Abstract
Jalisco fir (Abies jaliscana) is an endemic tree with restricted distribution, and is considered a Pliocene relict inhabiting the cloud montane forests in the humid-temperate upper zone of western Jalisco Mountains. Despite its ecological importance, to date, there are no studies for this taxon aimed to understand its vulnerability to regional climate variability. We used dendrochronological techniques to investigate tree-ring patterns and the effects of climate variability on the radial growth of Jalisco fir through its elevational range in both southern and northern aspects. We developed tree-ring series that ranged in length from 79 to 152 years. High-elevation trees (2400 m.a.s.l.) showed higher annual radial growth compared to those present in low-elevation sites. Over the last 150 years, Abies jaliscana trees have had higher annual radial growth rates compared to those exhibited by other Abies species. Both winter and early spring precipitation are the main factors determining the growth rates at all the elevation sites for both aspects. In the cold season, mean maximum temperature negatively affected ring-width growth at most of the elevation zones except for two study sites. We conclude that the radial growth of A. jaliscana appears to be smaller during lower rainfall periods in the dry seasons (winter and early spring), and A. jaliscana annual radial growth responds differently to regional climate variability along the elevational gradient and mountain aspects.
1. Introduction
In high-elevation mountains, climatic change may affect more rapidly the population structure, the distribution, and the annual growth rates of tree species. In these areas, the radial growth of trees can also be subject to environmental gradients related to elevation [1]. In temperate environments, growth conditions in tree species can vary widely along altitudinal gradients, and these variations are recorded in tree rings [2,3,4]. Previous studies have found that the relationship between tree growth and climate frequently varies in response to elevation [5,6]. In some cases, the temperature is the most limiting factor of growth in subalpine and timberline zones [1,6,7], in other cases both temperature and precipitation affect ring width (RW), which is a measure of radial growth [8]. In contrast, the growth of tree species at lower elevations usually shows a significant and positive correlation with precipitation [9,10]. Nonetheless, forests and tree species do not always conform to these patterns; for instance, tree growth at mid-elevation (2270–2800 m.a.s.l.) and high-elevation (>3000 m.a.s.l.) sites in north and central Mexico (20–30° N) is mainly correlated with winter-spring and total regional precipitation, not temperature [11,12,13,14,15]. Additionally, in other montane forests located in subtropical and temperate geographic regions, different growth trends have been observed at low-, middle-, and high-elevation belts; both precipitation and temperature have been reported as highly correlated at different elevations [2,16]. Therefore, variations in tree radial growth along elevational gradients are mostly influenced by regional climate variability.
Jalisco fir (Abies jaliscana (Martínez) Mantilla, Shalisko & A. Vázquez) is a Pliocene relict taxon and one of the dominant tree species that inhabit the cloud montane forests in the humid-temperate upper zone of the mountain ranges of western Jalisco, Mexico [17,18]. In this region, A. jaliscana occurs from approximately 1700 to 2447 m.a.s.l., and it is dominant above 2260 m. Studies in western, south, and central Mexico have used dendrochronological techniques to develop climatic reconstructions and determine the climatic response in the radial growth of Abies and other conifers. For instance, Astudillo-Sánchez et al. [8] used a ring-width chronology of Pinus hartwegii to explain the climate states during agricultural crises and climate phenomena in central Mexico. Additionally, using growth rings of Abies religiosa, Cerano-Paredes et al. [13] reconstructed the winter-spring precipitation in Tancítaro Peak National Park, Michoacán, Mexico. Carlón-Allende et al. [19,20] studied the effect of precipitation and temperature on the radial growth, earlywood, and latewood of Pinus pseudostrobus and A. religiosa in the Monarch Butterfly Biosphere Reserve, Mexico. Aquino-Ramírez et al. [21] determined the response of A. guatemalensis Rehder to climatic variability. Villanueva-Díaz et al. [22] developed a dendrochronological network for the Armería-Coahuayana basin in southern Jalisco, Mexico. Given that the region of western Mexico is located in the transitional zone of two morphotectonic mountainous provinces (Trans-Mexican Volcanic Belt (TMVB) and Sierra Madre del Sur (SMS)), understanding the growth response of A. jaliscana along elevation gradients is of foremost importance for sustainable forest management and conservation in the context of climatic change; however, to date, there are no studies for Abies jaliscana using RW chronologies along elevational gradients with respect to climate variability. Despite the importance and potential of the high-elevation mountains in TMVB, there is a lack of dendroecological studies focused on altitudinal gradients in this morphotectonic province, except for one study on A. religiosa and Pinus pseudostrobus; however, the research only addresses the issue of fire regime inferred from the age structure along an elevational gradient and not radial growth [23]. The present study represents the first effort focused on elevational gradients and climatic factors that influence the annual radial growth of Mexican fir species. It aims to contribute to a better understanding of how precipitation and temperature variables may impact the growth of conifer species that thrive in high-elevation areas.
We sampled A. jaliscana along its distributional range in elevation and both south and north aspects in the mountains of western Jalisco, Mexico, and used dendrochronological techniques in order to: (1) develop ring-width chronologies in trees of A. jaliscana; (2) examine the patterns of variation in tree-ring growth along the elevational gradient and in north and south aspects; and (3) identify and analyze the climatic variables that influence the radial growth of A. jaliscana in western Jalisco, Mexico.
