
Changes in Soil Substrate and Microbial Properties Associated with Permafrost Thaw Reduce Nitrogen Mineralization

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sampling and Plant Community Survey
2.3. Soil Incubation and Measurements
2.4. Statistical Analyses
3. Results
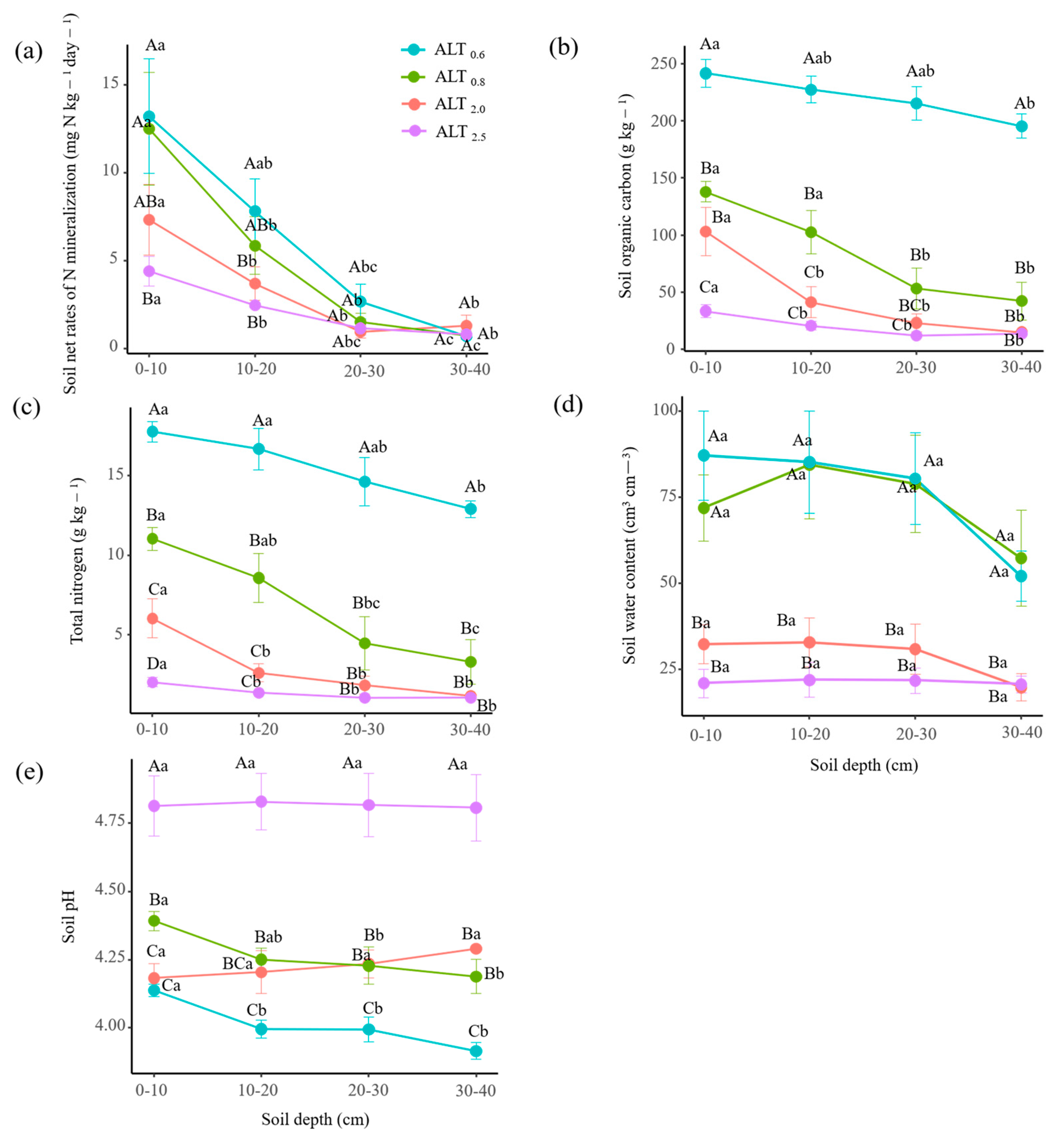
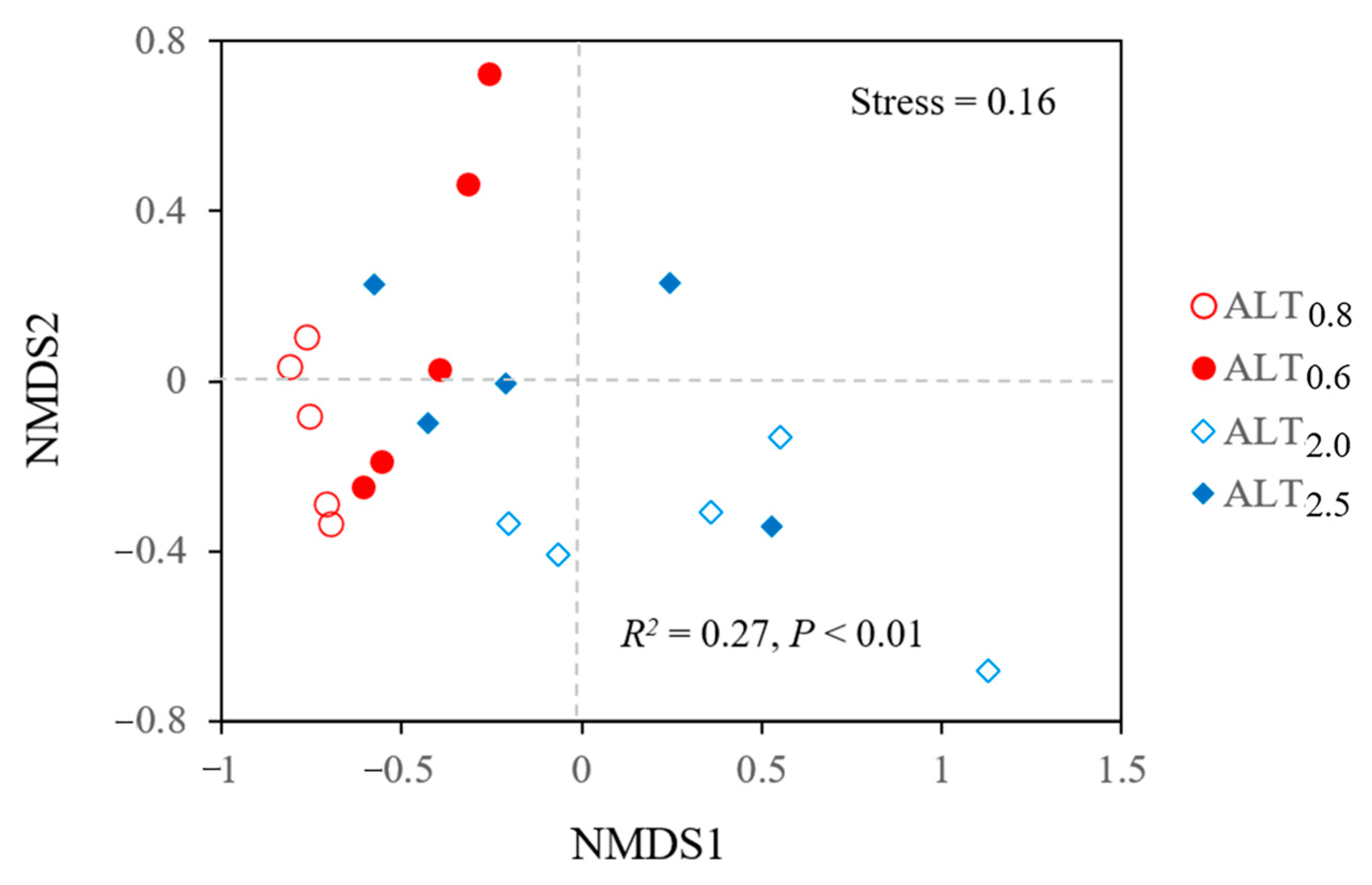
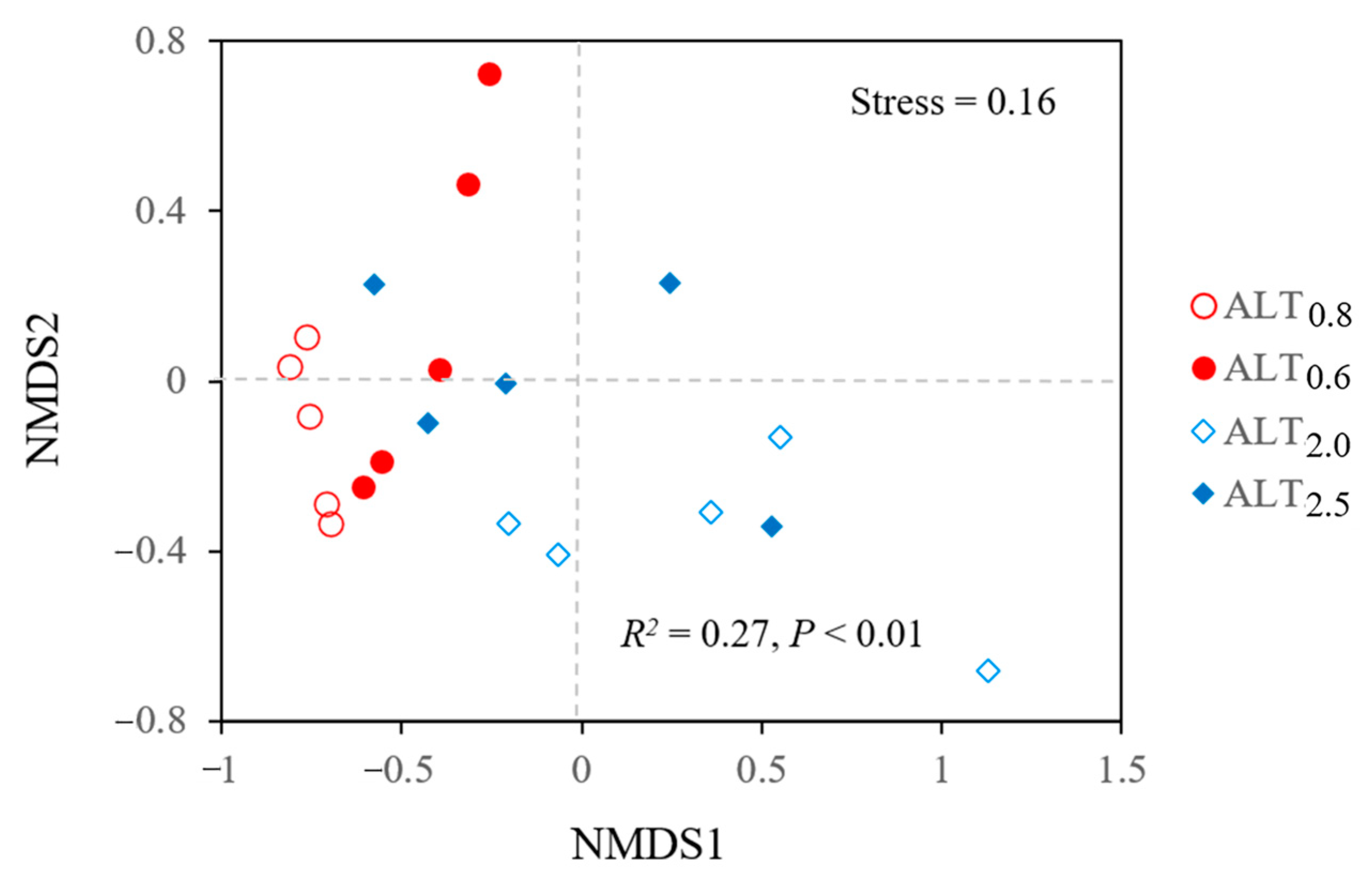
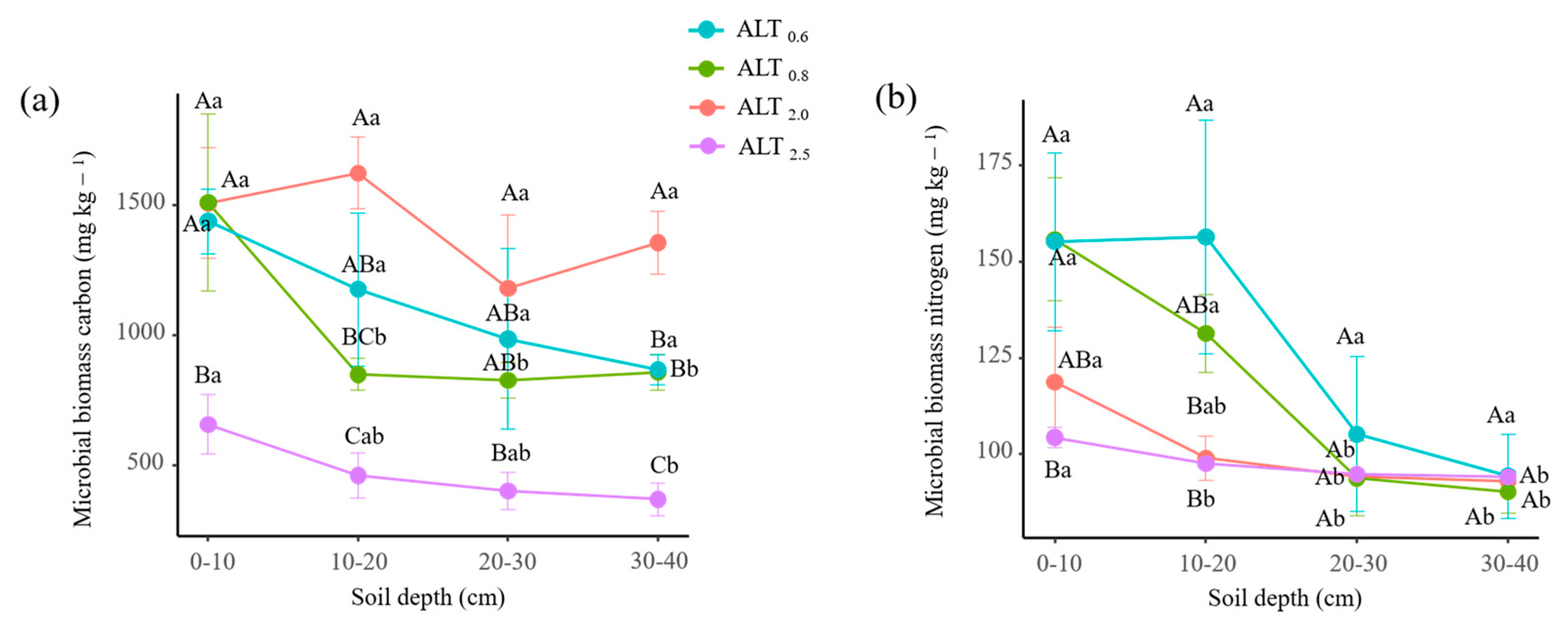
3.1. Variations in Nmin, Soil Physiochemical Properties, Microbial Biomass, and Vegetation
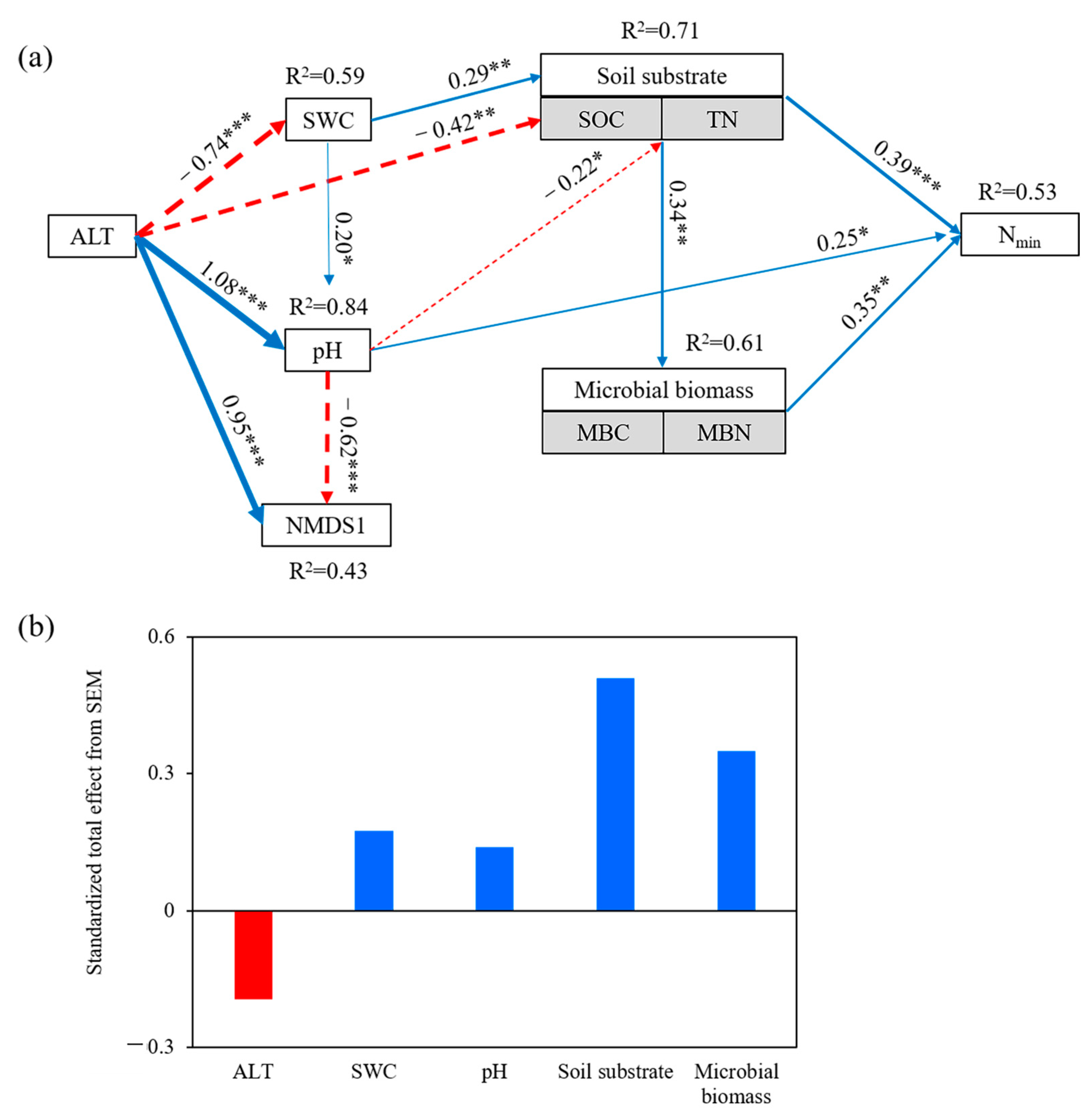
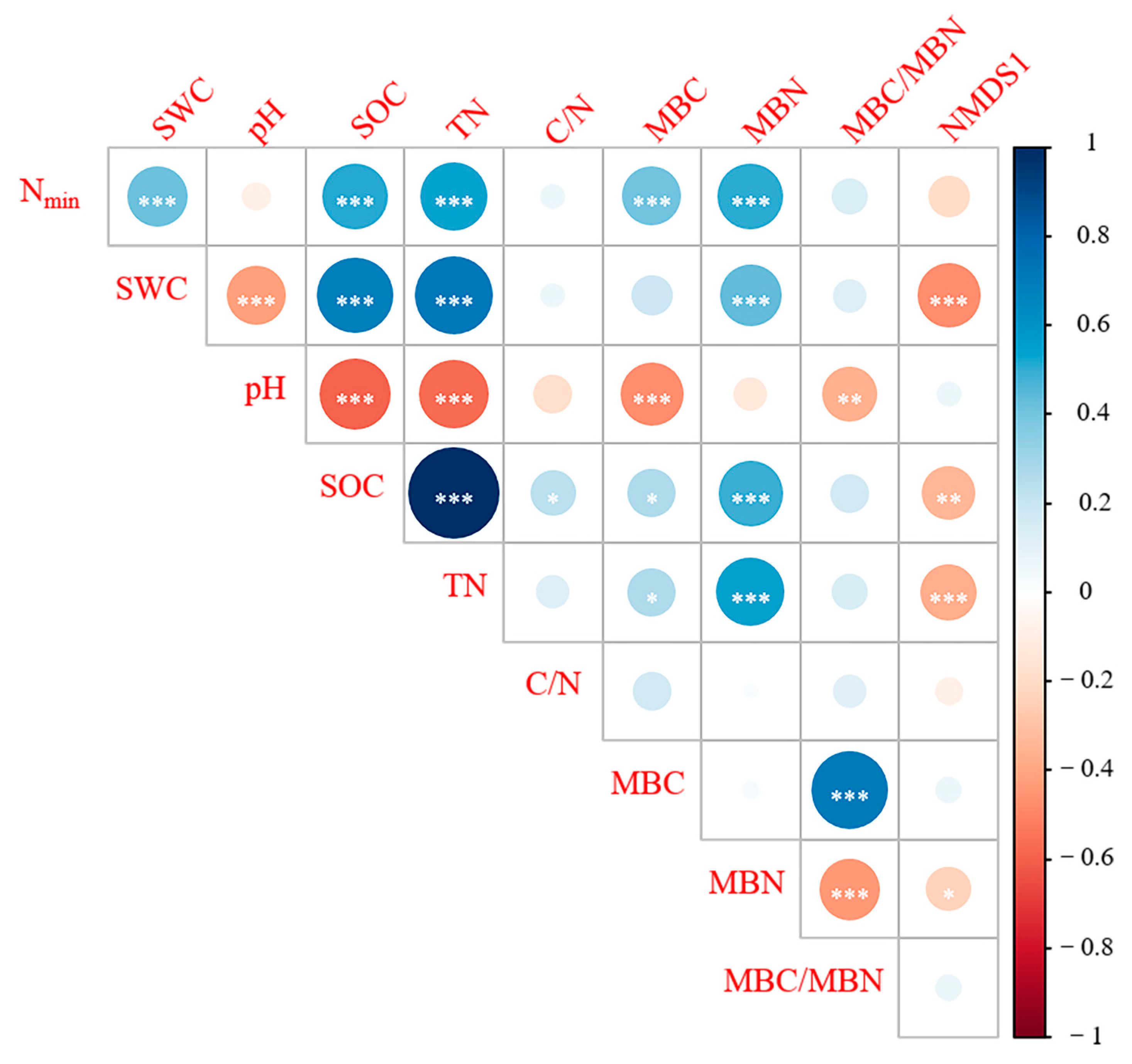
3.2. Relationships among Vegetation, Soil Physicochemical Properties, Microbial Biomass, and Nmin
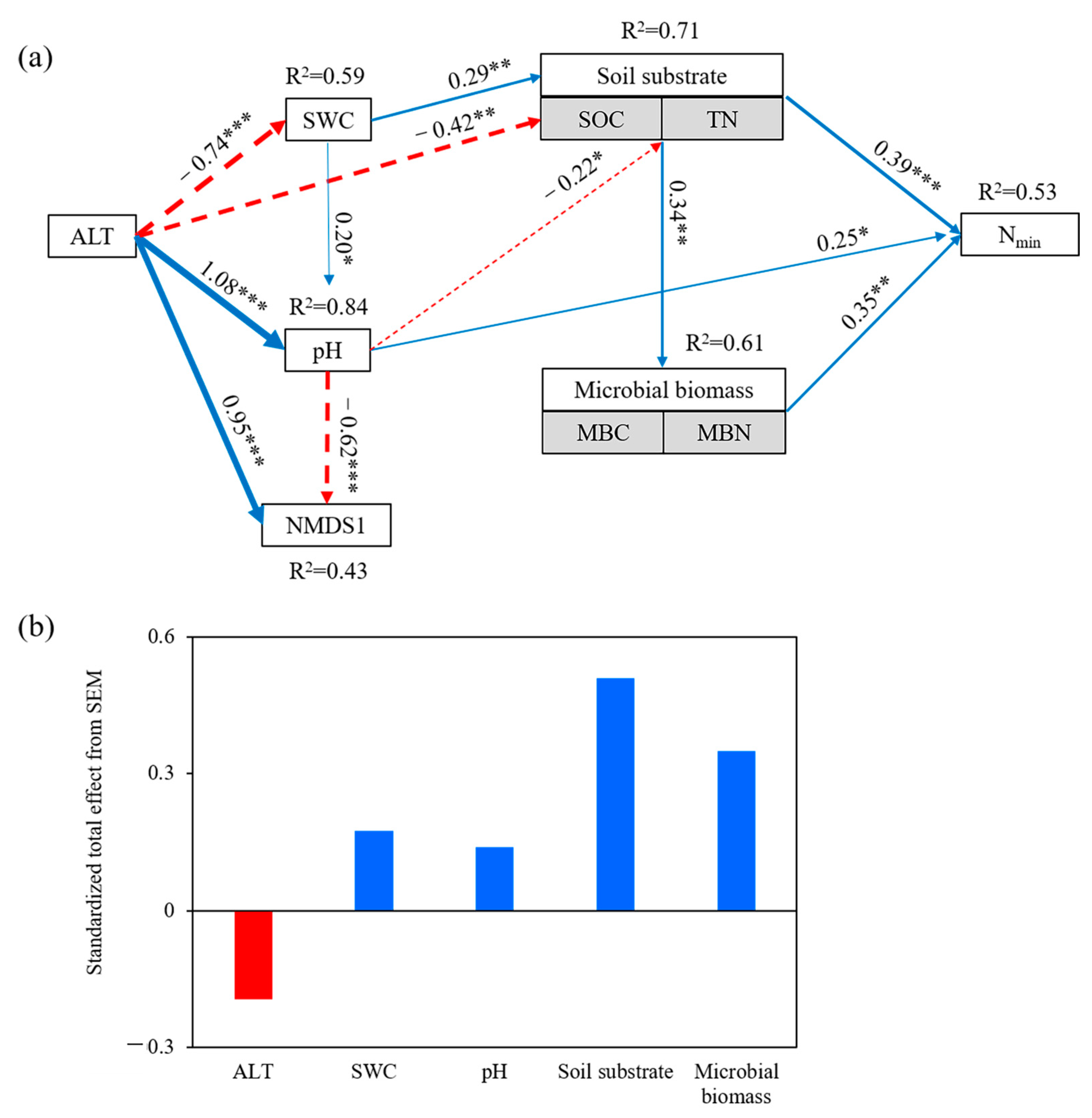
3.3. Direct and Indirect Effects of Nmin Drivers
4. Discussion
4.1. Changes in Soil Physiochemistry and Microbial Biomass with Permafrost Thaw
4.2. Drivers of Soil N Mineralization Rates (Nmin) with Permafrost Thaw
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Sites | Coordinate | Elevation (m a.s.l.) | Plant | Active Layer Thickness (ALT) m |
|---|---|---|---|---|
| Site 1 | 125°08′10.000″ E 51°07′54.970″ N | 445 | Carex tato and shrubs | 0.6 |
| Site 2 | 125°08′09.911″ E 51°07′54.970″ N | 445 | Carex tato | 0.8 |
| Site 3 | 125°08′09.104″ E 51°07′44.776″ N | 452 | Betula platyphylla and Larix gmelinii (Rupr.) Kuzen | 2.0 |
| Site 4 | 125°08′16.943″ E 51°07′27.471″ N | 474 | Larix gmelinii (Rupr.) Kuzen | 2.5 |
| Group | Variables | PC1 Score/% | PC2 Score/% |
|---|---|---|---|
