
Soil Microbial and Organic Carbon Legacies of Pre-Existing Plants Drive Pioneer Tree Growth during Subalpine Forest Succession

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Seed Collection
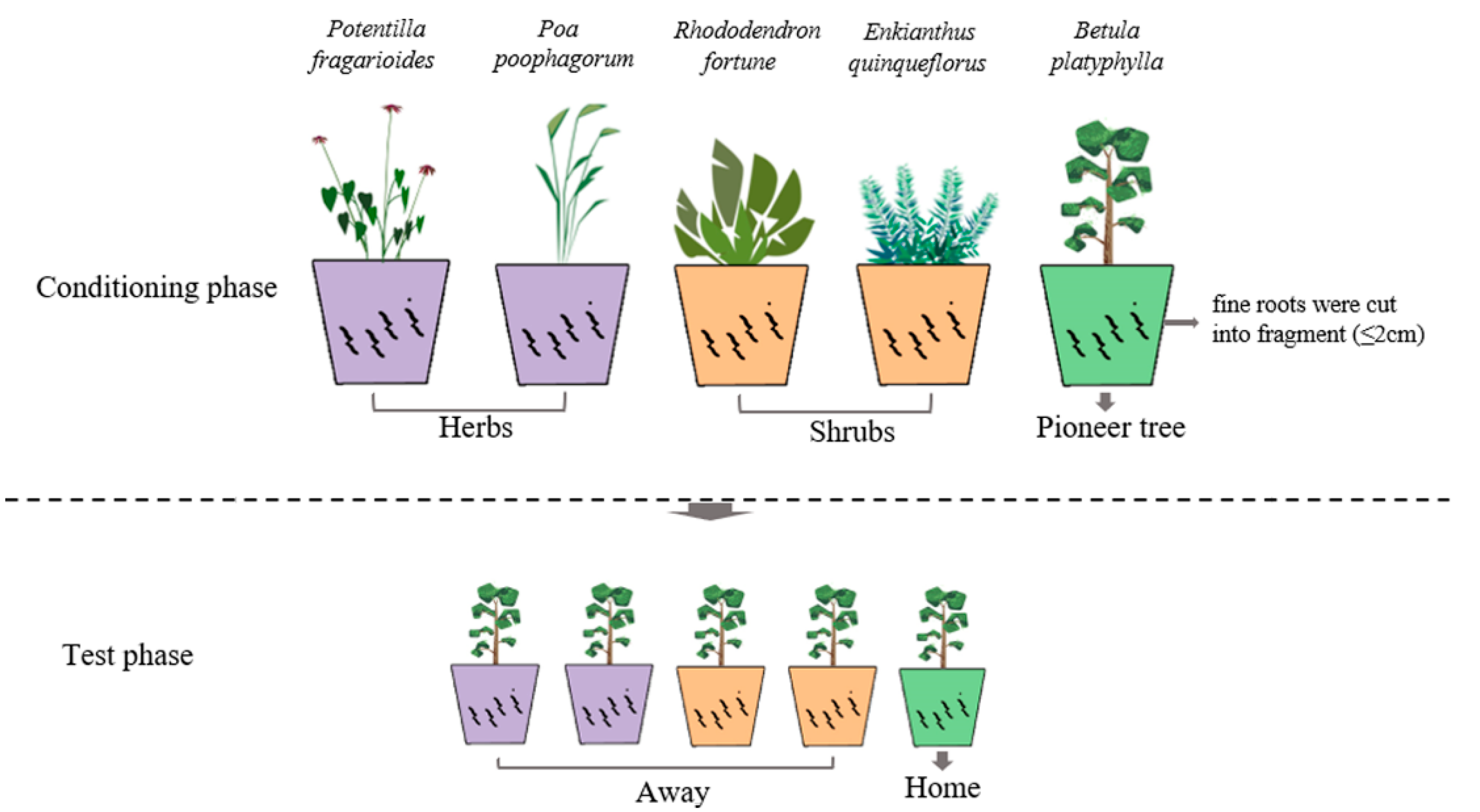
2.2. Greenhouse Experiment
2.2.1. Conditioning Phase
2.2.2. Test Phase
2.3. Soil DNA Extraction and Sequence Analyses
2.4. Soil Properties
2.5. Statistical Analysis
3. Results
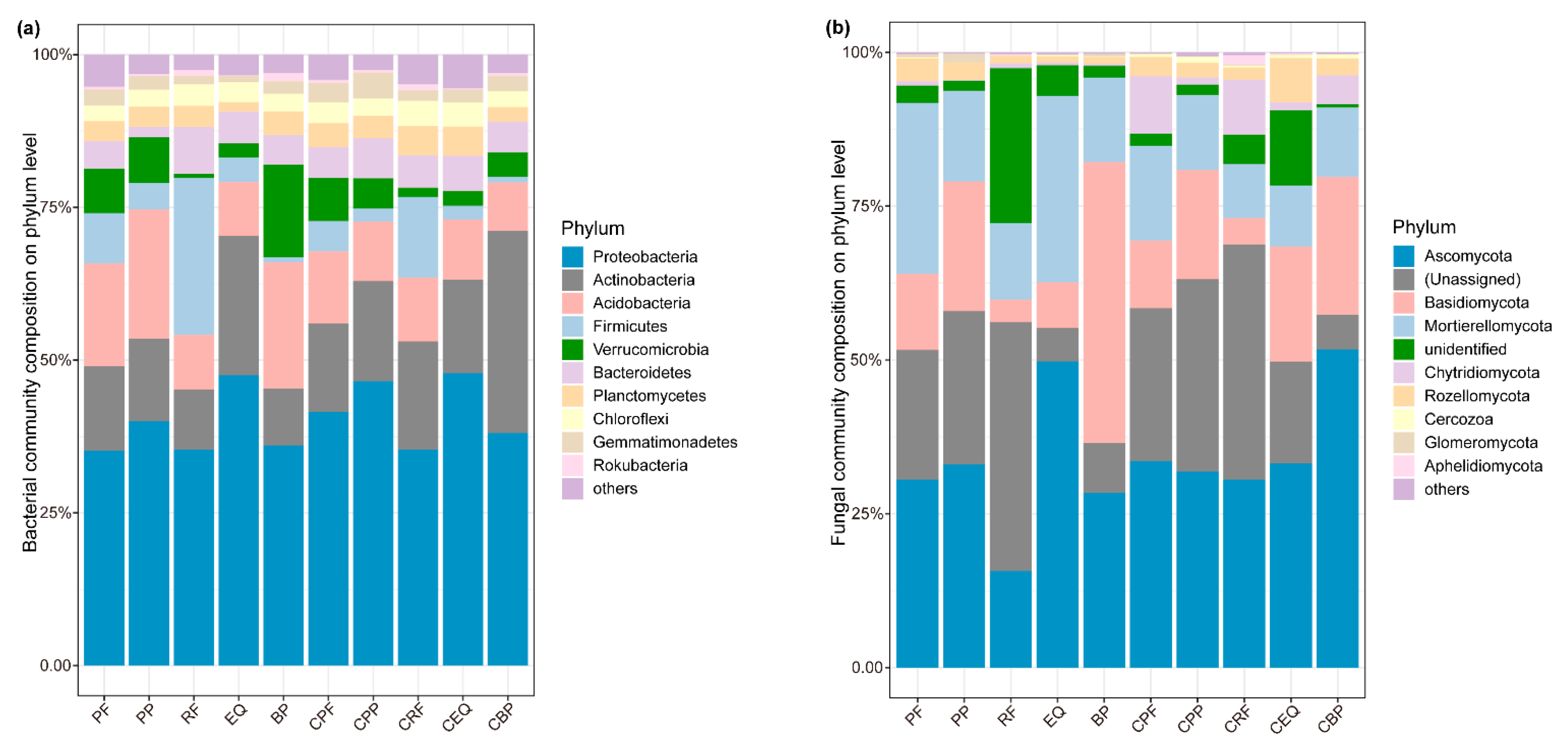
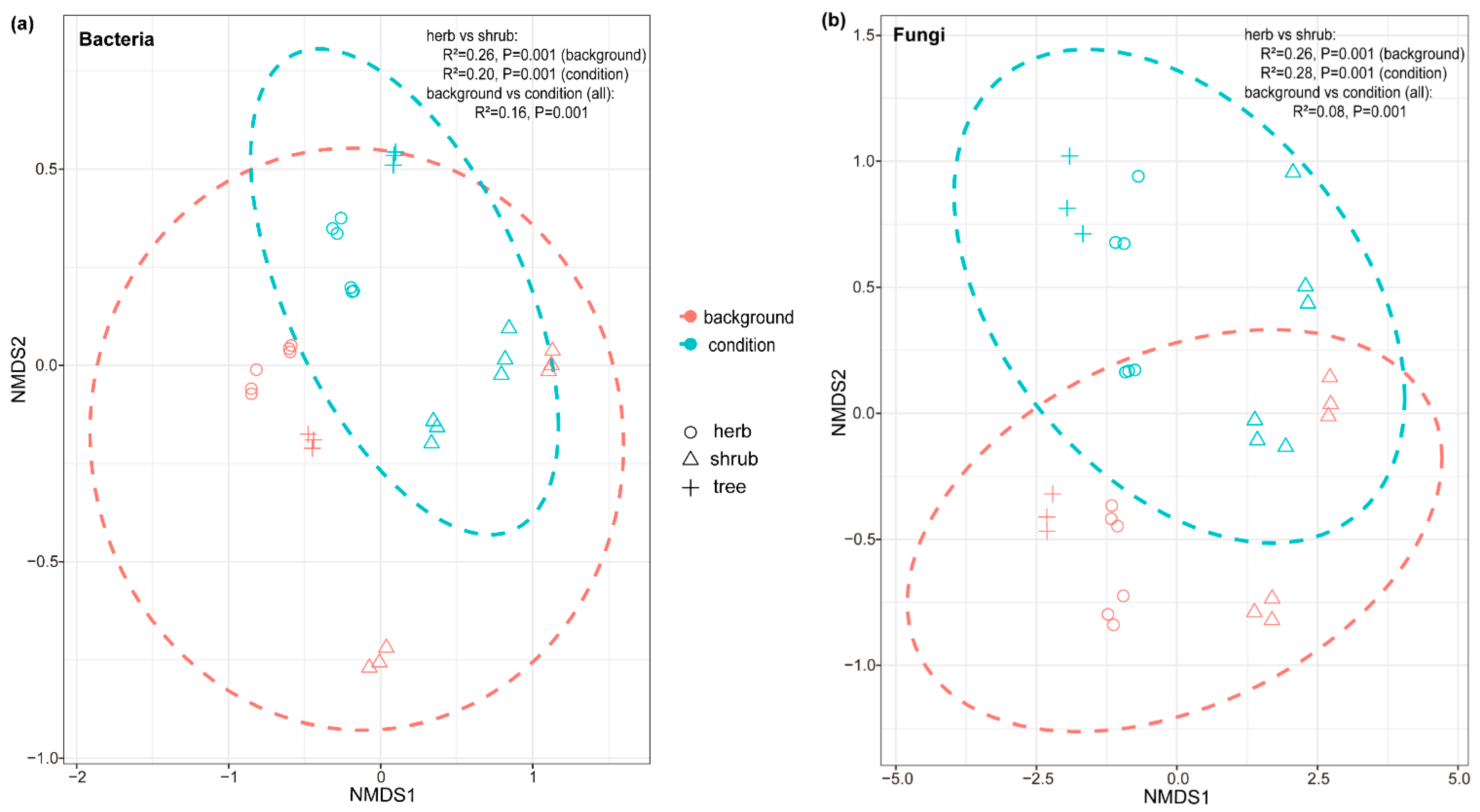
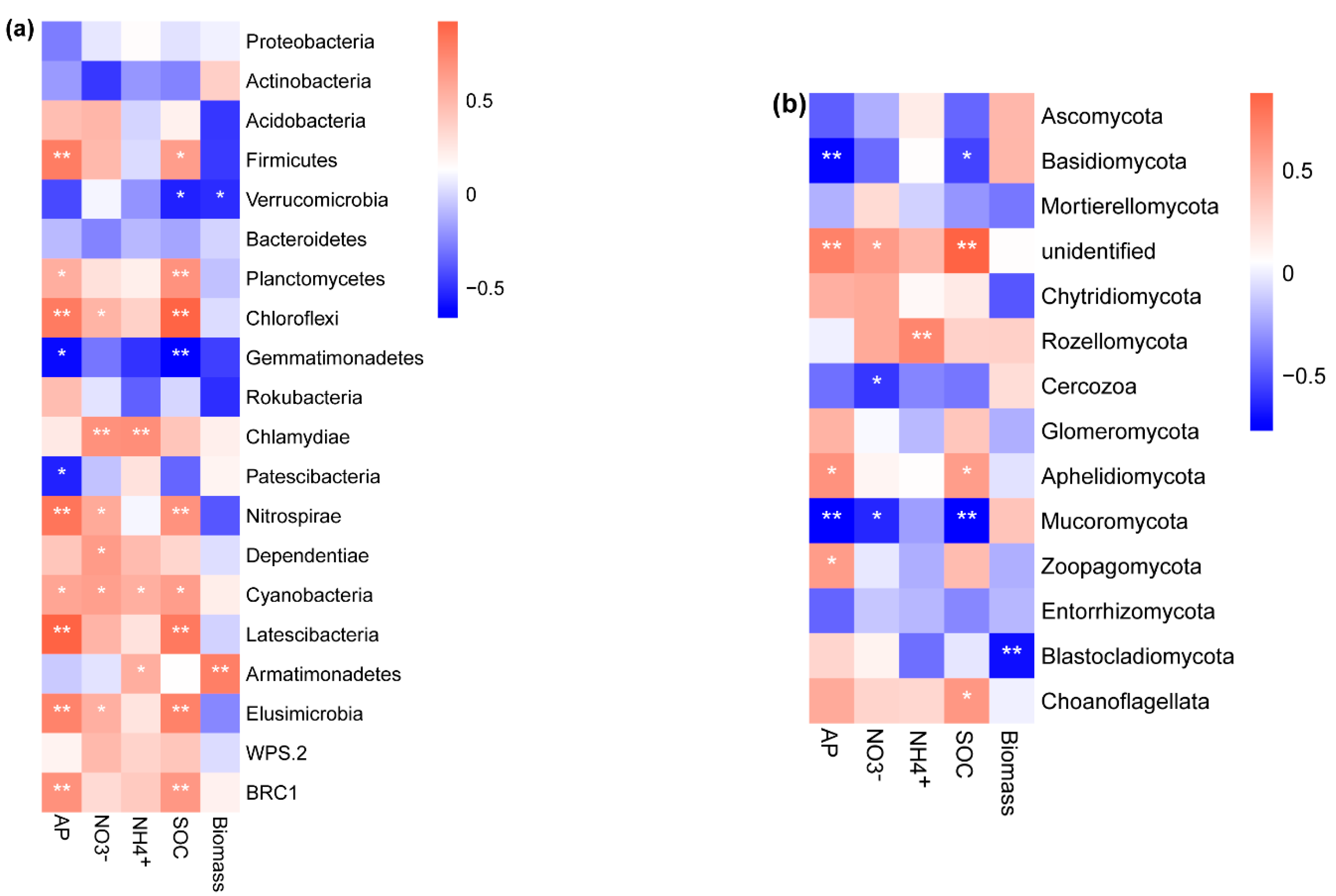
3.1. Soil Physicochemical Properties and Microbial Community Composition
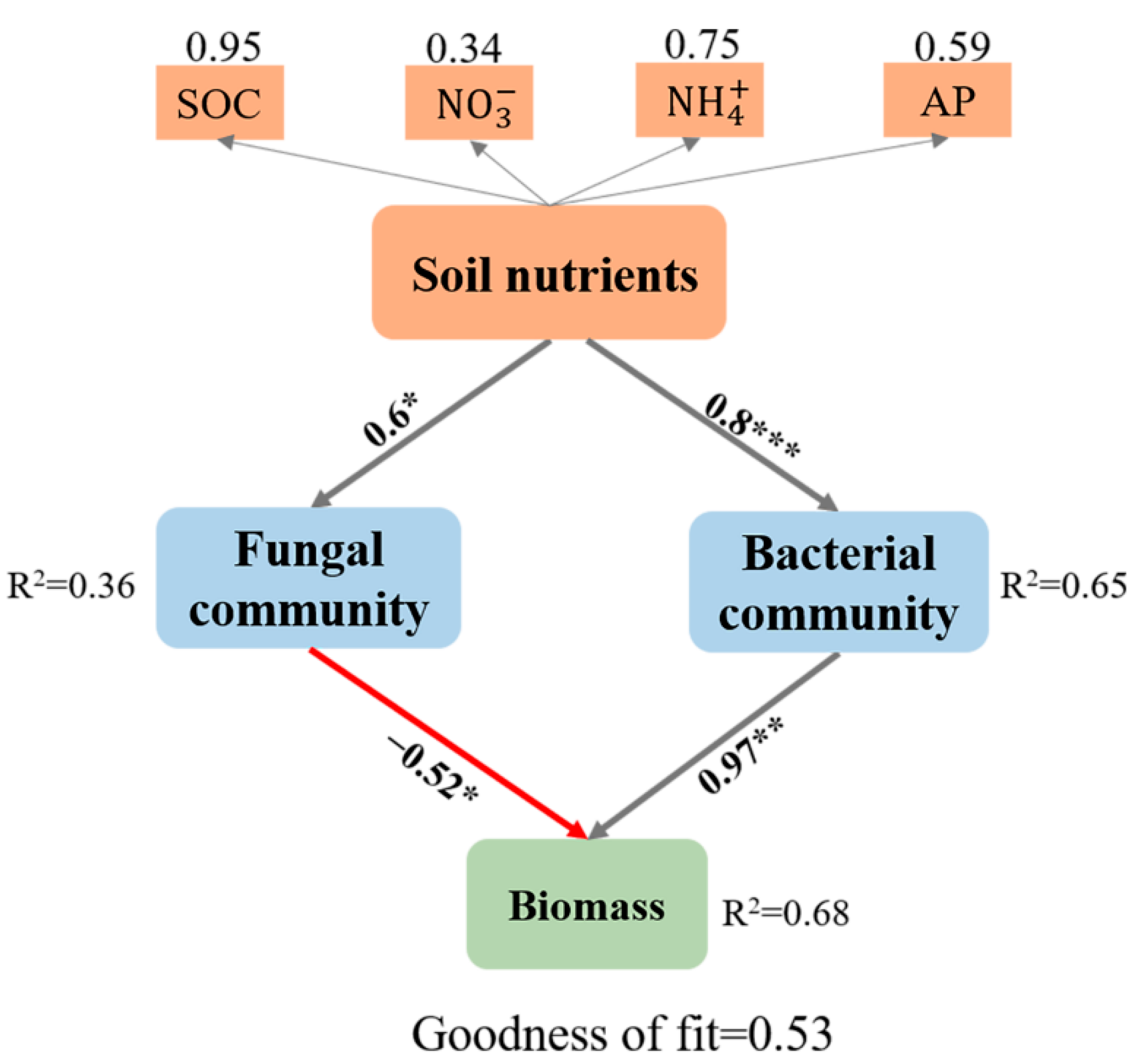
3.2. Linking Soil Biotic and Abiotic Legacies with Pioneer Tree Seedling Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gomez-Aparicio, L.; Zamora, R.; Gomez, J.M.; Hodar, J.A.; Castro, J.; Baraza, E. Applying plant facilitation to forest restoration: A meta-analysis of the use of shrubs as nurse plants. Ecol. Appl. 2004, 14, 1128–1138. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [Green Version]
- Mansourian, S.; Parrotta, J. From addressing symptoms to tackling the illness: Reversing forest loss and degradation. Environ. Sci. Policy 2019, 101, 262–265. [Google Scholar] [CrossRef]
- Harantová, L.; Mudrák, O.; Kohout, P.; Elhottová, D.; Frouz, J.; Baldrian, P. Development of microbial community during primary succession in areas degraded by mining activities. Land Degrad. Dev. 2017, 28, 2574–2584. [Google Scholar] [CrossRef]
- Gradel, A.; Haensch, C.; Ganbaatar, B.; Dovdondemberel, B.; Nadaldorj, O.; Günther, B. Response of white birch (Betula platyphylla Sukaczev) to temperature and precipitation in the mountain forest steppe and taiga of northern Mongolia. Dendrochronologia 2017, 41, 24–33. [Google Scholar] [CrossRef]
- Tiebel, K.; Huth, F.; Wagner, S. Soil seed banks of pioneer tree species in European temperate forests: A review. iForest 2018, 11, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Perala, D.A.; Alm, A.A. Reproductive ecology of birch: A review. Forest Ecol. Manag. 1990, 32, 1–38. [Google Scholar] [CrossRef]
- Ganade, G.; Miriti, M.N.; Mazzochini, G.G.; Paz, C.P. Pioneer effects on exotic and native tree colonizers: Insights for Araucaria forest restoration. Basic Appl. Ecol. 2011, 12, 733–742. [Google Scholar] [CrossRef]
