
Unique Land Cover Classification to Assess Day-Roost Habitat Selection of Northern Long-Eared Bats on the Coastal Plain of North Carolina, USA

Abstract
:1. Introduction
2. Materials and Methods
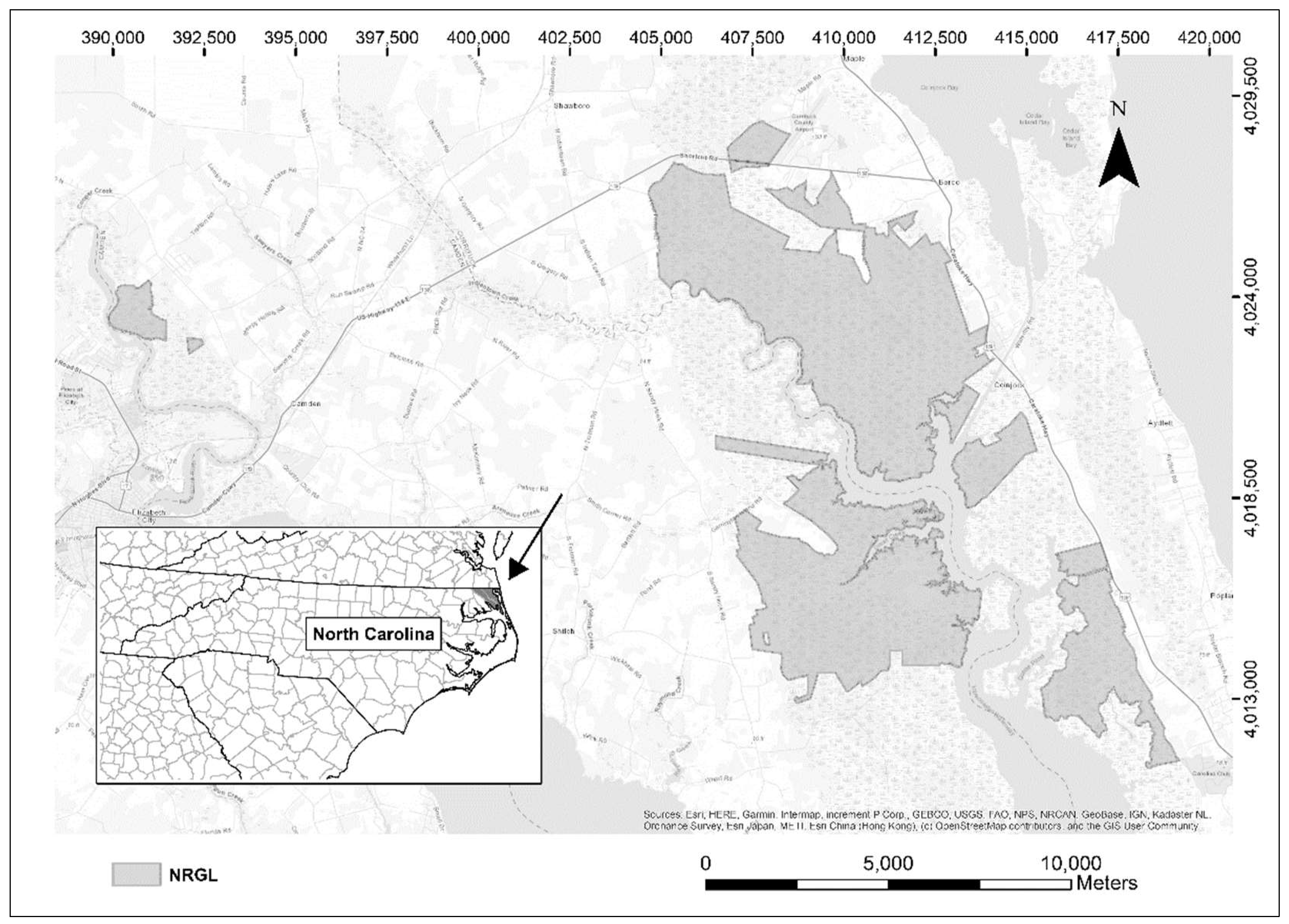
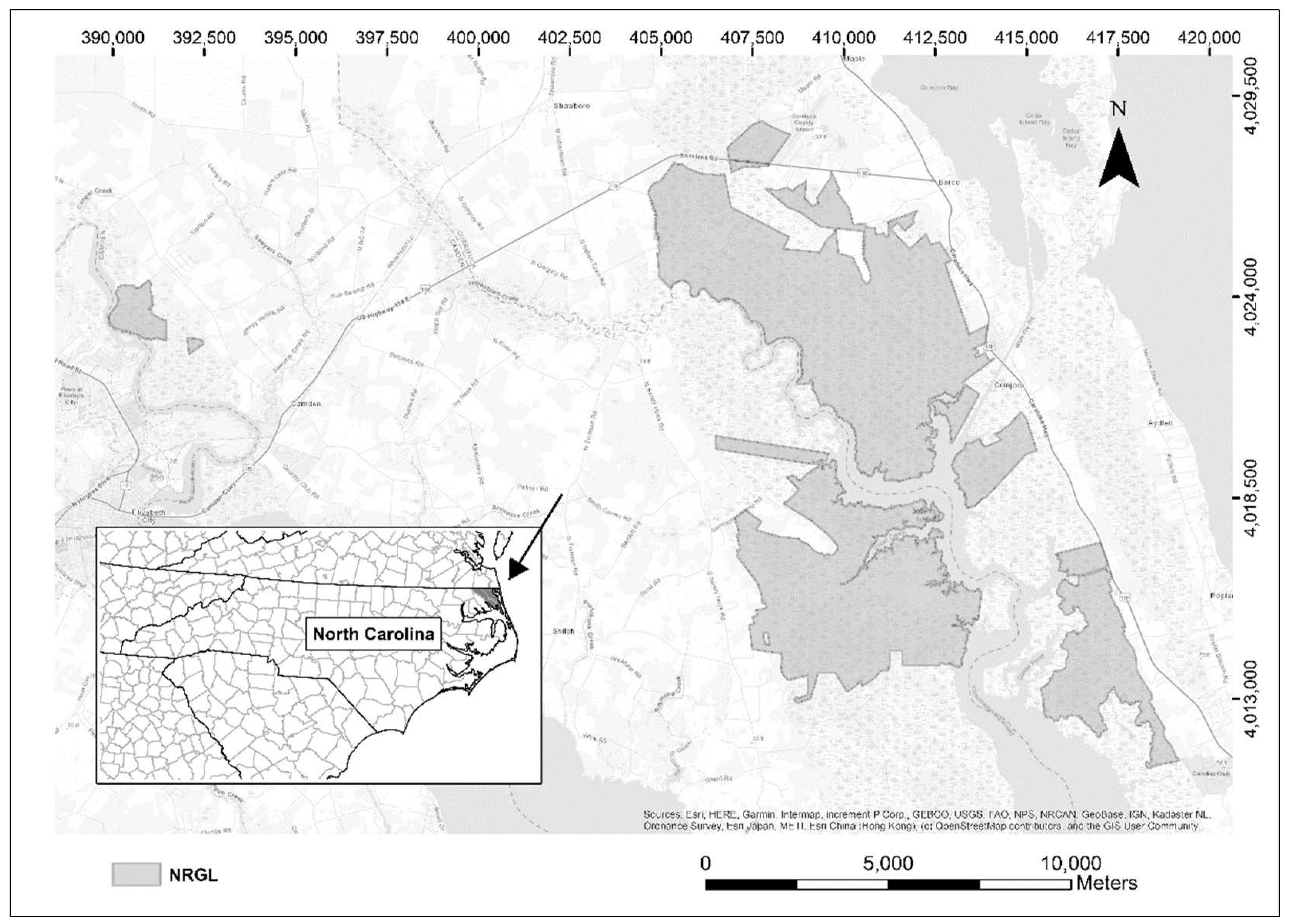
2.1. Study Area
2.2. Bat Sampling and Tracking
2.3. Roost Tree Selection and Roosting Home Range Estimates
2.4. Landscape-Scale Habitat Variables
2.5. Habitat Selection and Availability Analyses
3. Results
3.1. Roost Selection and Roosting Home Range Size
