
Dissolved Organic Carbon Flux Is Driven by Plant Traits More Than Climate across Global Forest Types

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Source
2.2. Statistical Analysis
3. Results
3.1. The Variation in DOC Fluxes in Coniferous and Broadleaved Forests
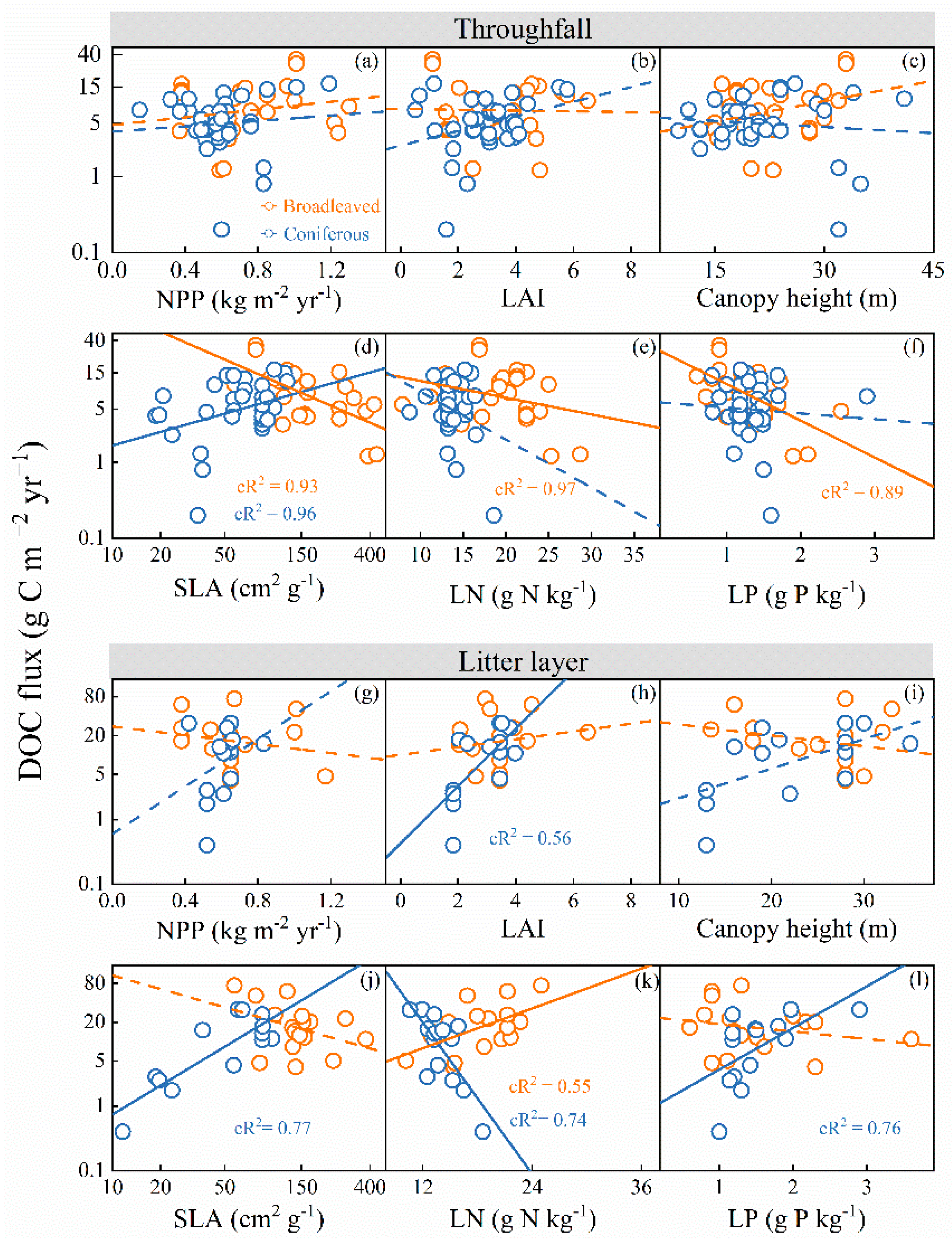
3.2. Effects of Climatic Variables on DOC Fluxes
3.3. Effects of Plant Traits on DOC Fluxes
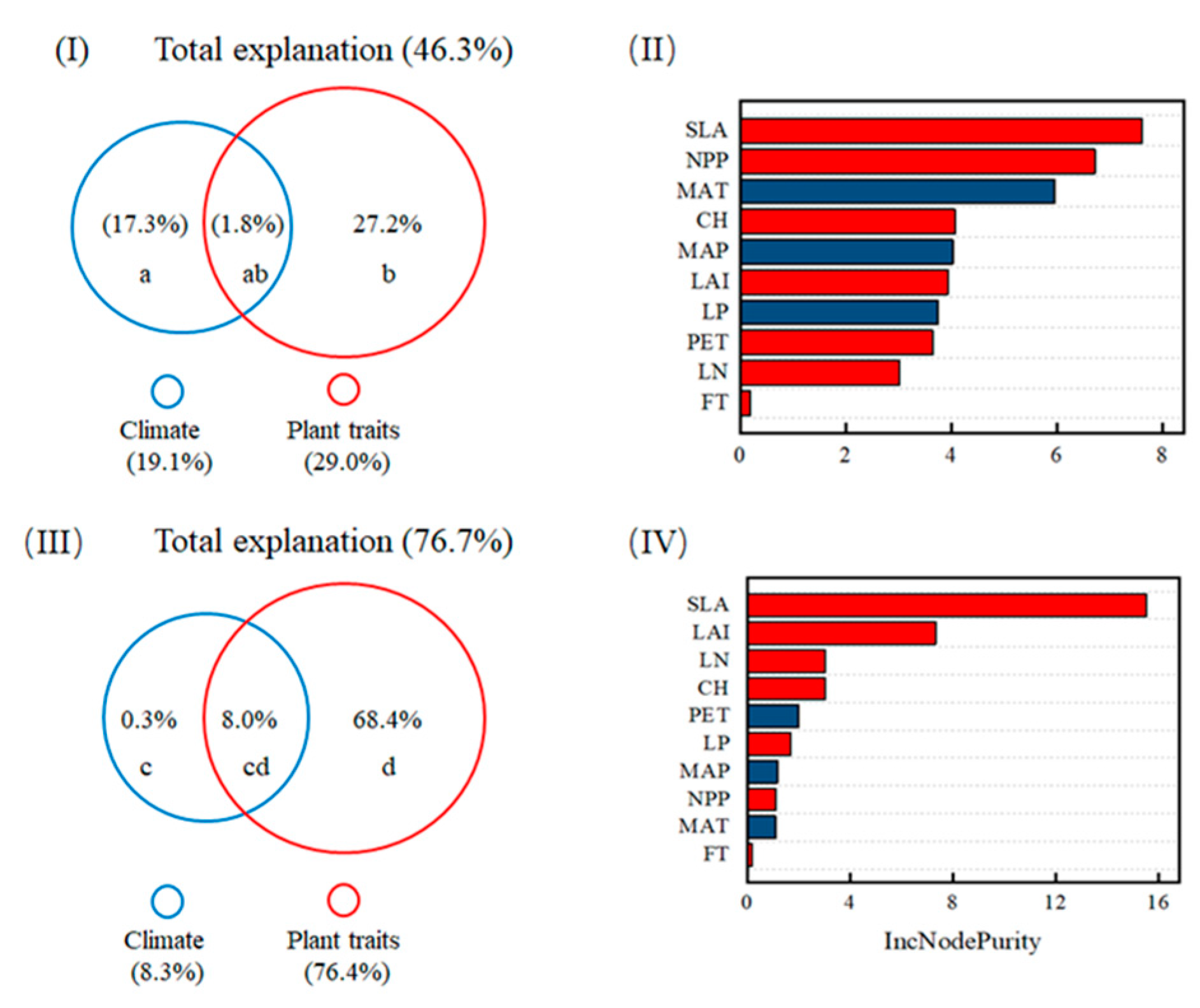
3.4. The Relative Contribution of Climate and Plant Traits to DOC Fluxes
4. Discussion
4.1. Differences in DOC Flux between Coniferous and Broadleaved Forests
4.2. The Dominant Role of Plant Traits in the Spatial Variability of DOC Fluxes Relative to Climate
4.3. Limitations and Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tranvik, L.J.; Jansson, M. Climate change—Terrestrial export of organic carbon. Nature 2002, 415, 861–862. [Google Scholar] [CrossRef]
- Kaiser, K.; Kalbitz, K. Cycling downwards—Dissolved organic matter in soils. Soil Biol. Biochem. 2012, 52, 29–32. [Google Scholar] [CrossRef]
- Wologo, E.; Shakil, S.; Zolkos, S.; Textor, S.; Ewing, S.; Klassen, J.; Spencer, R.G.M.; Podgorski, D.C.; Tank, S.E.; Baker, M.A.; et al. Stream dissolved organic matter in permafrost regions shows surprising compositional similarities but negative priming and nutrient effects. Glob. Biogeochem. Cycles 2021, 35, e2020GB006719. [Google Scholar] [CrossRef]
- Moore, T.R. Dissolved organic carbon in a northern boreal landscape. Glob. Biogeochem. Cycles 2003, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Battin, T.J.; Luyssaert, S.; Kaplan, L.A.; Aufdenkampe, A.K.; Richter, A.; Tranvik, L.J. The boundless carbon cycle. Nat. Geosci. 2009, 2, 598–600. [Google Scholar] [CrossRef]
- Pausch, J.; Kuzyakov, Y. Carbon input by roots into the soil: Quantification of rhizodeposition from root to ecosystem scale. Glob. Change Biol. 2018, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Keiser, A.D.; Davies, C.A.; Mersmann, C.A.; Strickland, M.S. Empirical evidence that soil carbon formation from plant inputs is positively related to microbial growth. Biogeochemistry 2013, 113, 271–281. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Change Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Michalzik, B.; Tipping, E.; Mulder, J.; Lancho, J.F.G.; Matzner, E.; Bryant, C.L.; Clarke, N.; Lofts, S.; Esteban, M.A.V. Modelling the production and transport of dissolved organic carbon in forest soils. Biogeochemistry 2003, 66, 241–264. [Google Scholar] [CrossRef]