- Pinard, M.; Howlett, B.; Davidson, D. Site conditions limit pioneer tree recruitment after logging of dipterocarp forests in Sabah, Malaysia. Biotropica 1996, 28, 2–12. [Google Scholar] [CrossRef]
- MacKenzie, W.H.; Mahony, C.R. An ecological approach to climate change-informed tree species selection for reforestation. Forest Ecol. Manag. 2021, 481, 118705. [Google Scholar] [CrossRef]
- Goodale, U.M.; Berlyn, G.P.; Gregoire, T.G.; Tennakoon, K.U.; Ashton, M.S. Differences in Survival and Growth Among Tropical Rain Forest Pioneer Tree Seedlings in Relation to Canopy Openness and Herbivory. Biotropica 2014, 46, 183–193. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 2009, 97, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Dalling, J.W.; Muller-Landau, H.C.; Wright, S.J.; Hubbell, S.P. Role of dispersal in the recruitment limitation of neotropical pioneer species. J. Ecol. 2002, 90, 714–727. [Google Scholar] [CrossRef]
- Xia, Q.; Ando, M.; Seiwa, K.; Kitajima, K. Interaction of seed size with light quality and temperature regimes as germination cues in 10 temperate pioneer tree species. Funct. Ecol. 2015, 30, 866–874. [Google Scholar] [CrossRef]
- Graves, J.H.; Peet, R.K.; White, P.S. The influence of carbon-nutrient balance on herb and woody plant abundance in temperate forest understories. J. Veg. Sci. 2006, 17, 217–226. [Google Scholar] [CrossRef]
- Skinner, A.K.; Lunt, I.D.; McIntyre, S.; Spooner, P.G.; Lavorel, S. Eucalyptus recruitment in degraded woodlands: No benefit from elevated soil fertility. Plant Ecol. 2009, 208, 359–370. [Google Scholar] [CrossRef]
- Molina, R.; Amaranthus, M. Rhizosphere biology: Ecological linkages between soil processes, plant-growth, and community dynamics. In Proceedings of the Conf on Management and Productivity of Western-Montane Forest Soils, Boise, ID, USA, 10–12 April 1990; pp. 51–58. [Google Scholar]
- Cuddington, K. Legacy Effects: The Persistent Impact of Ecological Interactions. Biol. Theor. 2012, 6, 203–210. [Google Scholar] [CrossRef]
- Peña, E.; Baeten, L.; Steel, H.; Viaene, N.; De Sutter, N.; De Schrijver, A.; Verheyen, K.; Bailey, J. Beyond plant-soil feedbacks: Mechanisms driving plant community shifts due to land-use legacies in post-agricultural forests. Funct. Ecol. 2016, 30, 1073–1085. [Google Scholar] [CrossRef] [Green Version]