| Soil substrate | SOC, TN | 0.99 | 0.01 |
| Microbial biomass | MBC, MBN | 0.52 | 0.48 |

References
- Obu, J.; Westermann, S.; Bartsch, A.; Berdnikov, N.; Christiansen, H.H.; Dashtseren, A.; Delaloye, R.; Elberling, B.; Etzelmüller, B.; Kholodov, A.; et al. Northern Hemisphere permafrost map based on TTOP modelling for 2000–2016 at 1 km2 scale. Earth-Sci. Rev. 2019, 193, 299–316. [Google Scholar] [CrossRef]
- Biskaborn, B.; Smith, S.; Noetzli, J.; Matthes, H.; Vieira, G.; Streletskiy, D.; Schoeneich, P.; Romanovsky, V.; Lewkowicz, A.; Abramov, A.; et al. Permafrost is warming at a global scale. Nat. Commun. 2019, 10, 264. [Google Scholar] [CrossRef]
- Chadburn, S.E.; Burke, E.J.; Cox, P.M.; Friedlingstein, P.; Hugelius, G.; Westermann, S. An observation-based constraint on permafrost loss as a function of global warming. Nat. Clim. Chang. 2017, 7, 340–345. [Google Scholar] [CrossRef]
- Schuur, E.A.; McGuire, A.D.; Schädel, C.; Grosse, G.; Harden, J.W.; Hayes, D.J.; Hugelius, G.; Koven, C.D.; Kuhry, P.; Lawrence, D.M.; et al. Climate change and the permafrost carbon feedback. Nature 2015, 520, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Kicklighter, D.W.; Melillo, J.M.; Monier, E.; Sokolov, A.P.; Zhuang, Q. Future nitrogen availability and its effect on carbon sequestration in Northern Eurasia. Nat. Commun. 2019, 10, 3024. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.E.; Nadelhoffer, K.J.; Högberg, P.A. Synthesis: The role of nutrients as constraints on carbon balances in boreal and arctic regions. Plant Soil 2002, 242, 163–170. [Google Scholar] [CrossRef]
