaa, Y. Winter—The optimal logging season to sustain growth and performance of retained epiphytic lichens in boreal forests. Biol. Conserv. 2014, 180, 108–114. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A. Differences in the susceptibility to light stress between epiphytic lichens of ancient and young boreal forest stands. Functi. Ecol. 1996, 10, 344–354. [Google Scholar] [CrossRef]
- Edman, M.; Eriksson, A.M.; Villard, M.-A. Effects of selection cutting on the abundance and fertility of indicator lichens Lobaria pulmonaria and Lobaria quercizans. J. Appl. Ecol. 2008, 45, 26–33. [Google Scholar] [CrossRef]
- Esseen, P.-A.; Renhorn, K.-E. Mass loss of epiphytic lichen litter in a boreal forest. Ann. Bot. Fenn. 1998, 35, 211–217. [Google Scholar]
- Baysal, A.; Yazici, K. Radioactive contamination in lichens collected from Akçaabat and Çamlihemşin in the Eastern Black Sea region of Turkey, 1994. Toxicol. Environ. Chem. 1995, 48, 145–148. [Google Scholar] [CrossRef]
- Loppi, S.; De Dominicis, V. Lichens as long-term biomonitors of air quality in central Italy. Acta Bot. Neerl. 1996, 45, 563–570. [Google Scholar] [CrossRef]
- Celik, N.; Cevik, U.; Celik, A.; Koz, B. Natural and artificial radioactivity measurements in Eastern Black Sea region of Turkey. J. Hazard. Mater. 2009, 162, 146–153. [Google Scholar] [CrossRef]
- Riga-Karandinos, A.N.; Karandinos, M.G. Caesium-137 concentrations and ecological half-lives in three epiphytic lichen species from Southern Greece (Megalopolis). Bryologist 1998, 101, 422–427. [Google Scholar] [CrossRef]
- Dragović, S.; Mandić, L.J. Transfer of radionuclides to ants, mosses and lichens in semi-natural ecosystems. Radiat. Environ. Biophys. 2010, 49, 625–634. [Google Scholar] [CrossRef]
- Kahraman, A.; Kaynak, G.; Akkaya, G.; Gürler, O.; Yalçın, S. Radioactivity measurements in epiphytic lichens of Uludaǧ Mountain in Western Anatolia. J. Radioanal. Nucl. Chem. 2013, 295, 1057–1066. [Google Scholar] [CrossRef]
- Hilmo, O.; Ely-Aastrup, H.; Hytteborn, H.; Holien, H. Population characteristics of old forest associated epiphytic lichens in Picea abies plantations in the boreal rainforest of Central Norway. Can. J. For. Res. 2011, 41, 1743–1753. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Lie, M.; Solhaug, K.A.; Ohlson, M. Growth and ecophysiological acclimation of the foliose lichen Lobaria pulmonaria in forests with contrasting light climates. Oecologia 2006, 147, 406–416. [Google Scholar] [CrossRef]
- Muir, P.S.; Shirazi, A.M.; Patrie, J. Seasonal growth dynamics in the lichen Lobaria pulmonaria. Bryologist 1997, 100, 458–464. [Google Scholar] [CrossRef]
- Zoller, S.; Lutzoni, F.; Scheidegger, C. Genetic variation within and among populations of the threatened lichen Lobaria pulmonaria in Switzerland and implications for its conservation. Mol. Ecol. 1999, 8, 2049–2059. [Google Scholar] [CrossRef] [PubMed]
- Otálora, M.G.; Martínez, I.; Belinchón, R.; Widmer, I.; Aragón, G.; Escudero, A.; Scheidegger, C. Remnants fragments preserve genetic diversity of the old forest lichen Lobaria pulmonaria in a fragmented Mediterranean mountain forest. Biodivers. Conserv. 2011, 20, 1239–1254. [Google Scholar] [CrossRef]
- Coxson, D.S.; Stevenson, S.K. Influence of high-contrast and low-contrast forest edges on growth rates of Lobaria pulmonaria in the inland rainforest, British Columbia. For. Ecol. Manag. 2007, 253, 103–111. [Google Scholar] [CrossRef]