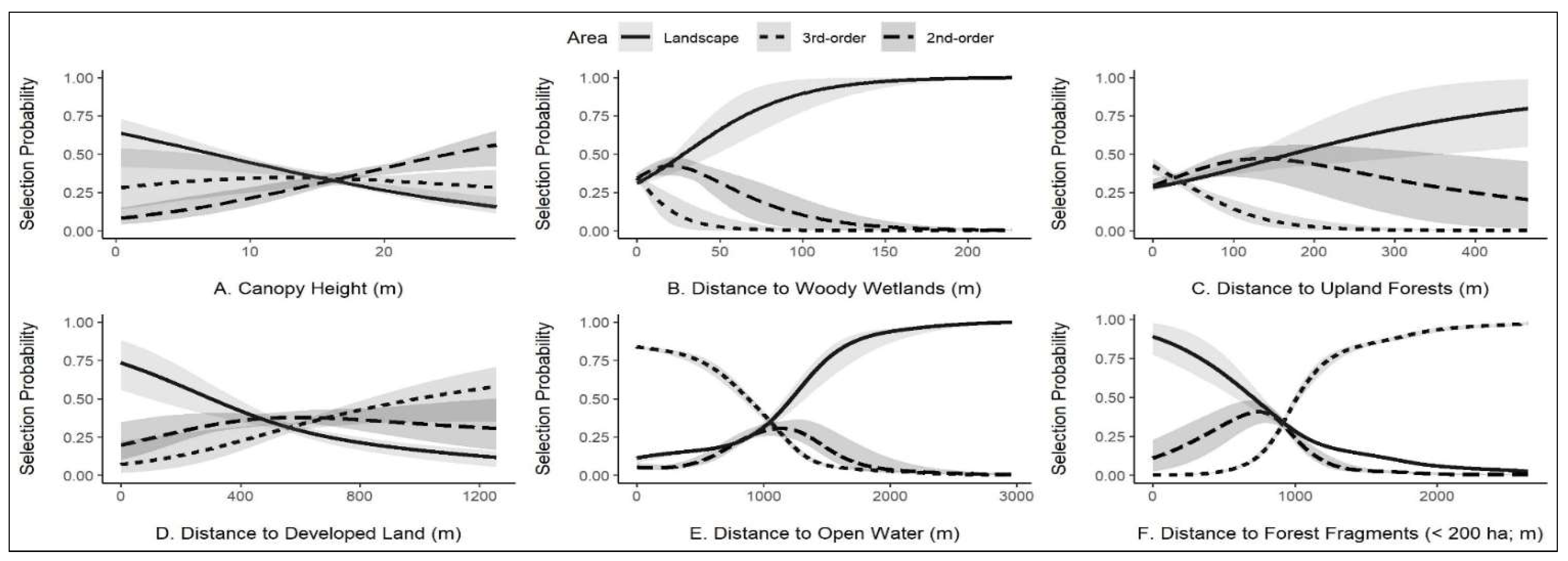
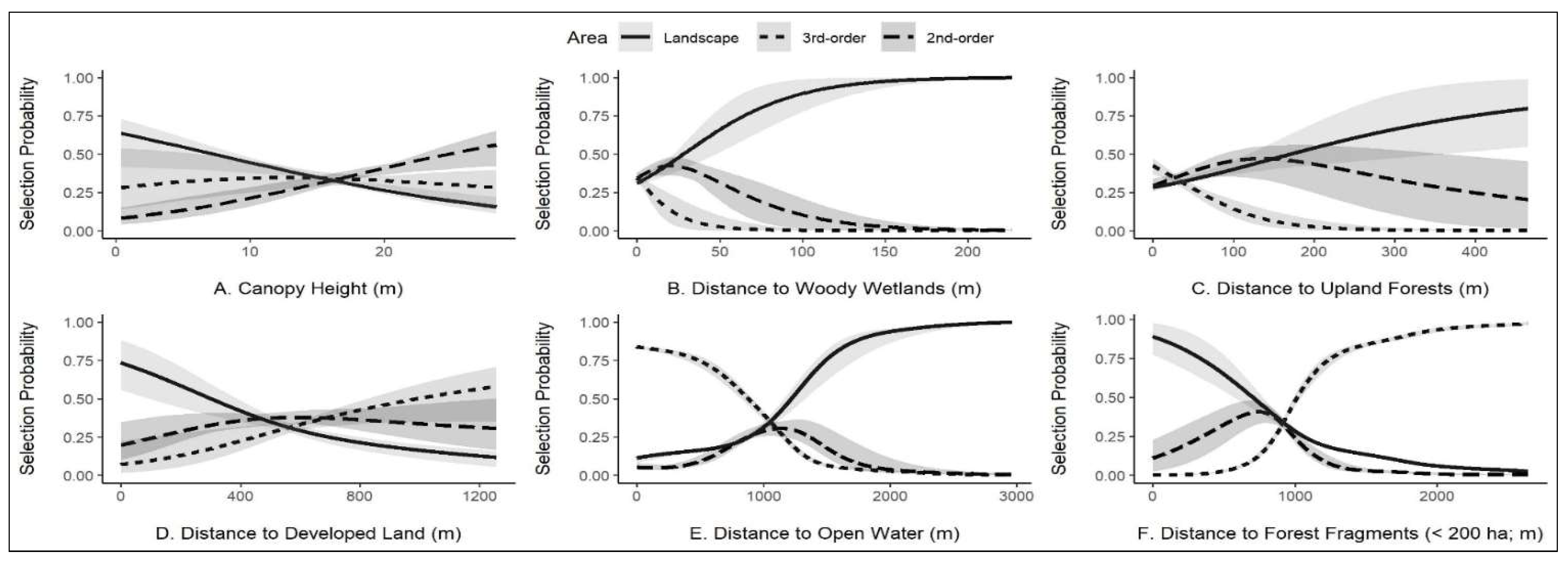
3.2. Image Classification, Habitat Selection, and Habitat Availability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frick, W.F.; Cheng, T.L.; Langwig, K.E.; Hoyt, J.R.; Janicki, A.F.; Parise, K.L.; Foster, J.T.; Kilpatrick, A.M. Pathogen dynamics during invasion and establishment of white-nose syndrome explain mechanisms of host persistence. Ecology 2017, 98, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvis, A.; Perry, R.; Ford, W.M. Relationships of Three Species of Bats Impacted by White-Nose Syndrome to Forest Condition and Management; USFS Gen. Tech. Rep. SRS–214; US Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2016; Volume SRS–214, p. 48.
- Johnson, J.B.; Rodrigue, J.L.; Ford, W.M. Nightly and Yearly Bat Activity before and after White-Nose Syndrome on the Fernow Experimental Forest in West Virginia; USDA For. Serv. North. Res. Stn. Res. Pap. NRS-Res. Pap.; USDA Forest Service: Newtown Square, PA, USA, 2013; Volume 24, pp. 1–17.
