
Effect of Seasonal Variation on Leaf Cuticular Waxes’ Composition in the Mediterranean Cork Oak (Quercus suber L.)



Abstract
1. Introduction
2. Material and Methods
2.1. Sampling
2.2. Morphological Variables
2.3. Extraction of Cuticular Waxes
2.4. Cuticular Wax Composition
2.5. Statistical Analysis
3. Results
3.1. Leaf Area and SLA
3.2. Cuticular Wax Content
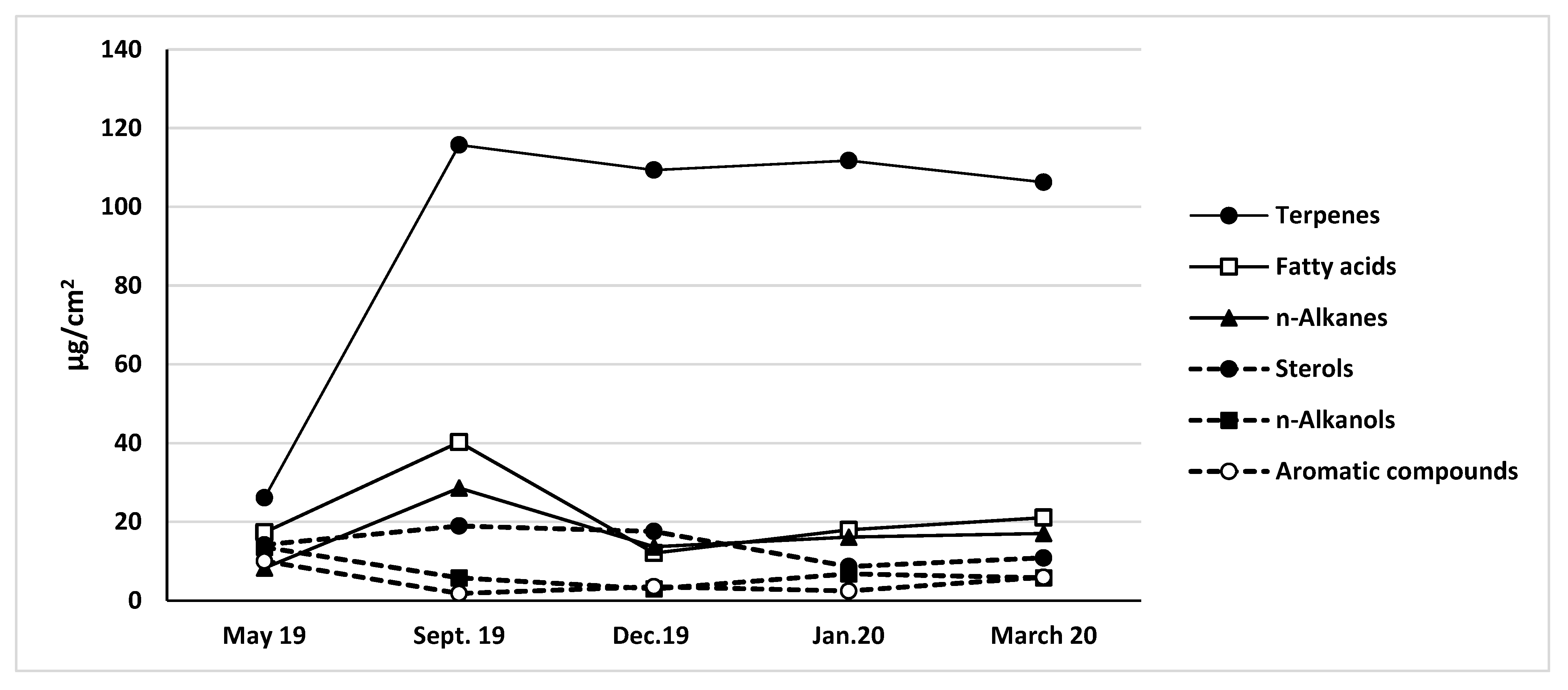
3.3. Cuticular Wax Composition
4. Discussion
4.1. Leaf Area and SLA
4.2. Cuticular Wax Content
4.3. Cuticular Wax Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of plant cuticular waxes. In Biology of the Plant Puticle; Riederer, M., Müller, C., Eds.; Blackwell: Oxford, MS, USA, 2006; pp. 145–181. [Google Scholar]
- Buschhaus, C.; Jetter, R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal surfaces? J. Exp. Bot. 2011, 62, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticleds. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Zeisler-Diehl, V.; Müller, Y.; Schreiber, L. Epicuticular wax on leaf cuticles does not establish the transpiration barrier, which is essentially formed by intracuticular wax. J. Plant Physiol. 2018, 227, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Seufert, P.; Staiger, S.; Arand, K.; Bueno, A.; Burghardt, M.; Riederer, M. Building a barrier: The influence of different wax fractions on the water transpiration barrier of leaf cuticles. Front. Plant Sci. 2022, 12, 766602. [Google Scholar] [CrossRef]
- Tsubaki, S.; Sugimura, K.; Teramoto, Y.; Yonemori, K.; Azuma, J.I. Cuticular membrane of Fuyu persimmon fruit is strengthened by triterpenoid nano-fillers. PLoS ONE 2013, 8, e75275. [Google Scholar] [CrossRef]
- Schuster, A.-C.; Burghardt, M.; Alfarhan, A.; Bueno, A.; Hedrich, R.; Leide, J.; Thomas, J.; Riederer, M. Effectiveness of cuticular transpiration barriers in a desert plant at controlling water loss at high temperatures. AoB Plants 2016, 8, plw027. [Google Scholar] [CrossRef]
- Pereira, H. Cork: Biology, Production and Uses; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Simões, R.; Rodrigues, A.; Ferreira-Dias, S.; Miranda, I.; Pereira, H. Chemical composition of cuticular waxes, pigments and morphology of leaves of Quercus suber trees from different provenances. Plants 2020, 9, 1165. [Google Scholar] [CrossRef]
- Simões, R.; Miranda, I.; Pereira, H. Chemical composition of leaf cutin in six Quercus suber provenances. Phytochemistry 2021, 181, 112570. [Google Scholar] [CrossRef]
- Pereira, J.S.; Beyschlag, G.; Lange, O.L.; Beyschlag, W.; Tenhunen, J.D. Comparative phenology of four mediterranean shrub species growing in Portugal. In Plant Response to Stress; Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel, W.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 503–514. [Google Scholar]
