
Picea wilsonii Transcription Factor PwNAC38 Promotes Drought and Salt Tolerance in Transgenic Arabidopsis and Apple Calli


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA Extraction and Gene Expression Analysis
2.2. In Silico Analysis
2.3. Vector Construction and Acquisition of Overexpression and Mutant Lines
2.4. Transcriptional Activation and Y2H Assays
2.5. Subcellular Localization
2.6. Salt and Drought Treatments
2.7. Biochemical Analysis
2.8. ABA Treatment
2.9. Measurement of Fresh Weight for Transgenic Apple Calli
2.10. Statistical Analysis
3. Results
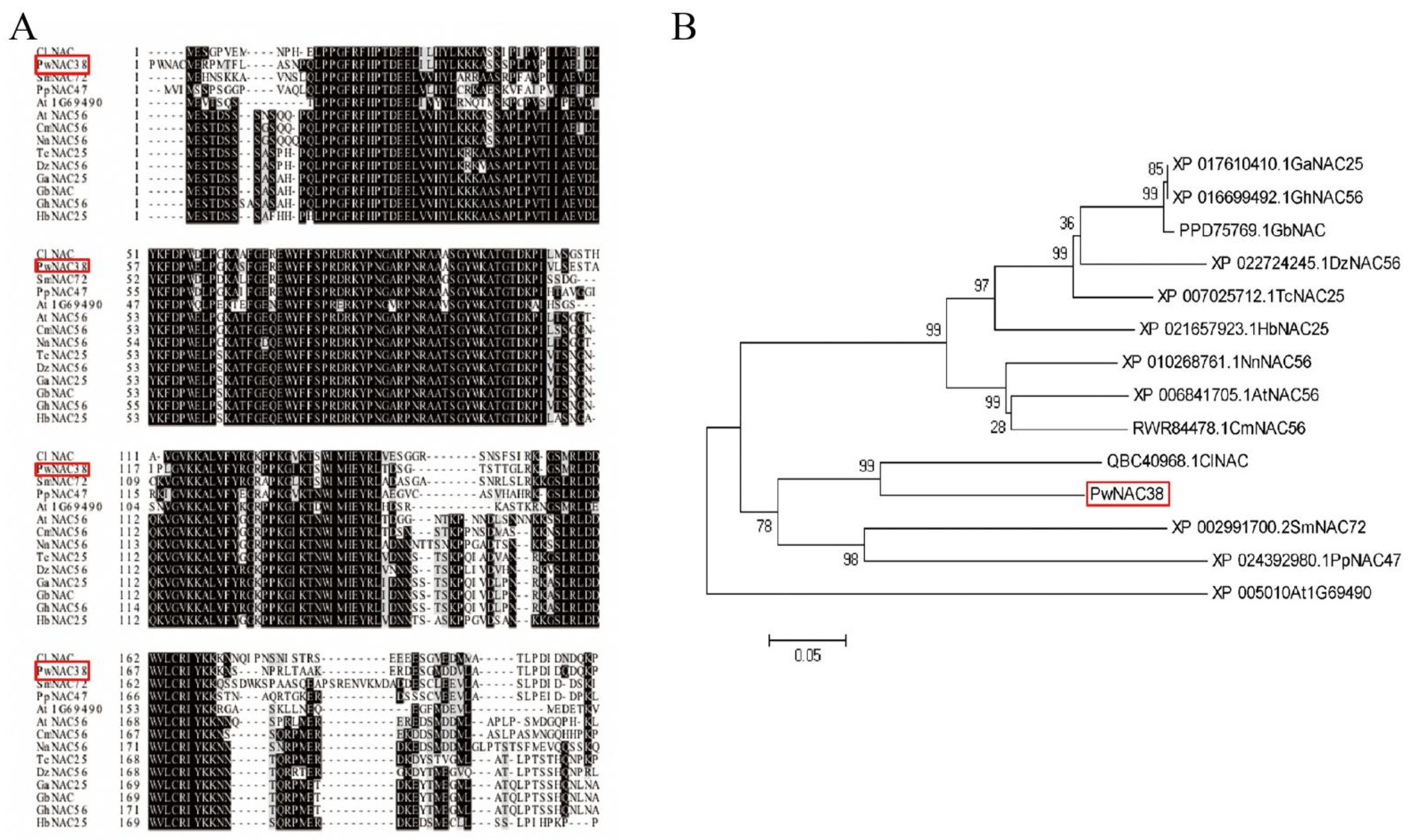
3.1. Analysis of PwNAC38 Gene
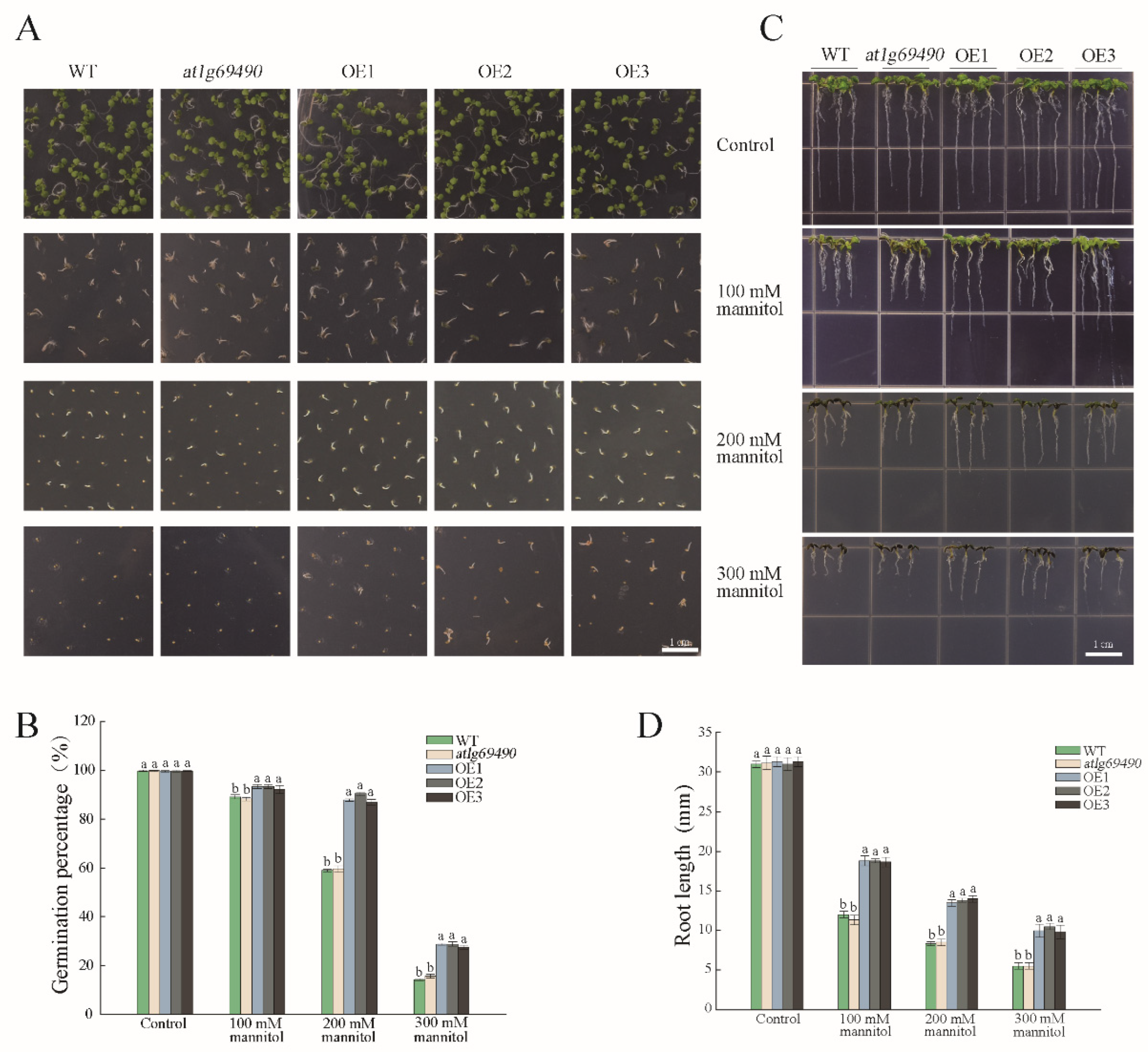
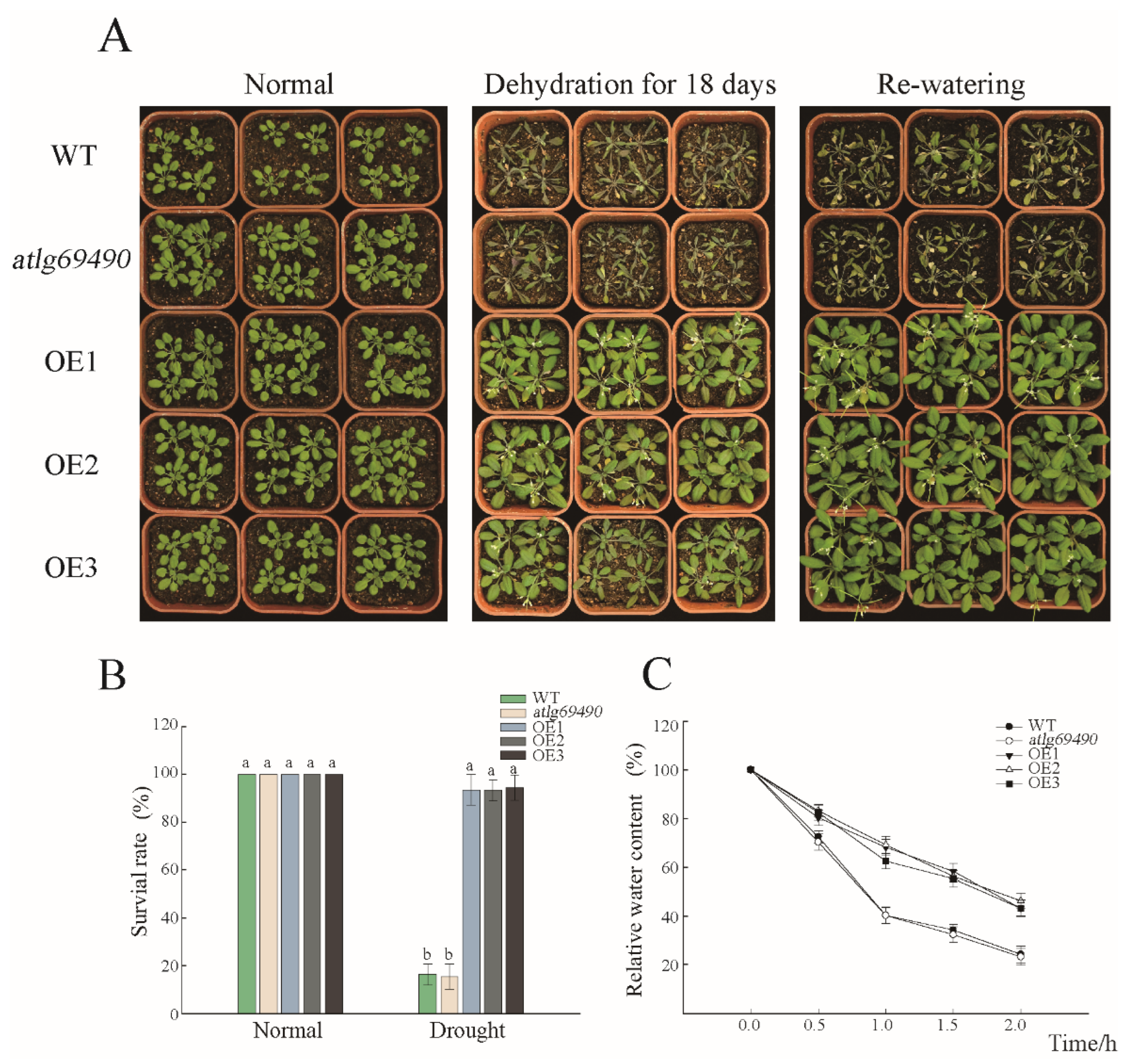
3.2. Transgenic Arabidopsis and Drought Phenotype Experiments
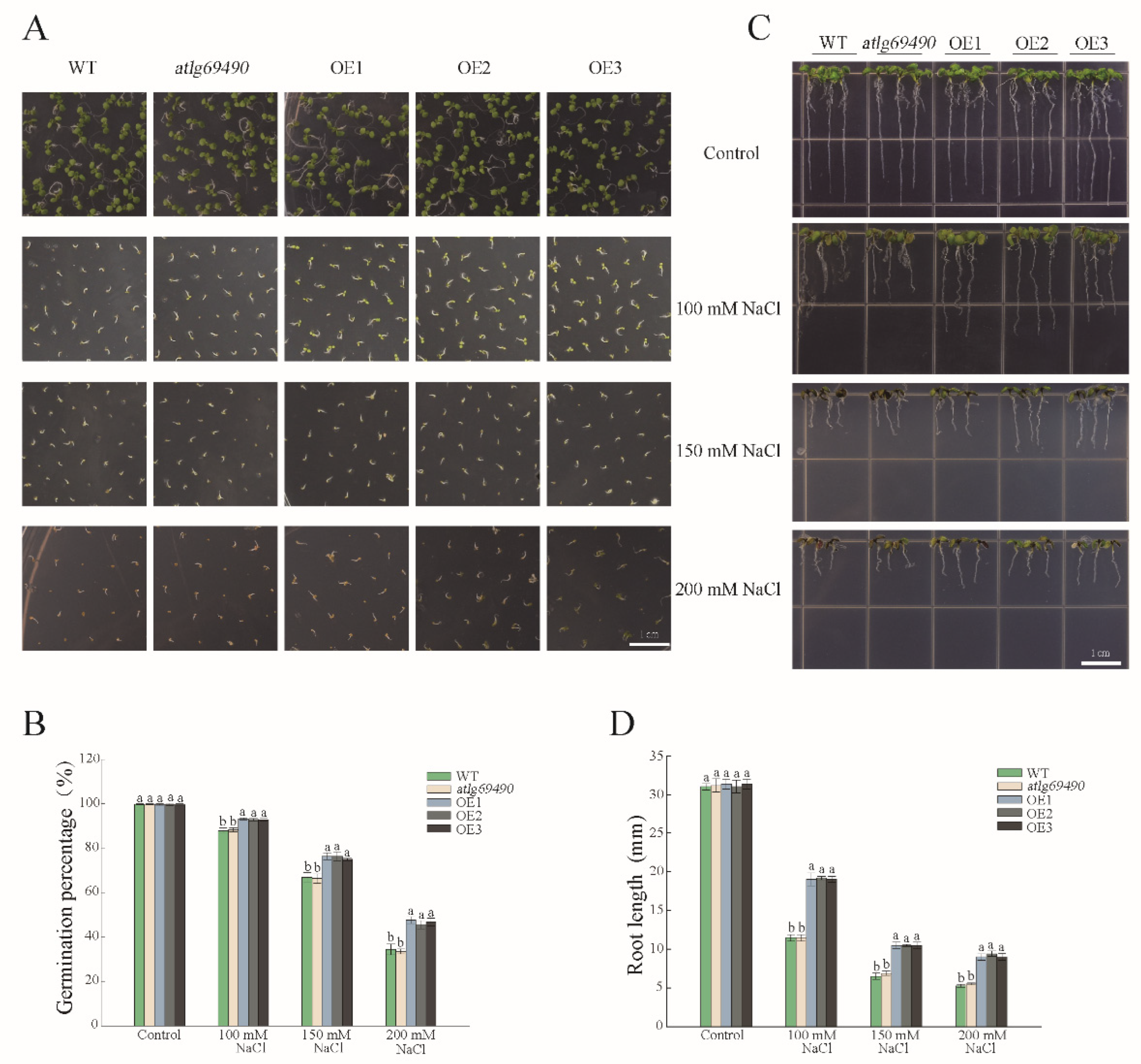
3.3. PwNAC38 Positively Regulated Plant Salt Tolerance
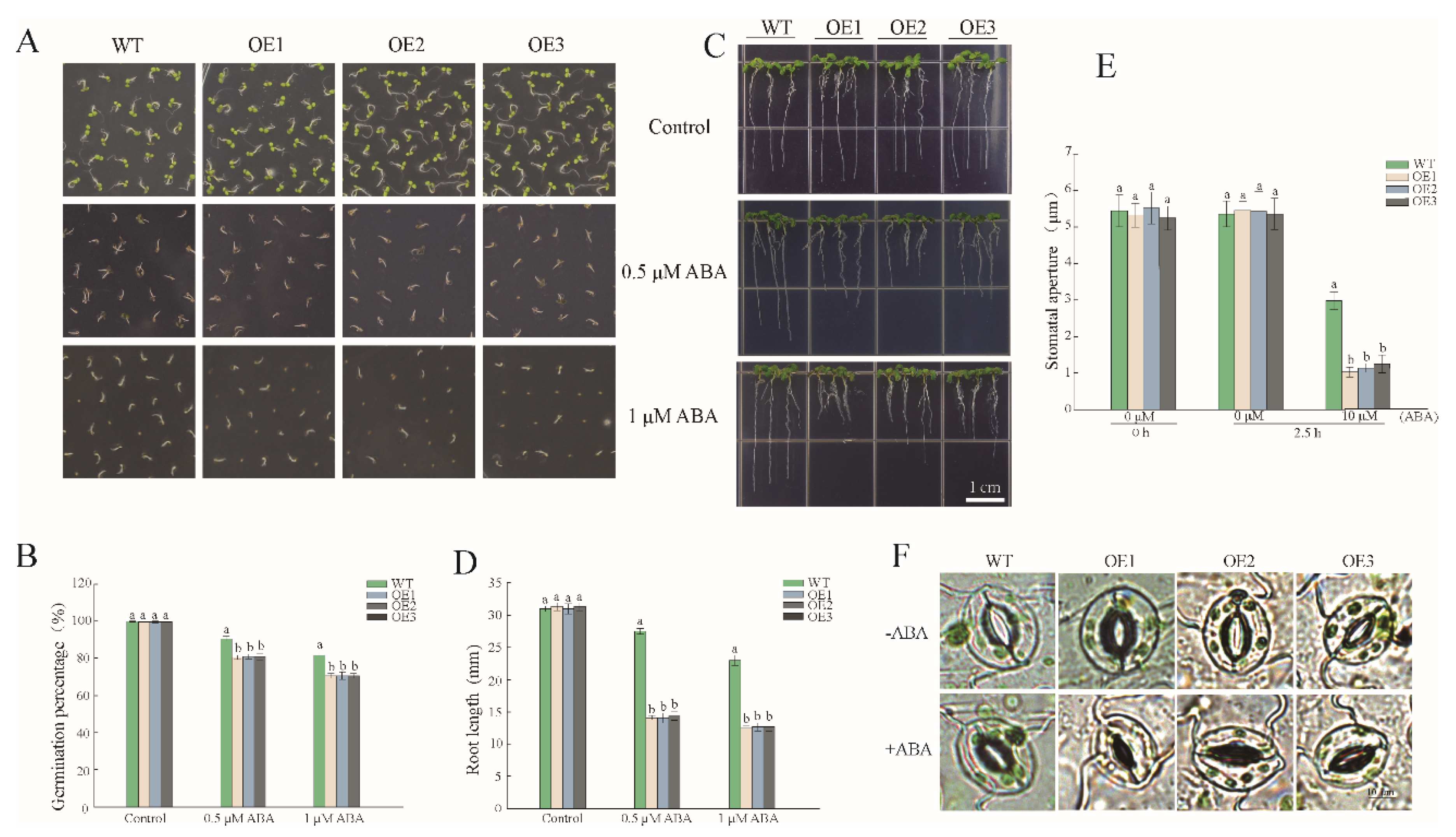
3.4. Overexpression of PwNAC38 Caused Susceptibility to ABA
3.5. ROS Scavenging Efficiency Was Enhanced in Transgenic Arabidopsis
3.6. Overexpression of PwNAC38 Improved Drought and Salt Tolerance of Transgenic Apple Calli
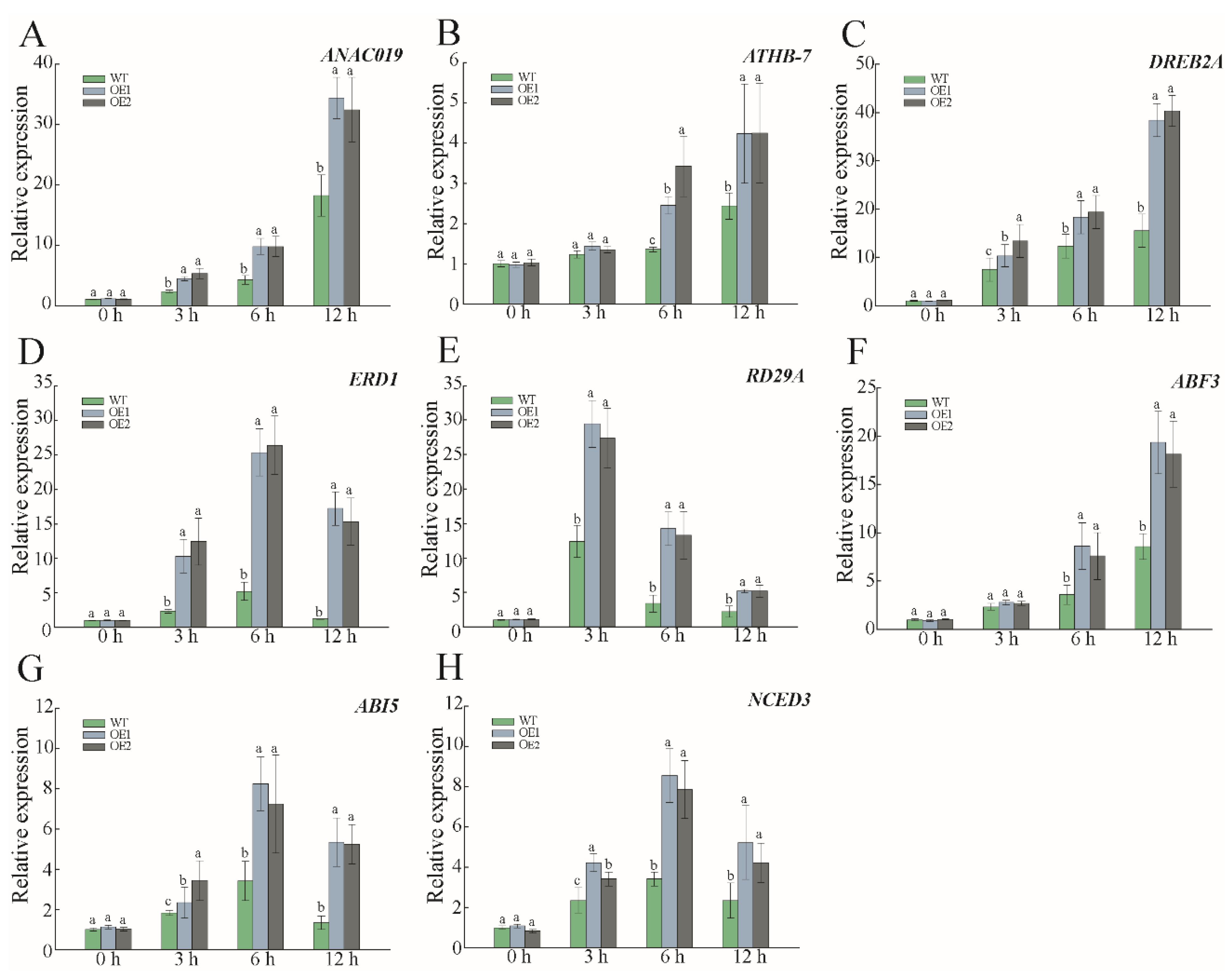
3.7. The Expression Level of Stress-Responsive Genes in PwNAC38 OE Lines under PEG Treatment
4. Discussion
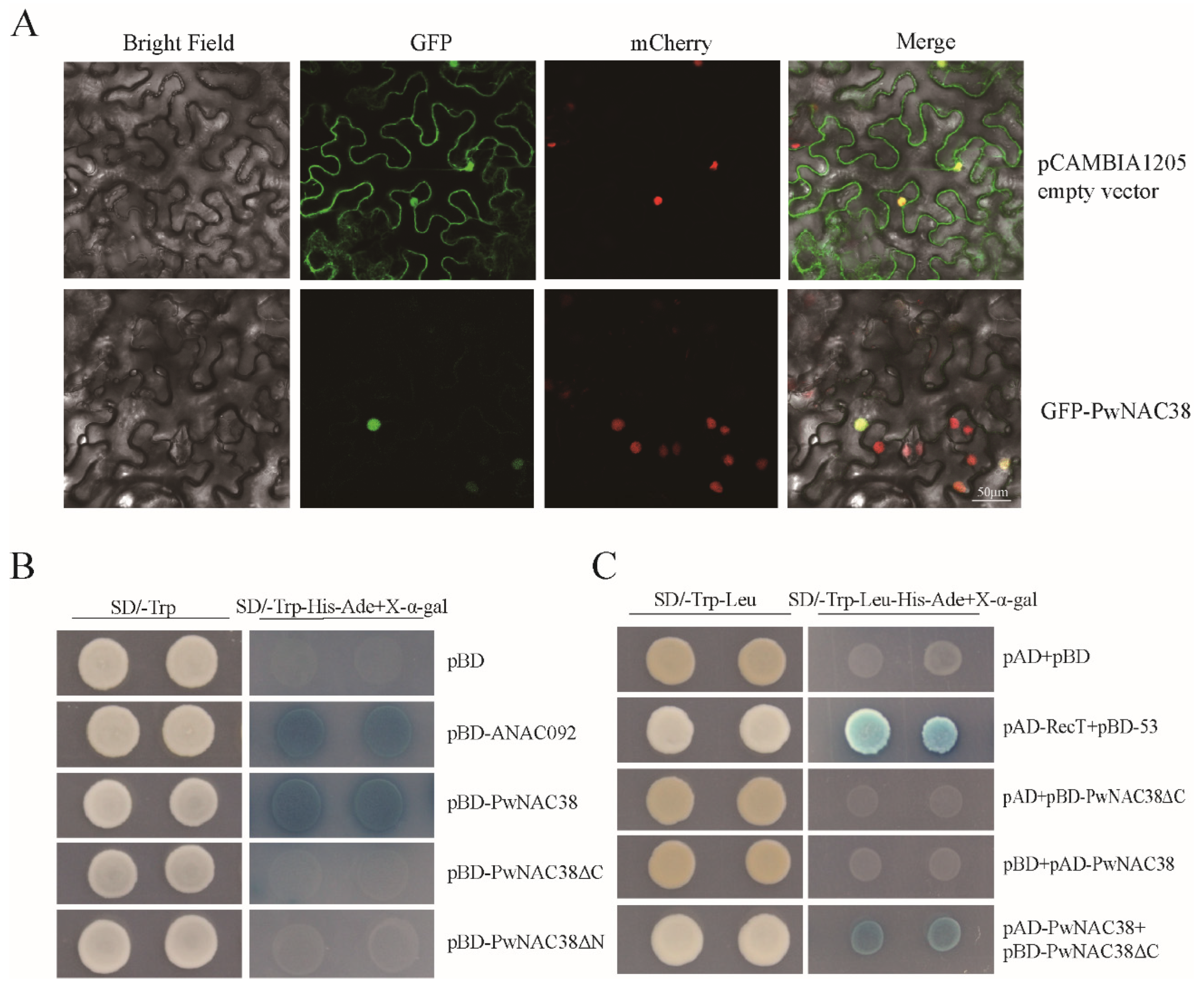
4.1. PwNAC38 Is a Nucleus-Located Transcription Factor That Can Form Homodimers
4.2. PwNAC38 Functions as a Positive Regulator to Drought and Salt Stress via ROS and ABA Pathways