2. Materials and Methods
2.1. Study Area
We carried out the study along two elevational gradients in the north and south aspects of the highlands of western Jalisco, Mexico (Figure 1); elevation in the study area ranges from 1700 to 2400 m. Based on the WorldClim database [24], annual precipitation ranges from 1462 to 1512 mm, most of which falls between June and September; whereas annual mean temperature ranges from 17 to 15 °C below 2000 m and from 14.8 to 13 °C above 2000 m.a.s.l. [17]. The nearest meteorological station for one fir forest stand is located in the Cumbre de Guadalupe village at 2120 m of elevation [25], which registered an annual mean temperature of 14.2 °C and annual precipitation of 2003 mm. Vegetation of the study area comprises the cloud forest, which occurs in humid ravines from 1700–2250 m of elevation, whereas mixed oak-pine forests and pine forests occur around 1700–2100 m. Conifer forests dominated by Abies jaliscana occupy humid areas above 2260 m in elevation, where the latter usually develops within mixed forests with some pine species (Pinus montezumae and P. pseudostrobus) or forms monospecific forests above 2300 m.a.s.l. [17,26]. Carpinus caroliniana, Cornus disciflora, Clethra hartwegii, Persea hintonii, Styrax argenteus, and Symplocos citrea are common understory species. The climate in these forests is humid temperate with a summer rainfall regime (Cm) [27], although moderate precipitation can occur throughout the year (Figure 2A). Following Holdridge life zones [28], these forests correspond to the warm-temperate moist forest zone, but the highest-elevation study sites are very close to the cool-temperate zone and are colder than other forests that thrive in these highlands. Following the global bioclimatic classification by Rivas-Martínez et al. [29], the area is located in the tropical pluviseasonal macrobioclimate whose main characteristic is that the rainfall regime occurs in summer (Figure 2), and the Mesotropical and Supratropical thermotypes belts, where humid ombrotype occurs. The locality Cumbre de Guadalupe (south aspect) exhibits a tropical pluvial macrobioclimate and a warm-temperate wet forest zone according to Holdridge’s scale.
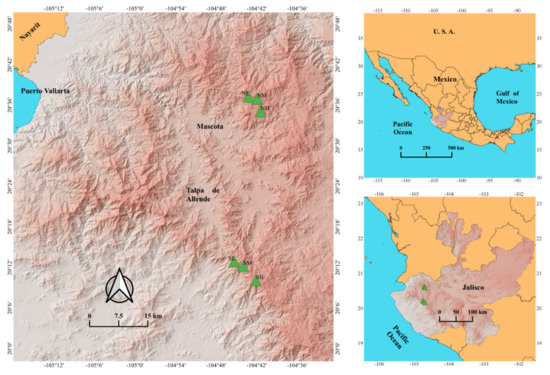
Figure 1.
Location of the sampling sites in the mountains of western Jalisco, Mexico. NL, low-elevation north aspect; NM, mid-elevation north aspect; NH, high-elevation north aspect; SL, low-elevation south aspect; SM, mid-elevation south aspect; SH, high-elevation south.
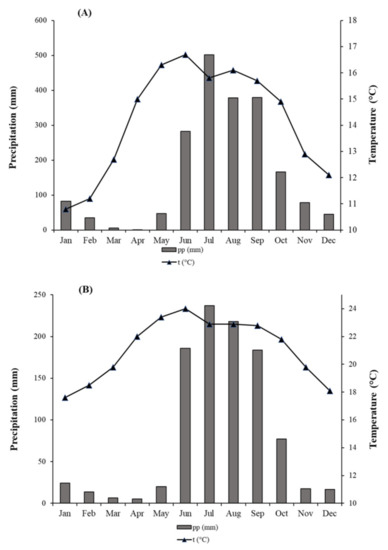
Figure 2.
Climatic characteristics at meteorological stations: (A) = Cumbre de Guadalupe; (B) = Mascota (1961–2014). Average monthly temperature (black lines) and average total monthly precipitation (gray bars).
2.2. Sample Collection
We sampled at three mountainous sites, on the north and south aspects (Figure 1) along an elevational range of the studied Jalisco fir: (1) in the cloud-forest zone at the lowest elevation, codominated by Acer binzayedii, A. jaliscana, and other cloud-forest species, (2) in the transitional zone between cloud forest and conifer forest at the middle elevation, where the dominance is distributed among species of Pinus, Quercus, and A. jaliscana, and (3) at the higher elevation where Abies jaliscana is the dominant species (Table 1). At each site, two or three increment cores of 5.1 mm width were extracted from 20 to 25 trees with a Pressler increment borer; sampled trees were located within a 500 m wide corridor parallel to the dominant slope direction. All the large and presumably old fir trees within the corridor were selected for increment core sampling at breast height (1.3 m above ground).

Table 1.
Characteristics of sampling sites in the mountains of western Jalisco, Mexico.
2.3. Tree-Ring Chronologies
The increment cores were mounted, sanded, and cross dated using standard procedures [30], and the tree-ring series were measured with a VELMEX TA stage micrometer at a 0.001 mm resolution. Crossdating and measurements were verified with COFECHA [31,32].
Ring-width measurements were standardized with ARSTAN [33]. All series were first detrended either with a negative exponential curve or a straight line with a negative slope and then second with a spline with a frequency response of 0.5 at wavelengths N/2, where N is the length of the time series [34]. The spline minimizes low-frequency trends still found in the detrended ring-width series and maximizes the high-frequency variability important to improve the climatic response of the chronology.
We used a double detrending standardization procedure based on the annual radial growth trend. The common growth trend for most of the conifers in Mexico including the Abies genus is a negative exponential curve, where annual radial growth is greater in the first years, but declines due to an increased basal area as the tree becomes older. Based on this concept, the first standardization option was a negative exponential curve that we considered suitable to eliminate the aged-related noise and maximize the climate signal [35].
Descriptive statistics were calculated for each chronology, to compare among the chronologies of sample plots. The mean sensitivity (MS) is a measure of year-to-year variability in growth [36]. First-order autocorrelation is a measure of the association between growth in the previous year and that in the current year [2]. RBAR is the mean correlation coefficient among the tree-ring series. Expressed population signal (EPS) assesses the degree to which a chronology represents a hypothetical chronology based on an infinite number of cores [37]. Standard deviation (SD) is an indicator of the interannual variation in tree-ring growth of a chronology. The signal-to-noise ratio (SNR) is a measure of the strength of the common signal relative to the uncommon signal of noise. A linear regression analysis was performed in order to describe the relationships among stand elevation, MS, and SD.