- Escudero, A.; Del Arco, J.M.; Sanz, I.C.; Ayala, J. Effects of leaf longevity and retranslocation efficiency on the retention time of nutrients in the leaf biomass of different woody species. Oecologia 1992, 90, 80–87. [Google Scholar] [CrossRef]
- Pausas, J.G.; Pereira, J.S.; Aronson, J. The tree. In Cork Oak Woodlands on the Edge: Conservation, Adaptive Management, and Restoration; Aronson, J., Pereira, J.S., Pausas, J.G., Eds.; Island Press: Washington, DC, USA, 2009; pp. 11–21. [Google Scholar]
- Oliveira, G.; Correia, O.; Martins-Loução, M.A.; Catarino, F. Phenological and growth patterns of the Mediterranean oak Quercus suber L. Trees 1994, 9, 41–46. [Google Scholar] [CrossRef]
- Fialho, C.; Lopes, F.; Perreira, H. The effect of cork removal on the radial growth and phenology of young cork oak trees. For. Ecol. Manag. 2001, 141, 251–258. [Google Scholar] [CrossRef]
- Sampaio, T.; Gonçalves, E.; Patrício, M.S.; Cota, T.M.; Almeida, M.H. Seed origin drives differences in survival and growth traits of cork oak (Quercus suber L.) populations. For. Ecol. Manag. 2019, 448, 267–277. [Google Scholar] [CrossRef]
- Varela, M.C. European Network for the Evaluation of Genetic Resources of Cork Oak for Appropriate Use in Breeding and Gene Conservation Strategies; INIA: Lisbon, Portugal, 2020. [Google Scholar]
- Kolattukudy, P.E.; Agrawal, V.P. Structure and composition of aliphatic constituents of potato tuber skin (suberin). Lipids 1974, 9, 682–691. [Google Scholar] [CrossRef]
- Ferreira, J.P.; Miranda, I.; Sen, A.; Pereira, H. Chemical and cellular features of virgin and reproduction cork from Quercus variabilis. Ind. Crops Prod. 2016, 94, 638–648. [Google Scholar] [CrossRef]
- Grant, O.M.; Tronina, L.; Ramalho, J.C.; Besson, C.K.; Lobo-Do-Vale, R.; Pereira, J.S.; Jones, H.G.; Chaves, M.M. The impact of drought on leaf physiology of Quercus suber L. trees: Comparison of an extreme drought event with chronic rainfall reduction. J. Exp. Bot. 2010, 61, 4361–4371. [Google Scholar] [CrossRef]
- Prats, K.A.; Brodersen, C.R.; Ashton, M.S. Influence of dry season on Quercus suber L. leaf traits in the Iberian Peninsula. Am. J. Bot. 2019, 106, 656–666. [Google Scholar] [CrossRef]
- Garcia-Plazaola, J.I.; Faria, T.; Abadía, J.; Abadía, A.; Chaves, M.M.; Pereira, J.S. Seasonal changes in xanthophyll composition and photosynthesis of cork oak (Quercus suber L.) leaves under Mediterranean climate. J. Exp. Bot. 1997, 48, 1667–1674. [Google Scholar] [CrossRef]