4.3. The Effects of PwNAC38 on Plant Growth and Functional Similarity among Plant Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kazuko, Y.S.; Kazuo, S. Transcriptional Regulatory Networks in Cellular Responses and Tolerance to Dehydration and Cold Stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. DNA-binding specificity and molecular functions of NAC transcription factors. Plant Sci. 2005, 169, 785–797. [Google Scholar] [CrossRef]
- Nakashima, K.; Tran, L.P.; Van Nguyen, D.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Shinozaki, K.Y. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J. 2007, 51, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Kawai, Y.; Nakamura, N.; Futaki, S. The Structure of the Promoter Region for Rat Inducible Nitric Oxide Synthase Gene. Life Sci. 1997, 61, 45–49. [Google Scholar] [CrossRef]
- Jeong, J.S.; Park, Y.T.; Jung, H.; Park, S.; Kim, J. Rice NAC proteins act as homodimers and heterodimers. Plant Biotechnol. Rep. 2009, 3, 127–134. [Google Scholar] [CrossRef]
- Hao, Y.; Song, Q.; Chen, H.; Zou, H.; Wei, W.; Kang, X.; Ma, B.; Zhang, W.; Zhang, J.; Chen, S. Plant NAC-type transcription factor proteins contain a NARD domain for repression of transcriptional activation. Planta 2010, 232, 1033–1043. [Google Scholar] [CrossRef]
- Phan, T.L.; Rie, N.; Kazuko, Y.; Kazuo, S. Potential utilization of NAC transcription factors to enhance abiotic stress tolerance in plants by biotechnological approach. GM Crops 2010, 1, 32–39. [Google Scholar] [CrossRef]
- Yang, S.; Seo, P.J.; Yoon, H.; Park, C. The Arabidopsis NAC Transcription Factor VNI2 Integrates Abscisic Acid Signals into Leaf Senescence via the COR/RD Genes. Plant Cell 2011, 23, 2155–2168. [Google Scholar] [CrossRef]