- Wu, H.; Peng, C.; Moore, T.R.; Hua, D.; Li, C.; Zhu, Q.; Peichl, M.; Arain, M.A.; Guo, Z. Modeling dissolved organic carbon in temperate forest soils: TRIPLEX-DOC model development and validation. Geosci. Model Dev. 2014, 7, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Nakhavali, M.; Lauerwald, R.; Regnier, P.; Guenet, B.; Chadburn, S.; Friedlingstein, P. Leaching of dissolved organic carbon from mineral soils plays a significant role in the terrestrial carbon balance. Glob. Change Biol. 2021, 27, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Neff, J.C.; Asner, G.P. Dissolved organic carbon in terrestrial ecosystems: Synthesis and a model. Ecosystems 2001, 4, 29–48. [Google Scholar] [CrossRef] [Green Version]
- Uselman, S.M.; Qualls, R.G.; Lilienfein, J. Quality of soluble organic C, N, and P produced by different types and species of litter: Root litter versus leaf litter. Soil Biol. Biochem. 2012, 54, 57–67. [Google Scholar] [CrossRef]
- Michalzik, B.; Kalbitz, K.; Park, J.H.; Solinger, S.; Matzner, E. Fluxes and concentrations of dissolved organic carbon and nitrogen—A synthesis for temperate forests. Biogeochemistry 2001, 52, 173–205. [Google Scholar] [CrossRef]
- Arisci, S.; Rogora, M.; Marchetto, A.; Dichiaro, F. The role of forest type in the variability of DOC in atmospheric deposition at forest plots in Italy. Environ. Monit. Assess. 2012, 184, 3415–3425. [Google Scholar] [CrossRef]
- Currie, W.S.; Aber, J.D.; McDowell, W.H.; Boone, R.D.; Magill, A.H. Vertical transport of dissolved organic C and N under long-term N amendments in pine and hardwood forests. Biogeochemistry 1996, 35, 471–505. [Google Scholar] [CrossRef]
- Cuss, C.; Gueguen, C. Impacts of microbial activity on the optical and copper-binding properties of leaf-litter leachate. Front. Microbiol. 2012, 3, 166. [Google Scholar] [CrossRef] [Green Version]
- Prior, S.A.; Runion, G.B.; Rogers, H.H.; Torbert, H.A.; Reeves, D.W. Elevated atmospheric CO2 effects on biomass production and soil carbon in conventional and conservation cropping systems. Glob. Change Biol. 2005, 11, 657–665. [Google Scholar] [CrossRef]
- Kalbitz, K.; Meyer, A.; Yang, R.; Gerstberger, P. Response of dissolved organic matter in the forest floor to long-term manipulation of litter and throughfall inputs. Biogeochemistry 2007, 86, 301–318. [Google Scholar] [CrossRef]
- Lapierre, J.-F.; Seekell, D.A.; del Giorgio, P.A. Climate and landscape influence on indicators of lake carbon cycling through spatial patterns in dissolved organic carbon. Glob. Change Biol. 2015, 21, 4425–4435. [Google Scholar] [CrossRef]
- Dorrepaal, E.; Toet, S.; van Logtestijn, R.S.P.; Swart, E.; van de Weg, M.J.; Callaghan, T.V.; Aerts, R. Carbon respiration from subsurface peat accelerated by climate warming in the subarctic. Nature 2009, 460, 616–619. [Google Scholar] [CrossRef]