- Mediavilla, S.; Martín, I.; Babiano, J.; Escudero, A. Foliar plasticity related to gradients of heat and drought stress across crown orientations in three Mediterranean Quercus species. PLoS ONE 2019, 14, e0224462. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Hobbs, R.J.; Mayfield, M.M. Specific leaf area responses to environmental gradients through space and time. Ecology 2014, 95, 399–410. [Google Scholar] [CrossRef]
- Aranda, I.; Pardos, M.; Puértolas, J.; Jiménez, M.D.; Pardos, J.A. Water-use efficiency in cork oak (Quercus suber) is modified by the interaction of water and light availabilities. Tree Physiol. 2007, 27, 671–677. [Google Scholar] [CrossRef]
- Gouveia, A.C.; Freitas, H. Modulation of leaf attributes and water use efficiency in Quercus suber along a rainfall gradient. Trees 2009, 23, 267–275. [Google Scholar] [CrossRef]
- Ramírez-Valiente, J.A.; Sánchez-Gómez, D.; Aranda, I.; Valladares, F. Phenotypic plasticity and local adaptation in leaf ecophysiological traitsof 13 contrasting cork oak populations under different water availabilities. Tree Physiol. 2010, 30, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Passarinho, J.A.P.; Lamosa, P.; Baeta, J.P.; Santos, H.; Ricardo, C.P.P. Annual changes in the concentration of minerals and organic compounds of Quercus suber leaves. Physiol. Plant. 2006, 127, 100–110. [Google Scholar] [CrossRef]
- Prasad, R.B.N.; Gülz, P.G. Surface structure and chemical composition of leaf waxes from Quercus robur L., Acer pseudoplatanus L. and JugIans regia L. Z. Naturforsch. C 1990, 45, 813–817. [Google Scholar] [CrossRef]
- Gülz, P.-G.; Müller, E. Seasonal variation in the composition of epicuticular waxes of Quercus robur leaves. Z. Naturforsch. C 1992, 47, 800–806. [Google Scholar] [CrossRef]
- Jenks, M.A.; Gaston, C.H.; Goodwin, M.S.; Keith, J.A.; Teusink, R.S. Seasonal variation in cuticular waxes on hosta genotypes differing in leaf surface glaucousness. HortScience 2002, 37, 637–677. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their applicationin drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Broeckling, C.D.; Blancaflor, E.B.; Sledge, M.K.; Sumner, L.W.; Wang, Z.Y. Overexpression of WXP1, a putativeMedicago truncatula AP2 domain-containing transcription fac-tor gene, increases cuticular wax accumulation and enhancesdrought tolerance in transgenic alfalfa (Medicago sativa). Plant J. 2005, 42, 689–707. [Google Scholar] [CrossRef]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased accumu- lation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar] [CrossRef]
- Hauke, V.; Schreiber, L. Ontogenetic and seasonal development of wax composition and cuticular transpiration of ivy (Hedera helix L.) sun and shade leaves. Planta 1998, 207, 67–75. [Google Scholar] [CrossRef]
