




Comparison of Chemosensory Receptor Genes in the Antennae Transcriptomes of Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Extraction and cDNA Synthesis
2.3. Sequence and Phylogenetic Analysis of Chemosensory Receptor Genes
2.4. Tissue-and Sex-Specific Expression Profiles of Chemosensory Receptor Genes
3. Results
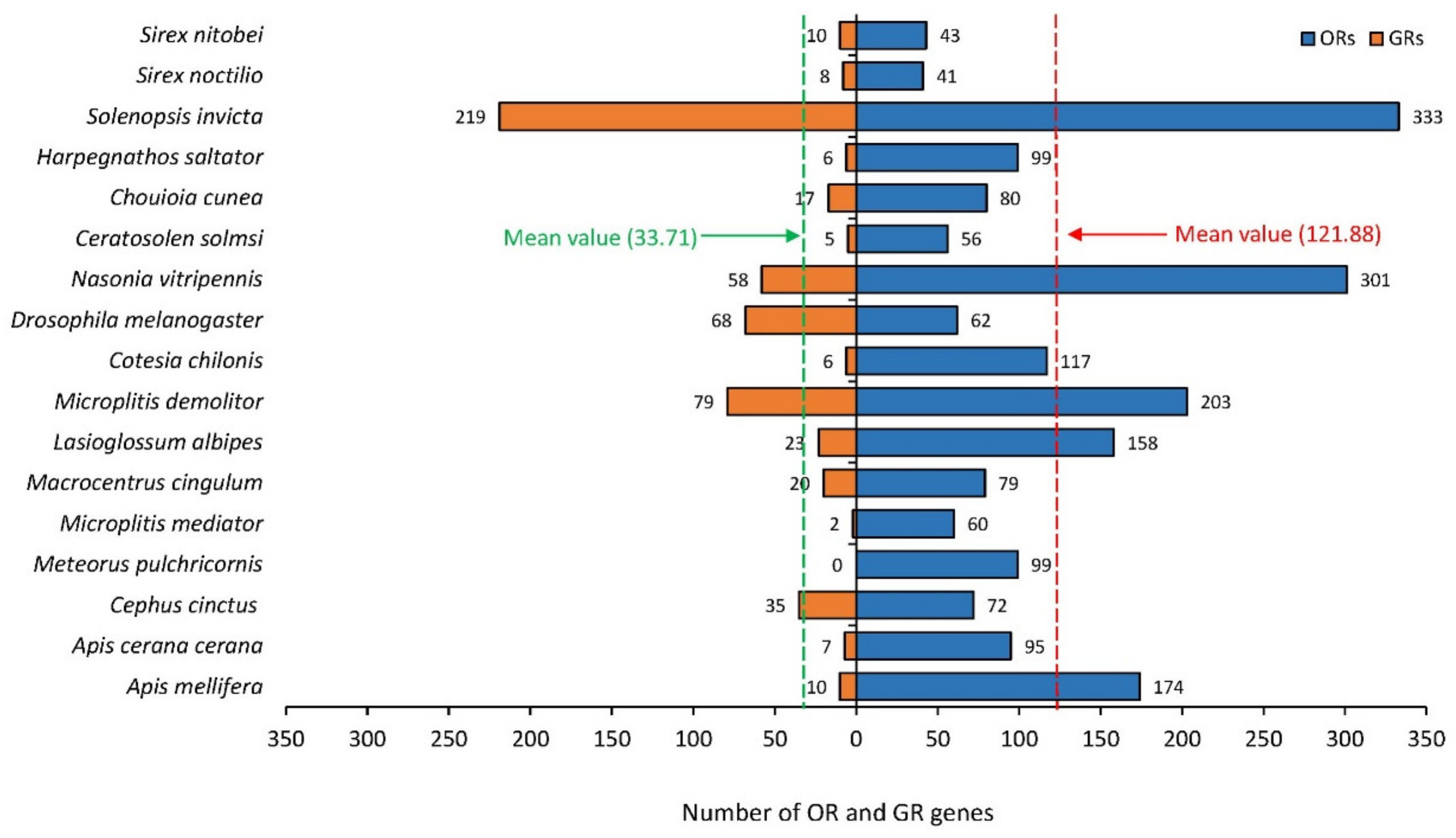
3.1. Analysis of Gene Expression Abundance from Antennal Transcriptome
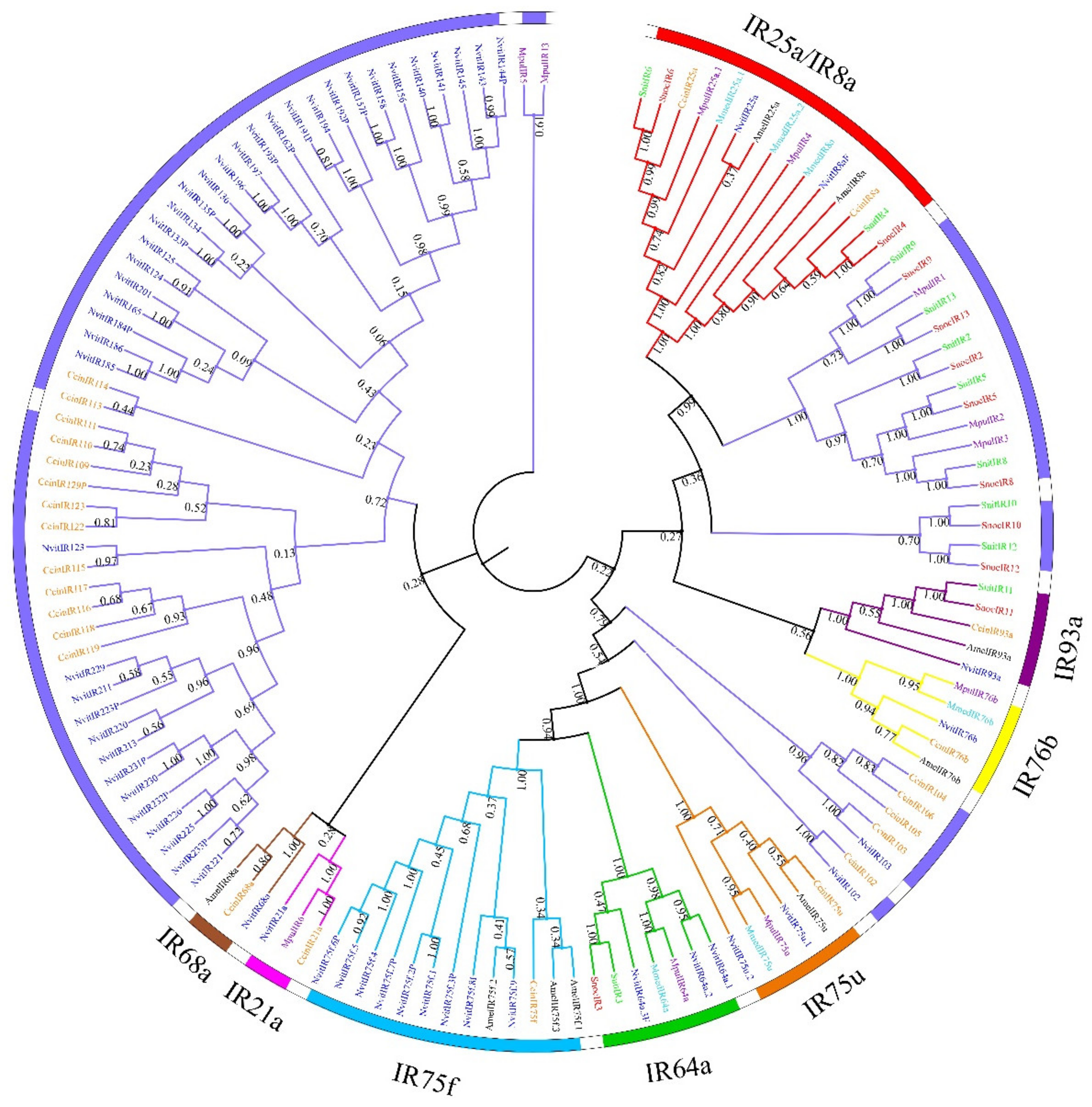
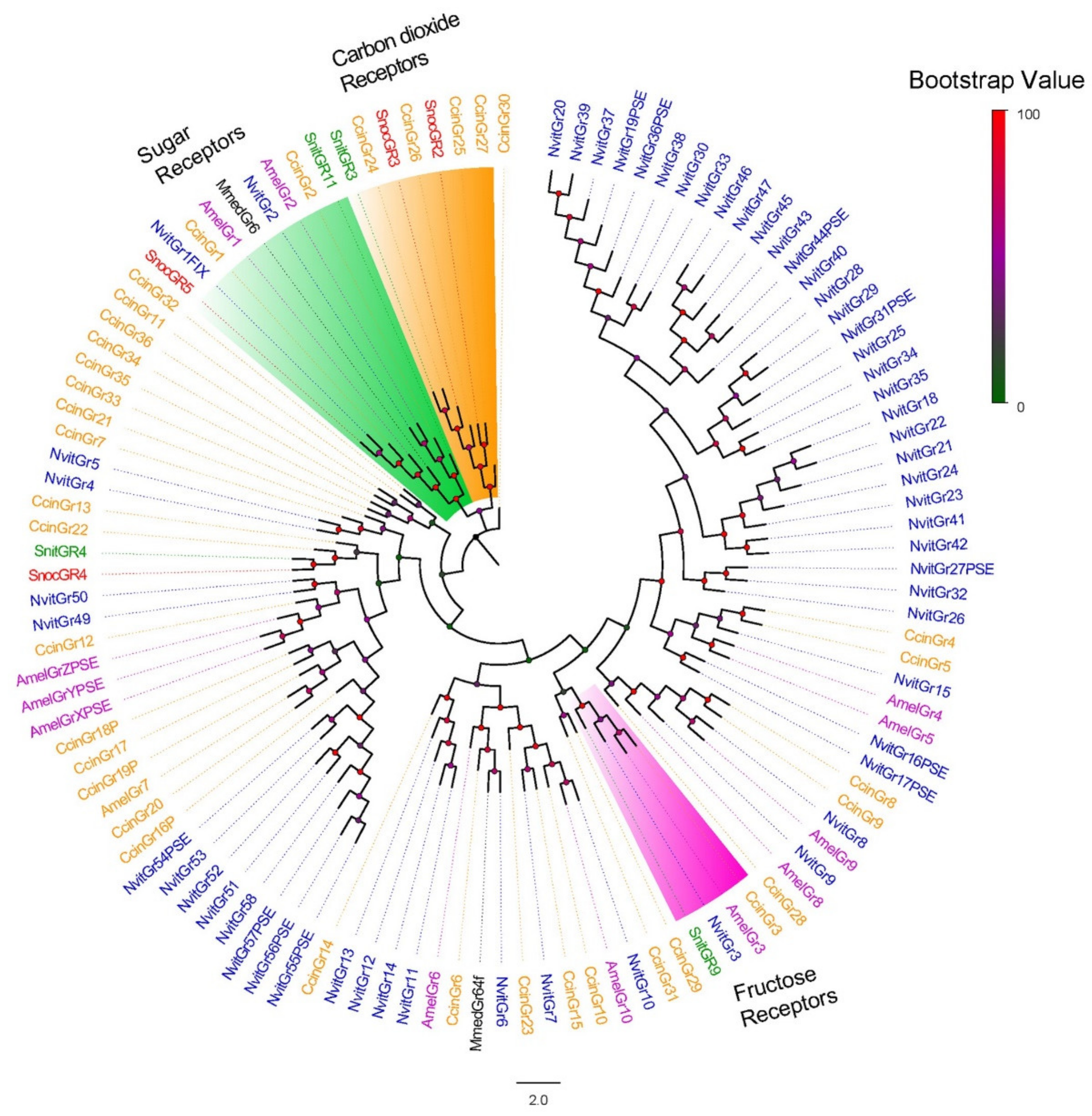
3.2. Phylogenetic Analysis of Chemosensory Receptors of S. noctilio and S. nitobei among Hymenoptera
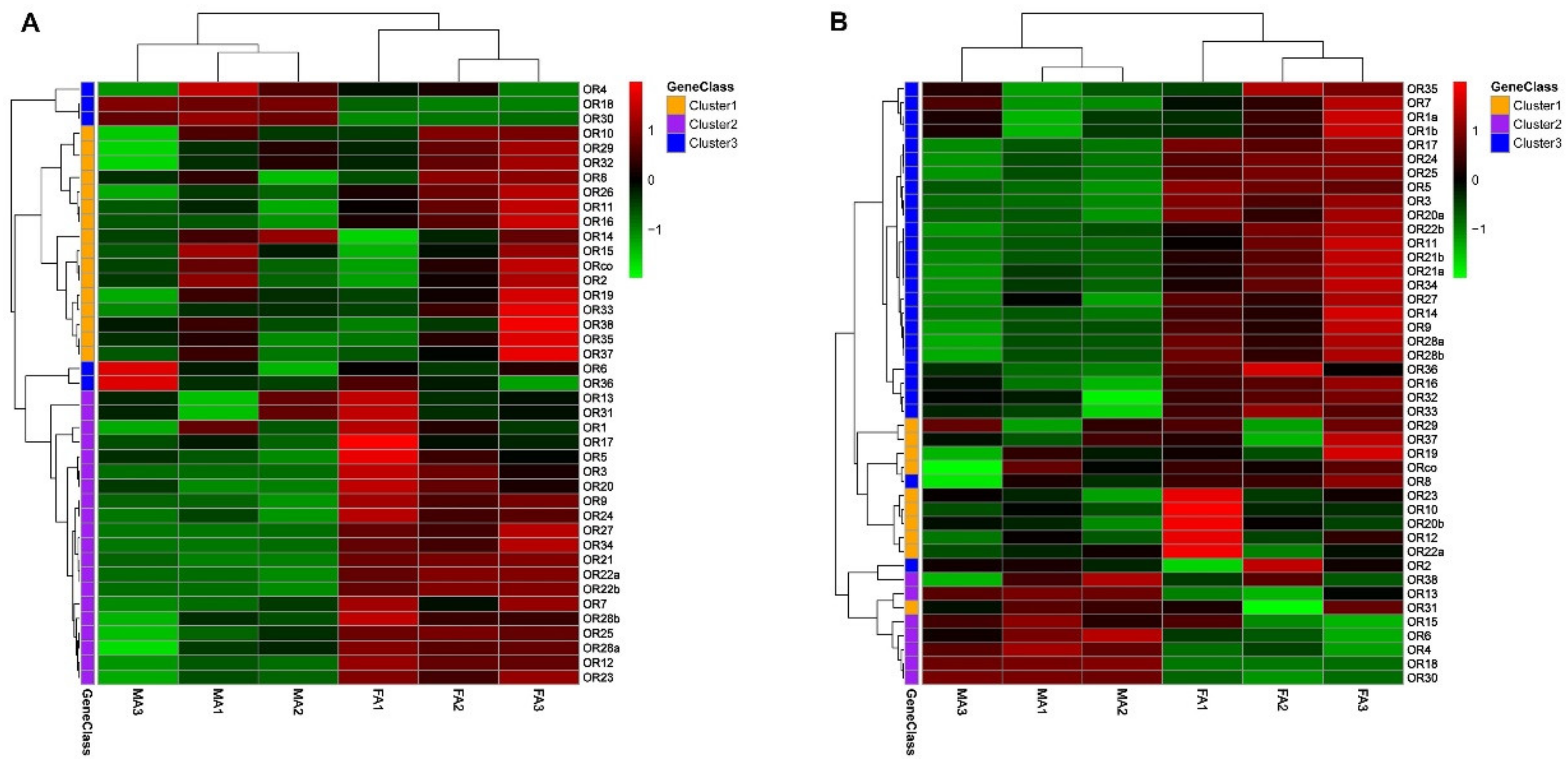
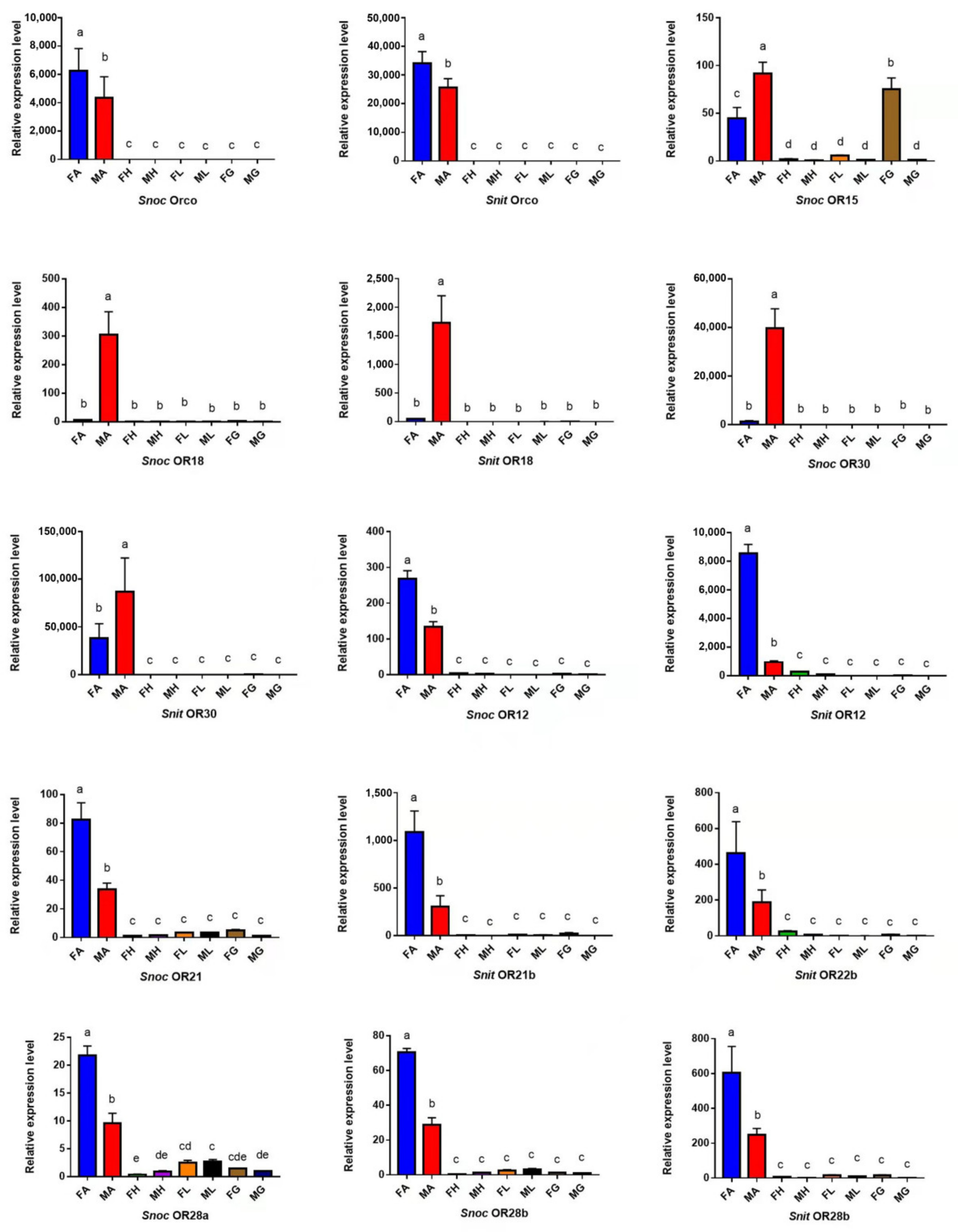
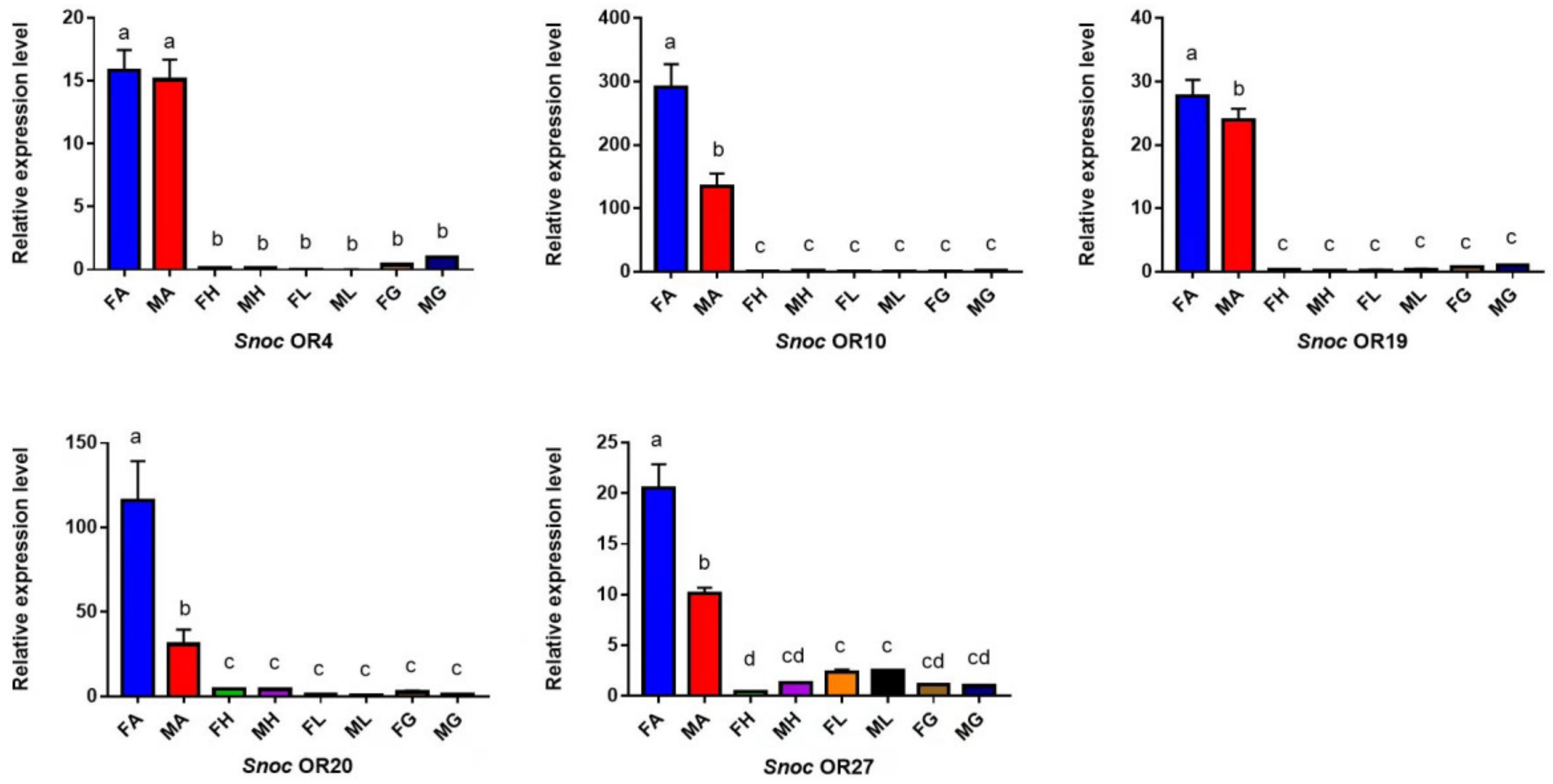
3.3. The Specific Expression Profiles of S. noctilio and S. nitobei OR Genes
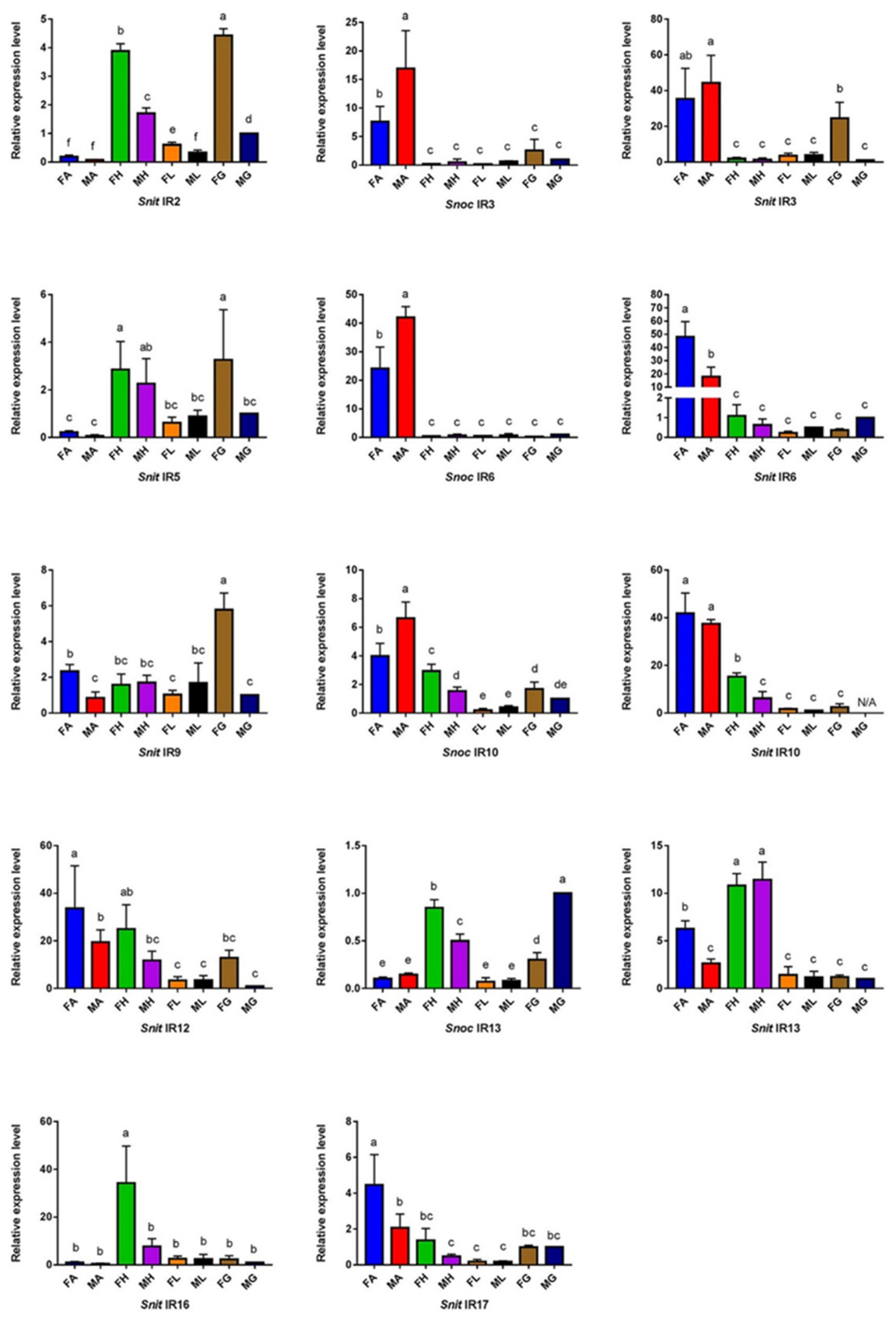
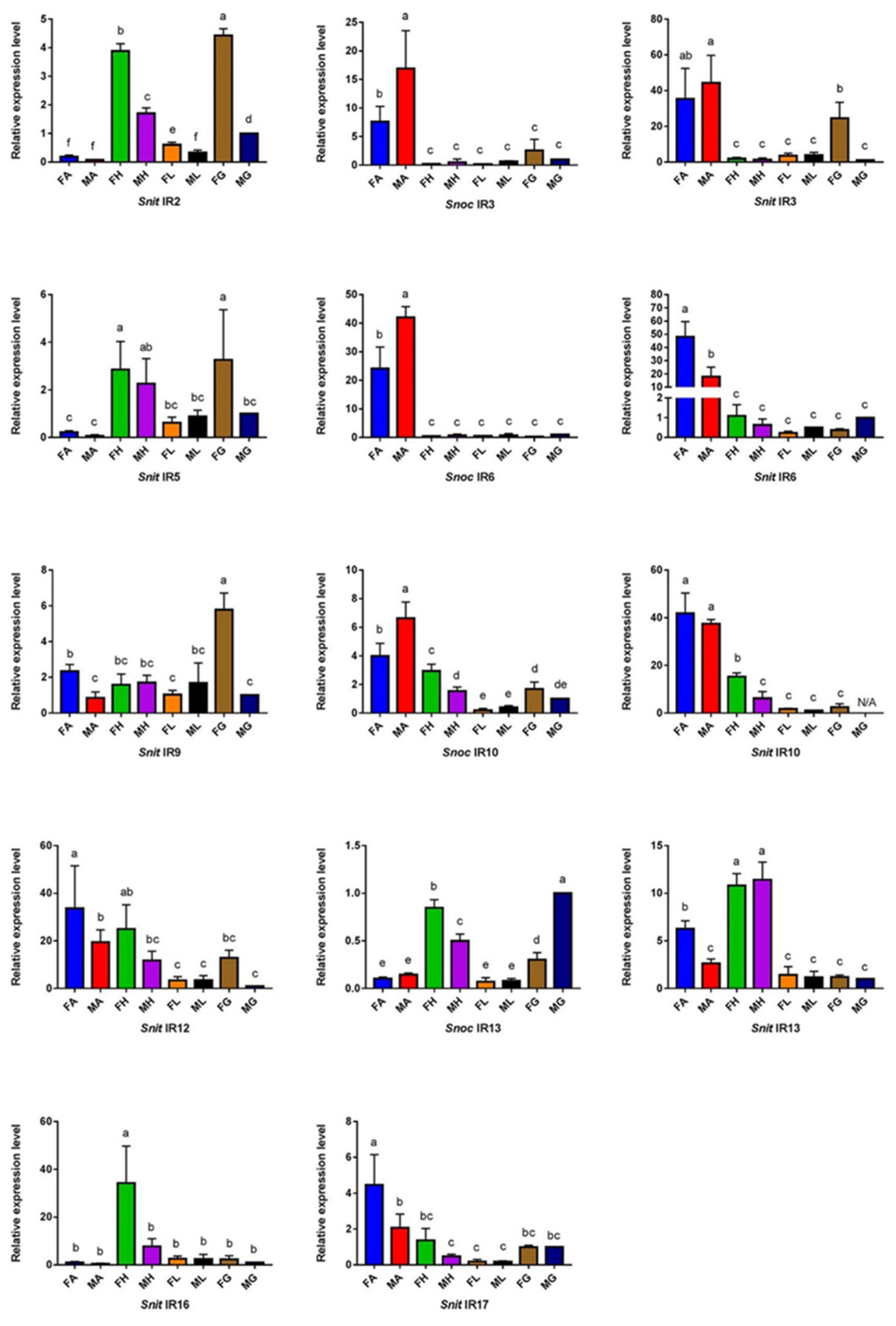
3.4. The Specific Expression Profiles of S. noctilio and S. nitobei IR Genes
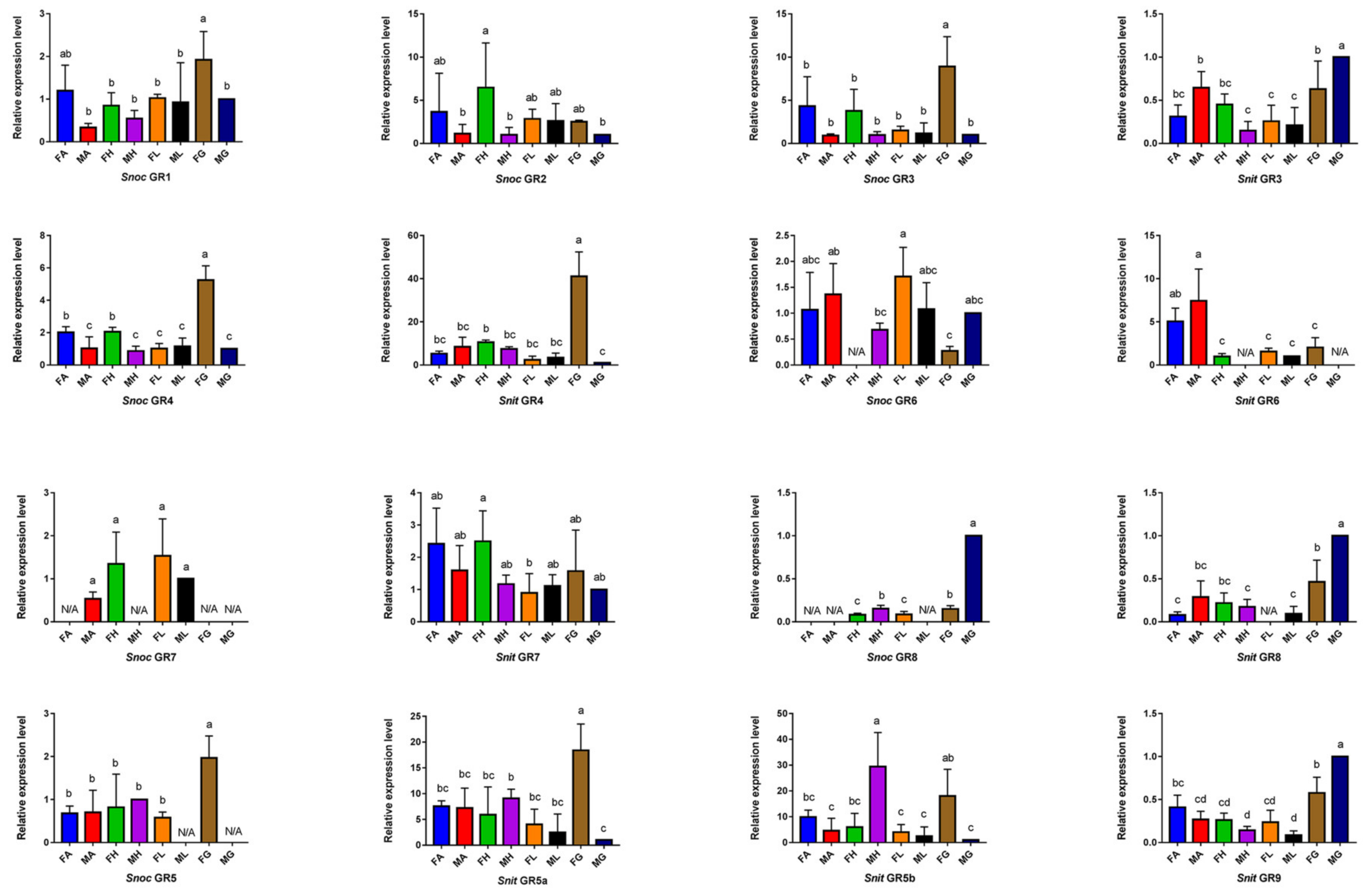
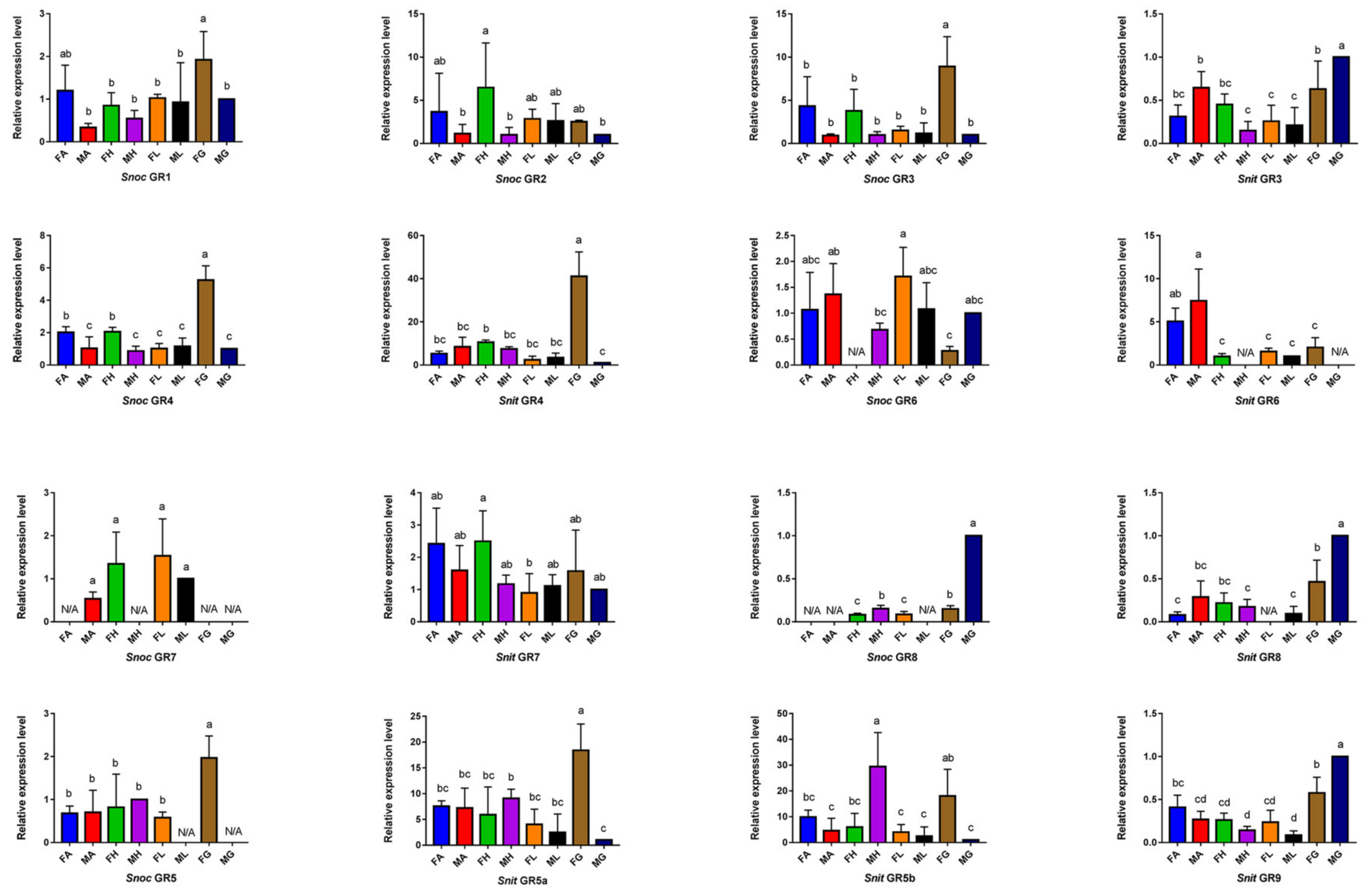
3.5. The Specific Expression Profiles of S. noctilio and S. nitobei GR Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ciesla, W.M. European Woodwasp: A potential threat to North America’s conifer forests. J. For. 2003, 101, 18–23. [Google Scholar]
- Spradbery, J.P.; Kirk, A.A. Aspects of the ecology of siricid woodwasps (Hymenoptera: Siricidae) in Europe, North Africa and Turkey with special reference to the biological control of Sirex noctilio F. in Australia. Bull. Entomol. Res. 1978, 68, 341–359. [Google Scholar] [CrossRef]
- Carnegie, A.J. Predicting the potential distribution of Sirex noctilio (Hymenoptera: Siricidae), a significant exotic pest of Pinus plantations. Ann. For. Sci. 2006, 63, 119–128. [Google Scholar] [CrossRef]
- Haugen, D.; Underdown, M. Release of parasitoids for Sirex noctilio control by transporting infested logs. Austral. For. 1990, 53, 266–270. [Google Scholar] [CrossRef]
- Slippers, B.; Hurley, B.P.; Wingfield, M.J. Sirex Woodwasp: A Model for Evolving Management Paradigms of Invasive Forest Pests. Annu. Rev. Èntomol. 2015, 60, 601–619. [Google Scholar] [CrossRef] [PubMed]
- Haack, R.A. Intercepted Scolytidae (Coleoptera) at U.S. ports of Entry: 1985–2000. Integr. Pest Manag. Rev. 2001, 6, 253–282. [Google Scholar] [CrossRef]
- Miller, D.; Clark, A.F. Sirex noctilio (Hym.) and its Parasite in New Zealand. Bull. Èntomol. Res. 1935, 26, 149–154. [Google Scholar] [CrossRef]