- Pries, C.E.H.; Castanha, C.; Porras, R.C.; Torn, M.S. The whole-soil carbon flux in response to warming. Science 2017, 355, 1420–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, F.; Joos, O. Experimental summer drought reduces soil CO2 effluxes and DOC leaching in Swiss grassland soils along an elevational gradient. Biogeochemistry 2014, 117, 395–412. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Reed, S.C.; Townsend, A.R. Nutrient regulation of organic matter decomposition in a tropical rain forest. Ecology 2006, 87, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Li, N.; Sun, X.; Hu, Z.; Bai, X.; Wang, G. Nitrogen addition reduces dissolved organic carbon leaching in a montane forest. Soil Biol. Biochem. 2018, 127, 31–38. [Google Scholar] [CrossRef]
- Gielen, B.; Neirynck, J.; Luyssaert, S.; Janssens, I.A. The importance of dissolved organic carbon fluxes for the carbon balance of a temperate Scots pine forest. Agric. For. Meteorol. 2011, 151, 270–278. [Google Scholar] [CrossRef]
- Mulholland, P.J. Large-scale patterns in dissolved organic carbon concentration, flux, and sources. In Aquatic Ecosystems; Findlay, S.E.G., Sinsabaugh, R.L., Eds.; Academic Press: Burlington, NJ, USA, 2003; pp. 139–159. [Google Scholar]
- Harrison, J.A.; Caraco, N.; Seitzinger, S.P. Global patterns and sources of dissolved organic matter export to the coastal zone: Results from a spatially explicit, global model. Glob. Biogeochem. Cycles 2005, 19, GB4S04. [Google Scholar] [CrossRef]
- Moatar, F.; Abbott, B.W.; Minaudo, C.; Curie, F.; Pinay, G. Elemental properties, hydrology, and biology interact to shape concentration-discharge curves for carbon, nutrients, sediment, and major ions. Water Resour. Res. 2017, 53, 1270–1287. [Google Scholar] [CrossRef]
- Raymond, P.A.; Saiers, J.E.; Sobczak, W.V. Hydrological and biogeochemical controls on watershed dissolved organic matter transport: Pulse-shunt concept. Ecology 2016, 97, 5–16. [Google Scholar] [CrossRef]
- Zarnetske, J.P.; Bouda, M.; Abbott, B.W.; Saiers, J.; Raymond, P.A. Generality of hydrologic transport limitation of watershed organic carbon flux across ecoregions of the United States. Geophys. Res. Lett. 2018, 45, 11702–11711. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Perez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- de la Riva, E.G.; Prieto, I.; Villar, R. The leaf economic spectrum drives leaf litter decomposition in Mediterranean forests. Plant Soil 2019, 435, 353–366. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Zak, D.R.; Burton, A.J.; Ashby, J.A.; MacDonald, N.W. Chronic nitrate additions dramatically increase the export of carbon and nitrogen from northern hardwood ecosystems. Biogeochemistry 2004, 68, 179–197. [Google Scholar] [CrossRef]