2.4. Climate Records and Correlation Analysis
Precipitation and temperature data spanning the period 1961–2014 from a single climate station (Mascota) (Figure 2B) were obtained from CLICOM [38] and used for north and south aspects because of their close geographical proximity. This climate station is located ca. 14 and 39 km apart from each aspect, north and south, respectively. The station was selected based on geographic coverage, correlation of climate data with tree growth, length of records, and data quality. We discarded the Cumbre de Guadalupe station due to its short length (12 years) (Figure 2A).
Standard Pearson correlation function analysis was applied to determine correlations between the derived chronologies and the climatic variables [36]. Analyses were performed with the Dendroclim software [39], using the derived mean monthly temperature and total monthly precipitation. Correlation analyses between the ring-width index (RWI) of each residual chronology and climatic variables were performed for 18 months in total, starting in July of the previous growing season, and ending in December of the current growing season.
3. Results
3.1. Tree Ring-Width Chronologies and Statistics
Comparisons of ring-width chronologies for Abies jaliscana across elevational gradients are shown in Figure 3 and Figure 4, and their corresponding statistics in Table 2. The mean tree age across all of the forest stands ranged from 52 to 84.3 years, with younger trees at the highest-elevation stand (ca. 2400 m.a.s.l.), and the oldest individual recorded (152 years old) located at the lowest-elevation site (SL). The total length for the combined ring-width chronologies extends back to the middle 19th century. The mean sensitivity and standard deviation decreased along the elevational gradient (Table 2); however, linear regression models did not show significant differences (Supplementary Figure S1). Most of the chronologies show that growth declined during the beginning of the present century (years 2000–2010) followed by increased growth in recent years (2013–2017).
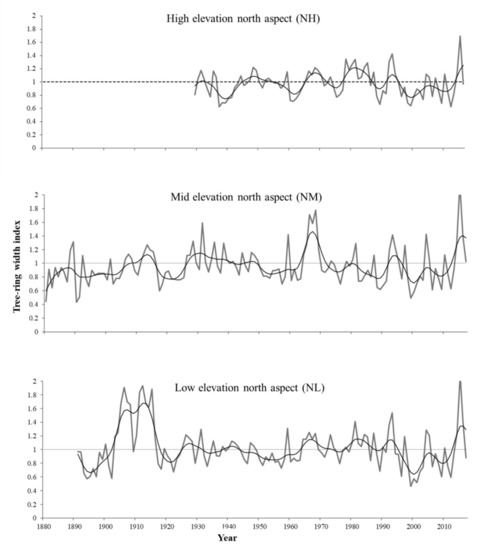
Figure 3.
Ring-width chronologies of A. jaliscana in the north aspect of the mountains in western Jalisco, Mexico. The gray line corresponds to the annual ring-width indices; the flexible line is a decadal spline to highlight low-frequency events, and the dotted line is the mean of the ring-width series.
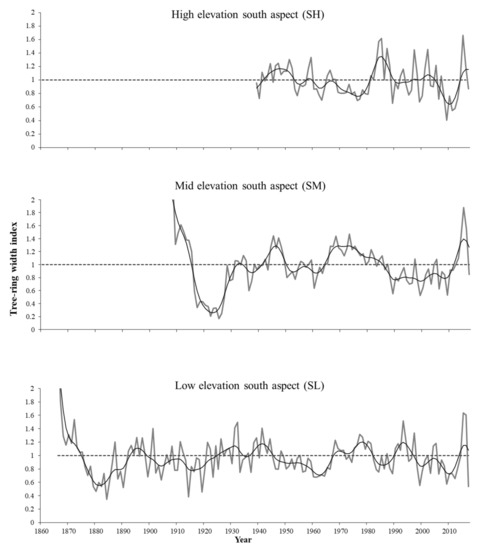
Figure 4.
Ring-width chronologies of A. jaliscana in the south aspect of the mountains in western Jalisco, Mexico. The gray line corresponds to the annual ring-width indices; the flexible line is a decadal spline to highlight low-frequency events, and the dotted line is the mean of the ring-width series.
Table 2.
Dendrochronological parameters of Abies jaliscana ring-width chronologies in the mountains of western Jalisco, Mexico.
The largest mean sensitivity and standard deviations occurred at the lower sites (NL and SL) and the middle distribution zone (NM) in the northern and southern aspects (Table 2). The mean ring widths tend to increase along the elevational gradient in the north aspect, whereas in the south aspect there is not a consistent variation pattern. The north sites NH and NM had a larger signal-to-noise ratio than the southern sites; only one site (NL) in the north aspect had a value lower than two sites in the south aspect. The first-order autocorrelations of chronologies range from 0.392 to 0.704, with the largest values in the NL site at the northern aspect, and the SM site at the southern aspect (Table 2). RBAR ranged from 0.288 to 0.403, and EPS varied from 0.689 to 0.9. Only one site (NM) reached an EPS ≥ 0.85, and for tree sensitivity, only a minimum sampled depth of four trees was required to reach that EPS in one stand where the elevation was 2260 m.a.s.l., in comparison to the other sites that demanded a greater sampled depth.
3.2. Response to Climatic Conditions
Maximum temperatures in winter and spring of the current year were negatively correlated with the ring-width index at low-, mid-, and high-elevation zones in the north aspect. In addition, the minimum temperature in February of the current year was negatively correlated with RWI in the three distribution zones of the northern slopes (NL, NM, and NH) (Figure 5). Similarly, precipitation in the early and late autumn of the previous year, and winter-spring rainfall of the current year, show positive correlations with RWI at the low and mid distribution zones; while precipitation in late autumn of the previous year, winter, and august of the current year are positively correlated at the high-elevation zone. In contrast, precipitation in July of the current year is negatively correlated with NH (Figure 5).