- Gilbert, J.M.; Miller, L.W. An Outbreak of Sirex noctilio F. In Tasmania. Aust. For. 1952, 16, 63–69. [Google Scholar] [CrossRef]
- Hoebeke, E.R.; Haugen, D.A.; Haack, R.A. Sirex noctilo: Discovery of a palearctic siricid woodwasp in New York (revised on 7 July 2005). Newsl. Mich. Entomol. Soc. 2005, 50, 24–25. [Google Scholar]
- Li, D.; Shi, J.; Lu, M.; Ren, L.; Zhen, C.; Luo, Y. Detection and Identification of the Invasive Sirex noctilio (Hymenoptera: Siricidae) Fungal Symbiont, Amylostereum areolatum (Russulales: Amylostereacea), in China and the Stimulating Effect of Insect Venom on Laccase Production by A. areolatum YQL03. J. Econ. Èntomol. 2015, 108, 1136–1147. [Google Scholar] [CrossRef]
- Wang, L.; Ren, L.; Li, C.; Gao, C.; Liu, X.; Wang, M.; Luo, Y. Effects of endophytic fungi diversity in different coniferous species on the colonization of Sirex noctilio (Hymenoptera: Siricidae). Sci. Rep. 2019, 9, 5077. [Google Scholar] [CrossRef]
- Coutts, M. The Mechanism of Pathogenicity of Sirex noctilio on Pinus Radiata II. Effects of S. Noctilio Mucus. Aust. J. Biol. Sci. 1969, 22, 1153–1162. [Google Scholar] [CrossRef]
- Taylor, K.L.; Kitching, R.L.; Jones, R.E. The Sirex woodwasp: Ecology and control of an introduced forest insect. In The Ecology of Pests: Some Australian Case Histories; CSIRO: Canberra, Australia, 1981; pp. 231–248. [Google Scholar]
- Talbot, P.H.B. The Sirex-Amylostereum-Pinus Association. Annu. Rev. Phytopathol. 1977, 15, 41–54. [Google Scholar] [CrossRef]
- Parkin, E.A. Symbiosis and Siricid Woodwasps. Ann. Appl. Biol. 1942, 29, 268–274. [Google Scholar] [CrossRef]
- Thompson, B.M.; Grebenok, R.J.; Behmer, S.T.; Gruner, D.S. Microbial Symbionts Shape the Sterol Profile of the Xylem-Feeding Woodwasp, Sirex noctilio. J. Chem. Ecol. 2013, 39, 129–139. [Google Scholar] [CrossRef]
- Madden, J.; Coutts, M. The role of fungi in the biology and ecology of woodwasps (Hymenoptera: Siricidae). In Insect-Fungus Symbiosis: Nutrition, Mutualism, and Commensalism; Allanheld Osmun: London, UK, 1979; pp. 165–174. [Google Scholar]
- Zondag, R. A nematode infection of Sirex noctilio (F.) in New Zealand. N. Z. J. For. Sci. 1969, 12, 732–747. [Google Scholar]
- Ryan, K.; De Groot, P.; Smith, S.M. Evidence of interaction between Sirex noctilio and other species inhabiting the bole of Pinus. Agric. For. Èntomol. 2012, 14, 187–195. [Google Scholar] [CrossRef]