- Zhong, R.Q.; Lee, C.H.; Ye, Z. Global Analysis of Direct Targets of Secondary Wall NAC Master Switches in Arabidopsis. Mol. Plant 2010, 3, 1087–1103. [Google Scholar] [CrossRef]
- Christianson, J.A.; Dennis, E.S.; Llewellyn, D.J.; Wilson, I.W. ATAF NAC transcription factors: Regulators of plant stress signaling. Plant Signal. Behav. 2010, 5, 428–432. [Google Scholar] [CrossRef]
- Yu, X.W.; Liu, Y.M.; Wang, S.; Yuan, T.; Zhankui, W.; Yingjie, S.; Hui, P.; Abudoukeyumu, M.; Ze, W.; Hua, Z.; et al. CarNAC4, a NAC-type chickpea transcription factor conferring enhanced drought and salt stress tolerances in Arabidopsis. Plant Cell Rep. 2016, 35, 613–627. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, H.; Liu, S.; Wang, X.; Zhang, Y.; Meng, Y.; Luo, D.; Chen, R. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.M.; Mi-Jeong, P.; Joon, S.P.; Jin-Su, S.; Hie-Joon, K.; Chung-Mo, P. Controlled nuclear import of the transcription factor NTL6 reveals a cytoplasmic role of SnRK2.8 in the drought-stress response. Biochem. J. 2012, 448, 353–363. [Google Scholar] [CrossRef]
- Liu, C.; Ma, H.Z.; Zhou, J.; Li, Z.X.; Peng, Z.H.; Guo, F.; Zhang, J.R. TsHD1 and TsNAC1 cooperatively play roles in plant growth and abiotic stress resistance of Thellungiella halophile. Plant J. 2019, 99, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Sanjana, N.; Himanshu, T.; Ganapathi, T.R. A banana NAC transcription factor (MusaSNAC1) impart drought tolerance by modulating stomatal closure and H2O2 content. Plant Mol. Biol. 2018, 96, 457–471. [Google Scholar] [CrossRef]
- Lu, M. Monitoring Radial Growth of Three Conifer Species in the Eastern Qilian Mountains. Master’s Thesis, Lanzhou University, Lanzhou, China, 2016. [Google Scholar]