- Keuper, F.; van Bodegom, P.M.; Dorrepaal, E.; Weedon, J.T.; van Hal, J.; van Logtestijn, R.S.; Aerts, R. A frozen feast: Thawing permafrost increases plant-available nitrogen in subarctic peatlands. Glob. Chang. Biol. 2012, 18, 1998–2007. [Google Scholar] [CrossRef]
- Salmon, V.G.; Soucy, P.; Mauritz, M.; Celis, G.; Natali, S.M.; Mack, M.C.; Schuur, E.A.G. Nitrogen availability increases in a tundra ecosystem during five years of experimental permafrost thaw. Glob. Chang. Biol. 2016, 22, 1927–1941. [Google Scholar] [CrossRef] [PubMed]
- Rousk, K.; Michelsen, A.; Rousk, J. Microbial control of soil organic matter mineralization responses to labile carbon in subarctic climate change treatments. Glob. Chang. Biol. 2016, 22, 4150–4161. [Google Scholar] [CrossRef]
- Ramm, E.; Liu, C.; Ambus, P.; Butterbach-Bahl, K.; Hu, B.; Martikainen, P.J.; Marushchak, M.E.; Mueller, C.W.; Rennenberg, H.; Schloter, M.; et al. A review of the importance of mineral nitrogen cycling in the plant-soil-microbe system of permafrost-affected soils-changing the paradigm. Environ. Res. Lett. 2022, 17, 013004. [Google Scholar] [CrossRef]
- Mao, C.; Kou, D.; Chen, L.; Qin, S.; Zhang, D.; Peng, Y.; Yang, Y. Permafrost nitrogen status and its determinants on the Tibetan Plateau. Glob. Chang. Biol. 2020, 26, 5290–5302. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Hogues, H.; Whyte, L.G.; Greer, C.W. The functional potential of high Arctic permafrost revealed by metagenomic sequencing, qPCR and microarray analyses. ISME J. 2010, 4, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.J.E.; Wanek, W.; Zappe, A.; Richter, A.; Svenning, M.M.; Schleper, C.; Urich, T. Nitrification rates in Arctic soils are associated with functionally distinct populations of ammonia-oxidizing archaea. ISME J. 2013, 7, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Hultman, J.; Waldrop, M.P.; Mackelprang, R.; David, M.M.; McFarland, J.; Blazewicz, S.J.; Harden, J.; Turetsky, M.R.; McGuire, A.D.; Shah, M.B.; et al. Multi-omics of permafrost, active layer and thermokarst bog soil microbiomes. Nature 2015, 521, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Keuper, F.; Dorrepaal, E.; van Bodegom, P.M.; van Logtestijn, R.; Venhuizen, G.; van Hal, J.; Aerts, R. Experimentally increased nutrient availability at the permafrost thaw front selectively enhances biomass production of deep-rooting subarctic peatland species. Glob. Chang. Biol. 2017, 23, 4257–4266. [Google Scholar] [CrossRef]
- Standen, K.M.; Baltzer, J.L. Permafrost condition determines plant community composition and community-level foliar functional traits in a boreal peatland. Ecol. Evol. 2021, 11, 10133–10146. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D.; Hobbie, S.E. Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J. Ecol. 2009, 97, 48–56. [Google Scholar] [CrossRef]
- Yun, H.; Zhu, Q.; Tang, J.; Zhang, W.; Chen, D.; Ciais, P.; Wu, Q.; Elberling, B. Warming, permafrost thawing and nitrogen availability are drivers of increasing plant growth and species richness on the Tibetan Plateau. Soil Biol. Biochem. 2023, 182, 109041. [Google Scholar] [CrossRef]