- Hurley, B.P.; Slippers, B.; Wingfield, M.J. A comparison of control results for the alien invasive woodwasp, Sirex noctilio, in the southern hemisphere. Agric. For. Èntomol. 2007, 9, 159–171. [Google Scholar] [CrossRef]
- Zylstra, K.E.; Dodds, K.J.; Francese, J.A.; Mastro, V. Sirex noctilio in North America: The effect of stem-injection timing on the at-tractiveness and suitability of trap trees. Agric. For. Entomol. 2010, 12, 243–250. [Google Scholar]
- Bashford, R. The development of static trapping systems to monitor for wood-boring insects in forestry plantations. Aust. For. 2008, 71, 236–241. [Google Scholar] [CrossRef]
- Cooperband, M.F.; Böröczky, K.; Hartness, A.; Jones, T.H.; Zylstra, K.E.; Tumlinson, J.H.; Mastro, V.C. Male-Produced Pheromone in the European Woodwasp, Sirex noctilio. J. Chem. Ecol. 2012, 38, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Èntomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Tsitoura, P.; Andronopoulou, E.; Tsikou, D.; Agalou, A.; Papakonstantinou, M.P.; Kotzia, G.A.; Labropoulou, V.; Swevers, L.; Georgoussi, Z.; Iatrou, K. Expression and Membrane Topology of Anopheles gambiae Odorant Receptors in Lepidopteran Insect Cells. PLoS ONE 2010, 5, e15428. [Google Scholar] [CrossRef]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Structure-Function of the G Protein–Coupled Receptor Superfamily. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 531–556. [Google Scholar] [CrossRef]
- Neuhaus, E.M.; Gisselmann, G.; Zhang, W.; Dooley, R.; Störtkuhl, K.; Hatt, H. Odorant receptor heterodimerization in the olfactory system of Drosophila melanogaster. Nat. Neurosci. 2005, 8, 15–17. [Google Scholar] [CrossRef]
- Nakagawa, T.; Sakurai, T.; Nishioka, T.; Touhara, K. Insect Sex-Pheromone Signals Mediated by Specific Combinations of Olfactory Receptors. Science 2005, 307, 1638–1642. [Google Scholar] [CrossRef]
- Di, C.; Ning, C.; Huang, L.Q.; Wang, C.Z. Design of larval chemical attractants based on odorant response spectra of odorant re-ceptors in the cotton bollworm. Insect. Biochem. Mol. Biol. 2017, 84, 48–62. [Google Scholar] [CrossRef]
- Yang, K.; Huang, L.-Q.; Ning, C.; Wang, C.-Z. Two single-point mutations shift the ligand selectivity of a pheromone receptor between two closely related moth species. eLife 2017, 6, e29100. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Hansson, B.S. A Unified Nomenclature System for the Insect Olfactory Coreceptor. Chem. Sens. 2011, 36, 497–498. [Google Scholar] [CrossRef]