- Heinen, R.; Hannula, S.E.; De Long, J.R.; Huberty, M.; Jongen, R.; Kielak, A.; Steinauer, K.; Zhu, F.; Bezemer, T.M. Plant community composition steers grassland vegetation via soil legacy effects. Ecol. Lett. 2020, 23, 973–982. [Google Scholar] [CrossRef]
- Endo, M.; Yamamura, Y.; Tanaka, A.; Nakano, T.; Yasuda, T. Nurse-Plant Effects of a Dwarf Shrub on the Establishment of Tree Seedlings in a Volcanic Desert on Mt. Fuji, Central Japan. Arct. Antarct. Alp. Res. 2008, 40, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Hannula, S.E.; Heinen, R.; Huberty, M.; Steinauer, K.; De Long, J.R.; Jongen, R.; Bezemer, T.M. Persistence of plant-mediated microbial soil legacy effects in soil and inside roots. Nat. Commun. 2021, 12, 5686. [Google Scholar] [CrossRef] [PubMed]
- Lekberg, Y.; Bever, J.D.; Bunn, R.A.; Callaway, R.M.; Hart, M.M.; Kivlin, S.N.; Klironomos, J.; Larkin, B.G.; Maron, J.L.; Reinhart, K.O.; et al. Relative importance of competition and plant-soil feedback, their synergy, context dependency and implications for coexistence. Ecol. Lett. 2018, 21, 1268–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyens, N.; van der Lelie, D.; Taghavi, S.; Newman, L.; Vangronsveld, J. Exploiting plant-microbe partnerships to improve biomass production and remediation. Trends Biotechnol. 2009, 27, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, L.M.; Nilsson, M.C.; Wardle, D.A.; Zackrisson, O. Context dependent effects of ectomycorrhizal species richness on tree seedling productivity. Oikos 2001, 93, 353–364. [Google Scholar] [CrossRef]
- Garcia-Guzman, G.; Heil, M. Life histories of hosts and pathogens predict patterns in tropical fungal plant diseases. New Phytol. 2014, 201, 1106–1120. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi-potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Heijden, M.G.A.; Verkade, S.; De Bruin, S.J. Mycorrhizal fungi reduce the negative effects of nitrogen enrichment on plant community structure in dune grassland. Global Chang. Biol. 2008, 14, 2626–2635. [Google Scholar] [CrossRef]
- Wei, G.; Li, M.; Shi, W.; Tian, R.; Chang, C.; Wang, Z.; Wang, N.; Zhao, G.; Gao, Z. Similar drivers but different effects lead to distinct ecological patterns of soil bacterial and archaeal communities. Soil Biol. Biochem. 2020, 144, 107759. [Google Scholar] [CrossRef]