- Belinchón, R.; Martínez, I.; Aragón, G.; Dimas, J.; Escudero, A. Fragment quality and matrix affect epiphytic performance in a mediterranean forest landscape. Am. J. Bot. 2009, 96, 974–1982. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Bartemucci, P.; Solhaug, K.A. Forest edge-induced damage of cephalo- and cyanolichens in northern temperate rainforests of British Columbia. Can. J. For. Res. 2019, 49, 434–439. [Google Scholar] [CrossRef]
- Barry, M.; McMullin, R.T.; Horn, A. Edge effects on the lichen genus Lobaria in Atlantic Canadian Forests. For. Chron. 2015, 91, 534–540. [Google Scholar] [CrossRef]
- Belinchón, R.; Harrison, P.J.; Mair, L.; Várkonyi, G.; Snäll, T. Local epiphyte establishment and future metapopulation dynamics in landscapes with different spatiotemporal properties. Ecology 2017, 98, 741–750. [Google Scholar] [CrossRef]
- Coxson, D.S.; Stevenson, S.K. Growth rate responses of Lobaria pulmonaria to canopy structure in even-aged and old-growth cedar-hemlock forests of central-interior British Columbia, Canada. For. Ecol. Manag. 2007, 242, 5–16. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Aleffi, M.; Marignani, M.; Rosati, L.; Burrascano, S.; Ravera, S. Lichens and bryophytes as indicators of old-growth features in Mediterranean forests. Plant Biosyst. 2010, 144, 221–233. [Google Scholar] [CrossRef]
- Benesperi, R.; Nascimbene, J.; Lazzaro, L.; Bianchi, E.; Tepsich, A.; Longinotti, S.; Giordani, P. Successful conservation of the endangered forest lichen Lobaria pulmonaria requires knowledge of fine-scale population structure. Fungal Ecol. 2018, 33, 65–71. [Google Scholar] [CrossRef]
- Kiebacher, T.; Keller, C.; Scheidegger, C.; Bergamini, A. Epiphytes in wooded pastures: Isolation matters for lichen but not for bryophyte species richness. PLoS ONE 2017, 12, e0182065. [Google Scholar] [CrossRef]
- Jüriado, I.; Karu, L.; Liira, J. Habitat conditions and host tree properties affect the occurrence, abundance and fertility of the endangered lichen Lobaria pulmonaria in wooded meadows of Estonia. Lichenologist 2012, 44, 263–276. [Google Scholar] [CrossRef]
- Eaton, S.; Ellis, C.J. High demographic rates of the model epiphyte Lobaria pulmonaria in an oceanic hazelwood (western Scotland). Fungal Ecol. 2014, 11, 60–70. [Google Scholar] [CrossRef]
- Öckinger, E.; Niklasson, M.; Nilsson, S. Is local distribution of the epiphytic lichen Lobaria pulmonaria limited by dispersal capacity or habitat quality? Biodivers. Conserv. 2005, 14, 759–773. [Google Scholar] [CrossRef]
- Werth, S.; Wagner, H.H.; Gugerli, F.; Holderegger, R.; Csencsics, D.; Kalwij, J.M.; Scheidegger, C. Quantifying dispersal and establishment limitation in a population of an epiphytic lichen. Ecology 2006, 87, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Jüriado, I.; Liira, J.; Csencsics, D.; Widmer, I.; Adolf, C.; Kohv, K.; Scheidegger, C. Dispersal ecology of the endangered woodland lichen Lobaria pulmonaria in managed hemiboreal forest landscape. Biodivers. Conserv. 2011, 20, 1803–1819. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Holien, H.; Ohlson, M.; Solhøy, T. Does snail grazing affect growth of the old forest lichen Lobaria pulmonaria? Lichenologist 2006, 38, 587–593. [Google Scholar] [CrossRef]
- Asplund, J.; Gauslaa, Y. Content of secondary compounds depends on thallus size in the foliose lichen Lobaria pulmonaria. Lichenologist 2007, 39, 273–278. [Google Scholar] [CrossRef]
- Asplund, J.; Gauslaa, Y. Mollusc grazing limits growth and early development of the old forest lichen Lobaria pulmonaria in broadleaved deciduous forests. Oecologia 2008, 155, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Vatne, S.; Solhøy, T.; Asplund, J.; Gauslaa, Y. Grazing damage in the old forest lichen Lobaria pulmonaria increases with gastropod abundance in deciduous forests. Lichenologist 2006, 42, 615–619. [Google Scholar] [CrossRef]