- Jones, W.; Nguyen, T.-A.; Kloss, B.; Lee, K.; Vosshall, L. Functional conservation of an insect odorant receptor gene across 250 million years of evolution. Curr. Biol. 2005, 15, R119–R121. [Google Scholar] [CrossRef]
- Gu, S.-H.; Zhou, J.-J.; Gao, S.; Wang, D.-H.; Li, X.-C.; Guo, Y.-Y.; Zhang, Y.-J. Identification and comparative expression analysis of odorant binding protein genes in the tobacco cutworm Spodoptera litura. Sci. Rep. 2015, 5, 13800. [Google Scholar] [CrossRef]
- Cattaneo, A.M.; Gonzalez, F.; Bengtsson, J.M.; Corey, E.A.; Jacquin-Joly, E.; Montagné, N.; Salvagnin, U.; Walker, W.B.; Witzgall, P.; Anfora, G. Candidate pheromone receptors of codling moth Cydia pomonella respond to pheromones and kairomones. Sci. Rep. 2017, 7, 41105. [Google Scholar] [CrossRef] [Green Version]
- Clyne, P.J.; Warr, C.G.; Carlson, J.R. Candidate Taste Receptors in Drosophila. Science 2000, 287, 1830–1834. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A Spatial Map of Olfactory Receptor Expression in the Drosophila Antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Wanner, K.W.; Nichols, A.S.; Walden, K.K.O.; Brockmann, A.; Luetje, C.W.; Robertson, H.M. A honey bee odorant receptor for the queen substance 9-oxo-2-decenoic acid. Proc. Natl. Acad. Sci. USA 2007, 104, 14383–14388. [Google Scholar] [CrossRef]
- Hallem, E.A.; Dahanukar, A.; Carlson, J.R. Insect odor and taste receptors. Annu. Rev. Èntomol. 2006, 51, 113–135. [Google Scholar] [CrossRef]
- Silbering, A.F.; Rytz, R.; Grosjean, Y.; Abuin, L.; Ramdya, P.; Jefferis, G.; Benton, R. Complementary Function and Integrated Wiring of the Evolutionarily Distinct Drosophila Olfactory Subsystems. J. Neurosci. 2011, 31, 13357–13375. [Google Scholar] [CrossRef]
- Guo, M.; Krieger, J.; Große-Wilde, E.; Mißbach, C.; Zhang, L.; Breer, H. Variant Ionotropic Receptors Are Expressed in Olfactory Sensory Neurons of Coeloconic Sensilla on the Antenna of the Desert Locust (Schistocerca gregaria). Int. J. Biol. Sci. 2014, 10, 1–14. [Google Scholar] [CrossRef]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic Receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient Protostome Origin of Chemosensory Ionotropic Glutamate Receptors and the Evolution of Insect Taste and Olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef] [PubMed]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant Ionotropic Glutamate Receptors as Chemosensory Receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.V.; Ni, J.; Montell, C. The Molecular Basis for Attractive Salt-Taste Coding in Drosophila. Science 2013, 340, 1334–1338. [Google Scholar] [CrossRef] [Green Version]