- Celano, G.; D’Auria, M.; Xiloyannis, C.; Mauriello, G.; Baldassarre, M. Composition and seasonal variation of soluble cuticular waxes in Actinidia deliciosa leaves. Nat. Prod. Res. 2006, 20, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Neinhuis, C.; Koch, K.; Barthlott, W. Movement and regeneration of epicuticlar waxes through plant cuticles. Planta 2001, 213, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Neinhuis, C.; Ensikat, H.-J.; Barthlott, W. Self assembly of epicuticular waxes on living plant surfaces imaged by atomic force microscopy (AFM). J. Exp. Bot. 2004, 55, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Bhushan, B.; Ensikat, H.-J.; Barthlott, W. Self-healing of voids in the wax coating on plant surfaces. Philos. Trans. Royal Society A 2009, 367, 1673–1688. [Google Scholar] [CrossRef] [PubMed]
- Jetter, R.; Schäffer, S. Chemical composition of the Prunus laurocerasus leaf surface. dynamic changes of the epicuticular wax film during leaf development. Plant Physiol. 2001, 126, 1725–1737. [Google Scholar] [CrossRef]
- Huth, M.A.; Huth, A.; Koch, K. Morphological diversity of β-diketone wax tubules on Eucalyptus gunnii leaves and real time observation of self-healing of defects in the wax layer. Aust. J. Bot. 2018, 66, 313–324. [Google Scholar] [CrossRef]
- Bueno, A.; Sancho-Knapik, D.; Gil-Pelegrín, E.; Leide, J.; Peguero-Pina, J.J.; Burghardt, M.; Riederer, M. Cuticular wax coverage and its transpiration barrier properties in Quercus coccifera L. leaves: Does the environment matter? Tree Physiol. 2020, 40, 827–840. [Google Scholar] [CrossRef]
- Huang, H.; Burghardt, M.; Schuster, A.; Leide, J.; Lara, I.; Riederer, M. Chemical compositions and water permeabilities of fruit and leaf cuticles of Olea europaea L. J. Agric. Food Chem. 2017, 65, 8790–8797. [Google Scholar] [CrossRef]
- Jetter, R.; Riederer, M. Localization of the transpiration barrier in the epi- and intracuticular waxes of eight plant species: Water transport resistances are associated with fatty acyl rather than alicyclic components. Plant Physiol. 2016, 170, 921–934. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| May 2019 | September 2019 | December 2019 | January 2020 | March 2020 | |
|---|---|---|---|---|---|
| Wax Content (µg/cm2) | 115.7 ± 52.6 | 235.6 ± 33.6 | 182.1 ± 31.6 | 215.1 ± 48.5 | 220.3 ± 56.0 |
| n-Alkanes | |||||
| Hexacosane (C26) | - | 0.16 ± 0.04 | 0.13 ± 0.15 | 0.13 ± 0.05 | 0.15 ± 0.17 |
| Heptacosane (C27) | 0.63 ± 0.24 | 1.06 ± 0.45 | 0.55 ± 0.45 | 0.68 ± 0.30 | 0.70 ± 0.17 |
| Octacosane (C28) | 0.21 ± 0.07 | 0.72 ± 0.20 | 0.46 ± 0.12 | 0.60 ± 0.19 | 0.46 ± 0.14 |
| Nonacosane (C29) | 4.75 ± 0.94 | 8.69 ± 3.38 | 7.23 ± 3.45 | 5.07 ± 2.61 | 5.24 ± 2.21 |
| Triacontane (C30) | 1.43 ± 0.54 | 1.38 ± 0.32 | 0.84 ± 0.40 | 0.93 ± 0.40 | 1.13 ± 0.29 |