- Ford, W.M.; Silvis, A.; Rodrigue, J.L.; Kniowski, A.B.; Johnson, J.B. Deriving habitat models for northern long-eared bats (Myotis septentrionalis) from historical detection data: A case study using the Fernow Experimental Forest. J. Fish Wildl. Manag. 2016, 7, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Austin, L.V.; Silvis, A.; Ford, W.M.; Muthersbaugh, M.; Powers, K.E. Bat activity following restoration prescribed burning in the central Appalachian upland and riparian habitats. Nat. Areas J. 2018, 38, 183–195. [Google Scholar] [CrossRef]
- Committee on the Status of Endangered Wildlife in Canada (COSEWIC). Assessment and Status Report on the Little Brown Myotis (Myotis lucifugus), Northern Myotis (Myotis septentrionalis), Tri-Colored Bat (Perimyotis subflavus) in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2013; p. 93.
- United States Fish and Wildlife Service Endangered and threatened wildlife and plants; endangered species status for northern long-eared bat. Fed. Regist. 2022, 87, 16442–16452.
- Straw, B.R.; Martin, J.A.; Reichard, J.D.; Reichert, B.E. Analytical assessments in support of the U.S. Fish and Wildlife Service 3-bat species status assessment. Fish Wildl. Serv. Cat. 2022, 271. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Endangered and threatened wildlife and plants; threatened species status for the northern long-eared bat with 4(d) rule; final rule and interim rule. Fed. Regist. 2015, 80, 17974–18033. [Google Scholar]
- Francl, K.E.; Ford, W.M.; Sparks, D.W.; Brack, V. Capture and reproductive trends in summer bat communities in West Virginia: Assessing the impact of white-nose syndrome. J. Fish Wildl. Manag. 2012, 3, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, R.J.; Powers, K.E.; Orndorff, W.; Ford, W.M.; Hobson, C.S. Changes in rates of capture and demographics of Myotis septentrionalis (northern long-eared bat) in western Virginia before and after onset of white-nose syndrome. Northeast. Nat. 2016, 23, 195–204. [Google Scholar] [CrossRef]
- Kalen, N.J.; Muthersbaugh, M.S.; Johnson, J.B.; Silvis, A.; Ford, W.M. Northern long-eared bats in the central Appalachians following white-nose syndrome: Failed maternity colonies? J. Southeast. Assoc. Fish Wildl. Agencies. 2022, 9, 159–167. [Google Scholar]
- Silvis, A.; Kniowski, A.B. Distribution of Indiana Bats (Myotis sodalis) and Northern Long-Eared Bats (M. septentrionalis) in Virginia – Final Project Report to the U.S. Fish and Wildlife Service; U.S. Fish and Wildlife Service, Virginia Field Office: Glouchester, VA, USA, 2017; pp. 1–68.
- Grider, J.F.; Larsen, A.L.; Homyack, J.A.; Kalcounis-Rueppell, M.C. Winter activity of coastal plain populations of bat species affected by white-nose syndrome and wind energy facilities. PLoS ONE 2016, 11, e0166512. [Google Scholar] [CrossRef]
- Morris, A.D.; Vonhof, M.J.; Miller, D.A.; Kalcounis-Rueppell, M.C. Myotis septentrionalis Trouessart (northern long-eared bat) records from the coastal plain of North Carolina. Southeast. Nat. 2009, 8, 355–362. [Google Scholar] [CrossRef]
- Kindel, J. Northern Long-Eared Bat Project: At Santee Coastal Reserve and Wildlife Management Area and the Nature Conservancy Washo Reserve; South Carolina State Library: Columbia, SC, USA, 2019; 32p. [Google Scholar]
- Stevens, R.D.; Garcia, C.J.; Madden, M.A.; Gregory, B.B.; Perry, R.W. Seasonal changes in the active bat community of the Kisatchie National Forest, Louisiana. Southeast. Nat. 2020, 19, 524–536. [Google Scholar] [CrossRef]
- Crnkovic, A.C. Discovery of northern long-eared myotis, Myotis septentrionalis (Chiroptera: Vespertilionidae), in Louisiana. Southwest. Nat. 2003, 48, 715–717. [Google Scholar] [CrossRef]
- Ford, W.M.; Silvis, A.; Johnson, J.B.; Edwards, J.W.; Karp, M. Northern long-eared bat day-roosting and prescribed fire in the central Appalachians, USA. Fire Ecol. 2016, 12, 13–27. [Google Scholar] [CrossRef]
- Deeley, S.M.; Kalen, N.J.; Freeze, S.R.; Barr, E.L.; Ford, W.M. Post-white-nose syndrome passive acoustic sampling effort for determining bat species occupancy within the mid-Atlantic region. Ecol. Indic. 2021, 125, 107489. [Google Scholar] [CrossRef]
- Jordan, G.W. Status of an anomalous population of northern long-eared bats in coastal North Carolina. J. Fish Wildl. Manag. 2020, 11, 665–678. [Google Scholar] [CrossRef]
- Deeley, S.M.; Johnson, J.B.; Ford, W.M.; Gates, J.E. White-nose syndrome-related changes to Mid-Atlantic bat communities across an urban-to-rural gradient. BMC Zool. 2021, 6, 12. [Google Scholar] [CrossRef]
- Dowling, Z.R.; O’Dell, D.I. Bat use of an island off the coast of Massachusetts. Northeast. Nat. 2018, 25, 362–382. [Google Scholar] [CrossRef]