- Crava, C.M.; Ramasamy, S.; Ometto, L.; Anfora, G.; Rota-Stabelli, O. Evolutionary Insights into Taste Perception of the Invasive Pest Drosophila suzukii. G3 Genes Genomes Genet. 2016, 6, 4185–4196. [Google Scholar] [CrossRef]
- Scott, K.; Brady, R.; Cravchik, A.; Morozov, P.; Rzhetsky, A.; Zuker, C.; Axel, R. A chemosensory gene family encoding candidate gustatory and olfactory receptor in Drosophila. Cell 2001, 104, 661–673. [Google Scholar] [CrossRef]
- Robertson, H.M.; Wanner, K.W. The chemoreceptor superfamily in the honey bee, Apis mellifera: Expansion of the odorant, but not gustatory, receptor family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef]
- Chyb, S.; Dahanukar, A.; Wickens, A.; Carlson, J.R. Drosophila Gr5a encodes a taste receptor tuned to trehalose. Proc. Natl. Acad. Sci. USA 2003, 100, 14526–14530. [Google Scholar] [CrossRef]
- Dahanukar, A.; Lei, Y.-T.; Kwon, J.Y.; Carlson, J.R. Two Gr Genes Underlie Sugar Reception in Drosophila. Neuron 2007, 56, 503–516. [Google Scholar] [CrossRef]
- Jiao, Y.; Moon, S.J.; Wang, X.; Ren, Q.; Montell, C. Gr64f Is Required in Combination with Other Gustatory Receptors for Sugar Detection in Drosophila. Curr. Biol. 2008, 18, 1797–1801. [Google Scholar] [CrossRef]
- Thorne, N.; Chromey, C.; Bray, S.; Amrein, H. Taste Perception and Coding in Drosophila. Curr. Biol. 2004, 14, 1065–1079. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Singhvi, A.; Kong, P.; Scott, K. Taste Representations in the Drosophila Brain. Cell 2004, 117, 981–991. [Google Scholar] [CrossRef]
- Jones, W.; Cayirlioglu, P.; Kadow, I.G.; Vosshall, L. Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature 2007, 445, 86–90. [Google Scholar] [CrossRef]
- Montell, C. A taste of the Drosophila gustatory receptors. Curr. Opin. Neurobiol. 2009, 19, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Stocker, R.F. The organization of the chemosensory system in Drosophila melanogaster: A rewiew. Cell Tissue Res. 1994, 275, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Hao, E.H.; Qiao, H.L.; Wang, J.Z.; Wu, W.W.; Zhou, J.J.; Lu, P.F. Antennal transcriptome analysis of olfactory genes and char-acterizations of odorant binding proteins in two woodwasps, Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). BMC Genom. 2021, 22, 172. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Wilkinson, L.; Friendly, M. The History of the Cluster Heat Map. Am. Stat. 2009, 63, 179–184. [Google Scholar] [CrossRef]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sheng, S.; Liao, C.-W.; Zheng, Y.; Zhou, Y.; Xu, Y.; Song, W.-M.; He, P.; Zhang, J.; Wu, F.-A. Candidate chemosensory genes identified in the endoparasitoid Meteorus pulchricornis (Hymenoptera: Braconidae) by antennal transcriptome analysis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 20–31. [Google Scholar] [CrossRef]
- Werren, J.H.; Richards, S.; Desjardins, C.A.; Niehuis, O.; Gadau, J.; Colbourne, J.K.; The Nasonia Genome Working Group. Functional and Evolutionary Insights from the Genomes of Three Parasitoid Nasonia Species. Science 2010, 327, 343–348. [Google Scholar] [CrossRef]
- Ahmed, T.; Zhang, T.-T.; Wang, Z.-Y.; He, K.-L.; Bai, S.-X. Identification and expression pattern analysis of chemosensory receptor genes in the Macrocentrus cingulum (Hymenoptera: Braconidae) antennae. Eur. J. Èntomol. 2016, 113, 76–83. [Google Scholar] [CrossRef]
- Wang, S.-N.; Peng, Y.; Lu, Z.-Y.; Dhiloo, K.H.; Gu, S.-H.; Li, R.-J.; Zhou, J.-J.; Zhang, Y.-J.; Guo, Y.-Y. Identification and Expression Analysis of Putative Chemosensory Receptor Genes in Microplitis mediator by Antennal Transcriptome Screening. Int. J. Biol. Sci. 2015, 11, 737–751. [Google Scholar] [CrossRef]
- Sun, Y.L.; Dong, J.F.; Ning, C.; Ding, P.P.; Huang, L.Q.; Sun, J.G.; Wang, C.Z. An odorant receptor mediates the attractiveness of cis-jasmone to Campoletis chlorideae, the endoparasitoid of Helicoverpa armigera. Insect Mol. Biol. 2019, 28, 23–34. [Google Scholar] [CrossRef]
- Robertson, H.M.; Waterhouse, R.M.; Walden, K.K.O.; Ruzzante, L.; Reijnders, M.J.M.F.; Coates, B.S.; Legeai, F.; Gress, J.C.; Biyiklioglu, S.; Weaver, D.K.; et al. Genome sequence of the wheat stem sawfly, Cephus cinctus, representing an early-branching lineage of the Hymenoptera, illuminates evolution of hymenopteran chemoreceptors. Genome Biol. Evol. 2018, 10, 2997–3011. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, F.; Zhang, X.; Zhang, S.; Guo, S.; Zhu, G.; Qiang, L.; Min, L. Transcriptome and expression patterns of chemosensory genes in antennae of the parasitoid wasp Chouioia cunea. PLoS ONE 2016, 11, e0148159. [Google Scholar] [CrossRef]