| n-Alkanols | |||||
| Hexadecan-1-ol (C16OH) | 0.10 ± 0.03 | 0.15 ± 0.05 | 0.14 ± 0.10 | 0.11 ± 0.09 | 0.12 ± 0.06 |
| Docosan-1-ol (C22OH) | 0.89 ± 0.29 | 0.21 ± 0.10 | 0.12 ± 0.02 | 0.25 ± 0.10 | 0.18 ± 0.14 |
| Tretracosan-1-ol (C24OH) | 9.92 ± 6.1 | 1.80 ± 0.89 | 1.14 ± 0.68 | 2.23 ± 1.05 | 1.84 ± 0.46 |
| Pentacosan-1-ol (C25OH | 0.11 ± 0.07 | - | - | - | - |
| Hexacosan-1-ol (C26OH) | 0.35 ± 0.30 | - | - | - | - |
| Octacosn-1-ol (C28OH) | - | - | - | 0.32 ± 0.22 | 0.25 ± 0.26 |
| Dotriacontan-1-ol (C32OH) | - | 0.11 ± 0.10 | - | - | - |
| Fatty acids | |||||
| Saturated | |||||
| Decanoic acid (C10:0) | - | - | - | - | 0.21 ± 0.07 |
| Dodecanoic acid (C12:0) | 0.10 ± 0.03 | - | - | 0.10 ± 0.07 | 0.22 ± 0.04 |
| Tetradecanoic acid (C14:0) | 0.30 ± 0.16 | 0.12 ± 0.07 | 0.22 ± 0.11 | 0.15 ± 0.14 | 0.19 ± 0.23 |
| Hexadecanoic acid (C16:0) | 4.97 ± 2.76 | 1.32 ± 0.65 | 4.31 ± 2.62 | 2.07 ± 0.60 | 2.02 ± 0.42 |
| Octadecanoic acid (C18:0) | 0.51 ± 0.14 | 0.23 ± 0.09 | 0.17 ± 0.05 | 0.26 ± 0.14 | 0.20 ± 0.08 |
| Eicosanoic acid (C20:0) | 0.20 ± 0.05 | 0.21 ± 0.11 | - | 0.18 ± 0.04 | 0.17 ± 0.08 |
| Docosanoic acid (C22:0) | 0.32 ± 0.10 | 0.21 ± 0.17 | - | 0.26 ± 0.07 | 0.30 ± 0.17 |
| Tetracosanoic acid (C24:0) | 0.80 ± 0.24 | 0.24 ± 0.15 | - | 0.24 ± 0.08 | 0.20 ± 0.17 |
| Hexacosanoic acid (C26:0) | 0.71 ± 0.34 | 1.01 ± 0.43 | - | 0.56 ± 0.19 | 0.45 ± 0.16 |
| Octacosanoic acid (C28:0) | 1.49 ± 1.09 | 4.65 ± 2.89 | 0.18 ± 0.09 | 1.46 ± 0.44 | 1.69 ± 0.36 |
| Triacontanoic acid (C30:0) | 0.95 ± 1.01 | 8.15 ± 6.18 | 0.96 ± 0.74 | 2.28 ± 1.23 | 2.37 ± 0.61 |
| Dotriacontanoic acid (C32:0) | 0.13 ± 0.09 | 0.41 ± 0.15 | 0.10 ± 0.7 | 0.32 ± 0.07 | 0.31 ± 0.12 |
| Unsaturated | |||||
| 9,12-Octadecadienoic acid (C18:2) | 1.42 ± 1.14 | 0.10 ± 0.05 | - | - | 0.15 ± 0.08 |
| 9,12,15-Octadecatrienoic acid (C18:3) | 2.54 ± 2.19 | 0.26 ± 0.16 | 0.24 ± 0.12 | 0.17 ± 0.12 | 0.51 ± 0.14 |
| Glycerides | |||||
| Glycerol | 3.52 ± 1.32 | 0.12 ± 0.05 | 0.16 ± 0.07 | 0.33 ± 0.38 | 1.03 ± 0.39 |
| 4-Hydroxyphenylglycolic acid | 0.19 ± 0.21 | 0.35 ± 0.40 | 1.01 ± 0.45 | 1.18 ± 0.71 | - |
| 2-Palmitoglycerol | 0.63 ± 0.09 | 0.21 ± 0.10 | - | 0.21 ± 0.26 | -. |
| Glycerol monostearate | 0.17 ± 0.07 | - | - | - | - |
| 1-Monolinoleate Glycerol | 0.33 ±0.15 | - | - | - | - |
| Linolenoylglycerol | 0.13 ± 0.07 | - | - | - | 0.18 ± 0.12 |
| Sterols | |||||
| β-Systosterol | 11.94 ± 5.28 | 6.29 ± 1.86 | 9.64 ± 4.26 | 4.01 ± 1.11 | 4.91 ± 1.03 |
| Terpenes | |||||
| Diterpenes | |||||
| Phytol | 4.00 ± 2.40 | 0.61 ± 0.28 | 0.35 ± 0.39 | 0.49 ± 0.36 | 1.01 ±0.23 |
| Pentacylic triperpenes | |||||
| α-Amyrin | 1.61 ± 0.53 | 1.34 ± 0.40 | 0.72 ± 0.47 | 0.76 ± 0.29 | 0.64 ± 0.21 |