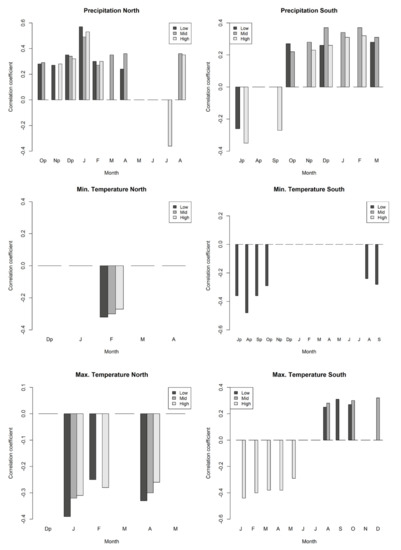
Figure 5.
Correlation coefficients between tree-ring width and mean monthly climatic variables (precipitation, minimum temperature, and maximum temperature). All values shown are statistically significant (p < 0.05), Dendroclim only shows significant values. Capital letters are months initials. The letter “p” next to some month initials denotes “previous”.
In the south aspect, the maximum temperature in winter and spring of the current year shows negative correlations with RWI at the high distribution zone (site SH), whereas maximum temperatures in late summer and autumn of the current year are positively correlated with RWI at the low and mid distribution zones (sites SL and SM) (Figure 5). Minimum temperature shows no significant correlation with RWI chronologies at the mid and high distribution zones, but radial growth is negatively correlated with minimum temperature in the summer and early autumn of the previous year and late summer of the current year at the low distribution zone (site SL) (Figure 5). Precipitation in autumn of the previous year and winter of the current year shows positive correlations with RWI at the three distribution zones, whereas precipitation in July of the previous year shows significant negative correlations with RWI at the high- and low-elevation zones, as is the case for precipitation in September of the previous year in SH (Figure 5).
4. Discussion
Trends in tree growth and their interrelations between rings and climate conditions vary widely among regions, elevations, and over time [2]. This research documents the variability in growth of Abies jaliscana in relation to elevation range and considers two aspects (north and south). Although some differences in the radial growth of A. jaliscana were found at each elevational belt, differences between tree ring parameters and elevation were not statistically significant. Even though in the north aspect there was a clear increase in mean ring-width in the site with the highest elevation (Table 2), this increase is not related to precipitation since this does not vary along the elevational gradient (Table 1). Therefore, we can infer that the increase in ring width at the high-elevation site may be related to less evapotranspiration due to a lower temperature that occurs at high elevations, and greater retention of moisture in the soil by having flatter reliefs (lesser slopes, Table 1) and deeper soils. The chronologies obtained (Figure 3 and Figure 4) are similar in length to those reported for other Abies species from the TMVB [13,20], however, a chronology from Abies guatemalensis (in all probability being A. hickelii the correct species) in the Sierra Norte de Oaxaca had a reported length of 206 years [21]. Other studies reported chronologies extended for 300 years in Abies guatemalensis from Los Cuchumatanes, Guatemala [40], and for 353 years in Abies concolor from San Pedro Mártir National Park, Baja California [41]. The shorter length of the chronologies observed in the TMVB could be explained by a high incidence of commercial and illegal logging since it is a region with greater anthropic pressure than other regions.
According to the estimated values of mean sensitivity, the NM and SL sites (northern and southern aspects, respectively) exhibit the strongest climatic signal in terms of seasonal precipitation and temperature (monthly maximum average temperature and monthly minimum average temperature) [42]. These sites had the longest chronologies and the oldest trees; the lowest values of mean ring width were also found at the NM site. On the other hand, the dominance of young trees in this mountain range could be attributed to illegal logging and extensive land-use changes, a situation that may prevent the development of longer chronologies [20,43], although it is accepted that stressful environmental conditions favor the presence of the oldest trees and the development of the longest chronologies [36]. For example, in colder subalpine areas (i.e., more stressful temperature conditions) other Abies species such as A. fargesii, A. georgei var. smithii, A. lasiocarpa, and A. spectabilis, exhibit chronologies over 200 years in length [1,2,6,44]. Conversely, in these subalpine firs, the mean ring width is lower than the values recorded in most of our studied sites (NH, SH, SM, and SL), which could be attributed to better site conditions (i.e., high atmospheric humidity and temperate environment throughout the year). In addition, the mean ring width in A. jaliscana is larger than the values reported for other temperate fir species such as A. borisii-regis in southeast Albania and A. veitchii in central Japan [4,16], which reflects more constraining environmental conditions at high latitudes (for example, high seasonality with very high average temperatures in summer or extremely cold conditions in winter), probably eliciting adaptive responses from these fir species in terms of size and radial growth. It should be noted that the length of Abies veitchii chronologies is very similar to those recorded for the high-altitude sites (NH and SH) in our study area; in addition, the Abies borisii-regis chronologies are similar in length to those in our study for the NM, NL, SM, and SL sites. The mean ring width found in the NH site suggests that environmental conditions at this high elevation (2400 m.a.s.l.) are less stressful for the growth of A. jaliscana, resulting in many large trees with dbh > 1.0 m, but with middle age and younger than those in the other sites; similarly, the NH site had the second shortest chronology (Table 2). The value of the signal-to-noise ratio was higher in NM compared to the other sites, which suggests that fir trees on this site show a high environmental signal in the annual growth rings due to a high-stress condition. Similarly, the highest values of EPS and RBAR were found in the NM site, and only in this stand, the EPS was ≥0.85, which corroborates the strong climatic signal. When a small sample depth corresponds with an EPS > 0.85 and the RBAR value is high and constant along the tree-ring series, it is an indication of the excellent potential of the species for climate reconstruction. On the other hand, an EPS < 0.85 may represent a limiting factor for proper reconstruction of climate variability, given that the trees may not be sensitive enough to capture the dominant climate signal of the area. This situation could be attributed to different causes, such as low sample depth, high frequency of complacent rings, or trees with irregular growth due to the presence of buttress. Increasing sample depth or selecting individuals in moisture-limited sites could be an option to improve the EPS value. Possibly, climate conditions and competition with other canopy and understory species have a negative impact on the growth of firs at this site, whereas in the south aspect (site SL) the climatic signal is weak, even though the forest is codominated with cloud-forest elements (Acer, Carpinus, Juglans, and Ostrya), which represent a strong competition. At this site, the ravine–stream condition may induce differences in the microenvironment, reducing the response to climatic variability and resulting in relatively low SNR and MS (trees that inhabit this condition can be considered as complacent). In general, montane temperate-humid belts allow a greater increase in fir radial growth compared to that observed in subalpine forest belts [7,45].