- Robertson, H.M.; Gadau, J.; Wanner, K.W. The insect chemoreceptor superfamily of the parasitoid jewel wasp Nasonia vitripennis. Insect Mol. Biol. 2010, 19, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Zhang, T.; Wang, Z.; He, K.; Bai, S. Gene set of chemosensory receptors in the polyembryonic endoparasitoid Macro-centrus cingulum. Sci. Rep. 2016, 6, 24078. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, J.; Cui, M.; Tao, J.; Luo, Y. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 2016, 6, 26652. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.-Z.; Min, S.-F.; Mi, F.; Zhou, S.-S.; Wang, M.-Q. Analysis of chemosensory gene families in the beetle Monochamus alternatus and its parasitoid Dastarcus helophoroides. Comp. Biochem. Physiol. Part D Genom. Proteom. 2014, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mamidala, P.; Wijeratne, A.J.; Wijeratne, S.; Poland, T.; Qazi, S.S.; Doucet, D.; Cusson, M.; Beliveau, C.; Mittapalli, O. Identification of Odor-Processing Genes in the Emerald Ash Borer, Agrilus planipennis. PLoS ONE 2013, 8, e56555. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Huertas, B.; Mitchell, R.F.; Lapoint, R.T.; Faucher, C.P.; Hildebrand, J.G.; Whiteman, N.K. Evolution of herbivory in Drosophilidae linked to loss of behaviors, antennal responses, odorant receptors, and ancestral diet. Proc. Natl. Acad. Sci. USA 2015, 112, 3026–3031. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, J.; Liu, N.-Y.; Zhang, Y.-N.; Yang, K.; Dong, S.-L. Distinct Expression Profiles and Different Functions of Odorant Binding Proteins in Nilaparvata lugens Stål. PLoS ONE 2011, 6, e28921. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-J.; Vieira, F.G.; He, X.-L.; Smadja, C.; Liu, R.; Rozas, J.; Field, L.M. Genome annotation and comparative analyses of the odorant-binding proteins and chemosensory proteins in the pea aphid Acyrthosiphon pisum. Insect Mol. Biol. 2010, 19, 113–122. [Google Scholar] [CrossRef]
- Shanbhag, S.R.; Singh, K.; Singh, R.N. Fine structure and primary sensory projections of sensilla located in the sacculus of the antenna of Drosophila melanogaster. Cell Tissue Res. 2010, 282, 237–249. [Google Scholar] [CrossRef]
- Ai, M.; Min, S.; Grosjean, Y.; Leblanc, C.; Bell, R.; Benton, R.; Suh, G.S.B. Acid sensing by the Drosophila olfactory system. Nature 2010, 468, 691–695. [Google Scholar] [CrossRef]
- Wang, S.-N.; Peng, Y.; Lu, Z.-Y.; Dhiloo, K.H.; Zheng, Y.; Shan, S.; Li, R.-J.; Zhang, Y.-J.; Guo, Y.-Y. Cloning and expression profile of ionotropic receptors in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). J. Insect Physiol. 2016, 90, 27–35. [Google Scholar] [CrossRef]