- O’Donnell, J.A.; Jorgenson, M.T.; Harden, J.W.; McGuire, A.D.; Kanevskiy, M.Z.; Wickland, K.P. The effects of permafrost thaw on soil hydrologic, thermal, and carbon dynamics in an Alaskan peatland. Ecosystems 2012, 15, 213–229. [Google Scholar] [CrossRef]
- Finger, R.A.; Turetsky, M.R.; Kielland, K.; Ruess, R.W.; Mack, M.C.; Euskirchen, E.S. Effects of permafrost thaw on nitrogen availability and plant-soil interactions in a boreal Alaskan lowland. J. Ecol. 2016, 104, 1542–1554. [Google Scholar] [CrossRef]
- Che, L.; Cheng, M.; Xing, L.; Cui, Y.; Wan, L. Effects of permafrost degradation on soil organic matter turnover and plant growth. Catena 2022, 208, 105721. [Google Scholar] [CrossRef]
- Jin, H.; Li, S.; Cheng, G.; Wang, S.; Li, X. Permafrost and climatic change in China. Glob. Planet. Chang. 2000, 26, 387–404. [Google Scholar] [CrossRef]
- Jin, H.; Yu, Q.; Lu, L.; Guo, D.; He, R.; Yu, S.; Sun, G.; Li, Y. Degradation of permafrost in the Xing’anling Mountains, Northeastern China. Permafr. Periglac. Process. 2007, 18, 245–258. [Google Scholar] [CrossRef]
- Jin, H.; Chang, X.; Luo, D.; He, R.; Lu, L.; Yang, S.; Guo, D.; Chen, X.; Harris, S.A. Evolution of permafrost in North east China since the late Pleistocene. Sci. Cold Arid Reg. 2016, 84, 269–296. [Google Scholar]
- Zhang, Z.; Wu, Q.; Hou, M.; Tai, B.; An, Y. Permafrost change in Northeast China in the 1950s–2010s. Adv. Clim. Chang. Res. 2021, 12, 18–28. [Google Scholar] [CrossRef]
- Jin, H.; Sun, G.; Yu, S.; Jin, R.; He, R. Symbiosis of marshes and permafrost in Da and Xiao Hinggan Mountains in North eastern China. Chin. Geogr. Sci. 2008, 18, 62–69. [Google Scholar] [CrossRef]