- Ma, H. Functional Traits of Picea wilsonii Based on Tree Age and Altitude. Master’s Thesis, Shanxi Agricultural University, Jinzhong, China, 2019. [Google Scholar]
- Guo, Y.X.; Zhang, H.H.; Yuan, Y.H.; Cui, X.Y.; Zhang, L.Y. Identification and characterization of NAC genes in response to abiotic stress conditions in Picea wilsonii using transcriptome sequencing. Biotechnol. Biotec. Eq. 2020, 34, 93–103. [Google Scholar] [CrossRef]
- Zhang, H.H.; Cui, X.Y.; Guo, Y.X.; Luo, C.B.; Zhang, L.Y. Picea wilsonii transcription factor NAC2 enhanced plant tolerance to abiotic stress and participated in RFCP1-regulated flowering time. Plant Mol. Biol. 2018, 98, 471–493. [Google Scholar] [CrossRef]
- Liang, K.H.; Wang, A.B.; Yuan, Y.H.; Miao, Y.H.; Zhang, L.Y. Picea wilsonii NAC Transcription Factor PwNAC30 Negatively Regulates Abiotic Stress Tolerance in Transgenic Arabidopsis. Plant Mol. Biol. Rep. 2020, 38, 554–571. [Google Scholar] [CrossRef]
- Yu, M.X.; Liu, J.L.; Du, B.S.; Zhang, M.J.; Wang, A.B.; Zhang, L.Y. NAC Transcription Factor PwNAC11 Activates ERD1 by Interaction with ABF3 and DREB2A to Enhance Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 6952. [Google Scholar] [CrossRef]
- Joshi, P.S.; Parinita, A.; Agarwal, P.K. Overexpression of AlNAC1 from recretohalophyte Aeluropus lagopoides alleviates drought stress in transgenic tobacco. Environ. Exp. Bot. 2021, 181, 104277. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, L.; Gao, H.; Wang, X.; Li, Z.; Li, Y. The transcription factor MdMYB2 influences cold tolerance and anthocyanin accumulation by activating SUMO E3 ligase MdSIZ1 in apple. Plant Physiol. 