- Gallio, M.; Ofstad, T.A.; Macpherson, L.J.; Wang, J.W.; Zuker, C.S. The Coding of Temperature in the Drosophila Brain. Cell 2011, 144, 614–624. [Google Scholar] [CrossRef]
- Chauhan, P.; Hansson, B.; Kraaijeveld, K.; De Knijff, P.; Svensson, E.I.; Wellenreuther, M. De novo transcriptome of Ischnura elegans provides insights into sensory biology, colour and vision genes. BMC Genome 2014, 15, 808. [Google Scholar] [CrossRef] [PubMed]
- Barbarossa, I.T.; Muroni, P.; Dardani, M.; Casula, P.; Angioy, A.M. New insight into the antennal chemosensory function of Opius concolor (Hymenoptera: Braconidae). It. J. Zool. 1998, 65, 367–370. [Google Scholar] [CrossRef]
- De Brito Sanchez, M.G. Taste perception in honey bees. Chem. Sens. 2011, 36, 675–692. [Google Scholar] [CrossRef]
- Yang, C.-H.; Belawat, P.; Hafen, E.; Jan, L.Y.; Jan, Y.-N. Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes. Science 2008, 319, 1679–1683. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M.; Warr, C.G.; Carlson, J.R. Molecular evolution of the insect chemoreceptor gene superfamily in Drosophila mela-nogaster. Proc. Natl. Acad. Sci. USA 2003, 100, 14537–14542. [Google Scholar] [CrossRef] [PubMed]
- De Brito Sanchez, M.G.; Giurfa, M.; de Paula Mota, T.R.; Gauthier, M. Electrophysiological and behavioural characterization of gustatory responses to antennal ‘bitter’ taste in honeybees. Eur. J. Neurosci. 2005, 22, 3161–3170. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Hao, E.; Guo, B.; Yang, H.; Zhou, J.; Ma, M.; Lu, P.; Qiao, H. Comparison of Chemosensory Receptor Genes in the Antennae Transcriptomes of Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). Forests 2022, 13, 1495. https://doi.org/10.3390/f13091495
Wu W, Hao E, Guo B, Yang H, Zhou J, Ma M, Lu P, Qiao H. Comparison of Chemosensory Receptor Genes in the Antennae Transcriptomes of Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). Forests. 2022; 13(9):1495. https://doi.org/10.3390/f13091495
Chicago/Turabian StyleWu, Weiwei, Enhua Hao, Bing Guo, Huan Yang, Jingjiang Zhou, Mei Ma, Pengfei Lu, and Haili Qiao. 2022. "Comparison of Chemosensory Receptor Genes in the Antennae Transcriptomes of Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae)" Forests 13, no. 9: 1495. https://doi.org/10.3390/f13091495
APA StyleWu, W., Hao, E., Guo, B., Yang, H., Zhou, J., Ma, M., Lu, P., & Qiao, H. (2022). Comparison of Chemosensory Receptor Genes in the Antennae Transcriptomes of Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). Forests, 13(9), 1495. https://doi.org/10.3390/f13091495