| β-Amyrin | 2.33 ± 0.95 | 3.98 ± 1.38 | 4.90 ± 2.74 | 3.50 ± 1.32 | 3.97 ± 1.25 |
| Germanicol | 0.27 ± 0.42 | 8.16 ± 1.69 | 8.10 ± 6.50 | 8.40 ± 2.47 | 6.38 ± 1.49 |
| Lupeol | 7.38 ± 5.71 | 28.20 ± 10.41 | 36.59 ± 11.13 | 27.75 ± 7.06 | 24.11 ± 6.81 |
| Epifriedelanol | 0.43 ± 0.32 | 2.38 ± 2.26 | 3.75 ± 3.25 | 1.92 ± 2.22 | 3.12 ± 2.38 |
| Erythrodiol | 0.21 ± 0.17 | - | 0.14 ± 0.12 | 1.62 ± 1.27 | 0.39 ± 0.30 |
| Friedelin | 1.14 ±0.96 | 2.47 ± 2.73 | 2.82 ± 2.82 | 2.16 ± 2.00 | 2.38 ±1.85 |
| Betulin | 0.40 ± 1.09 | 0.36 ± 0.10 | 0.80 ± 0.63 | 1.09 ± 0.81 | 1.46 ± 0.71 |
| Betulinic acid | 0.46 ± 0.25 | 0.42 ± 0.15 | 0.75 ± 0.63- | 0.98 ± 0.38 | 1.21 ± 0.53 |
| Ursolic acid | 0.44 ± 0.16 | 0.27 ± 0.10 | 0.24 ± 0.19 | 0.27 ± 0.16 | 0.28 ± 0.15 |
| Aromatic compounds | |||||
| Benzoic acid | 0.33 ± 0.29 | 0.16 ± 0.07 | 0.33 ± 0.09 | 0.27 ± 0.05 | 0.36 ± 0.06 |
| Hexadecy-(E)-p-coumarate | 4.93 ± 2.87 | - | 0.20 ± 0.09 | - | - |
| Vanillin acid | 0.47 ± 0.17 | 0.11 ± 0.06 | 0.11 ± 0.06 | 0.40 ± 0.49 | 0.16 ± 0.15 |
| 4-(Hydroxymethyl)phenol | 0.66 ± 0.31 | 0.10 ± 0.08 | 0.51 ± 0.54 | - | 1.36 ± 0.24 |
| Other compounds | |||||
| Myo-inositol | 1.35 ± 0.99 | 0.34 ± 0.18 | 0.10 ± 0.09 | 1.35 ± 2.31 | 1.18 ±1.27 |
| D-Fructose | 0.67 ± 0.20 | 0.32 ± 0.09 | - | 3.31 ± 2.80 | 1.04 ±1.36 |
| α-Tocopherol | 1.62 ±1.11 | - | - | - | 1.04 ± 1.36 |
| β-Tocopherol | 0.33 ± 0.13 | 0.95 ± 0.50 | - | - | - |
| γ-Tocopherol | - | - | 0.55 ± 0.62 | - | - |
| α-Tocopherolquinone | 0.19 ± 0.17 | 0.13 ± 0.10 | 0.10 ± 0.16 | 0.40 ± 0.46 | 2.05 ± 2.1 |
| Erythrono-1,4-lactone | 0.33 ± 0.14 | - | - | 0.37 ± 0.22 | - |
| Ribonic acid, 1,4-lactone | 0.33 ± 0.13 | 0.10 ± 0.06 | - | 0.51 ± 0.34 | 0.31 ± 0.22 |
| Quinic acid | - | - | - | 1.93 ± 0.76 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, R.; Miranda, I.; Pereira, H. Effect of Seasonal Variation on Leaf Cuticular Waxes’ Composition in the Mediterranean Cork Oak (Quercus suber L.). Forests 2022, 13, 1236. https://doi.org/10.3390/f13081236
Simões R, Miranda I, Pereira H. Effect of Seasonal Variation on Leaf Cuticular Waxes’ Composition in the Mediterranean Cork Oak (Quercus suber L.). Forests. 2022; 13(8):1236. https://doi.org/10.3390/f13081236
Chicago/Turabian StyleSimões, Rita, Isabel Miranda, and Helena Pereira. 2022. "Effect of Seasonal Variation on Leaf Cuticular Waxes’ Composition in the Mediterranean Cork Oak (Quercus suber L.)" Forests 13, no. 8: 1236. https://doi.org/10.3390/f13081236
APA StyleSimões, R., Miranda, I., & Pereira, H. (2022). Effect of Seasonal Variation on Leaf Cuticular Waxes’ Composition in the Mediterranean Cork Oak (Quercus suber L.). Forests, 13(8), 1236. https://doi.org/10.3390/f13081236