- Asplund, J. Snails avoid the medulla of Lobaria pulmonaria and L. scrobiculata due to presence of secondary compounds. Fungal Ecol. 2011, 4, 356–358. [Google Scholar] [CrossRef]
- Clyne, A.B.; Cleavitt, N.L.; Fahey, T.J. Terrestrial Gastropod Grazing on Macrolichens in a Northern Broadleaf-Conifer Forest. Northeast. Nat. 2019, 26, 261–274. [Google Scholar] [CrossRef]
- Asplund, J.; Gauslaa, Y.; Merinero, S. The role of fungal parasites in tri-trophic interactions involving lichens and lichen-feeding snails. New Phytol. 2016, 211, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Asplund, J.; Solhaug, K.A.; Gauslaa, Y. Fungal depsidones—An inducible or constitutive defence against herbivores in the lichen Lobaria pulmonaria? Basic Appl. Ecol. 2009, 10, 273–278. [Google Scholar] [CrossRef]
- Vatne, S.; Asplund, J.; Gauslaa, Y. Contents of carbon based defence compounds in the old forest lichen Lobaria pulmonaria vary along environmental gradients. Fungal Ecol. 2011, 4, 350–355. [Google Scholar] [CrossRef]
- Asplund, J.; Wardle, D.A. Contrasting changes in palatability following senescence of the lichenized fungi Lobaria pulmonaria and L. scrobiculata. Fungal Ecol. 2012, 5, 710–713. [Google Scholar] [CrossRef]
- Asplund, J. Chemical races of Lobaria pulmonaria differ in palatability to gastropods. Lichenologist 2011, 43, 491–494. [Google Scholar] [CrossRef]
- Gauslaa, Y. Mollusc grazing may constrain the ecological niche of the old forest lichen Pseudocyphellaria crocata. Plant Biol. 2008, 10, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, C.; Scheidegger, C. Morphological aspects associated with repair and regeneration in Lobaria pulmonaria and L. amplissima (Peltigerales, Ascomycota). Lichenologist 2013, 45, 285–289. [Google Scholar] [CrossRef]
- Asplund, J.; Larsson, P.; Vatne, S.; Gauslaa, Y. Gastropod grazing shapes the vertical distribution of epiphytic lichens in forest canopies. J. Ecol. 2010, 98, 218–225. [Google Scholar] [CrossRef]
- Boch, S.; Prati, D.; Werth, S.; Rüetschi, J.; Fischer, M. Lichen Endozoochory by Snails. PLoS ONE 2011, 6, e18770. [Google Scholar] [CrossRef] [PubMed]
- Cansaran-Duman, D.; Altunkaynak, E.; Aslan, A.; Büyük, I.; Aras, S. Application of molecular markers to detect DNA damage caused by environmental pollutants in lichen species. Genet. Mol. Res. 2015, 14, 4637–4650. [Google Scholar] [CrossRef]
- Hamutoğlu, R.; Aslan, A.; Aras, S.; Cansaran-Duman, D. Environmental risk assessment under the pollutants exposure with using four lichen species and molecular assay in cement plant, Askale-Erzurum (Turkey). Turk. Hijyen Deneysel Biyoloji Dergisi 2016, 73, 253–266. [Google Scholar] [CrossRef]
- Farmer, A.M.; Bates, J.W.; Bell, J.N.B. The transplantation of four species of Lobaria lichens to demonstrate a field acid rain effect. Environ. Sci. Stud. 1992, 50, 295–300. [Google Scholar] [CrossRef]
- Farmer, A.M.; Bates, J.W.; Bell, J.N.B. Seasonal variations in acidic pollutant inputs and their effects on the chemistry of stemflow, bark and epiphyte tissues in three oak woodlands in N.W. Britain. New Phytol. 1991, 118, 441–451. [Google Scholar] [CrossRef]
- Ignatenko, R.V.; Tarasova, V.N. Assessment of the State of Lung Lichen, Lobaria pulmonaria (L.) Hoffm., in Forest Communities with Different Times Since Disturbance in the Northeast of European Russia. Russ. J. Ecol. 2018, 49, 277–285. [Google Scholar] [CrossRef]