- Shi, S.; Yang, M.; Hou, Y.; Peng, C.; Wu, H.; Zhu, Q.; Liang, Q.; Xie, J.; Wang, M. Simulation of dissolved organic carbon concentrations and fluxes in Chinese monsoon forest ecosystems using a modified TRIPLEX-DOC model. Sci. Total Environ. 2019, 697, 20. [Google Scholar] [CrossRef]
- Kattge, J.; Sandel, B. TRY plant trait database—Enhanced coverage and open access (vol 2020, pg 119, 2020). Glob. Change Biol. 2020, 26, 5343. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Finegan, B.; Pena-Claros, M.; de Oliveira, A.; Ascarrunz, N.; Bret-Harte, M.S.; Carreno-Rocabado, G.; Casanoves, F.; Diaz, S.; Eguiguren Velepucha, P.; Fernandez, F.; et al. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. J. Ecol. 2015, 103, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Shao, J.; Liu, H.; Du, Z.; Zhou, L.; Liu, R.; Bernhofer, C.; Gruenwald, T.; Dusek, J.; Montagnani, L.; et al. Relative importance of climatic variables, soil properties and plant traits to spatial variability in net CO2 exchange across global forests and grasslands. Agric. For. Meteorol. 2021, 307, 15. [Google Scholar] [CrossRef]
- Lefsky, M.A. A global forest canopy height map from the moderate resolution imaging spectroradiometer and the geoscience laser altimeter system. Geophys. Res. Lett. 2010, 37, 15. [Google Scholar] [CrossRef] [Green Version]
- Running, S.W.; Nemani, R.R.; Heinsch, F.A.; Zhao, M.S.; Reeves, M.; Hashimoto, H. A continuous satellite-derived measure of global terrestrial primary production. Bioscience 2004, 54, 547–560. [Google Scholar] [CrossRef]
- Clifford, P.; Richardson, S.; Hemon, D. Assessing the significance of the correlation between two spatial processes. Biometrics 1989, 45, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Veen, G.F.; Bonis, A.; Bradford, E.M.; Classen, A.T.; Cornelissen, J.H.C.; Crowther, T.W.; De Long, J.R.; Freschet, G.T.; Kardol, P.; et al. A test of the hierarchical model of litter decomposition. Nat. Ecol. Evol. 2017, 1, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chen, H.Y.H.; Yue, C.; Gong, X.Y.; Shao, J.; Zhou, G.; Wang, J.; Wang, M.; Xia, J.; Li, Y.; et al. Traits mediate drought effects on wood carbon fluxes. Glob. Change Biol. 2020, 26, 3429–3442. [Google Scholar] [CrossRef]