- Gorman, K.M.; Barr, E.L.; Ries, L.; Nocera, T.; Ford, W.M. Bat activity patterns relative to temporal and weather effects in a temperate coastal environment. Glob. Ecol. Conserv. 2021, 30, e01769. [Google Scholar] [CrossRef]
- Gorman, K.; Deeley, S.; Barr, E.; Freeze, S.; Kalen, N.; Muthersbaugh, M.; Ford, W. Broad-scale geographic and temporal assessment of northern long-eared bat (Myotis septentrionalis) maternity colony–landscape association. Endanger. Species Res. 2022, 47, 119–130. [Google Scholar] [CrossRef]
- De La Cruz, J.L.; Ward, R.L.; Schroder, E.S. Landscape characteristics related to use of artificial roosts by northern long-eared bats in north-central West Virginia. Northeast. Nat. 2018, 25, 487–501. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R.; Johnson, J.B. Association, roost use and simulated disruption of Myotis septentrionalis maternity colonies. Behav. Processes 2014, 103, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Silvis, A.; Ford, W.M.; Britzke, E.R.; Beane, N.R.; Johnson, J.B. Forest succession and maternity day roost selection by Myotis septentrionalis in a mesophytic hardwood forest. Int. J. For. Res. 2012, 2012, 148106. [Google Scholar]
- Garroway, C.J.; Broders, H.G. Nonrandom association patterns at northern long-eared bat maternity roosts. Can. J. Zool. 2007, 85, 956–964. [Google Scholar] [CrossRef]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Weber, T.C.; Sparks, D.W. Summer habitat identification of an endangered bat, Myotis sodalis, across its eastern range of the USA. J. Conserv. Plan. 2013, 9, 53–68. [Google Scholar]
- Loeb, S.C.; Winters, E.A. Indiana bat summer maternity distribution: Effects of current and future climates. Ecol. Evol. 2013, 3, 103–114. [Google Scholar] [CrossRef]
- Badin, H.A. Habitat Selection and Roosting Ranges of Northern Long-Eared Bats (Myotis septentrionalis) in an Experimental Hardwood Forest System. Master’s Thesis, Ball State University, Muncie, IN, USA, 2014; 90p. [Google Scholar]
- Broders, H.G.; Forbes, G.J.; Woodley, S.; Thompson, I.D. Range extent and stand selection for roosting and foraging in forest-dwelling northern long-eared bats and little brown bats in the Greater Fundy ecosystem, New Brunswick. J. Wildl. Manag. 2006, 70, 1174–1184. [Google Scholar] [CrossRef]
- Perry, R.W.; Thill, R.E.; Leslie, D.M. Selection of roosting habitat by forest bats in a diverse forested landscape. For. Ecol. Manag. 2007, 238, 156–166. [Google Scholar] [CrossRef]
- Johnson, J.B.; Ford, M.W.; Edwards, J.W. Roost networks of northern myotis (Myotis septentrionalis) in a managed landscape. For. Ecol. Manag. 2012, 266, 223–231. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R. Effects of hierarchical roost removal on northern long-eared bat (Myotis septentrionalis) maternity colonies. PLoS ONE 2015, 10, e0116356. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jin, S.; Danielson, P.; Homer, C.; Gass, L.; Bender, S.M.; Case, A.; Costello, C.; Dewitz, J.; Fry, J.; et al. A new generation of the United States National Land Cover Database: Requirements, research priorities, design, and implementation strategies. ISPRS J. Photogramm. Remote Sens. 2018, 146, 108–123. [Google Scholar] [CrossRef]
- Schold, E.K. Using a Custom Landscape Classification to Understand the Factors Driving Site Occupancy by a Rapidly Declining Migratory Songbird. Master’s Thesis, Virginia Commonwealth University, Richmond, VA, USA, 2018; 63p. [Google Scholar]
- U.S. Department of Agriculture. Aerial 1 m Orthophotos (2018 NAIP)—Natural Color and Color Infrared. Available online: https://nrcs.app.box.com/v/naip/folder/69930349663 (accessed on 13 September 2019).
- Maxwell, A.E.; Strager, M.P.; Warner, T.A.; Zégre, N.P.; Yuill, C.B. Comparison of NAIP orthophotography and rapideye satellite imagery for mapping of mining and mine reclamation. GIScience Remote Sens. 2014, 51, 301–320. [Google Scholar] [CrossRef]
- Hayes, M.M.; Miller, S.N.; Murphy, M.A. High-resolution landcover classification using random forest. Remote Sens. Lett. 2014, 5, 112–121. [Google Scholar] [CrossRef]
- Maxwell, A.E.; Warner, T.A.; Fang, F. Implementation of machine-learning classification in remote sensing: An applied review. Int. J. Remote Sens. 2018, 39, 2784–2817. [Google Scholar] [CrossRef] [Green Version]
- Hunter, W.C.; Peoples, L.; Collazo, J.A. Partners in flight bird conservation plan for the south Atlantic Coastal Plain (physiographic area #03); American Bird Conservancy: The Plains, VA, USA, 2001; pp. 1–166. [Google Scholar]
- Hefner, J.M.; Wilen, B.O.; Dahl, T.E.; Frayer, W.E. Southeast Wetlands: Status and Trends, Mid-1970′s to Mid-1980′s; US Fish and Wildlife Service and US Environmental Protection Agency: Atlanta, GA, USA, 1994; pp. 1–32.