- Hogberg, M.N.; Baath, E.; Nordgren, A.; Arnebrant, K.; Hogberg, P. Contrasting effects of nitrogen availability on plant carbon supply to mycorrhizal fungi and saprotrophs-a hypothesis based on field observations in boreal forest. New Phytol. 2003, 160, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Amoo, A.E.; Babalola, O.O. Ammonia-oxidizing microorganisms: Key players in the promotion of plant growth. J. Soil Sci. Plant Nut. 2017, 17, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Treseder, K.K. A meta-an.alysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- in ‘t Zandt, D.; Hoekstra, N.J.; Wagemaker, C.A.M.; Caluwe, H.; Smit-Tiekstra, A.E.; Visser, E.J.W.; Kroon, H.; Pineda, A. Local soil legacy effects in a multispecies grassland community are underlain by root foraging and soil nutrient availability. J. Ecol. 2020, 108, 2243–2255. [Google Scholar] [CrossRef]
- Liu, Q. Ecological Research on Subalpine Coniferous Forests in China; Sichuan University Press: Chengdu, China, 2002; pp. 6–13. [Google Scholar]
- Hu, Z.; Liu, S.; Liu, X.; Hu, J.; Luo, M.; Li, Y.; Shi, S.; Wu, D.; Xiao, J. Soil and soil microbial biomass contents and C:N:P stoichiometry at different succession stages of natural secondary forest in sub-alpine area of western Sichuan, China. Acta Ecol. Sin. 2021, 41, 4900–4912. [Google Scholar]
- Beck, P.; Caudullo, G.; de Rigo, D.; Tinner, W. Betula pendula, Betula pubescens and other birches in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species, 2nd ed.; San-Miguel-Ayanz, J., de Rigo, D., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 70–73. [Google Scholar]
- Jonczak, J.; Jankiewicz, U.; Kondras, M.; Kruczkowska, B.; Oktaba, L.; Oktaba, J.; Olejniczak, I.; Pawłowicz, E.; Polláková, N.; Raab, T.; et al. The influence of birch trees (Betula spp.) on soil environment-A review. Forest Ecol. Manag. 2020, 477, 118486. [Google Scholar] [CrossRef]
- Qiang, W.; He, L.; Zhang, Y.; Liu, B.; Liu, Y.; Liu, Q.; Pang, X. Aboveground vegetation and soil physicochemical properties jointly drive the shift of soil microbial community during subalpine secondary succession in southwest China. Catena 2021, 202, 105251. [Google Scholar] [CrossRef]