- Sokal, R.R.J.B. The principles and practice of statistics in biological research. Biometry 1995, 451–554. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Hu, Z.; Michaletz, S.T.; Johnson, D.J.; McDowell, N.G.; Huang, Z.; Zhou, X.; Xu, C. Traits drive global wood decomposition rates more than climate. Glob. Change Biol. 2018, 24, 5259–5269. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Kuussaari, M.; Pöyry, J. New insights into butterfly–environment relationships using partitioning methods. Proc. R. Soc. B Boil. Sci. 2005, 272, 2203–2210. [Google Scholar] [CrossRef] [Green Version]
- Jian, J.; Steele, M.K.; Thomas, R.Q.; Day, S.D.; Hodges, S.C. Constraining estimates of global soil respiration by quantifying sources of variability. Glob. Change Biol. 2018, 24, 4143–4159. [Google Scholar] [CrossRef]
- Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Controls over leaf litter decomposition in wet tropical forests. Ecology 2009, 90, 3333–3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perakis, S.S.; Matkins, J.J.; Hibbs, D.E. Interactions of tissue and fertilizer nitrogen on decomposition dynamics of lignin-rich conifer litter. Ecosphere 2012, 3, 1–12. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, G.; Gallardo, J.F.; Bussotti, F. Canopy modification of atmospheric deposition in oligotrophic Quercus pyrenaica forests of an unpolluted region (central-western Spain). For. Ecol. Manag. 2001, 149, 47–60. [Google Scholar] [CrossRef]
- Fowler, R.A.; Osburn, C.L.; Saros, J.E. Climate-driven changes in dissolved organic carbon and water clarity in Arctic lakes of west Greenland. J. Geophys. Res. Biogeosci. 2020, 125, 2. [Google Scholar] [CrossRef]
- Prescott, C.E.J.B. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Deng, M.; Liu, L.; Jiang, L.; Liu, W.; Wang, X.; Li, S.; Yang, S.; Wang, B. Ecosystem scale trade-off in nitrogen acquisition pathways. Nat. Ecol. Evol. 2018, 2, 1724–1734. [Google Scholar] [CrossRef]
- Matulich, K.L.; Martiny, J.B.H. Microbial composition alters the response of litter decomposition to environmental change. Ecology 2015, 96, 154–163. [Google Scholar] [CrossRef]
- Buzzard, V.; Michaletz, S.T.; Deng, Y.; He, Z.; Ning, D.; Shen, L.; Tu, Q.; Van Nostrand, J.D.; Voordeckers, J.W.; Wang, J.J.; et al. Continental scale structuring of forest and soil diversity via functional traits. Nat. Ecol. Evol. 2019, 3, 1298–1308. [Google Scholar] [CrossRef]
- Cubino, J.P.; Biurrun, I.; Bonari, G.; Braslavskaya, T.; Font, X.; Jandt, U.; Jansen, F.; Rašomavičius, V.; Škvorc, Ž.; Willner, W.J.E. The leaf economic and plant size spectra of European forest understory vegetation. Ecography 2021, 44, 1311–1324. [Google Scholar] [CrossRef]