2022, 189, 2044–2060. [Google Scholar] [CrossRef] [PubMed]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem Gene of Petunia Is Required for Pattern Formation in Embryos and Flowers and Is Expressed at Meristem and Primordia Boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef]
- Wang, B.; Wei, J.; Song, N.; Wang, N.; Zhao, J.; Kang, Z. A novel wheat NAC transcription factor, TaNAC30, negatively regulates resistance of wheat to stripe rust. J. Integr. Plant Biol. 2018, 60, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, H.; Li, D.; Huang, L.; Hong, Y.; Ding, X.S.; Nelson, R.S.; Zhou, X.; Song, F. Functions of rice NAC transcriptional factors, ONAC122 and ONAC131, in defense responses against Magnaporthe grisea. Plant Mol. Biol. 2013, 81, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Chen, L.; Xie, H. Molecular characterization of banana NAC transcription factors and their interactions with ethylene signalling component EIL during fruit ripening. J. Exp. Bot. 2012, 63, 5171–5187. [Google Scholar] [CrossRef]
- Jia, D.F.; Gong, X.Q.; Li, M.J.; Li, C.; Sun, T.T.; Ma, F.W. Overexpression of a Novel Apple NAC Transcription Factor Gene, MdNAC1, Confers the Dwarf Phenotype in Transgenic Apple (Malus domestica). Genes 2018, 9, 229. [Google Scholar] [CrossRef]
- Fu, C.C.; Han, Y.C.; Fan, Z.Q.; Jian-Ye, C.; Wei-Xin, C.; Wang-Jin, L.; Jian-Fei, K. The Papaya Transcription Factor CpNAC1 Modulates Carotenoid Biosynthesis through Activating Phytoene Desaturase Genes CpPDS2/4 during Fruit Ripening. J. Agric. Food Chem. 2016, 64, 5454–5463. [Google Scholar] [CrossRef]