- Zhang, N.; Van der Putten, W.H.; Veen, G.F. Effects of root decomposition on plant-soil feedback of early- and mid-successional plant species. New Phytol. 2016, 212, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Kivlin, S.N.; Bedoya, R.; Hawkes, C.V. Heterogeneity in arbuscular mycorrhizal fungal communities may contribute to inconsistent plant-soil feedback in a Neotropical forest. Plant Soil 2018, 432, 29–44. [Google Scholar] [CrossRef]
- He, H.; Yu, L.; Yang, X.; Luo, L.; Liu, J.; Chen, J.; Kou, Y.; Zhao, W.; Liu, Q. Effects of Different Soils on the Biomass and Photosynthesis of Rumex nepalensis in Subalpine Region of Southwestern China. Forests 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Schittko, C.; Wurst, S. Above- and belowground effects of plant-soil feedback from exotic Solidago canadensis on native Tanacetum vulgare. Biol. Invasions 2013, 16, 1465–1479. [Google Scholar] [CrossRef]
- Kempel, A.; Rindisbacher, A.; Fischer, M.; Allan, E. Plant soil feedback strength in relation to large-scale plant rarity and phylogenetic relatedness. Ecology 2018, 99, 597–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachelot, B.; Uriarte, M.; Muscarella, R.; Forero-Montana, J.; Thompson, J.; McGuire, K.; Zimmerman, J.; Swenson, N.G.; Clark, J.S. Associations among arbuscular mycorrhizal fungi and seedlings are predicted to change with tree successional status. Ecology 2018, 99, 607–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glockner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, 259–264. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, W.; He, H.; Kou, Y.; Liu, Q. Variations in the community patterns of soil nematodes at different soil depths across successional stages of subalpine forests. Ecol. Indic. 2022, 136, 108624. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 25–308. [Google Scholar]
- Lozano, Y.M.; Armas, C.; Hortal, S.; Casanoves, F.; Pugnaire, F.I. Disentangling above- and below-ground facilitation drivers in arid environments: The role of soil microorganisms, soil properties and microhabitat. New Phytol. 2017, 216, 1236–1246. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Sanchez, G.; Trinchera, L.; Russolillo, G. Plspm: Tools for Partial Least Squares Path Modeling (PLS-PM). 2015. Available online: https://github.com/gastonstat/plspm (accessed on 20 May 2022).