- Bolli, J.C.; Wagner, H.H.; Kalwij, J.M.; Silke, W.; Cherubini, P.; Scheidegger, C.; Rigling, A. Growth dynamics after historic disturbance in a montane forest and its implications for an endangered epiphytic lichen. Bot. Helv. 2008, 118, 111–127. [Google Scholar] [CrossRef][Green Version]
- Singh, G.; Dal Grande, F.; Werth, S.; Scheidegger, C. Long-term consequences of disturbances on reproductive strategies of the rare epiphytic lichen Lobaria pulmonaria: Clonality a gift and a curse. FEMS Microbiol. Ecol. 2015, 91, 1–11. [Google Scholar] [CrossRef]
- Walser, J.-C.; Gugerli, F.; Holderegger, R.; Kuonen, D.; Scheidegger, C. Recombination and clonal propagation in different populations of the lichen Lobaria pulmonaria. Heredity 2004, 93, 322–329. [Google Scholar] [CrossRef][Green Version]
- Werth, S.; Wagner, H.H.; Holderegger, R.; Kalwij, J.M.; Scheidegger, C. Effect of disturbances on the genetic diversity of an old-forest associated lichen. Mol. Ecol. 2006, 15, 911–921. [Google Scholar] [CrossRef]
- Wagner, H.H.; Werth, S.; Kalwij, J.M.; Bolli, J.C.; Scheidegger, C. Modelling forest recolonization by an epiphytic lichen using a landscape genetic approach. Landsc. Ecol. 2006, 21, 849–865. [Google Scholar] [CrossRef]
- Forman, R.T.T. Canopy lichens with blue-green algae: A nitrogen source in a Colombian rainforest. Ecology 1975, 56, 1176–1184. [Google Scholar] [CrossRef]
- Holub, S.M.; Lajtha, K. The fate and retention of organic and inorganic 15N nitrogen in an old-growth forest soil in western Oregon. Ecosystems 2004, 7, 368–380. [Google Scholar] [CrossRef]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Budel, B.; Andreae, M.O.; Poschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Culberson, C.F. Chemical studies in the genus Lobaria and the occurrence of a new tridepside, 4-O-methylgyrophoric acid. Bryologist 1969, 72, 19–27. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Rose, F. Qualitative scale for estimating sulphur dioxide air pollution in England and Wales using epiphytic lichens. Nature 1970, 227, 145–148. [Google Scholar] [CrossRef] [PubMed]
- James, P.W.; Wolseley, P.A. Acidification and the Lobarion: A case for biological monitoring. Br. Lich. Soc. Bull. 1992, 71, 4–12. [Google Scholar]
- Liška, J.; Pišút, I. An example of bioindication on a large scale: Mapping of lichens in Czechoslovakia. In Proceedings of the Vth International Conference Bioindicators Deteriorisations Regionis; Boháč, J., Růžička, V., Eds.; Institute of Landscape Ecology CAS: České Budějovice, Czech Republic, 1989; pp. 81–86. [Google Scholar]
- Farkas, E.; Varga, N.; Veres, K.; Matus, G.; Sinigla, M.; Lőkös, L. Distribution types of lichens in Hungary that indicate changing environmental conditions. J. Fungi 2022, 8, 600. [Google Scholar] [CrossRef]
- Root, H.T.; McGee, G.G.; Nyland, R.D. Effects of two silvicultural regimes with large tree retention on epiphytic macrolichen communities in Adirondack northern hardwoods, New York, USA. Can. J. For. Res. 2007, 37, 1854–1866. [Google Scholar] [CrossRef]
- Schiefelbein, U.; Arnfred, T.; Dolnik, C.; Neumann, P.; Ossowska, E.; Poulsen, R.; Søchting, U.; Thell, A. Lobaria pulmonaria (L.) Hoffm. in the southwestern Baltic–Kattegat area. Folia Cryptogam. Est. 2021, 58, 183–197. [Google Scholar] [CrossRef]
- Lõhmus, A.; Runnel, K. Ash dieback can rapidly eradicate isolated epiphyte populations in production forests: A case study. Biol. Conserv. 2014, 169, 185–188. [Google Scholar] [CrossRef]
- Young, A.G.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Franklin, C.M.; Harper, K.A.; Clarke, M.J. Trends in studies of edge influence on vegetation at human-created and natural forest edges across time and space. Can. J. For. Res. 2021, 51, 274–282. [Google Scholar] [CrossRef]