- Aitkenhead-Peterson, J.A.; Alexander, J.E.; Albrechtova, J.; Kram, P.; Rock, B.; Cudlín, P.; Hruška, J.; Lhotakova, Z.; Huntley, R.; Oulehle, F.J.P.; et al. Linking foliar chemistry to forest floor solid and solution phase organic C and N in Picea abies [L.] Karst stands in northern Bohemia. Plant Soil 2006, 283, 187–201. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Brant, A.N.; Seedre, M.; Brassard, B.W.; Taylor, A.R. The contribution of litterfall to net primary production during secondary succession in the Boreal forest. Ecosystems 2017, 20, 830–844. [Google Scholar] [CrossRef]
- Camino-Serrano, M.; Gielen, B.; Luyssaert, S.; Ciais, P.; Vicca, S.; Guenet, B.; De Vos, B.; Cools, N.; Ahrens, B.; Arain, M.A.; et al. Linking variability in soil solution dissolved organic carbon to climate, soil type, and vegetation type. Glob. Biogeochem. Cycles 2014, 28, 497–509. [Google Scholar] [CrossRef]
- He, N.; Liu, C.; Tian, M.; Li, M.; Yang, H.; Yu, G.; Guo, D.; Smith, M.D.; Yu, Q.; Hou, J. Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functions. Funct. Ecol. 2018, 32, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Kembel, S.W.; Mueller, R.C. Plant traits and taxonomy drive host associations in tropical phyllosphere fungal communities. Botany 2014, 92, 4. [Google Scholar] [CrossRef] [Green Version]
- Manning, D.W.P.; Rosemond, A.D.; Gulis, V.; Benstead, J.P.; Kominoski, J.S. Nutrients and temperature additively increase stream microbial respiration. Glob. Change Biol. 2018, 24, E233–E247. [Google Scholar] [CrossRef]
- Lustenhouwer, N.; Maynard, D.S.; Bradford, M.A.; Lindner, D.L.; Oberle, B.; Zanne, A.E.; Crowther, T.W. A trait-based understanding of wood decomposition by fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 11551–11558. [Google Scholar] [CrossRef]
- Williams, K.; Percival, F.; Merino, J.; Mooney, H.A. Estimation of Tissue Construction Cost from Heat of Combustion and Organic Nitrogen Content. Plant Cell Environ. 1987, 10, 725–734. [Google Scholar] [CrossRef]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [Green Version]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer Nature: Berlin, Germany, 2020. [Google Scholar]
- Hu, Z.; Xu, C.; McDowell, N.G.; Johnson, D.J.; Wang, M.; Luo, Y.; Zhou, X.; Huang, Z. Linking microbial community composition to C loss rates during wood decomposition. Soil Biol. Biochem. 2017, 104, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Weyhenmeyer, G.A.; Froberg, M.; Karltun, E.; Khalili, M.; Kothawala, D.; Temnerud, J.; Tranvik, L.J. Selective decay of terrestrial organic carbon during transport from land to sea. Glob. Change Biol. 2012, 18, 349–355. [Google Scholar] [CrossRef]