- Chen, S.; Zang, S.; Sun, L. Characteristics of permafrost degradation in Northeast China and its ecological effects: A review. Sci. Cold Arid. Reg. 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Q.; Ma, Z.; Jin, H.; Chang, X.; Marchenko, S.S.; Spektor, V.V. Seasonal variations in temperature sensitivity of soil respiration in a larch forest in the Northern Daxing’an Mountains in Northeast China. J. For. Res. 2021, 33, 1061–1070. [Google Scholar] [CrossRef]
- Zhou, Y.; Qiu, G.; Guo, D.; Cheng, G.; Li, S. Geocryology in China; Science Press: Beijing, China, 2000. (In Chinese) [Google Scholar] [CrossRef]
- He, R.; Jin, H.; Luo, D.; Li, X.; Zhou, C.; Jia, N.; Jin, X.; Li, X.; Che, T.; Yang, X.; et al. Permafrost changes in the Nanwenghe Wetlands Reserve on the southern slope of the Da Xing’anling-Yile’huli mountains, Northeast China. Adv. Clim. Chang. Res. 2021, 12, 696–709. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 May 2022).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models, R package version 3.1-153; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://CRAN.R-project.org/package=nlme> (accessed on 21 January 2022).
- Harrell, F.E., Jr. Hmisc: Harrell Miscellaneous, R package version 4.6-0; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 22 February 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package, R package version 2.5-7; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=vegan (accessed on 21 April 2021).