- PRISM Climate Group 30-Year Normals. Available online: https://prism.oregonstate.edu/normals/ (accessed on 27 December 2021).
- Huebschman, J.J. Bats in southwest Wisconsin during the era of white-nose syndrome. Northeast. Nat. 2019, 26, 168–182. [Google Scholar] [CrossRef]
- Menzel, M.A.; Menzel, J.M.; Castleberry, S.B.; Ozier, J.; Ford, W.M.; Edwards, J.W. Illustrated Key to Skins and Skulls of Bats in the Southeastern and Mid-Atlantic States; Res. Note NE-376 Illus; U.S. Department of Agriculture, Forest Service, Northeastern Research Station: Newtown Square, PA, USA, 2002; pp. 1–10.
- Hoying, K.M.; Kunz, T.H. Variation in size at birth and post-natal growth in the insectivorous bat Pipistrellus subflavus (Chiroptera: Vespertilionidae). J. Zool. 1998, 245, 15–27. [Google Scholar] [CrossRef]
- Sikes, R.S.; Gannon, W.L. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- Ford, W.M.; Owen, S.F.; Edwards, J.W.; Rodrigue, J.L. Robinia pseudoacacia (black locust) as day-roosts of male Myotis septentrionalis (northern bats) on the Fernow Experimental Forest, West Virginia. Northeast. Nat. 2006, 13, 15–24. [Google Scholar] [CrossRef]
- Walter, W.D.; Fischer, J.W.; Baruch-Mordo, S.; VerCauteren, K.C. What is the proper method to delineate home range of an animal using today’s advanced GPS telemetry systems: The initial step. Mod. Telem. 2011, 68, 249–268. [Google Scholar]
- Beyer, H.L. Geospatial Modelling Environment (Version 0.7.2.1). Available online: http://www.spatialecology.com/gme (accessed on 3 July 2019).
- Obbard, M.E.; Coady, M.B.; Pond, B.A.; Schaefer, J.A.; Burrows, F.G. A distance-based analysis of habitat selection by American black bears (Ursus americanus) on the Bruce Peninsula, Ontario, Canada. Can. J. Zool. 2010, 88, 1063–1076. [Google Scholar] [CrossRef]
- Vogt, P.; Riitters, K.H.; Estreguil, C.; Kozak, J.; Wade, T.G.; Wickham, J.D. Mapping spatial patterns with morphological image processing. Landsc. Ecol. 2007, 22, 171–177. [Google Scholar] [CrossRef]
- Aerial Photography Field Office National Agriculture Imagery Program (NAIP) Digital Aerial Photography 3d Point Cloud Product; USDA-FSA-APFO: Salt Lake City, UT, USA, 2018.
- R Core Team A Language and Environment for Statistical Computing. Available online: http://www.r-project.org (accessed on 1 October 2019).
- Breiman, L.; Cutler, A.; Liaw, A.; Wiener, M. Package ‘randomForest’—Breiman and Cutler’s random forests for classification and regression. CRAN Repos. 2018, 2, 29. [Google Scholar]
- Gislason, P.O.; Benediktsson, J.A.; Sveinsson, J.R. Random forests for land cover classification. Pattern Recognit. Lett. 2006, 27, 294–300. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăgu, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Cheng, Y. Mean shift, mode seeking, and clustering. IEEE Trans. Pattern Anal. Mach. Intell. 1995, 17, 790–799. [Google Scholar] [CrossRef] [Green Version]
- Laliberte, A.S.; Rango, A.; Havstad, K.M.; Paris, J.F.; Beck, R.F.; McNeely, R.; Gonzalez, A.L. Object-oriented image analysis for mapping shrub encroachment from 1937 to 2003 in southern New Mexico. Remote Sens. Environ. 2004, 93, 198–210. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Rango, A. Texture and scale in object-based analysis of subdecimeter resolution unmanned aerial vehicle (UAV) imagery. IEEE Trans. Geosci. Remote Sens. 2009, 47, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S.; Cushman, S.A. Gradient modeling of conifer species using random forests. Landsc. Ecol. 2009, 24, 673–683. [Google Scholar] [CrossRef]
- Ripley, B.; Venables, W. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Sukumal, N.; Gale, G.A.; Savini, T. Sub-montane habitat selection by lowland pheasants. Raffles Bull. Zool. 2010, 58, 391–401. [Google Scholar]
- Roerick, T.M.; Cain, J.W.; Gedir, J.V. Forest restoration, wildfire, and habitat selection by female mule deer. For. Ecol. Manag. 2019, 447, 169–179. [Google Scholar] [CrossRef]
- Wilson, S.E.; Nielsen, C.K. Habitat characteristics of raccoon daytime resting sites in southern Illinois. Am. Midl. Nat. 2007, 157, 175–186. [Google Scholar] [CrossRef]