Although not a statistically significant result, the radial growth of A. jaliscana increases in relation to elevation. At the low-, mid-, and high-elevation distribution areas in both aspects, precipitation in the winter and early spring of the current growing season showed a significant positive correlation with the ring-width indices of A. jaliscana. The result is consistent with previous studies in subtropical Mexico, indicating that precipitation, particularly in the winter-spring season (driest period in this region), is the main climatic factor influencing radial growth in temperate-like environments [11,12,13,15]. In the Mediterranean basin, a similar response was reported for Abies borisii-regis and A. cephalonica in the dry season (late spring-summer) [4,46]. The positive correlation of winter-spring precipitation (driest months of the year) with tree growth reflects the reduction in water stress; this pattern could also be related to earlier vascular cambium activity, reducing frost damage, which allows tissue formation and shoot growth. In contrast, we found a negative correlation between the previous precipitation of July and September (the wettest months of the year in this region) and tree growth; this response could be related to saturated soils and a lack of oxygen for physiological processes, even though most of the rainfall is lost by runoff when the soil infiltration capacity is exceeded [8]. This response is consistent with what has been observed in other high-elevation forests in Mexico [8,47]; however, in other similar forests, such as those dominated by A. durangensis and A. religiosa, the correlation of the ring-width chronologies of these two species with the summer rain is not significant [15,20]. In the Mediterranean basin, a similar response was reported for Abies cephalonica, where the humid season (autumn-winter) showed a negative correlation with tree growth [46].
In both aspects (except SM and SL), mean maximum temperatures in the winter and spring of the current growing season show significant negative correlations with the ring-width chronology of A. jaliscana. The result is partially consistent with other climate-growth relationships observed in conifers such as A. durangensis, A. guatemalensis, Picea chihuahuana, and Pinus hartwegii, and highly consistent with A. religiosa in the MBBR [8,15,20,40,48]. This correlation between tree growth and temperature could be associated with high evapotranspiration rates, due to increased winter-spring temperatures, reducing annual radial growth [48].
The positive relationship between radial growth and mean maximum temperatures of the middle summer–autumn season (August–December), is not consistent with most climate-growth relationships observed in conifer forests mentioned above. Similarly, the negative correlation between minimum mean temperatures in February of the current year and fir tree growth is not consistent with most climate-growth relationships observed in montane and subalpine forests [1,6,11,48]. The latter correlation may indicate that winter minimum temperatures in the warmest years (which could be due to wide climate variability in February) limit the tree growth. Consequently, the coolest years favor the radial growth of A. jaliscana, which could be related to low evaporation rates and more water availability. However, late frosts that occur in late winter and early spring (February and March) could affect the growth of the trees; for instance, a snowfall in 2016 caused necrosis in foliar shoots, but this would not necessarily have affected the growth.
The decrease in growth of Abies jaliscana at the beginning of the 21st century and its subsequent increase in the last recent years (2013–2017) (Figure 3 and Figure 4) could be related to global climate change, specifically to the interaction of the continuous increase in global mean surface temperature in the last four decades, and the sudden increase in global land precipitation in the 2010–2015 period after a decrease in 2000–2007 [49]. Furthermore, the climatological stations closest to the populations of A. jaliscana show an increase in 146 mm in mean annual precipitation for the period 2011–2019, compared to the decade 2001–2010 [38,50]. In this study, the growth of Abies jaliscana was mainly negatively correlated with temperature and positively correlated with precipitation, which supports the above statement about relations between growth and climatic change. It has been shown that the increase in temperature can affect negatively the growth of coniferous species adapted to temperate-cold and humid climates [2,3,4,11,51], but if this warming is accompanied by an increase in precipitation, the effect on growth can change and even could be positive, especially for species that require significant humidity throughout the year, such as Abies jaliscana. This is similar to the growth response of Abies alba to climate change in Europe: in the southwestern areas of its range where aridity has increased due to global climate change its growth has diminished; on the other hand, in more temperate and humid areas warming is enhancing growth [52]. This pattern has been observed in another way along an elevation gradient for two coniferous species in north-central China, where low-elevation trees (exposed to a relatively higher temperature and lower precipitation) have decreased their growth in response to global warming, whereas high-elevation trees (exposed to more humid environments) have increased their growth [53].
The regional climate change projections for the western mountains in Jalisco (A. jaliscana habitat) for the year 2050 predict a warming rate in the range of 1 to 2.5 °C, whereas spring-summer precipitation will decrease from −10 to 0 mm and autumn-winter precipitation will decrease between 10 and 50 mm [54,55]. According to our estimates, this predicted increase in temperature and decrease in precipitation will affect negatively the radial growth of A. jaliscana; furthermore, the current anthropogenic threats such as illegal logging, forest fires, increase in pests, and changes in land use will synergize with climate change, increasing the risk of extinction that this species already undergoes [17,56]. The distribution area of A. jaliscana represents an archipelago of temperate mountains (“sky islands”) isolated between valleys and ravines with a warm climate [17,26], so global warming will reduce the area of suitable habitat for this species and further isolate its populations, thus preventing its capacity for horizontal-latitudinal migration. Migration along the elevational gradient would also be profoundly affected by climate change since the western mountains where this species inhabits do not reach elevations > 2450 m, so this species cannot migrate to higher elevations. Owing to these migration difficulties, the increase in temperature and decrease in precipitation that are expected in the future will notably affect the growth and survival of A. jaliscana fir populations.