- Jin, R.; Zhu, Q.G.; Shen, X.Y.; Wang, M.M.; Jamil, W.; Grierson, D.; Yin, X.R.; Chen, K.S. DkNAC7, a novel high-CO2/hypoxia-induced NAC transcription factor, regulates persimmon fruit de-astringency. PLoS ONE 2018, 13, e0194326. [Google Scholar] [CrossRef]
- Ng, S.; Ivanova, A.; Duncan, O.; Law, S.R.; Van Aken, O.; De Clercq, I.; Wang, Y.; Carrie, C.; Xu, L.; Kmiec, B.; et al. A Membrane-Bound NAC Transcription Factor, ANAC017, Mediates Mitochondrial Retrograde Signaling in Arabidopsis. Plant Cell 2013, 25, 3450–3471. [Google Scholar] [CrossRef]
- Liu, X.H.; Lyu, Y.S.; Yang, W.; Yang, Z.T.; Lu, S.J.; Liu, J.X. A membrane-associated NAC transcription factor OsNTL3 is involved in thermotolerance in rice. Plant Biotechnol. J. 2020, 18, 1317–1329. [Google Scholar] [CrossRef]
- Sun, L.; Yang, Z.T.; Song, Z.T.; Wang, M.J.; Sun, L.; Lu, S.J.; Liu, J.X. The plant-specific transcription factor gene NAC103 is induced by bZIP60 through a new cis-regulatory element to modulate the unfolded protein response in Arabidopsis. Plant J. 2013, 76, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.F.; Wang, C.; Yan, J.L.; Guo, X.H.; Wu, F.F.; Yang, B.; Deyholos, M.K.; Jiang, Y. Functional characterization of NAC55 transcription factor from oilseed rape (Brassica napus L.) as a novel transcriptional activator modulating reactive oxygen species accumulation and cell death. Plant Mol. Biol. 2016, 92, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.G.; Zhang, H.Y.; Qian, X.Y.; Li, A.; Zhao, G.Y.; Jing, R.L. TaNAC2, a NAC-type wheat transcription factor conferring enhanced multiple abiotic stress tolerances in Arabidopsis. J. Exp. Bot. 2012, 63, 2933–2946. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K.; et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.S.; Yuan, Y.; Wu, Q.; Wang, J.; Li, J.G.; Zhao, M.L. LcEIL2/3 are involved in fruitlet abscission via activating genes related to ethylene biosynthesis and cell wall remodeling in litchi. Plant J. 2020, 103, 1338–1350. [Google Scholar] [CrossRef]
- An, J.; Zhang, X.; Liu, Y.; Wang, X.; You, C.; Hao, Y. ABI5 regulates ABA-induced anthocyanin biosynthesis by modulating the MYB1-bHLH3 complex in apple. J. Exp. Bot. 2021, 72, 1460–1472. [Google Scholar] [CrossRef]
- Truong, H.A.; Lee, S.; Trịnh, C.S.; Lee, W.J.; Chung, E.; Hong, S.; Lee, H. Overexpression of the HDA15 Gene Confers Resistance to Salt Stress by the Induction of NCED3, an ABA Biosynthesis Enzyme. Front. Plant Sci. 2021, 12, 640443. [Google Scholar] [CrossRef]
- Han, D.G.; Du, M.; Zhou, Z.Y.; Wang, S.; Li, T.M.; Han, J.X.; Xu, T.L.; Yang, G.H. Overexpression of a Malus baccata NAC Transcription Factor Gene MbNAC25 Increases Cold and Salinity Tolerance in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1198. [Google Scholar] [CrossRef]
- Takasaki, H.; Maruyama, K.; Takahashi, F.; Fujita, M.; Yoshida, T.; Nakashima, K.; Myouga, F.; Toyooka, K.; Yamaguchi Shinozaki, K.; Shinozaki, K. SNAC-As, stress-responsive NAC transcription factors, mediate ABA-inducible leaf senescence. Plant J. 2015, 84, 1114–1123. [Google Scholar] [CrossRef]
- Zhang, K.W.; Gan, S.S. An Abscisic Acid-AtNAP Transcription Factor-SAG113 Protein Phosphatase 2C Regulatory Chain for Controlling Dehydration in Senescing Arabidopsis Leaves. Plant Physiol. 2012, 158, 961–969. [Google Scholar] [CrossRef]