- De La Cruz, J.L.; Ward, R.L. Summer-habitat suitability modeling of Myotis sodalis (Indiana Bat) in the eastern mountains of West Virginia. Northeast. Nat. 2016, 23, 100–117. [Google Scholar] [CrossRef]
- White, T.M.; Walea, J.E.; Robinson, J. New record of northern long-eared bats in coastal South Carolina. Southeast. Nat. 2018, 17, N1–N5. [Google Scholar] [CrossRef]
- De La Cruz, J.L.; Ward, R.L.; Schroder, E.S.; Ford, W.M.; Barr, E.; Nocera, T. Post-WNS northern long-eared bat day-roosts in a residual population. In Proceedings of the North American Joint Bat Working Group Meeting & 28th Colloquium on Conservation of Mammals in the South, North American Joint Bat Working Group Meeting, Roanoke, VA, USA, 26–29 March 2018; pp. 13–14. [Google Scholar]
- Lacki, M.J.; Schwierjohann, J.H. Day-roost characteristics of northern bats in mixed mesophytic forest. J. Wildl. Manag. 2001, 65, 482. [Google Scholar] [CrossRef]
- Menzel, M.A.; Owen, S.F.; Ford, W.M.; Edwards, J.W.; Wood, P.B.; Chapman, B.R.; Miller, K.V. Roost tree selection by northern long-eared bat (Myotis septentrionalis) maternity colonies in an industrial forest of the central Appalachian mountains. For. Ecol. Manag. 2002, 155, 107–114. [Google Scholar] [CrossRef]
- Patriquin, K.J.; Leonard, M.L.; Broders, H.G.; Ford, W.M.; Britzke, E.R.; Silvis, A. Weather as a proximate explanation for fission–fusion dynamics in female northern long-eared bats. Anim. Behav. 2016, 122, 47–57. [Google Scholar] [CrossRef]
- Perry, R.W.; Thill, R.E. Roost selection by male and female northern long-eared bats in a pine-dominated landscape. For. Ecol. Manag. 2007, 247, 220–226. [Google Scholar] [CrossRef]
- Racey, P.A.; Swift, S.M. Variations in gestation length in a colony of pipistrelle bats (Pipistrellus pipistrellus) from year to year. J. Reprod. Fertil. 1981, 61, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Willis, C.K.R.; Brigham, R.M.; Geiser, F. Deep, prolonged torpor by pregnant, free-ranging bats. Naturwissenschaften 2006, 93, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Sasse, D.B.; Pekins, P.J. Summer roosting ecology of northern long-eared bats (Myotis septentrionalis) in the White Mountain National Forest. In Bats and Forests Symposium; Barclay, R.M.R., Bringham, R.M., Eds.; British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; 302p, pp. 91–101. [Google Scholar]
- Carter, T.C.; Feldhamer, G.A. Roost tree use by maternity colonies of Indiana bats and northern long-eared bats in southern Illinois. For. Ecol. Manag. 2005, 219, 259–268. [Google Scholar] [CrossRef]
- Henderson, L.E.; Broders, H.G. Movements and resource selection of the northern long-eared myotis (Myotis septentrionalis) in a forest-agriculture landscape. J. Mammal. 2008, 89, 952–963. [Google Scholar] [CrossRef] [Green Version]
- Owen, S.F.; Menzel, M.A.; Ford, W.M.; Chapman, B.R.; Miller, K.V.; Edwards, J.W.; Wood, P.B. Home-range size and habitat used by the northern Myotis (Myotis septentrionalis). Am. Midl. Nat. 2003, 150, 352–359. [Google Scholar] [CrossRef]
- Ford, W.M.; Menzel, M.A.; Rodrigue, J.L.; Menzel, J.M.; Johnson, J.B. Relating bat species presence to simple habitat measures in a central Appalachian forest. Biol. Conserv. 2005, 126, 528–539. [Google Scholar] [CrossRef]
- Johnson, J.B.; Ford, W.M.; Edwards, J.W.; Menzel, M.A. Bat community structure within riparian areas of northwestern Georgia, USA. Folia Zool. 2010, 59, 192–202. [Google Scholar] [CrossRef]
- Loeb, S.C.; O’Keefe, J.M. Habitat use by forest bats in South Carolina in relation to local, stand, and landscape characteristics. J. Wildl. Manag. 2006, 70, 1210–1218. [Google Scholar] [CrossRef]
- Johnson, J.B.; Gates, J.E.; Ford, W.M. Distribution and activity of bats at local and landscape scales within a rural-urban gradient. Urban Ecosyst. 2008, 11, 227–242. [Google Scholar] [CrossRef]
- Starbuck, C.A.; Amelon, S.K.; Thompson, F.R. Relationships between bat occupancy and habitat and landscape structure along a savanna, woodland, forest gradient in the Missouri Ozarks. Wildl. Soc. Bull. 2015, 39, 20–30. [Google Scholar] [CrossRef]
- Brooks, R.; Ford, M. Bat activity in a forest landscape of central Massachusetts. Northeast. Nat. 2005, 12, 446–447. [Google Scholar] [CrossRef]