- Macek, P.; Schöb, C.; Núñez-Ávila, M.; Hernández Gentina, I.R.; Pugnaire, F.I.; Armesto, J.J.; Ohlemuller, R. Shrub facilitation drives tree establishment in a semiarid fog-dependent ecosystem. Appl. Veg. Sci. 2018, 21, 113–120. [Google Scholar] [CrossRef]
- Semchenko, M.; Leff, J.W.; Lozano, Y.M.; Saar, S.; Davison, J.; Wilkinson, A.; Jackson, B.G.; Pritchard, W.J.; De Long, J.R.; Oakley, S.; et al. Fungal diversity regulates plant-soil feedbacks in temperate grassland. Sci. Adv. 2018, 4, eaau4578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolan, N.S.; Hedley, M.J.; White, R.E. Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Osburn, E.D.; Elliottt, K.J.; Knoepp, J.D.; Miniat, C.F.; Barrett, J.E. Soil microbial response to Rhododendron understory removal in southern Appalachian forests: Effects on extracellular enzymes. Soil Biol. Biochem. 2018, 127, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, L.; Hu, J.; Zhang, W.; Fu, X.; Le, Y.; Jin, F. Organic carbon accumulation capability of two typical tidal wetland soils in Chongming Dongtan, China. J. Environ. Sci. 2011, 23, 87–94. [Google Scholar] [CrossRef]
- Marando, G.; Jiménez, P.; Hereter, A.; Julià, M.; Ginovart, M.; Bonmatí, M. Effects of thermally dried and composted sewage sludges on the fertility of residual soils from limestone quarries. Appl. Soil Ecol. 2011, 49, 234–241. [Google Scholar] [CrossRef]
- Tscherko, D.; Rustemeier, J.; Richter, A.; Wanek, W.; Kandeler, E. Functional diversity of the soil microflora in primary succession across two glacier forelands in the Central Alps. Eur. J. Soil Sci. 2003, 54, 685–696. [Google Scholar] [CrossRef]
- Ma, H.-K.; Pineda, A.; Hannula, S.E.; Kielak, A.M.; Setyarini, S.N.; Bezemer, T.M. Steering root microbiomes of a commercial horticultural crop with plant-soil feedbacks. Appl. Soil Ecol. 2020, 150, 103468. [Google Scholar] [CrossRef]
- Bejarano-Bolívar, A.A.; Lamelas, A.; Aguirre von Wobeser, E.; Sánchez-Rangel, D.; Méndez-Bravo, A.; Eskalen, A.; Reverchon, F. Shifts in the structure of rhizosphere bacterial communities of avocado after Fusarium dieback. Rhizosphere 2021, 18, 100333. [Google Scholar] [CrossRef]
- Lee, K.C.Y.; Dunfield, P.F.; Stott, M.B. The Phylum Armatimonadetes. In The Prokaryotes, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 447–458. [Google Scholar]
- Han, L.; Zhou, X.; Zhao, Y.; Zhu, S.; Wu, L.; He, Y.; Ping, X.; Lu, X.; Huang, W.; Qian, J.; et al. Colonization of endophyte Acremonium sp. D212 in Panax notoginseng and rice mediated by auxin and jasmonic acid. J. Integr. Plant Biol. 2020, 62, 1433–1451. [Google Scholar] [CrossRef]
- Gafta, D.; Peet, R.K.; Morgan, J. Interaction of herbs and tree saplings is mediated by soil fertility and stand evergreenness in southern Appalachian forests. J. Veg. Sci. 2019, 31, 95–106. [Google Scholar] [CrossRef]
- Hoffman, Y.; Aflalo, C.; Zarka, A.; Gutman, J.; James, T.Y.; Boussiba, S. Isolation and characterization of a novel chytrid species (phylum Blastocladiomycota), parasitic on the green alga Haematococcus. Mycol. Res. 2008, 112, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.M.; Martin, W.; James, T.Y.; Longcore, J.E.; Gleason, F.H.; Adler, P.H.; Letcher, P.M.; Vilgalys, R. Molecular phylogeny of the Blastocladiomycota (Fungi) based on nuclear ribosomal DNA. Fungal Biol. 2011, 115, 381–392. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Nutrient dynamics along a precipitation gradient in European beech forests. Biogeochemistry 2014, 120, 51–69. [Google Scholar] [CrossRef]