- Liu, Y.; He, N.; Wen, X.; Xu, L.; Sun, X.; Yu, G.; Liang, L.; Schipper, L.A. The optimum temperature of soil microbial respiration: Patterns and controls. Soil Biol. Biochem. 2018, 121, 35–42. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Yu, Z.; Li, Y.; Tian, D.; Wang, B.; Li, Z.; Niu, S. Vegetation type controls root turnover in global grasslands. Glob. Ecol. Biogeogr. 2019, 28, 442–455. [Google Scholar] [CrossRef]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modeling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Salmon, V.G.; Schädel, C.; Bracho, R.; Pegoraro, E.; Celis, G.; Mauritz, M.; Mack, M.C.; Schuur, E.A.G. Adding depth to our understanding of nitrogen dynamics in permafrost soils. J. Geophys. Res-Biogeo. 2018, 123, 2497–2512. [Google Scholar] [CrossRef]
- Chang, X.; Jin, H.; Zhang, Y.; He, R.; Luo, D.; Wang, Y.; Lü, L.; Zhang, Q. Thermal impacts of boreal forest vegetation on active layer and permafrost soils in northern Da Xing’anling (Hinggan) Mountains, Northeast China. Arct. Antarct. Alp. Res. 2015, 47, 267–279. [Google Scholar] [CrossRef]
- Aalto, J.; Karjalainen, O.; Hjort, J.; Luoto, M. Statistical forecasting of current and future circum-Arctic ground temperatures and active layer thickness. Geophys. Res. Lett. 2018, 45, 4889–4898. [Google Scholar] [CrossRef]
- Voigt, C.; Marushchak, M.E.; Mastepanov, M.; Lamprecht, R.E.; Christensen, T.R.; Dorodnikov, M.; Jackowicz-Korczyński, M.; Lindgren, A.; Lohila, A.; Nykänen, H.; et al. Ecosystem carbon response of an Arctic peatland to simulated permafrost thaw. Glob. Chang. Biol. 2019, 25, 1746–1764. [Google Scholar] [CrossRef]
- Voigt, C.; Marushchak, M.E.; Abbott, B.W.; Biasi, C.; Elberling, B.; Siciliano, S.D.; Sonnentag, O.; Stewart, K.J.; Yang, Y.; Martikainen, P.J. Nitrous oxide emissions from permafrost-affected soils. Nat. Rev. Earth Env. 2020, 1, 420–434. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Ramm, E.; Liu, C.; Mueller, C.W.; Gschwendtner, S.; Yue, H.; Wang, X.; Bachmann, J.; Bohnhoff, J.A.; Ostler, J.; Schloter, J.; et al. Alder-induced stimulation of soil gross nitrogen turnover in a permafrost-affected peatland of Northeast China. Soil Biol. Biochem. 2022, 172, 108757. [Google Scholar] [CrossRef]
- Wu, M.; Qu, D.; Li, T.; Liu, F.; Gao, Y.; Chen, S.; Chen, T. Effects of permafrost degradation on soil microbial biomass carbon and nitrogen in the Shule River headwaters, the Qilian Mountains. Sci. Geogr. Sin. 2021, 41, 177–186. (In Chinese) [Google Scholar] [CrossRef]
- Xu, W.; Yuan, W. Responses of microbial biomass carbon and nitrogen to experimental warming: A meta-analysis. Soil Biol. Biochem. 2017, 15, 265–274. [Google Scholar] [CrossRef]
- Davidson, E.A.; Samanta, S.; Caramori, S.S.; Savage, K. The Dual Arrhenius and Michaelis-M enten kinetics model for decomposition of soil organic matter at hourly to seasonal time scales. Glob. Chang. Biol. 2012, 18, 371–384. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef]
- Sistla, S.A.; Moore, J.C.; Simpson, R.T.; Gough, L.; Shaver, G.R.; Schimel, J.P. Long-term warming restructures Arctic tundra without changing net soil carbon storage. Nature 2013, 497, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Yuan, W.; Dong, W.; Xia, J.; Liu, D.; Chen, Y. A meta-analysis of the response of soil moisture to experimental warming. Environ. Res. Lett. 2013, 8, 044027. [Google Scholar] [CrossRef]
- Deslippe, J.R.; Hartmann, M.; Simard, S.W.; Mohn, W.W. Long-term warming alters the composition of Arctic soil microbial communities. FEMS Microbiol. Ecol. 2012, 82, 303–315. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, X.; Yang, Q.; Luo, Y.; Fang, C.; Chen, J.; Yang, X.; Li, B. Responses of ecosystem carbon cycle to experimental warming: A meta-analysis. Ecology 2013, 94, 726–738. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, Z.; Fu, G. A meta-analysis of the effects of experimental warming on soil carbon and nitrogen dynamics on the Tibetan Plateau. Appl. Soil Ecol. 2015, 87, 32–38. [Google Scholar] [CrossRef]
- Xu, X.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Wu, X.; Fang, H.; Zhao, Y.; Smoak, J.M.; Li, W.; Shi, W.; Sheng, Y.; Zhao, L.; Ding, Y. A conceptual model of the controlling factors of soil organic carbon and nitrogen densities in a permafrost-affected region on the eastern Qinghai-Tibetan Plateau. J. Geophys. Res.-Biogeo. 2017, 122, 1705–1717. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, J.; Meng, L.; Müller, C.; Cai, Z. Variations of soil N transformation and N2O emissions in tropical secondary forests along an aridity gradient. J. Soil. Sediment. 2015, 15, 1538–1548. [Google Scholar] [CrossRef]
- Elrys, A.S.; Ali, A.; Zhang, H.; Cheng, Y.; Zhang, J.; Cai, Z.C.; Müller, C.; Chang, S.X. Patterns and drivers of global gross nitrogen mineralization in soils. Glob. Chang. Biol. 2021, 27, 5950–5962. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Wanek, W.; Haemmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon: Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Baligar, V.C.; Jones, C.A. Growth and Mineral Nutrition of Field Crops; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Majumdar, A.; Dubey, P.K.; Giri, B.; Moulick, D.; Srivastava, A.K.; Roychowdhury, T.; Bose, S.; Jaiswal, M.K. Combined effects of dry-wet irrigation, redox changes and microbial diversity on soil nutrient bioavailability in the rice field. Soil Till. Res. 2023, 232, 105752. [Google Scholar] [CrossRef]