- Matlack, G.R.; Litvaitis, J.A. Forest edges. In Maintaining Biodiversity in Forest Ecosystems; Hunter, M.L., Ed.; Cambridge University Press: Cambridge, UK, 1999; pp. 210–233. [Google Scholar] [CrossRef]
- Aragón, G.; Abuja, L.; Belinchón, R.; Martínez, I. Edge type determines the intensity of forest edge effect on epiphytic communities. Eur. J. For. Res. 2015, 134, 443–451. [Google Scholar] [CrossRef]
- Nascimbene, J.; Casazza, G.; Benesperi, R.; Catalano, I.; Cataldo, D.; Grillo, M.; Isocrono, D.; Matteucci, E.; Ongaro, S.; Potenza, G.; et al. Climate change fosters the decline of epiphytic Lobaria species in Italy. Biol. Conserv. 2016, 201, 377–384. [Google Scholar] [CrossRef]
- Otálora, M.G.; Belinchón, R.; Prieto, M.; Aragón, G.; Izquierdo, P.; Martínez, I. The threatened epiphytic lichen Lobaria pulmonaria in the Iberian Peninsula: Genetic diversity and structure across a latitudinal gradient. Fungal Biol. 2015, 119, 802–811. [Google Scholar] [CrossRef]
- Pezzi, G.; Gambini, S.; Buldrini, F.; Ferretti, F.; Muzzi, E.; Maresi, G.; Nascimbene, J. Contrasting patterns of tree features, lichen, and plant diversity in managed and abandoned old-growth chestnut orchards of the northern Apennines (Italy). For. Ecol. Manag. 2020, 470471, 118207. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ecosystem Functioning and Services | Air Pollution Monitoring | Forest Management |
|---|---|---|
| N2 fixation: 12 [27,28,29,30,31,32,33,34,35,36,37,38] | Monitoring L. pulmonaria: 3 [39,40,41] | Monitoring L. pulmonaria: 10 [19,20,28,42,43,44,45,46,47,48] |
| C pool: 2 [29,49] | Biomonitoring PTEs, PAHs, HCHO: 14 [50,51,52,53,54,55,56,57,58,59,60,61,62,63] | Biomonitoring forest treatments: 15 [9,24,25,64,65,66,67,68,69,70,71,72,73,74,75] |
| Biomass: 6 [27,28,29,30,31,76] | Biomonitoring radioactivity: 6 [77,78,79,80,81,82] | Biomonitoring environmental and structural variables: 22 [23,25,43,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101] |
| Feeding: 15 [102,103,104,105,106,107,108,109,110,111,112,113,114,115,116] | Biomonitoring phytotoxic pollutants and acid rains: 4 [117,118,119,120] | Biomonitoring historical or actual disturbances: 6 [121,122,123,124,125,126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravera, S.; Benesperi, R.; Bianchi, E.; Brunialti, G.; Di Nuzzo, L.; Frati, L.; Giordani, P.; Isocrono, D.; Nascimbene, J.; Vallese, C.; et al. Lobaria pulmonaria (L.) Hoffm.: The Multifaceted Suitability of the Lung Lichen to Monitor Forest Ecosystems. Forests 2023, 14, 2113. https://doi.org/10.3390/f14102113
Ravera S, Benesperi R, Bianchi E, Brunialti G, Di Nuzzo L, Frati L, Giordani P, Isocrono D, Nascimbene J, Vallese C, et al. Lobaria pulmonaria (L.) Hoffm.: The Multifaceted Suitability of the Lung Lichen to Monitor Forest Ecosystems. Forests. 2023; 14(10):2113. https://doi.org/10.3390/f14102113
Chicago/Turabian StyleRavera, Sonia, Renato Benesperi, Elisabetta Bianchi, Giorgio Brunialti, Luca Di Nuzzo, Luisa Frati, Paolo Giordani, Deborah Isocrono, Juri Nascimbene, Chiara Vallese, and et al. 2023. "Lobaria pulmonaria (L.) Hoffm.: The Multifaceted Suitability of the Lung Lichen to Monitor Forest Ecosystems" Forests 14, no. 10: 2113. https://doi.org/10.3390/f14102113
APA StyleRavera, S., Benesperi, R., Bianchi, E., Brunialti, G., Di Nuzzo, L., Frati, L., Giordani, P., Isocrono, D., Nascimbene, J., Vallese, C., & Paoli, L. (2023). Lobaria pulmonaria (L.) Hoffm.: The Multifaceted Suitability of the Lung Lichen to Monitor Forest Ecosystems. Forests, 14(10), 2113. https://doi.org/10.3390/f14102113