- Dussault, C.; Courtois, R.; Ouellet, J.P.; Girard, I. Space use of moose in relation to food availability. Can. J. Zool. 2005, 83, 1431–1437. [Google Scholar] [CrossRef]
- Koprowski, J.L.; King, S.R.B.; Merrick, M.J. Expanded home ranges in a peripheral population: Space use by endangered Mt. Graham red squirrels. Endanger. Species Res. 2008, 4, 227–232. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R. Day-roost tree selection by northern long-eared bats-What do non-roost tree comparisons and one year of data really tell us? Glob. Ecol. Conserv. 2015, 3, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Ellison, L.E.; O’Shea, T.J.; Neubaum, D.J.; Bowen, R.A. Factors influencing movement probabilities of big brown bats (Eptesicus fuscus) in buildings. Ecol. Appl. 2007, 17, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, S.I.; Palmeirim, J.M. Influence of temperature in roost selection by Pipistrellus pygmaeus (Chiroptera): Relevance for the design of bat boxes. Biol. Conserv. 2004, 119, 237–243. [Google Scholar] [CrossRef]
- Johnson, J.B.; Edwards, J.W.; Ford, W.M.; Gates, J.E. Roost tree selection by northern myotis (Myotis septentrionalis) maternity colonies following prescribed fire in a central Appalachian Mountains hardwood forest. For. Ecol. Manag. 2009, 258, 233–242. [Google Scholar] [CrossRef]
- Tarabon, S.; Bergès, L.; Dutoit, T.; Isselin-Nondedeu, F. Maximizing habitat connectivity in the mitigation hierarchy. A case study on three terrestrial mammals in an urban environment. J. Environ. Manag. 2019, 243, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.R.; Noss, R.F.; Diamond, D.D.; Myers, M.N. Conservation biology and forest certification working together toward ecological sustainability. J. For. 2001, 99, 18–25. [Google Scholar]
- Lorch, J.M.; Palmer, J.M.; Lindner, D.L.; Ballmann, A.E.; George, K.G.; Griffin, K.; Knowles, S.; Huckabee, J.R.; Haman, K.H.; Anderson, C.D.; et al. First detection of bat white-nose syndrome in western North America. mSphere 2016, 1, e00148-16. [Google Scholar] [CrossRef] [Green Version]
- De La Cruz, J.L.; Schroder, E.S. Kanawha State Forest Hibernaculum Suitability Determination and Spring Emergence Survey Report, Kanawha County, West Virginia—Report Submitted to the Mary Ingles Trail Blazers; Mary Ingles Trail Blazers: Charleston, WV, USA, 2015; pp. 1–52. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Variable | DF | Wald X2 | p > X2 | ||
|---|---|---|---|---|---|
| Canopy height | 2 | 24.38 | <0.01 | ||
| Woody wetland | 2 | 15.20 | <0.01 | ||
| Upland forests | 2 | 22.74 | <0.01 | ||
| Development | 2 | 11.52 | <0.01 | ||
| Open Water | 2 | 99.47 | <0.01 | ||
| Large core forests | 2 | 64.30 | <0.01 | ||
| Non-forest areas | 2 | 5.92 | 0.05 | ||
| Final model | Log-likelihood | p > X2 | ρ2 | R2 | Accuracy |
| Canopy height + woody wetland + upland forest + development + open water + large core forests + non-forest areas | −182.86 | <0.01 | 0.45 | 0.70 | 0.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De La Cruz, J.L.; True, M.C.; Taylor, H.; Brown, D.C.; Ford, W.M. Unique Land Cover Classification to Assess Day-Roost Habitat Selection of Northern Long-Eared Bats on the Coastal Plain of North Carolina, USA. Forests 2022, 13, 792. https://doi.org/10.3390/f13050792
De La Cruz JL, True MC, Taylor H, Brown DC, Ford WM. Unique Land Cover Classification to Assess Day-Roost Habitat Selection of Northern Long-Eared Bats on the Coastal Plain of North Carolina, USA. Forests. 2022; 13(5):792. https://doi.org/10.3390/f13050792
Chicago/Turabian StyleDe La Cruz, Jesse L., Michael C. True, Hila Taylor, Dorothy C. Brown, and W. Mark Ford. 2022. "Unique Land Cover Classification to Assess Day-Roost Habitat Selection of Northern Long-Eared Bats on the Coastal Plain of North Carolina, USA" Forests 13, no. 5: 792. https://doi.org/10.3390/f13050792
APA StyleDe La Cruz, J. L., True, M. C., Taylor, H., Brown, D. C., & Ford, W. M. (2022). Unique Land Cover Classification to Assess Day-Roost Habitat Selection of Northern Long-Eared Bats on the Coastal Plain of North Carolina, USA. Forests, 13(5), 792. https://doi.org/10.3390/f13050792