- Sawada, Y.; Aiba, S.-I.; Takyu, M.; Repin, R.; Nais, J.; Kitayama, K. Community dynamics over 14 years along gradients of geological substrate and topography in tropical montane forests on Mount Kinabalu, Borneo. J. Trop. Ecol. 2015, 31, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Velmala, S.M.; Rajala, T.; Smolander, A.; Petäistö, R.L.; Lilja, A.; Pennanen, T. Infection with foliar pathogenic fungi does not alter the receptivity of Norway spruce seedlings to ectomycorrhizal fungi. Plant Soil 2014, 385, 329–342. [Google Scholar] [CrossRef]
- Liu, E.; Takahashi, T.; Hitomi, T. Effect of pruning material compost on the nitrogen dynamic, soil microbial biomass, and plant biomass in different soil types. Landsc. Ecol. Eng. 2019, 15, 413–419. [Google Scholar] [CrossRef]
- Thiessen, S.; Gleixner, G.; Wutzler, T.; Reichstein, M. Both priming and temperature sensitivity of soil organic matter decomposition depend on microbial biomass-An incubation study. Soil Biol. Biochem. 2013, 57, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Lemmermeyer, S.; Lorcher, L.; van Kleunen, M.; Dawson, W. Testing the Plant Growth-Defense Hypothesis Belowground: Do Faster-Growing Herbaceous Plant Species Suffer More Negative Effects from Soil Biota than Slower-Growing Ones? Am. Nat. 2015, 186, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, J.; Verbruggen, E.; Heinze, J.; Xiang, D.; Chen, B.; Joshi, J.; Rillig, M.C. The interplay between soil structure, roots, and microbiota as a determinant of plant-soil feedback. Ecol. Evol. 2016, 6, 7633–7644. [Google Scholar] [CrossRef]
- Hassan, K.; Carrillo, Y.; Nielsen, U.N.; Pugnaire, F. The effect of prolonged drought legacies on plant–soil feedbacks. J. Veg Sci. 2021, 32, e13100. [Google Scholar] [CrossRef]
- Schmid, M.W.; van Moorsel, S.J.; Hahl, T.; De Luca, E.; De Deyn, G.B.; Wagg, C.; Niklaus, P.A.; Schmid, B. Effects of plant community history, soil legacy and plant diversity on soil microbial communities. J. Ecol. 2021, 109, 3007–3023. [Google Scholar] [CrossRef]
- Blom, D.; Fabbri, C.; Connor, E.C.; Schiestl, F.P.; Klauser, D.R.; Boller, T.; Eberl, L.; Weisskopf, L. Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions. Environ. Microbiol. 2011, 13, 3047–3058. [Google Scholar] [CrossRef]
- Chamkhi, I.; El Omari, N.; Balahbib, A.; El Menyiy, N.; Benali, T.; Ghoulam, C. Is the rhizosphere a source of applicable multi-beneficial microorganisms for plant enhancement? Saudi J. Biol. Sci. 2022, 29, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Nixon, S.L.; Daly, R.A.; Borton, M.A.; Solden, L.M.; Welch, S.A.; Cole, D.R.; Mouser, P.J.; Wilkins, M.J.; Wrighton, K.C. Genome-Resolved Metagenomics Extends the Environmental Distribution of the Verrucomicrobia Phylum to the Deep Terrestrial Subsurface. mSphere 2019, 4, e00613-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altshuler, I.; Hamel, J.; Turney, S.; Magnuson, E.; Levesque, R.; Greer, C.W.; Whyte, L.G. Species interactions and distinct microbial communities in high Arctic permafrost affected cryosols are associated with the CH4 and CO2 gas fluxes. Environ. Microbiol. 2019, 21, 3711–3727. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y. Priming effects: Interactions between living and dead organic matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Ren, J.; Liu, X.; Yang, W.; Yang, X.; Li, W.; Xia, Q.; Li, J.; Gao, Z.; Yang, Z. Rhizosphere soil properties, microbial community, and enzyme activities: Short-term responses to partial substitution of chemical fertilizer with organic manure. J. Environ. Manag. 2021, 299, 113650. [Google Scholar] [CrossRef]
- Paterson, E.; Gebbing, T.; Abel, C.; Sim, A.; Telfer, G. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytol. 2007, 173, 600–610. [Google Scholar] [CrossRef]