- Agren, G.I.; Kleja, D.B.; Bosatta, E. Modelling Dissolved organic carbon production in coniferous forest soils. Soil Sci. Soc. Am. J. 2018, 82, 1392–1403. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, S.; He, T.; Liu, L.; Wu, J. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Yu, X.; Dijkstra, F.A. Carbon and nitrogen dynamics affected by litter and nitrogen addition in a grassland soil: Role of fungi. Eur. J. Soil Biol. 2020, 100, 103211. [Google Scholar] [CrossRef]
- Spurr, S.H.; Barnes, B.V. Forest Ecology, 3rd ed.; John Wiley & Sons: North Miami Beach, FL, USA, 1980. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Throughfall | Model | Litter Layer | ||
|---|---|---|---|---|---|
| Variable | Coefficient | Variable | Coefficient | ||
| Full model | Intercept | 9.09 | Full model | Intercept | −0.13 |
| MAT | −1.54 * | MAT | 1.33 | ||
| MAP | 0.17 | MAP | −0.19 | ||
| SLA | −0.71 | LN | 0.10 ** | ||
| NPP | 0.31 | SLA | −0.35 | ||
| LN | −0.41 * | FT | −1.58 | ||
| FT | 0.51 * | LN × FT | 1.55 *** | ||
| LN × MAT | 0.12 * | SLA × FT | 0.33 *** | ||
| SLA × FT | 2.51 *** | mR2 | 76.7 | ||
| mR2 | 46.3 | ||||
| Climate model | Intercept | −1.37 | Climate model | Intercept | 1.48 |
| MAT | 0.79 * | MAT | −70 * | ||
| MAP | 0.12 | MAP | 2.00 | ||
| mR2 | 19.1 | mR2 | 8.3 | ||
| Traits model | Intercept | 5.71 | Traits model | Intercept | 2.49 |
| SLA | −0.75 * | LN | 0.12 ** | ||
| NPP | 0.75 * | SLA | −0.36 | ||
| LN | −0.03 | FT | −2.39 | ||
| FT | −0.71 *** | LN × FT | −0.37 *** | ||
| SLA × FT | 1.52 *** | SLA × FT | 1.79 *** | ||
| mR2 | 29.0 | mR2 | 76.4 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, Y.; He, Y.; Shao, J.; Liu, H.; Fu, Y.; Chen, X.; Chen, Y.; Liu, R.; Gao, J.; Li, N.; et al. Dissolved Organic Carbon Flux Is Driven by Plant Traits More Than Climate across Global Forest Types. Forests 2022, 13, 1119. https://doi.org/10.3390/f13071119
Ji Y, He Y, Shao J, Liu H, Fu Y, Chen X, Chen Y, Liu R, Gao J, Li N, et al. Dissolved Organic Carbon Flux Is Driven by Plant Traits More Than Climate across Global Forest Types. Forests. 2022; 13(7):1119. https://doi.org/10.3390/f13071119
Chicago/Turabian StyleJi, Yuhuang, Yanghui He, Junjiong Shao, Huiying Liu, Yuling Fu, Xinyue Chen, Yang Chen, Ruiqiang Liu, Jing Gao, Nan Li, and et al. 2022. "Dissolved Organic Carbon Flux Is Driven by Plant Traits More Than Climate across Global Forest Types" Forests 13, no. 7: 1119. https://doi.org/10.3390/f13071119
APA StyleJi, Y., He, Y., Shao, J., Liu, H., Fu, Y., Chen, X., Chen, Y., Liu, R., Gao, J., Li, N., Zhou, G., Zhou, L., & Zhou, X. (2022). Dissolved Organic Carbon Flux Is Driven by Plant Traits More Than Climate across Global Forest Types. Forests, 13(7), 1119. https://doi.org/10.3390/f13071119