Our results suggest that the observed precipitation patterns and their relation with ring-width indices might be dependent on regional climatic conditions. In addition, the six studied sites were similarly affected by the winter-spring precipitation fluctuations. However, differences in correlation values between temperature and radial growth at the north and south aspects are due to changes in microenvironment and microclimatic conditions in the mountains of western Jalisco, where the southern aspect is slightly wetter and warmer than the northern aspect (Table 1). The high incidence of fog above 2000 m in these forests [17], could cause slight changes in the regional thermal pattern. Although the growth of the fir species in the TMVB has been considered sensitive to temperature variations, our results show that precipitation is the major limiting factor positively affecting the radial growth of Abies jaliscana either in the low-, mid-, and high-elevation sites. In general, these results are consistent with previous reports in other conifers [8,19,48], showing that winter and spring precipitation is the primary climatic factor associated with the annual growth variability, as it is the case for A. jaliscana in the pluviseasonal climate region. Under these premises, if climate fluctuations lead to drier conditions, we anticipate a reduction in radial growth, which will cause a decline in the stem hydraulic conductivity and negative feedback on forest growth and productivity [48]. On the other hand, rainy conditions between November and April would favor tree radial growth; however, increases in temperature due to climate change may reduce the positive effect of precipitation.
5. Conclusions
Jalisco fir (A. jaliscana) shows distinct radial growth patterns in response to the climatic variability along an elevational gradient in the mountains of western Jalisco, Mexico. Similarly, it shows different radial growth in response to seasonal winter-spring precipitation variability, and mean monthly maximum temperature, due to elevational changes.
In the low, mid, and high elevations in both (north and south) aspects, winter-early spring precipitation of the current year shows a positive correlation with annual radial growth, while the mean maximum temperature in the current winter-spring season is the main factor affecting negatively ring width for most of the sites (except SL and SM). Increases in maximum temperature could limit the positive effects of cool seasonal precipitation and produce a reduction in growth, and decrease the relict populations of A. jaliscana. According to the increase in temperature and decrease in precipitation that are predicted for the future in the most probable scenarios of global and regional climate change, the growth of A. jaliscana fir and its suitable habitat will be negatively affected, which will significantly increase the risk of extinction of this species.
The present study provides new data on the radial growth response of Mesoamerican fir species to regional climatic fluctuations, representing the first research on elevational gradients and climatic factors that influence the annual radial growth of Mexican fir species. Similarly, it contributes to a better understanding of how precipitation, temperature, and microenvironmental variables may impact the growth of conifer species that thrive in high-elevation areas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13070981/s1, Figure S1. Mean sensitivity and standard deviation variations in Abies jaliscana tree-ring growth along altitudinal gradients of the mountains in western Jalisco, Mexico.
Author Contributions
Conceptualization, R.G.-H.; Data curation, R.G.-H. and J.V.-D.; Formal analysis, R.G.-H., J.V.-D., G.H.-V. and J.A.R.-C.; Funding acquisition, M.Á.M.-C.; Investigation, R.G.-H.; Methodology, R.G.-H. and J.V.-D.; Project administration, M.Á.M.-C.; Resources, M.Á.M.-C. and J.V.-D.; Software, J.V.-D.; Supervision, M.Á.M.-C.; Writing—original draft, R.G.-H.; Writing—review and editing, M.Á.M.-C., J.V.-D., G.H.-V., J.A.V.-G. and J.A.R.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The first author thanks the Doctoral Postgraduate Program in Biosystematics, Ecology, Natural and Agricultural Resources Management (BEMARENA) of the Universidad de Guadalajara for the support to conduct his graduate studies, and the CONACYT (Consejo Nacional de Ciencia y Tecnología) for the postgraduate scholarship provided (449479). Many thanks to the community “Ejido Cumbre de Guadalupe” and Modesto Peña for allowing access to his land properties. Carlos Verduzco, Dante Figueroa, and Rodrigo Mantilla contributed with sample processing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liang, E.; Wang, Y.; Xu, Y.; Liu, B.; Shao, X. Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan Plateau. Trees 2010, 24, 363–373. [Google Scholar] [CrossRef]
- Dang, H.; Jiang, M.; Zhang, Q.; Zhang, Y. Growth responses of subalpine fir (Abies fargesii) to climate variability in the Qinling Mountain, China. For. Ecol. Manag. 2007, 240, 143–150. [Google Scholar] [CrossRef]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Interacting effects of changes in climate and forest cover on mortality and growth of the southernmost European fir forests. Glob. Ecol. Biogeogr. 2009, 18, 485–497. [Google Scholar] [CrossRef]