- Wang, J.; Pan, F.; Soininen, J.; Heino, J.; Shen, J. Nutrient enrichment modifies temperature-biodiversity relationships in large-scale field experiments. Nat. Commun. 2016, 7, 13960. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [CrossRef]
- Heuck, C.; Weig, A.; Spohn, M. Soil microbial biomass C: N: P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Majumdar, A.; Pradhan, N.; Sadasivan, J.; Acharya, A.; Ojha, N.; Babu, S.; Bose, S. Food degradation and foodborne diseases: A microbial approach. In Microbial Contamination and Food Degradation; Elsevier: Amsterdam, The Netherlands, 2018; pp. 109–148. [Google Scholar] [CrossRef]
- Mu, C.; Zhang, T.; Zhang, X.; Li, L.; Guo, H.; Zhao, Q.; Cao, L.; Wu, Q.; Cheng, G. Carbon loss and chemical changes from permafrost collapse in the northern Tibetan Plateau. J. Geophys. Res. Biogeosci. 2016, 121, 1781–1791. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil. Till. Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Mu, C.; Abbott, B.W.; Zhao, Q.; Su, H.; Wang, S.; Wu, Q.; Zhang, T.; Wu, X. Permafrost collapse shifts alpine tundra to a carbon source but reduces N2O and CH4 release on the northern Qinghai-Tibetan Plateau. Geophys. Res. Lett. 2017, 44, 8945–8952. [Google Scholar] [CrossRef]
- Bonde, T.A.; Schnurer, J.; Rosswall, T. Microbial biomass as a faction of potentially mineralizable nitrogen in soils from long-term field experiments. Soil Biol. Biochem. 1988, 20, 447–452. [Google Scholar] [CrossRef]
- Wu, M.; Chen, S.; Chen, J.; Xue, K.; Chen, S.; Wang, X.; Chen, T.; Kang, S.; Rui, J.; Thies, J.E.; et al. Reduced microbial stability in the active layer is associated with carbon loss under alpine permafrost degradation. Proc. Natl. Acad. Sci. USA 2021, 118, e2025321118. [Google Scholar] [CrossRef]
- Dong, X.; Man, H.; Liu, C.; Wu, X.; Zhu, J.; Zheng, Z.; Ma, D.; Li, M.; Zang, S. Changes in soil bacterial community along a gradient of permafrost degradation in Northeast China. Catena 2023, 222, 106870. [Google Scholar] [CrossRef]
- Chapin, F.S.; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science and Business Media: New York, NY, USA, 2011. [Google Scholar] [CrossRef]
- Weedon, J.T.; Kowalchuk, G.A.; Aerts, R.; van Hal, J.; van Logtestijn, R.; Tas, N.; Röling, W.F.M.; van Bodegom, P.M. Summer warming accelerates sub-arctic peatland nitrogen cycling without changing enzyme pools or microbial community structure. Glob. Chang. Biol. 2012, 18, 138–150. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Koven, C.D.; Swenson, S.C.; Riley, W.J.; Slater, A.G. Permafrost thaw and resulting soil moisture changes regulate projected high-latitude CO2 and CH4 emissions. Environ. Res. Lett. 2015, 10, 094011. [Google Scholar] [CrossRef]
- Li, S.; Fang, X.; Xiang, W.; Sun, W.; Zhang, S. Soil microbial biomass carbon and nitrogen concentrations in four subtropical forests in hilly region of central Hunan Province, China. Sci. Silv. Sin. 2014, 50, 8–16. [Google Scholar]
- Kamimura, Y.; Hayano, K. Properties of protease extracted from tea-field soil. Biol. Fert. Soils 2000, 30, 351–355. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S. Size and functional diversity of microbe populations control plant persistence and long-term soil carbon accumulation. Ecol. Lett. 2005, 8, 1075–1087. [Google Scholar] [CrossRef]
- Liu, W.; Jiang, Y.; Su, Y.; Smoak, J.M.; Duan, B. Warming affects soil nitrogen mineralization via changes in root exudation and associated soil microbial communities in a subalpine tree species Abies fabri. J. Soil Sci. Plant Nut. 2022, 22, 406–415. [Google Scholar] [CrossRef]
- Hou, B.; Ma, F.; Wu, H.; Xin, S. Characteristics of soil nematodes communities at different succession stages of wetland in the Yellow River Delta, China, Chin. Chin. J. Appl. Environ. Biol. 2008, 14, 202–206. (In Chinese) [Google Scholar]

| Nmin | SWC | pH | SOC | TN | MBC | MBN | NMDSplant | |
|---|---|---|---|---|---|---|---|---|
| ALT | 18.69 *** | 65.49 *** | 25.87 *** | 214.74 *** | 157.69 *** | 0.76 ns | 14.65 *** | 15.54 *** |
| Soil depth | 30.73 *** | 3.16 * | 1.08 ns | 18.05 ** | 17.85 *** | 4.94 * | 9.74 *** | 0 ns |
| ALT × soil depth | 5.73 ** | 1.38 ns | 2.19 ns | 1.02 ns | 3.6 ns | 0.82 ns | 4.08 ** | 0 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Jin, X.; Yang, S.; Jin, H.; Wang, H.; Li, X.; He, R.; Wang, J.; Sun, Z.; Yun, H. Changes in Soil Substrate and Microbial Properties Associated with Permafrost Thaw Reduce Nitrogen Mineralization. Forests 2023, 14, 2060. https://doi.org/10.3390/f14102060
Yang X, Jin X, Yang S, Jin H, Wang H, Li X, He R, Wang J, Sun Z, Yun H. Changes in Soil Substrate and Microbial Properties Associated with Permafrost Thaw Reduce Nitrogen Mineralization. Forests. 2023; 14(10):2060. https://doi.org/10.3390/f14102060
Chicago/Turabian StyleYang, Xue, Xiaoying Jin, Sizhong Yang, Huijun Jin, Hongwei Wang, Xiaoying Li, Ruixia He, Junfeng Wang, Zhizhong Sun, and Hanbo Yun. 2023. "Changes in Soil Substrate and Microbial Properties Associated with Permafrost Thaw Reduce Nitrogen Mineralization" Forests 14, no. 10: 2060. https://doi.org/10.3390/f14102060
APA StyleYang, X., Jin, X., Yang, S., Jin, H., Wang, H., Li, X., He, R., Wang, J., Sun, Z., & Yun, H. (2023). Changes in Soil Substrate and Microbial Properties Associated with Permafrost Thaw Reduce Nitrogen Mineralization. Forests, 14(10), 2060. https://doi.org/10.3390/f14102060