- Moore-Kucera, J.; Dick, R.P. Application of 13C-labeled litter and root materials for in situ decomposition studies using phospholipid fatty acids. Soil Biol. Biochem. 2008, 40, 2485–2493. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Contrasting effects of glucose, living roots and maize straw on microbial growth kinetics and substrate availability in soil. Eur. J. Soil Sci. 2009, 60, 186–197. [Google Scholar] [CrossRef]
- Cheng, W. Rhizosphere priming effect: Its functional relationships with microbial turnover, evapotranspiration, and C-N budgets. Soil Biol. Biochem. 2009, 41, 1795–1801. [Google Scholar] [CrossRef]
- Sun, S.; Li, S.; Avera, B.N.; Strahm, B.D.; Badgley, B.D. Soil Bacterial and Fungal Communities Show Distinct Recovery Patterns during Forest Ecosystem Restoration. Appl. Environ. Microbiol. 2017, 83, e00966-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielson, R.M. Ectomycorrhiza formation by the operculate discomycete Sphaerosporella brunnea (Pezizales). Mycologia 1984, 76, 454–461. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fert. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Liang, B.; Yang, X.; Murphy, D.V.; He, X.; Zhou, J. Fate of 15 N-labeled fertilizer in soils under dryland agriculture after 19 years of different fertilizations. Biol. Fert. Soils 2013, 49, 977–986. [Google Scholar] [CrossRef]
- Nash, J.; Laushman, R.; Schadt, C. Ectomycorrhizal fungal diversity interacts with soil nutrients to predict plant growth despite weak plant-soil feedbacks. Plant Soil 2020, 453, 445–458. [Google Scholar] [CrossRef]
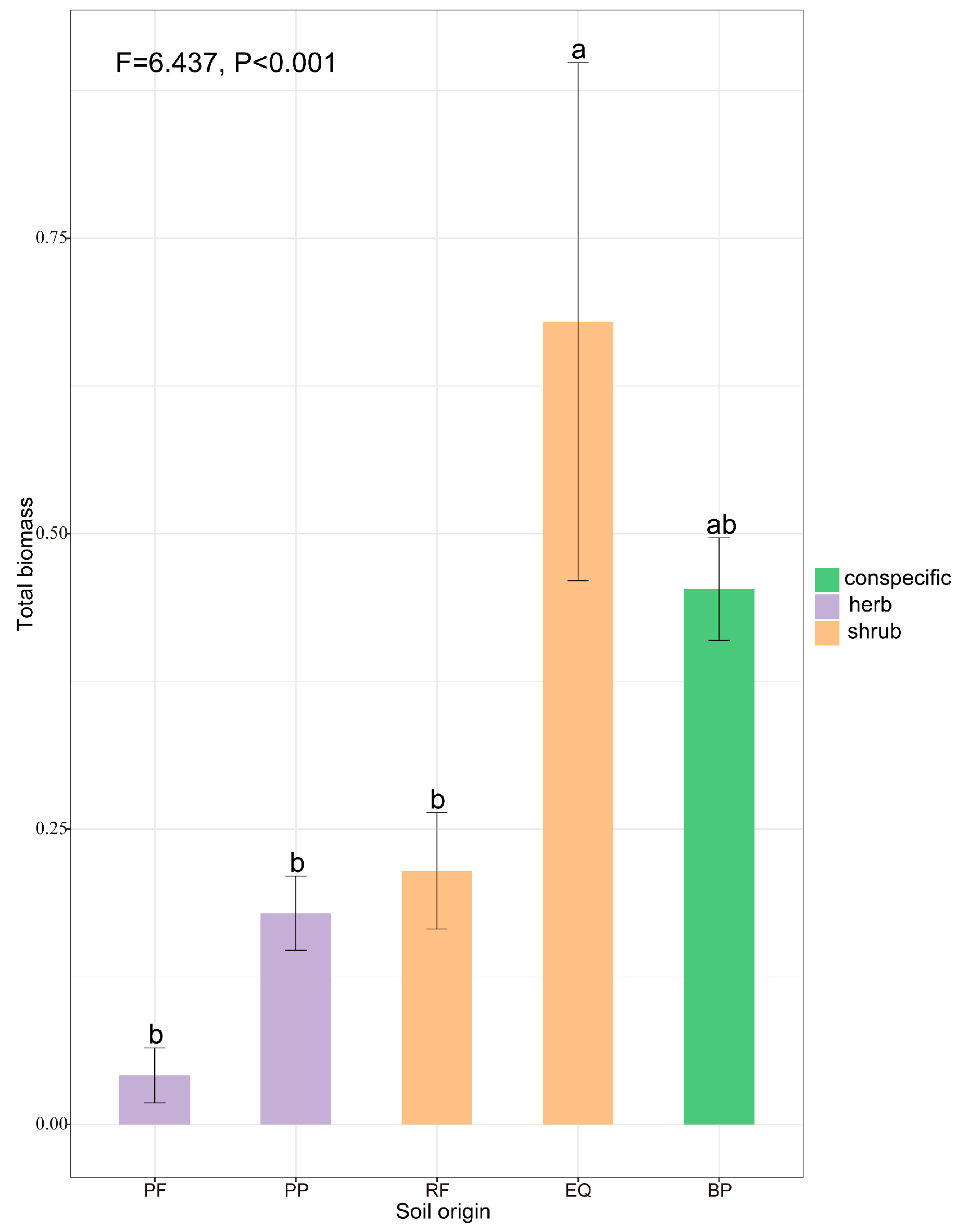

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AP (mg kg−1) | -N (mg kg−1) | -N (mg kg−1) | SOC (g kg−1) | |
|---|---|---|---|---|
| PP | 48.19 ± 1.5 bc | 4.04 ± 0.25 e | 1.06 ± 0.29 c | 18.58 ± 0.46 c |
| PF | 62.5 ± 8.13 ab | 30.29 ± 0.52 a | 3.95 ± 0.32 b | 22.73 ± 0.36 bc |
| RF | 82.58 ± 0.92 a | 8.13 ± 0.12 c | 3.4 ± 0.26 bc | 26.87 ± 3.0 ab |
| EQ | 56.97 ± 1.7 bc | 15.7 ± 0.39 b | 5.3 ± 0.05 b | 30.12 ± 1.48 a |
| BP | 47.19 ± 3.28 bc | 6.7 ± 0.14 cd | 3.97 ± 0.68 b | 18.11 ± 0.83 c |
| Str. | 37.13 ± 8.16 c | 6.26 ± 0.18 d | 21.04 ± 0.89 a | 26.11 ± 0.47 ab |
| Biomass | Independent Variables | Contribution of the Individual (%) | p Value | Adj. R2 of Full Model |
|---|---|---|---|---|
| Biomass | -N | 5.6 | 0.02 | 0.92 (p < 0.001) |
| SOC | 27.8 | <0.001 | ||
| Fungal communities (PC1) | 2.3 | 0.09 | ||
| Fungal communities (PC2) | 29.7 | <0.001 | ||
| Bacterial communities (PC1) | 27.3 | <0.001 | ||
| Bacterial communities (PC2) | 7.5 | 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.; Zhao, W.; Kou, Y.; Liu, J.; Liu, Q. Soil Microbial and Organic Carbon Legacies of Pre-Existing Plants Drive Pioneer Tree Growth during Subalpine Forest Succession. Forests 2022, 13, 1110. https://doi.org/10.3390/f13071110
Liang T, Zhao W, Kou Y, Liu J, Liu Q. Soil Microbial and Organic Carbon Legacies of Pre-Existing Plants Drive Pioneer Tree Growth during Subalpine Forest Succession. Forests. 2022; 13(7):1110. https://doi.org/10.3390/f13071110
Chicago/Turabian StyleLiang, Ting, Wenqiang Zhao, Yongping Kou, Jia Liu, and Qing Liu. 2022. "Soil Microbial and Organic Carbon Legacies of Pre-Existing Plants Drive Pioneer Tree Growth during Subalpine Forest Succession" Forests 13, no. 7: 1110. https://doi.org/10.3390/f13071110
APA StyleLiang, T., Zhao, W., Kou, Y., Liu, J., & Liu, Q. (2022). Soil Microbial and Organic Carbon Legacies of Pre-Existing Plants Drive Pioneer Tree Growth during Subalpine Forest Succession. Forests, 13(7), 1110. https://doi.org/10.3390/f13071110