- Pasho, E.; Toromani, E.; Alla, A.Q. Climatic impact on tree-ring widths in Abies borisii-regis forests from South-East Albania. Dendrochronologia 2014, 32, 237–244. [Google Scholar] [CrossRef]
- Fritts, H.C.; Smith, D.G.; Cardis, J.W.; Budelsky, C.A. Tree-ring characteristics along a vegetation gradient in northern Arizona. Ecology 1965, 46, 393–401. [Google Scholar] [CrossRef]
- Kharal, D.K.; Thapa, U.K.; George, S.S.; Meilby, H.; Rayamajhi, S.; Bhujo, D.R. Tree-climate relations along an elevational transect in Manang Valley, central Nepal. Dendrochronologia 2017, 41, 57–64. [Google Scholar] [CrossRef]
- Splechtna, B.E.; Dobry, J.; Klinka, K. Tree-ring characteristics of subalpine fir (Abies lasiocarpa (Hook) Nutt.) in relation to elevation and climatic fluctuations. Ann. For. Sci. 2000, 57, 89–100. [Google Scholar] [CrossRef]
- Astudillo-Sánchez, C.; Villanueva-Díaz, J.; Endara-Agramont, A.R.; Nava-Bernal, G.E.; Gómez-Albores, M.A. Climatic variability at the treeline of Monte Tlaloc, Mexico: A dendrochronological approach. Trees 2017, 31, 441–453. [Google Scholar] [CrossRef]
- Peng, J.; Gou, X.; Chen, F.; Li, J.; Liu, P.; Zhang, Y. Altitudinal variability of climate-tree growth relationships along a consistent slope of Anyemaqen Mountains, northeastern Tibetan Plateau. Dendrochronologia 2008, 26, 87–96. [Google Scholar] [CrossRef]
- Yang, B.; Minhui, M.; Melvin, T.M.; Zhao, Y.; Briffa, K.R. Climate control on tree growth at the upper and lower treelines: A case study in the Qilian Mountains, Tibetan Plateau. PLoS ONE 2013, 8, e69065. [Google Scholar] [CrossRef]
- González-Elizondo, M.; Jurado, E.; Navar, J.; González-Elizondo, M.S.; Villanueva-Díaz, J.; Aguirre, O.; Jiménez, J. Tree-rings and climate relationships for Douglas-fir chronologies from the Sierra Madre Occidental, Mexico: A 1681–2001 rain reconstruction. For. Ecol. Manag. 2005, 213, 39–53. [Google Scholar] [CrossRef]
- Villanueva-Diaz, J.; Stahle, D.W.; Luckman, B.H.; Cerano-Paredes, J.; Therrell, M.D.; Cleaveland, M.K.; Cornejo-Oviedo, E. Winter-spring precipitation reconstructions from tree rings for northeast Mexico. Clim. Chang. 2007, 83, 117–131. [Google Scholar] [CrossRef]
- Cerano-Paredes, J.; Villanueva-Díaz, J.; Cervantes-Martínez, R.; Vázquez-Selem, L.; Trucios-Caciano, R.; Guerra de la Cruz, V. Winter-spring precipitation reconstruction for the Pico de Tancítaro National Park, Michoacan. Investig. Geogr. Bol. Inst. Geogr. 2014, 83, 41–54. [Google Scholar]
- Villanueva-Díaz, J.; Cerano-Paredes, J.; Fulé, J.P.Z.; Cortés-Montaño, C.; Vázquez-Selem, L.; Yocom, L.L.; Ruiz-Corral, J.A. Four centuries of reconstructed hydroclimatic variability for Northwestern Chihuahua, Mexico, based in tree rings. Investig. Geogr. Bol. Inst. Geogr. 2015, 87, 141–153. [Google Scholar]
- González-Cásares, M.; Pompa-García, M.; Venegas-González, A. Climate signals from intra-annual wood density fluctuations in Abies durangensis. IAWA J. 2019, 40, 276–287. [Google Scholar] [CrossRef]
- Takahashi, K.; Azuma, H.; Yasue, K. Effects of climate on the radial growth of tree species in the upper and lower distribution limits of an altitudinal ecotone on Mount Norikura, central Japan. Ecol. Res. 2003, 18, 549–558. [Google Scholar] [CrossRef]
- Guerrero-Hernández, R.; Muñiz-Castro, M.A.; Vázquez-García, J.A.; Ruiz-Corral, A.; Hernández-Vera, G. Vegetation structure, classification and climatic characteristics in temperate-like mountain forests dominated by Abies jaliscana in western Mexico. Kast. Univ. J. For. Fac. 2019, 19, 235–258. [Google Scholar]
- Vázquez-García, J.A.; Shalisko, V.; Cuevas-Guzmán, R.; Muñiz-Castro, M.A.; Mantilla-Blandón, M.R. Abies jaliscana (Pinaceae): A new combination in section Grandis and a key to the species of Abies in western Mexico. Phytotaxa 2014, 183, 27–36. [Google Scholar] [CrossRef][Green Version]
- Carlón-Allende, T.; Mendoza, M.E.; Pérez-Salicrup, D.R.; Villanueva-Díaz, J.; Lara, A. Climatic responses of Pinus pseudostrobus and Abies religiosa in the Monarch Butterfly Biosphere Reserve, Mexico. Dendrochronologia 2016, 38, 103–116. [Google Scholar] [CrossRef]
- Carlón-Allende, T.; Villanueva-Díaz, J.; Mendoza, M.E.; Pérez-Salicrup, D.R. Climatic signal in earlywood and latewood in conifer forests in the Monarch Butterfly Biosphere Reserve, Mexico. Tree Ring Res. 2018, 74, 63–75. [Google Scholar] [CrossRef]
- Aquino-Ramírez, M.; Velázquez-Martínez, A.; Villanueva-Díaz, J.; Hervert-Zamora, H.L.; Gómez-Guerrero, A.; Reyes-Hernández, V.J.; Ramírez-Valverde, G. Climatic response of Abies guatemalensis Rehder in Ixtlán de Juárez, Oaxaca, Mexico. Madera Bosques 2019, 25, e2521773. [Google Scholar]
- Villanueva-Díaz, J.; Martínez-Sifuentes, A.R.; Rubio-Camacho, E.A.; Chávez-Durán, A.; Benavides-Solorio, J.D.D.; Cerano-Paredes, J.; Estrada-Ávalos, J. Dendrohydrological reconstruction of the streamflow of the Coahuayana hydrological sub-basin, Jalisco State. Rev. Mex. Cienc. For. 2021, 12, 157–181. [Google Scholar]
- Sáenz-Ceja, J.E.; Pérez-Salicrup, D.R. Modification of fire regimes inferred from the age structure of two conifer species in a Tropical Montane Forest, Mexico. Forests 2020, 11, 1193. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- SMN. Servicio Meteorológico Nacional. 2018. Available online: https://smn.conagua.gob.mx/es/informacion-climatologica-por-estado?estado=jal (accessed on 5 April 2018).