- Yu, Y.C.; Wang, L.; Chen, J.C.; Liu, Z.; Chung-Mo, P.; Xiang, F.N. WRKY71 Acts Antagonistically Against Salt-Delayed Flowering in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Pham-Thi, M.; Sug, K.J.; Songhwa, C.; Mi, J.K.; Gang-Seob, L.; Dong-Eun, K.; Jong-Joo, C.; Ik, S.S.; Hie, N.B.; Yeon-Ki, K. A WUSCHEL Homeobox Transcription Factor, OsWOX13, Enhances Drought Tolerance and Triggers Early Flowering in Rice. Mol. Cells 2018, 41, 781–798. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Liu, J.; Hu, J.; Zhou, J.; Zhang, M.; Cao, Y.; Zhang, L. Picea wilsonii Transcription Factor PwNAC38 Promotes Drought and Salt Tolerance in Transgenic Arabidopsis and Apple Calli. Forests 2022, 13, 1304. https://doi.org/10.3390/f13081304
Yu M, Liu J, Hu J, Zhou J, Zhang M, Cao Y, Zhang L. Picea wilsonii Transcription Factor PwNAC38 Promotes Drought and Salt Tolerance in Transgenic Arabidopsis and Apple Calli. Forests. 2022; 13(8):1304. https://doi.org/10.3390/f13081304
Chicago/Turabian StyleYu, Mingxin, Junling Liu, Jing Hu, Jing Zhou, Mengjuan Zhang, Yibo Cao, and Lingyun Zhang. 2022. "Picea wilsonii Transcription Factor PwNAC38 Promotes Drought and Salt Tolerance in Transgenic Arabidopsis and Apple Calli" Forests 13, no. 8: 1304. https://doi.org/10.3390/f13081304
APA StyleYu, M., Liu, J., Hu, J., Zhou, J., Zhang, M., Cao, Y., & Zhang, L. (2022). Picea wilsonii Transcription Factor PwNAC38 Promotes Drought and Salt Tolerance in Transgenic Arabidopsis and Apple Calli. Forests, 13(8), 1304. https://doi.org/10.3390/f13081304