- Guerrero-Hernández, R.; González-Gallegos, J.G.; Castro-Castro, A. Análisis florístico de un bosque de Abies y el bosque mesófilo de montaña adyacente en Juanacatlán, Mascota, Jalisco, México. Bot. Sci. 2014, 92, 541–562. [Google Scholar] [CrossRef]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen, 5th ed.; Instituto de Geografía, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2004. [Google Scholar]
- Holdridge, L. Determination of world plant formations from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A. Worldwide Bioclimatic Classification System. Glob. Geobot. 2011, 1, 1–634. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago: Chicago, IL, USA, 1968; p. 73. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010; pp. 186–209. [Google Scholar]
- Cook, E.R. The decomposition of tree-ring series for environmental studies. Tree Ring Bull. 1987, 47, 37–59. [Google Scholar]
- Cook, E.R.; Peters, K. The smoothing spline: A new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree Ring Bull. 1981, 41, 45–53. [Google Scholar]
- Fritts, H.C.; Mosimann, J.E.; Bottorff, C.P. A revised computer program for standardizing tree ring series. Tree Ring Bull. 1969, 29, 15–20. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976; p. 567. [Google Scholar]
- Briffa, K.R.; Jones, P.D. Basic chronology statistics and assessment. In Methods of Dendrochronology: Applications in the Environmental Sciences; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 137–152. [Google Scholar]
- CLICOM. Base de Datos Climatológica Nacional. Available online: http://clicom-mex.cicese.mx/ (accessed on 21 November 2020).
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Anchukaitis, K.J.; Taylor, M.J.; Leland, C.; Pons, D.; Martin-Fernandez, J.; Castellanos, E. Tree-ring reconstructed dry season is rainfall in Guatemala. Clim. Dyn. 2015, 45, 1537–1546. [Google Scholar] [CrossRef]
- Meko, D.M.; Touchan, R.; Villanueva-Díaz, J.; Griffin, D.; Woodhouse, C.A.; Castro, C.L.; Carillo, C.; Leavitt, S.W. Sierra San Pedro Mártir, Baja California, cool-season Precipitation reconstructed from earlywood width of Abies concolor tree rings. J. Geophys. Res. Biogeosci. 2013, 118, 1660–1673. [Google Scholar] [CrossRef]
- Cook, E.R.; Shiyatov, S.; Mazepa, V. Estimation of the mean chronology. In Methods of Dendrochronology: Application in the Environmental Sciences; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 123–132. [Google Scholar]
- Brower, L.P.; Slayback, D.A.; Jaramillo-López, P.; Ramírez, I.; Oberhauser, K.S.; Williams, E.H.; Fink, L.S. Illegal logging of 10 hectares of forest in the Sierra Chincua monarch butterfly overwintering area in Mexico. Am. Entomol. 2016, 62, 92–97. [Google Scholar] [CrossRef][Green Version]
- Whipple, S.A.; Dix, R.L. Age structure and Succesional Dynamics of a Colorado Subalpine Forest. Am. Midl. Nat. 1979, 101, 142–158. [Google Scholar] [CrossRef]
- Klinka, K.; Wang, Q.; Carter, R.E.; Chen, H.Y.H. Height growth-elevation relationships in subalpine forests of interior British Columbia. For. Chron. 1996, 72, 193–198. [Google Scholar] [CrossRef]
- Papadopoulos, A. Tree-ring patterns and climate response of Mediterranean fir populations in Central Greece. Dendrochronologia 2016, 40, 17–25. [Google Scholar] [CrossRef]
- Biondi, F.; Hartsough, P.C.; Galindo-Estrada, I. Daily weather and tree growth at the tropical treeline of North America. Arct. Antarct. Alp. Res. 2005, 37, 16–24. [Google Scholar] [CrossRef]
- Cabral-Alemán, C.; Pompa-García, M.; Acosta-Hernández, A.C.; Zúñiga-Vásquez, J.M.; Camarero, J.J. Earlywood and latewood widths of Picea chihuahuana show contrasting sensitivity to seasonal climate. Forests 2017, 8, 173. [Google Scholar] [CrossRef]
- Gulev, S.K.; Thorne, P.W.; Ahn, J.; Dentener, F.J.; Domingues, C.M.; Gerland, S.; Gong, D.; Kaufman, D.S.; Nnamchi, H.C.; Quaas, J.; et al. Changing State of the Climate System. In Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 287–422. [Google Scholar] [CrossRef]
- SEMARNAT-CONAGUA. Estaciones Climatológicas en Google Earth. Estación de Mascota. Estadística. 2022. Available online: https://smn.conagua.gob.mx/tools/RESOURCES/Estadistica/14096.pdf (accessed on 9 May 2022).
- Gaire, N.P.; Dhakal, Y.R.; Lekhak, H.C.; Bhuju, D.R.; Shah, S.K. Dynamics of Abies spectabilis in relation to climate change at the treeline ecotone in Langtang National Park. Nepal J. Sci. Technol. 2011, 12, 220–229. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct effects of climate warming on populations of silver fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, W.; Wang, M.; Kang, M.; Dong, M. Radial growth of two dominant montane conifer tree species in response to climate change in North-Central China. PLoS ONE 2014, 9, e112537. [Google Scholar] [CrossRef]
- Ruiz-Corral, J.A.; Medina-García, G.; Rodríguez-Moreno, V.M.; Sánchez-González, J.D.J.; Villavicencio-García, R.; Durán-Puga, N.; Grajeda-Grajeda, J.; García-Romero, G.E. Regionalization of climate change in Mexico. Rev. Mex. Cien. Agríc. 2016, 7, 2451–2464. [Google Scholar]
- Davidova-Belitskaya, V. Clima, cambio climático y sus perspectivas en Jalisco. Cap. II. In La Gestión Climática en Jalisco; Davidova-Belitskaya, V., Ed.; Editorial Universidad de Guadalajara: Guadalajara, Mexico, 2020; pp. 34–52. [Google Scholar]
- SEMARNAT. Protección Ambiental-Especies Nativas de México de Flora y Fauna Silvestres-Categorías de Riesgo y Especificaciones Para su Inclusión, Exclusión o Cambio—Lista de Especies en Riesgo, Secretaria de Medio Ambiente y Recursos Naturales; Norma Oficial Mexicana NOM-059-SEMARNAT-2010; Diario Oficial de la Federación: Mexico City, Mexico, 2010.





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).